Maitohappokäyminen
Maitohappokäyminen on solujen aineenvaihdunnan käymisreaktio, jossa pyruvaatti eli palorypälehappo pelkistyy laktaattidehydrogenaasi-entsyymillä maitohapoksi. Pyruvaatti saadaan usein hajottamalla glukoosia glykolyysissä. Maitohappokäymistä tapahtuu mm. ihmisillä ja muilla eläimillä lihaksissa rasittavan liikunnan aikana ja maitohappobakteereissa näiden tuottaessa energiaa itselleen.[1] Soluissa käyminen tapahtuu usein solulimassa.[2]
Maitohappokäyminen on perinteinen ruoansäilömismentelmä, jota käytetään esimekiksi hapankaalin valmistukseen. Maitohappo lisää ruokien happamuutta. Hapattaminen muuttaa maun lisäksi myös ravintoarvoja.
Biokemia
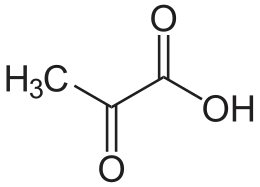
Maitohappokäymisessä pyruvaatti pelkistyy ihmisillä ja monilla eläimillä L-laktaattidehydrogenaasilla (LDH) L-maitohapoksi. Tämä on solujen olosuhteissa deprotonoitu anioni, L-laktaatti. Myös pyruvaatti on anionina (toisin kuin alla olevassa reaktiossa). NADH hapettuu reaktiossa NAD+:ksi. Tasapainoreaktio on L-laktaatin ja NAD+:n suuntaan, sillä reaktio on hyvin eksergoninen (energiallisesti suotuisa):[1]
 |
+ | NADH | + | H+ | 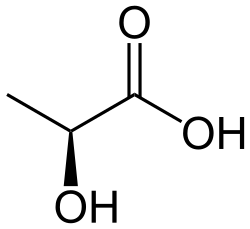 |
+ | NAD+ | |
| pyruvaatti | LDH | L-maitohappo |
Mm. joissain bakteereissa päätuotteena saadaan D-laktaatti-enantiomeeriä D-laktaattidehydrogenaasilla.[3]
Pyruvaatti saadaan usein glykolyysistä, jossa 1 glukoosi hajoaa 2:ksi pyruvaatiksi usean reaktion kautta. Ihmisten ja muiden eläinten solujen glykolyysin 6. vaiheessa glyseraldehydi-3-fosfaatti muuntuu 1,3-bisfosfoglyseraatiksi. Samalla 1 NAD+ pelkistyy NADH:ksi.[4] NADH hapettuu ihmisillä ja joillain muilla eliöillä normaalisti mitokondrioiden oksidatiivisessa fosforylaatiossa takaisin NAD+:ksi. Näin ei voi käydä liian vähähappisissa oloissa. Siksi NADH hapettuu NAD+:ksi maitohappokäymisessä. Tämä tapahtuu jotta NAD+ ei lopu, ja solulle energiaa tuottava glykolyysi voi jatkua taas vähintään 6. vaiheeseensa asti. Maitohappokäyminen ei siis tuota energiaa ATP:n muodossa.[1]

Glykolyysissä saadaan 2 ATP:tä per glukoosi. Aerobisessa eli happea kuluttavassa soluhengityksessä saadaan noin 32 ATP:tä per glukoosi. Maitohappokäymisellä toimivan solun ATP:n tuotto on siis noin 16 kertaa tehottomampaa kuin aerobisella soluhengityksellä, sillä glykolyysin ja maitohappokäymisen nettoreaktio on:[2]
Maitohappokäymistä tapahtuu mm. maitohappobakteereissa ja lihassoluissa rasittavassa liikunnassa, jossa lihasten hapen kulutus ylittää niihin hengittämällä saadun hapen määrän.[1] Käymistä tapahtuu myös monissa veden alle joutuneissa kasveissa.[5] Vähähappisuus ei ole edellytys: käymistä tapahtuu myös happirikkaissa olosuhteissa, kuten joissain syöpäsoluissa. Tällöin on kyse voi olla ns. Warburg-ilmiöstä.[6] Reaktio saattaa tapahtua myös jos solu ei pysty aerobiseen soluhengitykseen. Esim. monien eläinten kuten ihmisten punasoluissa ei ole mitokondrioita, joten ne tuottavat verensokerista energiaa maitohappokäymisellä.[1] Mm. lintujen punasoluissa tosin on toiminnalliset mitokondriot.[7]
Homofermentaatiossa kuusihiilisistä sokereista (heksooseista) muodostuu maitohappokäymisellä lähinnä vain L- tai D-maitohappoa. Heterofermentaatiossa heksooseista muodostuu runsaasti muitakin aineita kuten etikkahappoa, mannitolia ja etanolia esim. samanaikaisen etanolikäymisen kautta. Termejä käytetään jakamaan esim. maitohappobakteereita homo- ja heterofermentatiivisiin.[8]
Mm. ihmisillä lihaksissa liikunnassa muodostunut laktaatti poistuu pääosin Corin sykliin: lihasten laktaatti siirtyy vereen ja sieltä maksaan, jossa se muuntuu pääosin glukoosiksi. Tämä palaa verensokeriksi, joka voi muuntua lihaksissa taas laktaatiksi. Sykli toistuu. Laktaatin kertyminen vereen voi aiheuttaa maitohappoasidoosin – tätä ilmenee epätavallisissa tilanteissa, kuten joissain sairauksissa ja myrkytyksissä.[9]
Ruuan valmistus
Ruuan hapattaminen tehdään lisäämällä raaka-aineeseen tarkoitukseen sopiva maitohappobakteerikanta. Bakteerit käyttävät ravinnokseen raaka-aineen sokereita ja erittävät maitohappoa, joka happamoi ruuan. Bakteerien kasvua ja sen vaikutuksena maitohapon muodostumista säädellään kontrolloimalla lämpötilaa ja hapensaantia. Kun ruoka on tarpeeksi hapan, siinä eivät elä ihmiselle haitalliset bakteerilajit. Maitotuotteissa hapattaminen on maun lisäksi merkittävä myös rakenteen muodostamisessa.[10]
Tyypillisiä hapatettavia ruokia ovat
- maitotuotteista: piimä, viili, jogurtti, voi ja juusto syntyvät eri maitohappobakteerikantojen avulla[10]
- vihanneksista: valkokaalista valmistettava hapankaali sisältää tuoretta kaalia hiukan vähemmän C-vitamiinia kuin tuore kaali, mutta on helpommin sulavaa[11][12][13]. Korealaiseen keittiöön kuuluu kiinankaalista valmistettu kimchiksi kutsuttu hapankaali.
- leivistä: pelkkää täysjyväruista sisältävä ruisleipä on yleensä hapatettua
- makkaroista: meetvurstin maku ja osittain myös säilyvyys saadaan aikaan hapattamalla
Kulttuuri
Juutalaisen pesah-juhlan aikana on juutalaisilta tavallinen hapatettu leipä ankarasti kielletty. Sen sijaan tulee syödä happamatonta ja maustamatonta matsa-leipää, jota ei ole kohotettu.
Katso myös
- Malolaktinen käyminen
- Etanolikäyminen
- Raski eli leipäjuuri
- Etikkasäilöntä
Lähteet
- DL Nelson, MM Cox: Lehninger principles of biochemistry. 5. painos. W.H. Freeman, 2008. ISBN 9780716771081. Teoksen verkkoversio.
Viitteet
- Nelson, s. 546–547
- LA Urry et al: Campbell biology, s. 179–181. 11. painos. Pearson Education, Inc., 2017. ISBN 9780134093413.
- C Zhang et al: Non-sterilized fermentation of high optically pure d-lactic acid by a genetically modified thermophilic Bacillus coagulans strain. Microbial Cell Factories, 25.11.2017, 16. vsk, nro 1, s. 213. PubMed:29178877. doi:10.1186/s12934-017-0827-1. ISSN 1475-2859. Artikkelin verkkoversio.
- Nelson, s. 529
- FZ Bertolde, AF Almeida, CP Pirovani: Analysis of Gene Expression and Proteomic Profiles of Clonal Genotypes from Theobroma cacao Subjected to Soil Flooding. PLoS ONE, 7.10.2014, 9. vsk, nro 10. PubMed:25289700. doi:10.1371/journal.pone.0108705. ISSN 1932-6203. Artikkelin verkkoversio.
- MGV Heiden, LC Cantley, CB Thompson: Understanding the Warburg Effect: The Metabolic Requirements of Cell Proliferation. Science, 22.5.2009, 324. vsk, nro 5930, s. 1029–1033. PubMed:19460998. doi:10.1126/science.1160809. ISSN 0036-8075. Artikkelin verkkoversio.
- A Stier et al: Avian erythrocytes have functional mitochondria, opening novel perspectives for birds as animal models in the study of ageing. Frontiers in Zoology, 8.6.2013, nro 10, s. 33. PubMed:23758841. doi:10.1186/1742-9994-10-33. ISSN 1742-9994. Artikkelin verkkoversio.
- LC McDonald et al: A Differential Medium for the Enumeration of Homofermentative and Heterofermentative Lactic Acid Bacteria. Applied and Environmental Microbiology, kesäkuu 1987, 53. vsk, nro 6, s. 1382–1384. PubMed:16347367. ISSN 0099-2240. Artikkelin verkkoversio.
- LW Andersen et al: Etiology and therapeutic approach to elevated lactate. Mayo Clinic proceedings, 2013-10, 88. vsk, nro 10, s. 1127–1140. PubMed:24079682. doi:10.1016/j.mayocp.2013.06.012. ISSN 0025-6196. Artikkelin verkkoversio.
- Hapanmaitotuotteiden valmistus Ruokatieto
- Elintarvike - Fineli fineli.fi. Viitattu 30.9.2022.
- Hapansäilöntä Martat
- Terveyttä keräkaalista YLE Akuutti