Kasvit
Kasvit ovat monisoluisia, pääasiassa yhteyttämällä ravintonsa saavia eliöitä. Kuuden eliökunnan järjestelmässä ne muodostavat kasvikunnan (Plantae), yhden aitotumaisten eliöiden neljästä pääryhmästä eläinten, sienten ja alkueliöiden ohella. Kasveiksi on eri aikoina tulkinnasta riippuen voitu määritellä vaihteleva joukko erilaisia eliöitä. Kasveja tutkiva tieteen ala on kasvitiede eli botaniikka.
| Kasvit | |
|---|---|
_version_2.png.webp) Kasvien monimuotoisuutta |
|
| Tieteellinen luokittelu | |
| Luonto: | Eliökunta Biota |
| Yläkunta: | Aitotumaiset Eucarya |
| Kunta: |
Kasvit Plantae Haeckel (1866) |
| Kehityslinjat tai alakunnat | |
|
|
| Katso myös | |
Kasvit poikkeavat eläimistä ja sienistä monella tavalla. Ne eivät yleensä kykene liikkumaan paikasta toiseen, vaan pysyvät koko elämänsä tietyllä kasvualustalla. Kasvit ovat yleensä omavaraisia eli autotrofisia, sillä ne hankkivat tarvitsemansa aineet yhteyttämisen avulla. Kasvisolut poikkeavat eläinsoluista muun muassa kiinteällä, selluloosasta koostuvalla soluseinällään ja väriaineita sisältävillä plastideillaan. Sienten soluissa on kitiinin jäykistämä soluseinä, ja bakteerien soluseinässä on mureiinia.
Kasvit muodostavat levien kanssa suurimman osan planeettamme biomassasta. Lähes ainoina omavaraisina eliöinä kasvit ovat ravintoketjun perusta, ja ne ovat luoneet ilmakehän runsaan happipitoisuuden tuottamalla kaasua yhteyttämisellä. Nykyisen kaltainen elämä maapallolla on pitkälti kasvien ansiota. Maapallo voidaan jakaa alueella kasvavien kasvien mukaan eri kasvillisuusvyöhykkeisiin.
Kasvit voidaan jaotella monen tavan mukaan. Kehityksen ja sukulaisuussuhteiden mukaan kasvit voidaan ryhmitellä taksonomisesti tieteellisen luokittelun avulla. Kasvumuotojen perusteella kasvit voidaan luokitella esimerkiksi puihin, pensaisiin, köynnöksiin, ruohoihin, mehikasveihin ja päällyskasveihin.
Rakenne
Kasvin osat
Putkilokasveilla on varsi, joka toimii kasvin tukena, rakenteen perustana ja johtojänteiden ensisijaisena paikkana. Jotkut lajit kasvattavat maanpinnan alla kulkevan maavarren, jota pitkin kasvi voi levitä kasvullisesti. Paksuimmillaan varsi on vanhoilla puilla, joilla sen ympärysmitta saattaa harvoin kasvaa jopa muutamaan kymmeneen metriin. Juuri on kasvin maanalainen osa, joka ankkuroi kasvin paikalleen ja imee maaperästä ravinteita sekä vettä kasvin käyttöön. Päällyskasveilla juuret ovat usein paljaat ja niiden avulla päällyskasvi kiinnittyy isäntäkasviinsa. Viherlevillä ja eräillä sammalilla on erikoistuneiden varren, juurten ja lehtien sijaan erikoistumaton sekovarsi.
Lehti on useimmilla kasveilla pääasiallinen yhteyttävä elin. Sen muoto ja ulkonäkö vaihtelevat erittäin paljon kasviryhmästä riippuen. Tyypillinen korkean kehitystason kasvin lehti on pitkulainen, teräväkärkinen ja melko pehmeä. Havupuiden neulasiksi kutsutut lehdet ovat usein hyvin kapeita, jäykkiä ja teräviä; kaktusten lehdet taas ovat surkastuneet piikeiksi. Lihansyöjäkasveilla osa lehdistä on erikoistunut esimerkiksi suppiloiksi.
Kukka on koppisiemenisen kasvin lisääntymiselin. Se sisältää emiön ja hetiön, jotka voivat sijaita samassa kukassa, erikseen saman kasvin kukissa tai kokonaan eri kasveissa. Koristeelliset ja värikkäät terälehdet houkuttelevat hyönteisiä ja muita pölyttäjiä vierailemaan kukissa pölytyksen aikaansaamiseksi. Tuulipölytteiset kukat ovat yleensä pieniä ja vaatimattoman näköisiä. Hedelmä on koppisiemenisillä kasveilla hedelmöityksen jälkeen paisuneesta emiöstä muodostunut elin, joka sisältää kasvin siemenet.




Solutason rakenne

- Pääartikkeli: solu
Kasvisolu on aitotumallinen eli eukaryoottinen solu, kooltaan keskimäärin 10–100 μm (eläinsolut 10–30 μm). Soluseinä koostuu selluloosasta sekä muista polymeereistä, kuten pektiinistä ja ligniinistä. Se on rakenteeltaan jäykkä ja vankka, ja sen avulla solu pystyy luomaan nestepaineen, joka pitää koko kasvia muodossaan. Mikäli kasvi ei saa tarpeeksi vettä, nestepaine vähenee ja kasvi nuutuu. Viherhiukkaset eli kloroplastit ovat solulimassa sijaitsevia soluelimiä, jotka suorittavat yhteyttämisen. Niiden rakenne sisältää liukoisen perusaineksen eli strooman sekä päällekkäisistä pussimaisista kalvoista koostuvan yhteyttämis- eli tylakoidivesikkelikalvoston. Pusseissa sijaitsevat yhteyttämisreaktion aikaansaavat väripigmentit, niistä tärkeimpänä vihreä klorofylli. Ne kasvit, jotka pudottavat lehtensä talveksi, imevät ennen sitä lehdistä klorofyllin talteen sisuksiinsa, jolloin muut väriaineet tulevat näkyviin. Siten syntyy syksyinen ruska. Vakuoli eli solunesterakkula sisältää veden lisäksi entsyymejä, ravintoaineita, jäteaineksia tai jopa myrkkyjä. Nuorilla soluilla saattaa olla useita pieniä rakkuloita, jotka vanhemmiten yhdistyvät vieden jopa 90% solun tilavuudesta. Kasvisoluilla ei ole keskusjyväsiä, lysosomeja tai värekarvoja (poikkeuksena eräät yksisoluiset levät).
 Viherhiukkanen valomikroskooppikuvassa
Viherhiukkanen valomikroskooppikuvassa Viherhiukkasen mallikuva
Viherhiukkasen mallikuva Solunesterakkulan laajentuessa ja kutistuessa kasvisolu pysyy soluseinän avulla samankokoisena.
Solunesterakkulan laajentuessa ja kutistuessa kasvisolu pysyy soluseinän avulla samankokoisena.
Kasvisolukot

Solukot muodostuvat keskenään samaan tehtävään erikoistuneista soluista. Ne ovat syntyneet, kun kehittyneiden monisoluisten eliöiden solujen työnjako on johtanut solujen erilaistumiseen. Lähes kaikilla putkilokasveilla on vähintään kolmenlaisia solukkoja: Pintasolukko on tavallisesti yhden solukerroksen vahvuinen, ja se verhoaa kasvia tiiviisti. Solukon ulkoseinä on yleensä paksu ja kestävä. Sen päällä on vielä ohut vahainen kalvo, kutikula, joka suojaa kasvia liialta haihtumiselta. Perussolukko muodostaa pääosan kasvista. Solut voivat olla pyöreähköjä tai lähes suorakulmaisia, pitkiä ja paksuseinäisiä, jolloin ne ovat tukisoluja. Johtosolukko muodostaa johtojänteet.
Johtojänteet

Putkilokasveille on kehittynyt erityinen johtojänteiden verkosto, joka toimii veden ja ravinteiden tuojana soluille sekä yhteyttämistuotteiden viejänä soluista pois. Jänteet ovat selkeimmin nähtävissä lehtien suonina. Varressa jänteiden sijainti vaihtelee: yksisirkkaisilla kasveilla ne ovat sekaisin. Kaksisirkkaisilla ja havupuilla ne ovat järjestäytyneet renkaaksi poikkileikkauksen tasossa. Ytimenpuoleinen johtojänteen osa on puu ja ulompi osa nila; lisäksi niiden välissä on vielä ohut jälsi.
Ohuet jälsisolut jakaantuvat ja muodostavat uusia soluja lähinnä sisäänpäin, ja myös hieman ulospäin kuoreksi. Tähän perustuu varren paksuuskasvu. Puuosassa sijaitsevat putkilot, jotka ovat syntyneet päällekkäisten kuolleiden solujen seinämistä. Niitä pitkin vesi kulkeutuu juurista aina kasvin kärkiosiin saakka. Havupuilla on putkiloiden sijaan kapeita ja pitkiä soluja. Nilassa sijaitsevat siiviläputket. Ne muodostuvat päällekkäisistä elävistä soluista, joiden väliseinät ovat täynnä reikiä. Siiviläputket kuljettavat yhteyttämistuotteet lehdistä kasvin muihin osiin.
Puuvartisilla kasveilla johtojänteet ovat kasvaneet kiinni toisiinsa ydintä ympäröiväksi putkeksi. Niiden solut ovat paksuseinäisiä ja vahvoja, ja niiden puuosan solujen selluloosaseinissä on kerrostuneena puuainetta. Keväällä syntyvät uudet solut ovat suurikokoisempia kuin myöhemmin kesällä syntyvät, mistä aiheutuu vuosirenkaiden näkyminen puun poikkileikkauksessa.
Lisääntyminen
Kasvit voivat lisääntyä suvullisesti siementen avulla tai suvuttomasti itiöiden avulla tai kasvullisesti. Kasveille on ominaista sukupolvenvuorottelu, jossa vuorottelee suvullinen ja suvuton lisääntyminen.
Kasvien suvullisen lisääntymisen kierrossa kasvin sporofyyttisukupolvi, joka muodostaa siemenkasveilla varsinaisen näkyvän kasvin, kuten puun tai ruohon, lisääntyy suvuttomasti tuottamalla itiöitä, joista kehittyy gametofyyttejä.[1] Gametofyyttisukupolvi, joka koostuu siemenkasveilla käytännössä koiraspuolisista siitepölyhiukkasista ja naaraspuolisista siemenaiheen alkiorakoista, puolestaan tuottaa koiraspuolisia siittiöitä ja naaraspuolisia munasoluja. Varsinainen suvullinen lisääntyminen tapahtuu aina gametofyyttien kesken.[2] Koiras- ja naarasgametofyytit saattavat kehittyä samassa sporofyytin kukassa tai kukinnossa (kaksineuvoisuus) tai erillisissä hede- ja emikukissa tai -kukinnoissa (yksineuvoisuus).[3][4][5] Prosessi alkaa pölytyksellä, jossa itiöseinän suojaamista koirasgametofyyteistä koostuva siitepöly kulkeutuu joko saman kasvin (itsepölytys) tai toisen kasvin (ristipölytys) heteen ponnesta emin luotille.[6][7] Usein yksittäinen kaksineuvoinen kukka saattaa pölyttää itse itsensä.[4] Eläinpölytteiset kasvit houkuttelevat kukkien värin ja tuoksun sekä meden tai siitepölyn avulla eläimiä luokseen, jotka sitten levittävät siitepölyn muiden kukkien emiin. Useimmat eläinpölytteiset kasvit ovat hyönteispölytteisiä, mutta pölyttäjinä voivat toimia muutkin eläimet, kuten linnut tai lepakot.[8][9] Tuulipölytteiset kasvit, joita ovat monet puut ja heinät, tuottavat suuria määriä siitepölyä tuulen levitettäväksi.[10]


 Siitepöly tarttuu hyönteiseen
Siitepöly tarttuu hyönteiseen
Koppisiemenisillä kasveilla emin luotille päätynyt siitepölyhiukkanen kasvattaa pitkän siiteputken emin vartalon läpi sikiäimeen, jossa siemenaihe sijaitsee. Samalla putkessa oleva siittävä tuma on jakautunut kahtia. Niistä toinen sulautuu yhteen munasolun ja toinen keskussolun tuman kanssa. Näin hedelmöitys on tapahtunut. Sikiäin muuttuu suuremmaksi hedelmäksi, siemenaihe siemeneksi ja siemenaiheen kalvot siemenkuoriksi. Munasolusta on kehittynyt alkio siemenen sisälle ja keskussolusta yleensä varastoainesolukkoa. Paljassiemenisillä kasveilla siitepölyhiukkanen kiinnittyy suoraan siemenaiheen pinnalla olevaan nestepisaraan. Jos siitepölyä joutuu toisen lähisukua olevan kasvin emin luotille, saattaa hedelmöitys tapahtua, jolloin siemenistä kasvaa lajien risteymä eli hybridi. (Katso partenokarpia).
Siementen on epäedullista kehittyä emokasvin juurelle, joten kasveille on kehittänyt erilaisia keinoja siementensä levittämiseksi. Monien kasvien siemenissä on lisäke, jonka tuuli helposti tempaa mukaansa (esimerkiksi maitohorsma ja voikukka). Monet hedelmät taas tarjoavat syöjälleen ravintoa vastineeksi sen sisältämien siementen kuljettamisesta. Jotkut siemenet ja hedelmät takertuvat helposti turkkiin tai vaatteisiin koukkumaisten piikkien avulla (takiaiset, rusokit). Osa kasveista osaa itse singota siemenensä kauemmas, esimerkiksi käenkaali ja aho-orvokki. Aho-orvokki saattaa ampua siemeniään jopa viiden metrin päähän.
Siementen koko vaihtelee suuresti. Seychellienpalmun (Lodoicea maldivica) yhden siemenen sisältävä hedelmä painaa jopa 20 kg, kun taas kämmekkäkasvien eli orkideoiden siemeniä voi mahtua yhteen grammaan 992,25 miljoonaa.[11] Ne eivät sisällä lainkaan vararavintoa, vain pelkän alkion, joka tarvitsee sienirihmaston apua itääkseen.
 Turkkiin tarttuvia siemeniä
Turkkiin tarttuvia siemeniä Tuulen mukana kulkevia siemeniä
Tuulen mukana kulkevia siemeniä Kiivin siemenet kulkeutuvat syöjän mukana
Kiivin siemenet kulkeutuvat syöjän mukana Seychellienpalmun (Lodoicea maldivica) suuri siemen
Seychellienpalmun (Lodoicea maldivica) suuri siemen Avokadon siemenen kaaviokuva
Avokadon siemenen kaaviokuva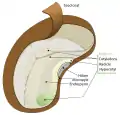 Pavun kaaviokuva
Pavun kaaviokuva
Suvuton lisääntyminen on kasveilla hyvin tavallista. Yksisoluiset levät lisääntyvät jakautumalla. Sanikkaisten ja sammalten itiöt itävät ilman hedelmöitystä. Muillakin kasveilla on runsaasti keinoja suvuttomaan lisääntymiseen, sillä kukkaa lukuun ottamatta lähes mikä tahansa kasvin osa voi kehittyä uudeksi yksilöksi. Tällaisia ovat muun muassa katkenneet oksat ja varrenpätkät (monet pensaat), juuristot (leskenlehti, peltokorte), maavarret (kielo), rönsyt (mansikka, rönsylilja), lehdet (voikukka, paavalinkukat), sipulit ja silmut. Tällä tavalla syntyneet kasvit ovat aina emokasvinsa klooneja, ellei tapahdu mutaatiota.
 Itiöinti
Itiöinti Maavarsi
Maavarsi Rönsyävä kasvi
Rönsyävä kasvi Sipulista kasvaa uutta
Sipulista kasvaa uutta Silmu
Silmu
Kasvu ja elämä
Useimmilla siemenkasveilla siemenen sisällä oleva alkio on kuivahorroksessa siihen asti, kunnes se pääsee kosketuksiin veden, lämmön ja hapen kanssa. Solut alkavat imeä vettä, jolloin ne turpoavat ja siemenkuori halkeaa. Sirkkajuuri kasvaa pääjuureksi (kaksisirkkaiset) tai surkastuu ja sen tyvestä kasvaa hajajuuria (yksisirkkaiset). Sirkkalehdet (tai sirkkalehti yksisirkkaisilla) ja sirkkavarsi alkavat kasvaa. Lehdet alkavat yhteyttää saatuaan valoa. Sirkkavarren kärjessä on silmu, josta kehittyvät kasvin muut osat. Kasvin pituuskasvu tapahtuu kasvuvyöhykkeissä, jotka ovat juurien kärjissä 5–10 mm:n pituisena alueena sekä heti verson kärjen alapuolella. Heinäkasveilla kasvuvyöhykkeitä on useita pitkin vartta. Aluksi kasvuvyöhykkeen solut jakaantuvat voimakkaasti, jonka jälkeen ne suurentuvat, kehittyvät ja erilaistuvat tehtäviinsä. Puuvartisten kasvien rungot ja juuret kasvavat myös paksuutta. Joidenkin kasvien siemenet tarvitsevat kylmä-, kuuma- tai happokäsittelyn itääkseen.
Elinkaarensa perusteella kasvit jaetaan yksivuotisiin, kaksivuotisiin ja monivuotisiin kasveihin. Yksivuotiset (monet heinä- ja viljelykasvit) kasvavat keväällä, lisääntyvät kesällä ja kuolevat talven tuloon mennessä. Kaksivuotiset (monet kaalit ja malvakasvit) kasvavat ensimmäisenä vuonna ja kukkivat sekä lisääntyvät vasta seuraavana. Monivuotiset kasvit saattavat olla lisääntymiskykyisiä vasta monien vuosien kuluttua itämisestään (esimerkiksi puut), jonka jälkeen ne tuottavat siemeniä tai itiöitä vuosittain kuolemaansa asti, mikäli olosuhteet niin sallivat. Lauhkean ja sitä kylmempien vyöhykkeiden kasvit menettävät usein lehtensä tai koko maanpäällisen osansa talveksi, poikkeuksena havupuut, jotka pitävät lehtensä ympäri vuoden. Ne ovat siten ikivihreitä. Lämpimällä vyöhykkeellä jotkut kasvit menettävät lehtensä kuivan kauden ajaksi.
Yksittäisten kasviyksilöiden elinikä vaihtelee yhdestä kasvukaudesta satoihin vuosiin. Kaikkein pisimpään elävät eräät havupuut, jotka karuilla seuduilla, kuten vuoristoissa ja levinneisyysalueensa pohjoisrajalla, ovat erittäin hidaskasvuisia. Amerikkalaiset okakäpymännyt (Pinus longaeva) ovat eläneet jopa noin 5 000-vuotiaiksi.[12] Vuonna 2022 Länsi-Australiasta löytyi Posidonia australis -meriruoho, jonka laajuus oli 180 kilometriä. Kasvin iäksi arvioitiin 4500 vuotta.[13]
 kurkun verso
kurkun verso Suuri mammuttipetäjä
Suuri mammuttipetäjä Pystyyn kuollut puu eli kelo
Pystyyn kuollut puu eli kelo
Yhteyttäminen
- Pääartikkeli: Fotosynteesi
Kasvien ravinnonsaannin perusta on yhteyttäminen, suurin osa kasveista yhteyttää fotosynteesin avulla mutta harvat kasvit käyttävät samaan tarkoitukseen kemosynteesiä. Kemosynteesissä kasvit valmistavat hiilestä ja vedystä orgaanisia yhdisteitä. Kasvit saavat energiansa kemosynteesiin hapottamalla epäorgaanisia yhdisteitä. Fotosynteesi on evolutiivisesti nuorempi ja enemmän kasveja käyttävä sitä. Siinä kasvisolut tuottavat auringon säteilyenergian avulla sokeria ja happea vedestä ja hiilidioksidista (6H2O + 6CO2 + valo → C6H12O6 (glukoosi) + 6O2). Yhteyttävä pigmentti, lehtivihreä eli klorofylli, sijaitsee kasvisolun viherhiukkasissa eli kloroplasteissa.
On olemassa noin sata varsinaisten kasvien lajia, joilla ei ole yhteyttäviä soluja. Niistä Suomessa tunnetuimpia ovat humalanvieras, pesäjuuri ja mäntykukka. Lehtivihreättömät kasvit ovat muiden kasvien loisia tai saavat ravintonsa yksinomaan sienirihmastojen avulla.

Vesi

Kuten muillekin eliöille, myös kasveille vesi on välttämätön aine. Ne tarvitsevat sitä ravintoaineiden kuljetukseen juurista lehtiin sekä lehtien valmistamien aineiden kuljetukseen niistä pois. Vettä tarvitaan myös yhteyttämisessä sokerin muodostamiseen. Solut taas imevät itseensä vettä siinä määrin, että syntyy painetta soluseinämiä vasten (nestejännitys). Sen avulla myös pehmeävartiset kasvit pysyvät jäykkinä. Solujen elintoiminnot tapahtuvat solulimassa, joka on suurimmaksi osaksi vettä. Kasvit saavat vettä juurien avulla.
Kasvin imemässä vedessä on vain noin yksi promille erilaisia suoloja. Kasvi tarvitsee suuria määriä vettä saadakseen tarvittavan määrän aineita, ja niiden päästyä lehtiin vesi käy tarpeettomaksi. Niinpä kasvi haihduttaa veden erityisten ilmarakojen kautta (putkilokasvit), ja myös suoraan lehtien pinnan kautta nuorilla ja ohuilla lehdillä. Sitä haihdunnan osuutta, joka kulkee kasvien läpi osana niiden elintoimintoja, kutsutaan transpiraatioksi.[14] Jos vettä ei ole saatavilla tarpeeksi, kasvi sulkee ilmarakonsa. Niin se tekee myös yöllä, kun yhteyttäminen lakkaa. Kuivilla seuduilla kasvit saattavat turvautua liiallista haihtumista vastaan esimerkiksi paksuilla pyöreähköillä lehdillä, jotka varastoivat vettä hyvin ja niiden haihtumispinta-ala on pieni. Lehtien pinnalla saattaa lisäksi olla paksu karva- tai vahapeite. Erittäin kuivien olojen kasveilla, kuten kaktuksilla, on erityinen aineenvaihduntajärjestelmä, CAM-yhteyttäminen, jonka turvin ne voivat pitää ilmarakonsa auki öisin ja kiinni päivisin, ja näin minimoida haihtumisen. Ne varastoivat yöllä hiilidioksidia, jolloin niiden yhteyttävät solut muuttuvat happamiksi. Päivällä ne yhteyttävät tämän varastoituneen hiilidioksidin turvin.[15]
Hengitys
Kasvit tuottavat happea, mutta myös tarvitsevat sitä omaan hengitykseensä. Prosessi on jotakuinkin vastakkainen yhteyttämiselle: kasvi hapettaa eli polttaa sokeria, josta se saa energiaa. Samalla vapautuu vettä ja hiilidioksidia. Palaminen on yhteyttämiseen nähden pientä, ja niinpä kasvit ovat siitä huolimatta hapen tuottajia. Koska kasveilta puuttuvat hengitys- ja verenkiertoelimistö, huolehtii jokainen kasvin osa omasta hapensaannistaan. Happea kulkeutuu lähinnä ilmarakojen ja kuoren huokosten kautta kasvin sisään, jossa se kulkeutuu edelleen soluväleissä. Myös juuret ottavat happea maasta, jossa sitä yleensä on tarpeeksi. Vesikasvit sen sijaan saavat happea vain lehdillään; lehtiruotiin onkin usein kehittynyt erityinen ilmakäytävä, jota pitkin happi kulkeutuu aina juuriin asti.
Symbioosi

Monet kasvit elävät symbioosissa toisen kasvin tai muuhun eliöryhmään kuuluvan lajin kanssa. Ehkä pitkälle viedyin symbioosi on jäkälä, jossa levä ja sieni muodostavat ikään kuin kaksoiseliön. Sieni imee vettä, jota rihmastojen välissä olevat levät käyttävät ja muodostavat yhteyttämällä sienen tarvitsemia aineita.[16]
Eräs tärkeä symbioosi on palkokasvien ja typpibakteerien välillä. Typpibakteerit tunkeutuvat mullasta kasvin juuriin muodostaen niihin nystyröitä. Ne alkavat muodostaa ilman typestä typpiyhdisteitä, joita syntyy yli kasvin oman tarpeen ja jotka leviävät multaan lannoitteeksi myös muiden kasvien käytettäväksi. Palkokasvi taas tuottaa bakteereille hiilihydraatteja.[17] Myös harmaalepällä on samankaltainen symbioosi. Kanervat ja kämmekkäkasvit ovat hyvin riippuvaisia niiden kanssa yhteiselämää viettävistä sienirihmastoista. Myös muilla kasveilla sienirihmasto saattaa yhteistyötarkoituksessa ympäröidä kasvin juuren muodostaen niin kutsutun sienijuuren.[18]
.jpg.webp) Sienijuuri eli mykorritsa
Sienijuuri eli mykorritsa
Lihansyöjäkasvit
Lihansyöjäkasvit ovat kehittäneet omalaatuisen tavan typen hankkimiseen; ne pyydystävät pieniä hyönteisiä houkuttelemalla ne esimerkiksi tahmeille lehdilleen (kihokit) tai erityisille suppilonmuotoisiksi erikoistuneille lehdille (kannukasvi), jonne hyönteinen putoaa. Äärimmillään kasvit saattavat pyydystää hyönteisiä aktiivisesti, kuten kärpäsloukku (Dionaea muscipula).[19] Suomessa tavataan lihansyöjäkasveista soilla kasvavia kihokkeja (Drosera), yökönlehtiä (Pinguicula) ja vesiherneitä (Utricularia)[20].
 Lihansyöjäkasvi
Lihansyöjäkasvi
Kasvit ja ihminen
Ihminen on käyttänyt kasveja ravinnokseen koko olemassaolonsa ajan. Ajalta noin 8500 eaa. ovat peräisin ensimmäiset todisteet maanviljelykulttuurin synnystä, jolloin viljeltiin aluksi lähinnä villivehnää ja pian myös ohraa, hernettä, linssejä ja pellavaa. Maanviljelyn avulla ihmisten lukumäärä alkoi kasvaa nopeasti ja he saattoivat jäädä aloilleen tietylle seudulle. Jalostuksen myötä ihminen on muokannut viljelykasveista tuottavampia ja kestävämpiä. Ihmisten ravinnoksi sopivat erityisesti helposti sulavat kasvinosat, kuten hedelmät ja juurekset.
Puusta ihminen on saanut materiaalia muun muassa asumusten ja veneiden rakentamiseen sekä paperin valmistukseen. Puuvillasta ja pellavasta hän on valmistanut kankaita, hampusta ja sisalista köysiä. Kasveista jalostetaan myös monia kemian teollisuuden tuotteita, kuten tärpättiä ja muita pesuaineita, aromaattisia öljyjä, hiusvärejä ja muuta kosmetiikkaa.
Esteettisen mielihyvän tuottamiseen ihminen kasvattaa koristekasveja puutarhoissa ja huonekasveja sisätiloissa. Kasveilla on symbolista merkitystä kansalliskukkana ja kansallispuuna, joissakin kulttuureissa puilla on maagisia ominaisuuksia. Kasveilla on myös monissa uskonnoissa suuri merkitys, kuten juutalaisuuden ja kristinuskon Hyvän ja pahan tiedon puu tai skandinaavisen mytologian Maailmanpuu sekä usean eri uskonnon pyhät lehdot.
Lääkekasveja on käytetty vuosituhansien ajan erilaisten vaivojen hoitamiseen, ja myös modernissa lääketieteessä niiden parantaviin ainesosiin on kiinnitetty yhä enemmän huomiota. Kasveista saadaan myös päihteitä, kuten tupakkaa, marihuanaa ja oopiumia.
Jotkut kasvit ovat myrkyllisiä syötyinä tai kosketuksen kautta, joidenkin siitepöly aiheuttaa allergiaa. Rikkaruohoiksi kutsutaan kasveja paikoissa, joissa niistä on haittaa, vieraslajit tai tulokaslajit voivat olla uhka alkuperäiselle kasvistolle.[21][22]
 Vehnä on yksi ihmisen ravinnokseen viljelemistä kasveista.
Vehnä on yksi ihmisen ravinnokseen viljelemistä kasveista. Hedelmiä
Hedelmiä Kaadettuja puita rakentamiseen ja paperin valmistukseen
Kaadettuja puita rakentamiseen ja paperin valmistukseen Puuvillasta valmistetaan vaatteita
Puuvillasta valmistetaan vaatteita Puutarha
Puutarha Koristekasvi
Koristekasvi Voikukkaa pidetään usein rikkaruohona
Voikukkaa pidetään usein rikkaruohona
Systemaattinen luokittelu
Vielä ennen 1950-lukua kaikki eliöt jaettiin kasveihin ja eläimiin. Kasveja olivat kaikki ne eliöt, jotka eivät täyttäneet eläimen tuntomerkkejä, kuten sienet ja jopa bakteerit. 1900-luvun puolivälin jälkeen oli käytössä niin sanottu viiden eliökunnan järjestelmä, jossa kasvit saivat oman määritelmänsä, mutta jätti kuitenkin monet levät kasvikunnan ulkopuolelle silloiseen tarkemmin määrittelemättömään alkueliökuntaan (Protista). Määritelmä sisällytti kasvikuntaan versokasvit eli alkiolliset kasvit (Embryophyta), joita ovat sammalet ja putkilokasvit ja joille on tunnusomaista omiin tehtäviinsä erikoistuneet varsi ja lehdet erotuksena sekovartisista kasveista. Kun 1900-luvun loppupuolelle tultaessa ymmärrys kasvien polveutumishistoriasta oli kasvanut, muuttui kasvikunnan käsite hieman laveammaksi.
Johtuen kasvi-käsitteen löyhästä liitoksesta nykyisen systematiikan kanssa, ei käsitteelle voida asettaa täysin yksiselitteisiä rajoja. Nykyisessä kielessä esiintyvä kasvikunnan käsite voi tarkoittaa käytännössä kolmea eri taksonia. Suppeimmasta laajimpaan rajaukseen nämä taksonit ovat:
- Versokasvit eli alkiolliset kasvit (Embryophyta), eli Plantae sensu stricto.
- Vihreät kasvit (Viridiplantae[23], Chlorobionta tai Chloroplastida[24]), johon kuuluvat varsinaiset viherlevät (Chlorophyta) sekä näkinpartaislevät (Charophyta) ja versokasvit käsittävä Streptophytina-ryhmä. Tieteellisessä luokittelussa käytetään usein tätä rajausta.[25]
- Archaeplastida eli kehityslinja, josta nykytutkimusten perusteella ovat ilmeisesti kehittyneet vihreät kasvit, punalevät (Rhodophyta) sekä glaukofyytit (Glaucophyta). Tästä taksonista on mahdollista käyttää myös termiä Plantae sensu lato.
Arviot maapallon kasvilajien määrästä vaihtelevat suuresti. Eri tutkijoiden esittämiä lukuja koppisiemenisten kasvien määrälle ovat 231 413 (vuonna 1974),[26] nykyään yleisesti hyväksytty 270 000, 422 127 (vuonna 2001)[27] ja edellisestä riippumaton tulos 421 968[28]. Tämän lisäksi erilaisia sammalia tunnetaan noin 24 000, sanikkaisia yli 12 000 ja paljassiemenisiä yli 800.
| Alakunta / ryhmä | Kaari | Alakaari | Luokka |
|---|---|---|---|
| Viherlevät, Chlorophyta | |||
| Näkinpartaislevät, Charophyta | |||
| Versokasvit, Embryophyta | Sammalet, Bryophyta | Maksasammalet, Hepaticopsida | |
| Sarvisammalet, Anthocerotae | |||
| Lehtisammalet, Bryopsida | |||
| Liekomaiset, Lycopodiophyta | Liekokasvit, Lycopodiopsida | ||
| Mähkäkasvit, Selaginellopsida | |||
| Lahnanruohokasvit, Isoëtopsida | |||
| Sanikkaiset, Pteridophyta | Haarusanikkaiset, Psilotopsida | ||
| Kortemaiset, Sphenopsida | |||
| Saniaismaiset, Pteropsida | |||
| Siemenkasvit, Spermatophyta | Paljassiemeniset, Gymnospermae | Käpypalmut, Cycadinae | |
| Neidonhiuspuut, Ginkgoinae | |||
| Havupuut, Pinopsida | |||
| Gnetinae | |||
| Koppisiemeniset, Angiospermae | Kaksisirkkaiset, Magnoliatae | ||
| Yksisirkkaiset, Liliatae | |||
Taulukkoon on merkitty nykyisin elävien vihreiden kasvien suuntaa antava luokittelu.[29] Versokasvien osalta myös luokat on merkitty. Liekomaiset kasvit on nostettu sanikkaisten luokasta omaksi kaarekseen, sillä nykykäsityksen mukaan ne eivät ole läheistä sukua muille versokasveille.[30] Siemenkasvit jakautuvat perinteisen näkemyksen mukaan viiteen nykyään esiintyvään kaareen (käpypalmut, neidonhiuspuut, havupuut, Gnetophyta ja koppisiemeniset), mutta nykytiedon mukaan neljä ensiksi mainittua ryhmää muodostavat myös yhtenäisen paljassiemenisten ryhmän.[31] Yhteisnimitystä itiökasvit käytetään kaikista muista kasveista paitsi siemenkasveista.[29] Taksonomisesti itiökasvit eivät kuitenkaan muodosta omaa ryhmäänsä.
Kasvit voidaan luokitella myös seuraavalla tavalla 18 kaareen:[32]
- Anthocerotophyta – sarvisammalet
- Bacillariophyta – piilevät
- Bryophyta – lehtisammalet
- Chlorophyta – viherlevät
- Cyanidiophyta
- Cycadophyta – käpypalmut
- Equisetophyta
- Ginkgophyta
- Glaucophyta – glaukofyytit
- Gnetophyta
- Lycopodiophyta – liekomaiset
- Magnoliophyta
- Marchantiophyta – maksasammalet
- Pinophyta
- Prasinophyta
- Psilophyta – haarusanikkaiset
- Pteridophyta – sanikkaiset
- Rhodophyta – punalevät
Levät

- Pääartikkeli: Levä
Nykyinen tieteellinen luokittelu ei enää pidä muita leviä kuin viherleviä kasvikuntaan kuuluvina eliöinä, mutta morfologisesti ja elintavoiltaan niiden yhteneväisyydet kasvien kanssa ovat niin suuret, että kaikkia levämäisiä eliöitä on aikoinaan tällä perusteella pidetty alkukantaisina kasveina. Myöhemmin on todettu, että viherleviä lukuun ottamatta muut levämäiset eliöt eivät kuulu kasvien kehityslinjaan. Esimerkiksi ruskolevät kuuluvat eräiden muiden levämäisten eliöryhmien kanssa omaan kuntaansa, josta on lähteestä riippuen käytetty mm. nimityksiä Chromalveolata ja Chromista.[33] Sinilevien eli syanobakteerien ei katsota nykyisin kuuluvan leviin, vaan bakteereihin.[34] Levät ovat hyvin moninainen, kehityshistoriallisesti täysin epäyhtenäinen ja alkujaan varsin keinotekoisesti luokiteltu joukko eliöitä, ja nimitystä käytetäänkin nykyään lähinnä käytännöllisistä syistä. Nimitys viittaa nykyään lähinnä tiettyihin morfologisiin ominaisuuksiin tai elintapoihin, eikä tiettyyn itsenäiseen eliöryhmään.
Monisoluiset levät, kuten ruskolevät ja punalevät, ovat sekovartisia, kuten jotkut alkeelliset varsinaiset kasvitkin, eli niiden rakenteesta ei ole erotettavissa lehtiä, juuria eikä kukkia. Levät pystyvät yhteyttämään – levät muodostavatkin noin 90% kaikesta maapallolla tapahtuvasta yhteyttämisestä. Lehtivihreän ohella levät saattavat sisältää muitakin pigmenttejä, kuten keltaisia tai ruskeita karotenoideja ja sinisiä tai punaisia fykobiliinejä, jotka antavat leville niiden ominaisvärin. Myös monien levien solurakenne muistuttaa kasveja, poikkeuksena piilevät, joiden soluseinä koostuu piidioksidista. Monilla yksisoluisilla levillä on soluseinässään uintisiimoja, joiden avulla ne pystyvät uimaan vedessä. Muutamat lajit, joilla ei ole soluseinää tai se ei ole jäykkä, pystyvät lisäksi ryömimään amebojen tapaan.
Useimmat levät elävät makeissa ja suolaisissa vesistöissä, joissa ne joko ajelehtivat irrallaan tai ovat kiinnittyneinä pohjaan. Lisäksi ne ovat metsien kosteiden paikkojen tavallisia asukkeja. Leviä on kuitenkin tavattu myös lumesta, merellä olevan jääpeitteen alta ja läpikuultavien kvartsilohkareiden alta Namibin aavikolla. Ne muodostavat usein symbiooseja muiden eliöryhmien kanssa: jäkälät ovat sienten ja levien symbioottisia kaksoiseliöitä, ja korallieläimet taas tarvitsevat zooxanthellae-nimistä yksisoluista levää muodostaakseen koralliriuttoja.
Kehitys
- Katso myös: Kasvien evoluution aikajana
Kaaviossa on esitetty vihreiden kasvien polveutuminen ja keskinäiset sukulaisuussuhteet eräiden lähteiden mukaan.[30][35][36] Sammalten tarkka polveutumishistoria on toistaiseksi ollut epäselvä, ja siitä on esitetty useita eri vaihtoehtoja. Sukupuuttoon kuolleet taksonit on merkitty †-merkillä.
| Viridiplantae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

Ensimmäiset yhteyttävät eliöt olivat noin 2700 miljoonaa vuotta sitten eläneitä esitumallisia syanobakteereja. Monet niistä muodostivat yhdyskuntia, stromatoliitteja, joita on yhä olemassa esimerkiksi Shark Bayn matalissa rannikkovesissä läntisessä Australiassa. Endosymbioottisen teorian mukaan levät syntyivät, kun syanobakteerit muodostivat solunsisäisen symbioosin, endosymbioosin, eräiden yksisoluisten alkueliöiden kanssa ja kehittyivät leväsolun viherhiukkasiksi. Uskotaan myös, ettei näin syntyneitä kantamuotoja ollut vain yksi, vaan endosymbioosi muodostui samoihin aikoihin useiden eri lajien kanssa, jotka sitten kehittyivät muun muassa rusko-, kulta-, puna- ja viherleviksi. Ensimmäinen tunnettu aitotumainen levä oli Grypania, joka ilmestyi noin 2100 miljoonaa vuotta sitten.[37]
Seuraavat 1500 miljoonaa vuotta eliöt pysyttelivät merissä, sillä tappavat UV-säteet pääsivät pinnalle saakka otsonikerroksen puuttuessa. Vähitellen levien yhteyttämisen ansiosta ilmakehän happipitoisuus nousi riittävälle tasolle synnyttääkseen otsonikerroksen, ja näin elämä myös maanpinnalla tuli mahdolliseksi. Luonnolla on taipumus täyttää tyhjät ekologiset lokerot, ja niinpä ordovikikaudella 490–450 miljoonaa vuotta sitten kasvit alkoivat siirtyä myös maalle.[38] DNA-ajoituksen perusteella on kuitenkin arveltu ensimmäisten maakasvien ilmestyneen jo selvästi aiemmin, noin 700 miljoonaa vuotta sitten.[39]
Maalle siirtyneet ensimmäiset pioneerikasvit olivat kehittyneitä leviä, syanobakteereja ja jäkäliä. Viherlevien Charophyta-luokan lajeista kehittyivät sammalet, ensimmäiset varsinaiset kasvit. Sammalet olivat edelleen sidoksissa kosteisiin paikkoihin, sillä ne ovat poikilohydrisiä, eli niiden kosteustasapaino määräytyy ympäristön mukaan. Kuivuuden koittaessa myös sammalet kuivuvat, siirtyvät horrosmaiseen tilaan, ja elpyvät taas nopeasti olojen muututtua suotuisammiksi.[40]
Siluurikauden alkupuolella noin 425 miljoonaa vuotta sitten ilmaantuivat ensimmäiset putkilokasvit, joista fossiililöytöjen perusteella vanhin tunnettu on muutaman senttimetrin korkuinen Cooksonia, jonka lehdettömien varsien päässä oli itiöpesäke. Rakenteeltaan paljon monimutkaisempi Baragwanathia oli lähes yhtä varhainen laji, ja sillä oli spiraalimaisesti järjestäytyneet lehdet. Putkilokasveille kehittyi kuivumiselta suojaava vahakerros niiden pinnalle, säädeltävät ilmaraot lehtien alapinnoille sekä puuosa, joiden putkilot tuovat juurien imemän veden ja ravinteet kasvin käytettäväksi. Putkilokasvit olivat siis homoiohydrisiä, ne pystyivät ylläpitämään kosteustasapainonsa ympäristön heilahteluista huolimatta. Kilpailu valosta sai aikaan lehtien kehittymisen, jotta kasvit saisivat kerättyä auringon energiaa tehokkaammin; samalla vedenottomekanismien oli edelleen kehityttävä haihtumisen lisääntyessä.

Devonikaudella 408–362 miljoonaa vuotta sitten tapahtui uusien evolutiivisten kehitysaskelten myötä voimakas putkilokasvien runsastuminen ja niiden moninaisuuden lisääntyminen. Lieot ja kortteet ilmestyivät näihin aikoihin, samoin saniaiset, sinettipuut, käpypalmut sekä Calamites-puut. Itiökasveista kehittyi niiden sekä siemenkasvien välimuoto Progymnospermae. Kauden lopulla kehittyivät ensimmäiset liekomaiset puut ja paljassiemeniset siemenkasvit. Vanhimmat siementen fossiilit ovat noin 365 miljoonaa vuotta vanhoja, ja niiden esimuotoja tavattiin ainakin 20 miljoonaa vuotta aiemmin.
Kivihiilikaudella 362–290 miljoonaa vuotta sitten ilmasto oli lämmin, lämpötilanvaihtelut pieniä ja maaperä soista. Olosuhteet olivat ihanteelliset korte- ja saniaispuille, joiden itiöt tarvitsivat kosteita olosuhteita itääkseen. Ne muodostivat ensimmäiset metsät. Myös monet muut devonikaudella kehittyneet kasviryhmät kukoistivat. Suuri osa niistä kuoli kuitenkin sukupuuttoon paleotsooisen maailmankauden loppuun tultaessa olosuhteiden muututtua epäsuotuisammiksi.
Permikaudella 290–245 miljoonaa vuotta sitten ilmasto oli viileä ja kuiva. Mannerjäätiköt valtasivat alaa eteläisellä pallonpuoliskolla. Ensimmäiset havupuut ilmaantuivat. Tyypillisimpiä lajeja tälle aikakaudelle olivat Gondwanamantereelle levinneet Glossopteris-siemensaniaiset, jotka kuitenkin hävisivät kauden lopulla. Triaskaudella monentyyppiset siemensaniaiset sekä paljassiemeniset havupuut, käpypalmut ja neidonhiuspuut yleistyivät ja muodostivat valtaosan metsistä koko Pangaean suurmantereella. Niiden – erityisesti käpypalmujen – valtakausi jatkui vielä jurakaudella 208–145 miljoonaa vuotta sitten.
Liitukaudella (145–65 miljoonaa vuotta sitten) koitti koppisiemenisten (Magnoliophytina) vallankumous. Ne kehittyivät yhteisevoluutiossa niitä pölyttävien hyönteisten kanssa nopeammin kuin muut kasvit, ja ne valtasivatkin nopeasti alaa. Kauden puolivälissä jo noin 90 prosenttia maakasveista oli kukkakasveja.
Kenotsooisella maailmankaudella (alkaen 65 miljoonaa vuotta sitten) yleistyivät heinäkasvit (Poaceae), jotka vaikuttivat kasvinsyöjänisäkkäiden evoluutioon. Maapallon keskilämpötilan laskiessa mioseenilla (23,2–5,2 miljoonaa vuotta sitten) heinäarot alkoivat syrjäyttää metsiä, ja plioseenin (5,2–1,6 miljoonaa vuotta sitten) alkuun mennessä esiintyi jo aavikoita.
Katso myös
- kasvitiede eli botaniikka
- kasvillisuusvyöhyke
- puutarhanhoito
- kasvien lämpötilastressi
- kasveja kasvutavan mukaan: puu, pensas, köynnös, ruoho, mehikasvi
- kasveja kasvupaikan mukaan: päällyskasvi
Lähteet
- Soveri, Ulvinen, Kalliola: Kasvioppi. Otava, 1970.
- Uusi Pikkujättiläinen. WSOY, 1989.
- CD-Facta 2004. WSOY. ISBN 951-0-28676-1.
- Plant evolution University of Waikato. (englanniksi)
- Rikkinen, Jouko: Leviä, sieniä ja leväsieniä. Yliopistopaino, 1999. ISBN 9789515703637
Viitteet
- Kasvitiede: sporofyytti Tieteen termipankki. Helsingin yliopisto. Viitattu 25.8.2017.
- Kasvitiede: gametofyytti Tieteen termipankki. Helsingin yliopisto. Viitattu 25.8.2017.
- Kasvitiede: yksineuvoinen Tieteen termipankki. Helsingin yliopisto. Viitattu 25.8.2017.
- Kasvitiede: yksineuvoinen Tieteen termipankki. Helsingin yliopisto. Viitattu 25.8.2017.
- Kasvitiede: kaksineuvoinen Tieteen termipankki. Helsingin yliopisto. Viitattu 25.8.2017.
- Kasvitiede: pölytys Tieteen termipankki. Helsingin yliopisto. Viitattu 25.8.2017.
- Kasvitiede: siitepölyhiukkanen Tieteen termipankki. Helsingin yliopisto. Viitattu 25.8.2017.
- Kasvitiede: eläinpölytys Tieteen termipankki. Helsingin yliopisto. Viitattu 25.8.2017.
- Kasvitiede: hyönteispölytys Tieteen termipankki. Helsingin yliopisto. Viitattu 25.8.2017.
- Kasvitiede: tuulipölytys Tieteen termipankki. Helsingin yliopisto. Viitattu 25.8.2017.
- Guinness World Records 2005, s. 74–75. Sanoma Magazines Finland, 2004.
- Miten vanha on vanhin puu? National Geographic Suomi. 4.12.2013. Viitattu 20.2.2019.
- Australiassa löydettiin maailman suurin kasvi – meriruoho on levittäytynyt yli 180 kilometriä leveälle alueelle Yle. 2022.
- Heikki Pajula ja Lasse Järvenpää (toim.): Maankuivatuksen ja kastelun suunnittelu – Työryhmän mietintö – osa 2. Suomen ympäristökeskus, syyskuu 2007. ISBN 978-952-11-2805-9 (PDF) ISSN 1796-1726 (verkkoj.). Teoksen verkkoversio. (suomeksi)
- Crassulacean Acid Metabolism (CAM) Hyperphysics. Georgia State University.
- Symbiosis Lichens Ireland. Arkistoitu 2.6.2013. Viitattu 26.9.2011. (yksityinen sivusto?)
- Typen Kierto Solunetti
- Wilhelmiina Hämäläinen: Sienijuuret – maan verisuonet Helsingin yliopisto. Arkistoitu 16.2.2013. Viitattu 26.9.2011. (yksityisen henkilön omat nettisivut?)
- Dionaea muscipula – The Venus Flytrap The Botanical Society of America. Arkistoitu 27.9.2011. Viitattu 27.9.2011. (englanniksi)
- Mehtola, Johanna: Syömme lihaa! – Opi tuntemaan lihansyöjäkasvit Suomen Luonto. Viitattu 6.1.2021.
- Terho E.O.: Allergiaa aiheuttavat siitepölyt Terveyskirjasto. 2009. Arkistoitu 14.5.2013. Viitattu 3.5.2012. (suomeksi)
- Vieraslajit Suomen ympäristökeskus. Viitattu 3.5.2012. (suomeksi)
- Cavalier-Smith (1981). "Eukaryote Kingdoms: Seven or Nine?". BioSystems 14 (3-4): 461–481. doi:10.1016/0303-2647(81)90050-2
- Adl et al., 2005: The New Higher Level Classification of Eukaryotes with Emphasis on the Taxonomy of Protists. J. Eukaryot. Microbiol., 52(5), 2005 pp. 399–451
- UniProt Taxonomy: Viridiplantae
- Stebbins GL. 1974 Flowering plants: evolution above the species level. London: Arnold xviii, 399p. ISBN 0674306856
- Govaerts R: How many species of plants are there?. Taxon, 2001, 2001. vsk, nro 50 (4). Artikkelin verkkoversio. (englanniksi)
- Plant population estimated BBC News. 2.7.2002. (englanniksi)
- Tirri, R., Lehtonen, J., Lemmetyinen, R., Pihakaski, S. & Portin, P.: Biologian sanakirja. Helsinki: Otava, 2001. ISBN 951-1-17618-8.
- Smith, Alan R., Kathleen M. Pryer, E. Schuettpelz, P. Korall, H. Schneider, & Paul G. Wolf. (2006). "A classification for extant ferns". Taxon 55(3): 705–731 http://www.pryerlab.net/publication/fichier749.pdf
- Stevens, P. F. (2001 onwards). Angiosperm Phylogeny Website. Version 9, June 2008 [and more or less continuously updated since]: Cycadales: Seedplants
- Catalogue of Life catalogueoflife.org. Viitattu 7.5.2012. [vanhentunut linkki]
- Macroalgae Itämeriportaali. Arkistoitu 8.9.2013. Viitattu 3.5.2012.
- Syanobakteerit eli sinilevät Suomen Ympäristökeskus. Viitattu 3.5.2012.
- Kenrick, Paul & Peter R. Crane. 1997. The Origin and Early Diversification of Land Plants: A Cladistic Study. (Washington, D.C.: Smithsonian Institution Press). ISBN 1-56098-730-8.
- Lewis, Louise A.; Richard M. McCourt:. Green algae and the origin of land plants. American Journal of Botany 2004: 91 (10): 1535–1556. http://dx.doi.org/10.3732/ajb.91.10.1535
- Eukaryotes and the First Multicellular Life Forms The Department of Paleobiology, Smithsonian
- Ordovician UCMP Berkeley
- http://sci.waikato.ac.nz/evolution/plantEvolution.shtml
- Sammalet Yle.fi oppiminen. YLE. Viitattu 3.5.2012. (suomeksi)
Aiheesta muualla
- Green plants. (englanniksi)
- Identification de plantes medicinales et plantes toxiues. (ranskaksi)
- La structure d'une fleur. (ranskaksi)
- Botanisk og plantefysiologisk leksikon. (norjaksi)