Evoluutio
Evoluutio viittaa sukupolvien myötä tapahtuviin muutoksiin biologisten populaatioiden periytyvissä ominaisuuksissa.[1] Evoluutioprosessit tuottavat monimuotoisuutta biologisen hierarkian jokaisella tasolla, mukaan lukien lajien tasolla, yksittäisten eliöiden tasolla ja molekyylievoluution tasolla.[2]
Kaikki elämä maapallolla polveutuu universaalista esivanhemmasta joka eli noin 3,5–3,8 miljardia vuotta sitten.[3][4] Toistuvat uusien lajien synnyt eli lajiutumiset, lajien sisäiset erilaistumiset sekä lajien menetykset eli sukupuutot voidaan päätellä yhteisistä biokemiallisista ja morfologisista piirteistä sekä yhteisten DNA-jaksojen sekvensoinnista.[5] Nämä homologiset piirteet ja jaksot ovat samankaltaisimpia lajeissa, joiden sukuhaarojen eroamiseen yhteisestä esivanhemmasta on lyhin aika. Tätä tietoa voidaan käyttää rekonstruoimaan evoluution historiaa käyttäen sekä nykyisiä lajeja että fossiileja. Nykyisen biodiversiteetin kuviot ovat muotoutuneet sekä lajiutumisten että sukupuuttojen kautta.[6] On arvioitu, että enemmän kuin 99 % maapallolla eläneistä lajeista on kohdannut sukupuuton.[7][8] Maapallolla on arvioitu olevan tällä hetkelläselvennä 10–14 miljoonaa eliölajia,[9] joista noin 1,2 miljoonaa eli 14 % on dokumentoitu.[10]
1800-luvun puolivälissä Charles Darwin laati luonnonvalinnan kautta tapahtuvalle evoluutiolle tieteellisen teorian, jonka hän julkaisi kirjassaan Lajien synty (1859). Luonnonvalinnan kautta tapahtuva evoluutio on prosessi, joka voidaan päätellä kolmesta populaatioita koskevasta tosiasiasta: 1) morfologiaa, fysiologiaa ja käyttäytymistä määrittelevät piirteet vaihtelevat yksilöiden välillä, 2) eri piirteet antavat erisuuruiset eloonjäämisen ja lisääntymisen todennäköisyydet ja 3) piirteet voivat periytyä sukupolvelta toiselle.[11] Näin ollen populaation jäsenet korvautuvat peräkkäisissä sukupolvissa jälkeläisillä, joiden vanhemmat olivat paremmin sopeutuneita selviytymään ja lisääntymään ympäristössä, jossa luonnonvalinta tapahtuu. Tämä prosessi tuottaa ja säilyttää piirteitä, jotka ovat näennäisesti sopeutuneita niiden suorittamiin toiminnallisiin tehtäviin.[12] Luonnonvalinta on ainoa tunnettu syy adaptaatiolle, mutta se ei ole ainoa tunnettu syy evoluutiolle. Muita ei-adaptiivisia syitä mikroevoluutiolle ovat mutaatiot ja geneettinen ajautuminen.[13]
1900-luvun alkupuolella synteettinen evoluutioteoria sisällytti perinnöllisyystieteen Darwinin teoriaan luonnonvalinnan kautta tapahtuvasta evoluutiosta populaatiogenetiikan tieteenhaaran avulla. Luonnonvalinnan tärkeys yhtenä evoluution aiheuttajista hyväksyttiin osaksi muita biologian tieteenhaaroja. Tämän lisäksi aiemmat käsitykset evoluutiosta, kuten ortogeneesi, hylättiin.[14] Tieteilijät jatkavat evoluutiobiologian eri osa-alueiden tutkimista muodostamalla ja testaamalla hypoteeseja, kehittämällä matemaattisia malleja biologisista prosesseista, käyttämällä havaintotietoja ja suorittamalla kokeita sekä kentällä että laboratorioissa. Biologien keskuudessa on tieteellinen konsensus siitä, että polveutuminen muutoksien kera on yksi luotettavimmin perustelluista tosiasioista tieteessä. Evoluutiota tukevat biologisten todisteiden lisäksi antropologiset, psykologiset, astrofyysiset, kemialliset, fyysiset, geologiset sekä yhteiskuntatieteiden havainnot. Evoluution ymmärtämisellä on ollut suuri vaikutus ihmiskunnalle muun muassa sairauksien ennaltaehkäisyssä ja hoidossa, maatalouden kehittämisessä sekä tietokonetekniikassa evoluutioon pohjautuvan evoluutiolaskennan muodossa.[15][16][17] Biologian lisäksi evoluution havainnot ovat vieneet eteenpäin monia muita tieteenaloja, kuten biologista antropologiaa sekä evoluutiopsykologiaa, ja yhteiskuntaa kokonaisuudessaan.[18][19]
Evoluutioteorian historia




- Pääartikkeli: Evoluutioteorian historia
Ehdotus että eläinlaji voisi polveutua toisen lajin eläimestä voidaan jäljittää ensimmäisiin esisokraattisiin kreikkalaisiin filosofeihin, kuten Anaksimandrokseen ja Empedoklekseen.[21] Tämänkaltaiset ehdotukset selviytyivät antiikin Roomaan asti. Runoilija ja filosofi Lucretius seurasi Empedokleksen jalanjäljissä Maailmankaikkeudesta (lat. De rerum natura) -nimisellä teoksella.[22][23] Vastapainona näille materialistisille näkemyksille Aristoteles käsitti kaikkien luonnollisten asioiden – ei vain elävien asioiden – olevan epätäydellisiä aktualisaatioita eri pysyvistä luonnollisista mahdollisuuksista, jotka tunnetaan "ideoina".[24][25] Tämä oli osa hänen teleologista ymmärrystä luonnosta, jossa kaikilla asioilla on tietty rooli jumalallisessa kosmisessa järjestyksessä. Tähän ajattelutapaan perustuvista variaatioista tuli vakiokäsitys keskiajalla ja niitä integroitiin kristinuskon oppeihin.[26] Aristoteles ei kuitenkaan vaatinut, että oikeat eläinlajit vastaisivat yksi-yhteen täsmällisten metafyysisten ideoiden kanssa, ja antoi esimerkkejä siitä miten uusia lajeja voisi syntyä.[26]
1600-luvulla modernin tieteen uusi menetelmä hylkäsi Aristoteleen lähestymistavan ja etsi selityksiä luonnollisille ilmiöille kaikille näkyville asioille yhteisten luonnonlakien muodossa. Menetelmän ei tarvinnut olettaa mitään pysyviä luonnollisia luokkia tai jumalallista kosmista järjestystä. Tämä uusi lähestymistapa oli kuitenkin hidas juurtumaan biologisissa tieteissä, joista tuli lopulta viimeinen turvapaikka pysyvien luonnollisten tyyppien aatteelle. John Ray käytti yhtä aiemmin yleisemmässä käytössä olevista määritelmistä pysyvistä luonnollisista tyypeistä, "lajeista", viittaamaan eläin- ja kasvityyppeihin. Hän piti kuitenkin kaikkia eläviä tyyppejä jäykästi erillisinä lajeina ja ehdotti, että jokainen laji voitaisiin tunnistaa niiden sukupolvelta toiselle periytyvistä piirteistä.[27] Nämä lajit olivat Jumalan suunnittelemia mutta näyttivät erovaisuuksia paikallisten olosuhteiden takia. Myös Carl von Linnén vuonna 1735 esittämä biologinen luokittelujärjestelmä katsoi lajeja jumalallisen suunnitelman mukaisesti pysyvinä.[28]
Muut sen ajan naturalistit spekuloivat, että lajit voisivat ajan myötä muuttua evolutiivisesti luonnonlakeja seuraten. Pierre Louis Moreau de Maupertuis kirjoitti vuonna 1751, että lisääntymisen yhteydessä ilmenevät luonnolliset muutokset voisivat kasaantua useiden sukupolvien aikana ja tuottaa uusia lajeja.[29] Georges Leclerc de Buffon ehdotti, että lajit voisivat degeneroitua eri eliöiksi, ja Erasmus Darwin ehdotti, että kaikki tasalämpöiset eläimet olisivat voineet polveutua yksittäisestä mikrobista (tai "filamentista").[30] Ensimmäinen täysin kehittynyt evolutiivinen järjestelmä oli Jean-Baptiste Lamarckin "transmutaatioteoria" vuodelta 1809.[31] Tämän järjestelmän mukaan, spontaani sikiäminen tuotti jatkuvasti yksinkertaisia elämänmuotoja, joilla oli luontainen taipumus progressiiviseen kehitykseen ja jotka kehittyivät monimutkaisemmiksi eliöiksi paralleeleissa sukulinjoissa. Paikallisella tasolla, nämä sukulinjat adaptoituivat ympäristöön perimällä ominaisuudet, jotka olivat vanhempien käytössä.[32][33] (Jälkimmäistä prosessia kutsuttiin myöhemmin Lamarckismiksi.)[32][34][35][36] Vakiintuneet naturalistit tuomitsivat nämä ajatukset spekulaatioina, joilta puuttui empiirinen tuki. Erityisesti Georges Cuvier piti kiinni siitä, että lajit olivat erillisiä ja pysyviä ja että niiden yhtäläisyydet heijasti jumalallista suunnitelmaa, joka otti huomioon eliöiden toiminnalliset tarpeet. Samaan aikaan, William Paley oli käyttänyt Rayn ajatuksia hyväntahtoisesta suunnitelmasta kehittääkseen Natural Theology or Evidences of the Existence and Attributes of the Deity (1802) -nimisen, myöhemmin Charles Darwinin ihaileman, kirjan, jossa hän ehdotti, että monimutkaiset adaptaatiot olivat todisteita jumalallisesta suunnitelmasta.[37][38][39]
Ratkaiseva irtautuminen kiinteiden biologisten lajien käsitteestä alkoi, kun Charles Darwin laati luonnonvalinnan kautta tapahtuvalle evoluutiolle tieteellisen teorian. Osittain Thomas Malthusin kirjoittaman An Essay on the Principle of Population (1798) -nimisen kirjan vaikutuksen alla, Darwin totesi, että populaation kasvu johtaisi "taisteluun olemassaolon puolesta", jossa suotuisat variaatiot voisivat selviytyä kun toiset menehtyivät. Jokaisessa sukupolvessa, lukuisat jälkeläiset eivät selviydy hengissä lisääntymisikään rajallisten resurssien takia. Tämä voisi selittää kasvien ja eläinten monimuotoisuus ja yhteinen alkuperä luonnonlakeihin perustuvilla prosesseilla, jotka toimivat samalla tavalla kaikentyyppisiin eliöihin.[40][41][42][43] Darwin kehitti "luonnonvalinnaksi" kutsumaansa teoriaa vuodesta 1838 eteenpäin ja oli kirjoittamassa "isoa kirjaa" aiheesta kun Alfred Russel Wallace lähetti hänelle samankaltaisen teorian vuonna 1858. Molemmat miehet esittivät erilliset tutkielmansa Lontoon Linnean Society -tutkimusseuralle.[44] Vuoden 1859 loppupuolella, Darwinin julkaisema Lajien synty selitti luonnonvalinnan yksityiskohtaisesti ja tavalla, joka johti evoluutiokäsitteiden yhä laajempaan hyväksymiseen. Thomas Henry Huxley sovelsi Darwinin ajatuksia ihmisiin ja käytti paleontologiaa ja komparatiivista anatomiaa esittääkseen vahvaa näyttöä sille, että ihmisillä ja ihmisapinoilla on yhteiset esivanhemmat. Jotkut häiriintyivät tästä, sillä se merkitsi, että ihmisillä ei ole erityistä asemaa maailmankaikkeudessa.[45]
Perinnöllisyyden täsmälliset mekanismit ja uusien piirteiden alkuperä olivat edelleen arvoituksia. Tätä varten Darwin kehitti väliaikaisen pangeneesi (engl. pangenesis) -nimisen teorian.[46] Gregor Mendel kirjoitti vuonna 1865, että piirteet periytyivät ennakoitavalla tavalla elementtien (myöhemmin geenien) vapaan yhdistymisen ja erkanemisen kautta. Mendelin perinnöllisyyssäännöt syrjäyttivät lopulta suurimman osan Darwinin pangeneesiteoriasta.[47] August Weismann teki tärkeän erotuksen kehossa olevien itusolujen (siittiöiden ja munasolujen) ja somaattisten solujen välille osoittaen, että perinnöllisyys tapahtuu vain ituradan kautta. Hugo de Vries yhdisti Darwinin pangeneesiteorian Weismannin itu/soma -solujen erotukseen ja ehdotti, että Darwinin pangeenit sijaitsivat tumassa ja ilmaistuna siirtyivät solulimaan ja muuttivat solun rakennetta. De Vries oli myös yksi tutkijoista, jotka tekivät Mendelin työn tunnetuksi uskoen, että Mendeeliset piirteet vastasivat ituradan kautta periytyviä variaatioita.[48] Selittääkseen kuinka uusia variaatioita syntyy, De Vries kehitti mutaatioteorian, joka aiheutti hetkellisen välirikon Darwinistisen evoluution tukijoiden ja De Vriesin puolella olevien biometriikoiden välille.[33][49][50] 20. vuosisadan taitteessa, populaatiogenetiikan edelläkävijät kuten J. B. S. Haldane, Sewall Wright ja Ronald Fisher asettivat evoluutioteorian perustan vankalle tilastolliselle filosofialle. Darwinin teorian, geneettisten mutaatioiden ja Mendelin perinnöllisyyssääntöjen välillä ollut perätön ristiriita oli siten ratkaistu.[51]
1920- ja 1930-luvuilla synteettinen evoluutioteoria yhdisti luonnonvalinnan, mutaatioteorian ja Mendelin perinnöllisyyssäännöt yhtenäiseksi teoriaksi, joka soveltui yleisesti kaikkiin biologian haaroihin. Moderni synteesi kykeni selittämään populaatioissa elävien lajien välisiä kuvioita paleontologiassa olevilla välimuotofossiileilla ja kehitysbiologiassa olevilla monimutkaisilla solumekanismeilla.[33][52] James Watsonin ja Francis Crickin vuonna 1953 julkaisema DNA:n rakenne osoitti, että periytyvyydellä on fyysinen perusta.[53] Molekyylibiologia paransi ymmärrystämme genotyypin ja fenotyypin välisestä suhteesta. Edistyksiä tehtiin myös fylogeneettisessä systematiikassa, jossa kartoitettiin piirteiden muuttuminen komparatiiviseen ja testattavissa olevaan kehykseen evoluutiopuiden julkaisemisen ja käytön kautta.[54][55] Evoluutiobiologi Theodosius Dobzhansky kirjoitti vuonna 1973, että "millään asialla biologiassa ei ole mitään järkeä ellei sitä katso evoluution näkökulmasta", koska se on tuonut esiin luonnonhistoriassa aiemmin hajanaisilta vaikuttavien tosiasioiden välisiä suhteita ja yhtenäistänyt ne johdonmukaiseen selittävään tietojoukkoon, joka kuvaa ja ennustaa monia havaittavissa olevia, tällä planeetalla olevaa elämää koskevia tosiasioita.[56]
Sen jälkeen modernia synteesiä on edelleen laajennettu selittämään biologisia ilmiöitä koko biologisen hierarkian mitalta, aina geeneistä lajeihin asti. Tämä evolutiivisena kehitysbiologiana (epävirallisesti "evo-devona") tunnettu laajennus painottaa sitä miten sukupolvien väliset eroavaisuudet (evoluutio) vaikuttavat yksittäisten eliöiden sisäisiin muutoskuvioihin (kehitykseen).[57][58]
Perinnöllisyys

Eliöiden evoluutio tapahtuu periytyvien ominaisuuksien muutoksien kautta. Esimerkiksi silmien väri on peritty ominaisuus ja yksilö voi periä "ruskeiden silmien ominaisuuden" yhdeltä vanhemmalta.[59] Geenit säätelevät perittyjä ominaisuuksia ja yksilön perimässä olevaa geenikokoelmaa kutsutaan yksilön genotyypiksi.[60]
Yksilön kaikkien havaittavien ominaisuuksien kokonaisuutta kutsutaan sen fenotyypiksi. Nämä ominaisuudet ovat genotyypin ja ympäristön vuorovaikutuksen tulos.[61] Monet yksilön fenotyyppiin liittyvät ominaisuudet eivät tämän takia periydy. Esimerkiksi ruskettunut iho johtuu henkilön genotyypin ja auringon välisestä vuorovaikutuksesta eikä rusketus tämän takia periydy henkilön lapsille. Jotkut henkilöt kuitenkin ruskettuvat helpommin kuin toiset, johtuen eroista heidän genotyypeissä. Esimerkiksi henkilöillä joilla on albinismin periytyvä ominaisuus eivät rusketu lainkaan ja ovat erittäin herkkiä auringonpolttamille.[62]
Periytyvät ominaisuudet siirtyvät yhdeltä sukupolvelta seuraavalle geneettisen informaation sisältävän DNA-molekyylin avulla.[60] DNA on pitkä polymeeri, joka koostuu neljän tyyppisestä emäksestä. Emästen järjestys tietyllä DNA-molekyylin jaksolla määrittelee geneettisen informaation samaan tapaan kuin kirjainten järjestys määrittelee lauseen. Ennen kuin solu jakautuu, DNA kopioidaan niin, että kummassakin syntyvässä solussa on sama emäsjärjestys. DNA-molekyylin osia, jotka voidaan määritellä yksittäisiksi toiminnallisiksi yksiköiksi kutsutaan geeneiksi; eri geeneillä on eri emäsjärjestys. Solujen sisällä olevat pitkät DNA-juosteet muodostavat kromosomeiksi kutsuttuja tiivistettyjä rakenteita. DNA-jakson tarkkaa sijaintia kromosomissa kutsutaan lokukseksi. Jos DNA-jakso lokuksessa eroaa yksilöiden välillä, tämän jakson eri muotoja kutsutaan alleeleiksi. DNA-jaksot voivat muuttua mutaatioiden kautta, tuottaen uusia alleeleja. Jos mutaatio tapahtuu geenissä, uusi alleeli voi vaikuttaa geenin säätelemään ominaisuuteen muuttaen eliön fenotyyppiä.[63] On kuitenkin huomioitava, että vaikka tämä yksinkertainen vuorovaikutus alleelin ja ominaisuuden välillä toimii joissain tapauksissa, valtaosa ominaisuuksista on monimutkaisempia ja niiden kehitystä säätelevät useat vuorovaikutuksessa olevat geenit.[64][65]
Hiljattaiset löydökset ovat vahvistaneet tärkeitä esimerkkejä periytyvistä muutoksista, joita ei voida selittää emäsjärjestyksien muutoksilla DNA-molekyylissä. Nämä ilmiöt luokitellaan epigeneettisiksi periytymisjärjestelmiksi.[66] Kromatiinin merkitseminen DNA-metylaatiolla, omavaraiset metaboliset kytkennät, geenien vaimentaminen RNA-interferenssillä ja prionien kaltaisten proteiinien kolmiulotteiset konformaatiot ovat alueita, joilla epigeneettisiä periytymisjärjestelmiä on löydetty eliötasolla.[67][68] Kehitysbiologit ovat ehdottaneet, että monimutkaiset vuorovaikutukset geneettisissä verkostoissa ja solujen välinen kommunikaatio voivat johtaa periytyviin variaatioihin, jotka voivat olla kehityksen plastisuuden ja kanalisaation mekanismien taustalla.[69] Perinnöllisyyttä voi esiintyä myös suuremmissa mittakaavoissa. Esimerkiksi, eliöiden tavanomaiset ja toistuvat käyttäytymismallit omissa ympäristöissään ohjaavat ekologista periytyvyyttä ekolokeroiden rakentamisen prosessin kautta. Tämä tuottaa kirjon ilmiöitä, jotka muuttavat seuraavien sukupolvien valintajärjestelmää. Jälkeläiset perivät geenien lisäksi esivanhempien ekologisten tekojen tuottamat ympäristötekijät.[70] Muita esimerkkejä evoluutiossa esiintyvistä periytyvyyksistä, jotka eivät ole suoraan geenien vaikutuksen alla, ovat kulttuuripiirteiden periytyminen ja symbiogeneesi.[71][72]
Variaatio


Yksittäisen eliön fenotyyppi johtuu sekä sen genotyypistä että ympäristöstä, jossa se on elänyt. Merkittävä osa populaatiossa olevien fenotyyppien variaatiosta johtuu eroista genotyypeissä.[65] Synteettinen evoluutioteoria määrittelee evoluution muutoksina, jotka tapahtuvat tässä geneettisessä variaatiossa ajan myötä. Tietyn alleelin frekvenssi tulee enemmän tai vähemmän yleiseksi suhteessa saman geenin eri muotoihin. Variaatio katoaa kun uusi alleeli saavuttaa fiksaation – se on joko kadonnut populaatiosta tai se on korvannut esivanhemmilta perityn geenimuodon täysin.[73]
Luonnonvalinta johtaa evoluutioon vain jos populaatiossa on riittävän paljon geneettistä variaatiota. Ennen Mendelin perinnöllisyyskokeita, yksi yleinen hypoteesi oli perinnöllinen sekoittuminen. Perinnöllinen sekoittuminen johtaisi kuitenkin nopeasti siihen, että geneettinen variaatio katoaisi, tehden luonnonvalinnan kautta tapahtuvasta evoluutiosta epätodennäköistä. Hardyn–Weinbergin laki tarjoaa ratkaisun sille, miten populaatiossa oleva variaatio pysyy yllä Mendelin sääntöjen mukaisesti. Alleelien frekvenssi (variaatiot geenissä) pysyy vakiona jos ei ole valintaa, mutaatiota, migraatiota tai geneettistä ajautumista.[74]
Variaatio tulee perimässä olevista mutaatioista, geenien uudelleenjärjestäytymisestä suvullisen lisääntymisen kautta ja populaatioiden välillä tapahtuvasta migraatiosta (geenivirtauksesta). Vaikka uutta variaatiota tulee jatkuvasti mutaatioiden ja geenivirtauksen kautta, suurin osa lajin perimästä on identtistä kaikissa kyseisen lajin yksilöissä.[75] Siitä huolimatta, suhteellisen pienikin muutos genotyypissä voi johtaa merkittäviin muutoksiin fenotyypissä: esimerkiksi simpanssien ja ihmisten perimässä on vain noin 5 % eroavaisuuksia.[76]
Mutaatio
- Pääartikkeli: Mutaatio

Mutaatiot ovat solun perimän DNA-jaksossa tapahtuvia muutoksia. Mutaatiot voivat olla vaikutuksettomia, muuttaa geenin tuotetta tai estää geenin toiminnan. Banaanikärpäsillä (Drosophila melanogaster) tehtyjen tutkimusten perusteella on ehdotettu, että jos mutaatio muuttaa geenin tuottamaa proteiinia, tämä tulee todennäköisesti olemaan vahingollista; noin 70 % näistä mutaatioista aiheuttavat haittaa ja loput ovat joko neutraaleja tai heikosti hyödyllisiä.[77]
Mutaatiot voivat aiheuttaa suurien kromosomin osien duplikaation eli kahdentumisen (yleensä rekombinaation kautta), mikä voi tuottaa ylimääräisiä kopioita geenistä perimään.[78] Ylimääräiset geenien kopiot ovat merkittävä raaka-ainelähde uusien geenien kehittymiselle.[79] Tämä on tärkeää, koska useimmat uudet geenit kehittyvät geeniperheissä jo olemassa olevista geeneistä, joilla on yhteiset esivanhemmat.[80] Esimerkiksi ihmisen silmä käyttää neljää geeniä tehdäkseen rakennelmia, jotka aistivat valoa: kolme värinäköön ja yksi hämäränäköön; kaikki neljä polveutuvat yhdestä ainoasta esivanhemman geenistä.[81]
Uusia geenejä voi kehittyä esivanhempien geenistä kun kahdentuneessa kopiossa tapahtuu mutaatio, ja geeni saa uuden funktion. Tämä prosessi on helpompaa kun geeni on kahdentunut koska se kasvattaa järjestelmän redundanssia; yksi parin geeneistä voi saada uuden funktion samalla kun toinen kopio jatkaa alkuperäistä funktiota.[82][83] Toisen tyyppiset mutaatiot voivat jopa kehittää kokonaan uusia geenejä aiemmin ei-koodaavasta DNA:sta.[84][85]
Uusia geenejä voi myös kehittyä useiden geenien pienten osien kahdentumisesta. Nämä osat voivat uudelleenjärjestäytyä muodostaen uusia yhdistelmiä, joilla on uusia funktioita.[86][87] Kun uusia geenejä muodostuu olemassa olevien osien uudelleenjärjestäytymisen kautta, proteiinidomeenit toimivat moduuleina joilla on yksinkertaiset itsenäiset funktiot. Nämä moduulit voivat sekoittua ja tuottaa uusia yhdistelmiä, joilla on uusia ja monimutkaisia funktioita.[88] Esimerkiksi polyketidisyntaasit ovat suuria entsyymejä, jotka tuottavat antibiootteja; ne sisältävät jopa sata itsenäistä domeenia, joista jokainen katalysoi yhden prosessissa olevan vaiheen, aivan kuten liukuhihnalla.[89]
Suvullinen lisääntyminen ja rekombinaatio
Suvuttomasti lisääntyvissä eliöissä, geenit periytyvät yhdessä – tai linkitettyinä – koska ne eivät voi sekoittua muiden eliöiden geenien kanssa lisääntymisen aikana. Sen sijaan suvullisten eliöiden jälkeläisillä on itsenäisen lajittelun kautta saavutetut satunnaiset sekoitukset vanhempien kromosomeista. Tähän liittyvän homologiseksi rekombinaatioksi kutsutun prosessin kautta, suvulliset eliöt voivat vaihtaa DNA:ta kahden vastaavan kromosomin välillä.[90] Rekombinaatio ja uudelleenjärjestely eivät muuta alleelien frekvenssejä, mutta muuttavat sen sijaan mitkä alleelit ovat vuorovaikutuksessa keskenään, tuottaen jälkeläisiä joilla on uusia alleeliyhdistelmiä.[91] Suvullinen lisääntyminen kasvattaa yleensä geneettistä variaatiota ja voi kasvattaa evoluution nopeutta.[92][93]

Suvullisen lisääntymisen kaksinkertaisesta kustannuksesta kirjoitti ensimmäisenä John Maynard Smith.[94] Ensimmäinen kustannus on se, että vain yksi kahdesta sukupuolesta voi tuottaa jälkeläisiä. Toinen kustannus on se, että suvullisesti lisääntyvä yksilö voi antaa vain 50 % omista geeneistään eteenpäin.[95] Tästä huolimatta, suvullinen lisääntyminen on ylivoimaisesti yleisin tapa lisääntyä useimpien aitotumaisten ja monisoluisten eliöiden keskuudessa. Punaisen Kuningattaren hypoteesia on käytetty selittämään suvullisen lisääntymisen merkittävyyden keinona mahdollistaa jatkuva evoluutio ja mukautuminen reaktiona muiden lajien koevoluutiolle jatkuvasti muuttuvassa ympäristössä.[95][96][97][98]
Geenivirta
- Pääartikkeli: Geenivirta
Geenivirta viittaa populaatioiden ja lajien välillä tapahtuvaan geenien siirtymiseen.[99] Se voi siksi olla uusi lähde populaation tai lajin sisäiselle variaatiolle. Geenivirran voi aiheuttaa populaatioiden välillä liikkuvat yksilöt, kuten hiirien liikkuminen sisämaan ja rannikkoalueiden populaatioiden välillä tai siitepölyn liikkuminen raskasmetalleja sietävien ja raskasmetalleja sietämättömien ruohopopulaatioiden välillä.
Risteymät ja horisontaaliset geeninsiirrot ovat lajien välisiä geeninsiirtoja. Horisontaalisella geeninsiirrolla viitataan geneettisen materiaalin siirtymiseen yhdeltä eliöltä toiselle, joka ei ole sen jälkeläinen; tämä on yleistä bakteerien keskuudessa.[100] Lääketieteessä, tämä edistää antibioottiresistenssin leviämistä; bakteeri voi nopeasti siirtää resistenssigeenit muihin lajeihin silloin kun se on hankkinut ne.[101] Horisontaalista geeninsiirtoa bakteereista aitotumaisiin kuten leivinhiivaan (Saccharomyces cerevisiae) ja Callosobruchus chinensis -nimiseen kovakuoriaiseen on tapahtunut.[102][103] Esimerkkinä suuremman mittakaavan siirroista ovat aitotumaiset Bdelloidea -nimiset rataseläimet, jotka ovat saaneet kirjon geenejä bakteereilta, sieniltä ja kasveilta.[104] Virukset voivat myös siirtää DNA:ta eliöiden välillä, mahdollistaen geenien siirtymisen jopa biologisten domeenien välillä.[105]
Suuren mittakaavan geeninsiirtoja on tapahtunut myös aitotumaisten solujen esivanhempien ja bakteerien välillä, kun viherhiukkaset ja mitokondriot tulivat aitotumaisiin soluihin. On mahdollista, että aitotumaiset itse saivat alkunsa bakteerien ja arkeonien välisistä horisontaalisista geeninsiirroista.[106]
Mekanismit

Synteettisen evoluutioteorian näkökulmasta, evoluutiota tapahtuu kun keskenään lisääntyvien yksilöiden populaatiossa olevien alleelien frekvensseissä tapahtuu muutoksia.[74] Esimerkkinä voidaan pitää koiperhosten populaatiossa olevan mustan värin alleelin yleistyminen. Luonnonvalinta, geneettinen ajautuminen, geneettinen liftaaminen, mutaatio ja geenivirta ovat mekanismeja, jotka voivat johtaa muutoksiin alleelien frekvensseissä.
Luonnonvalinta
- Pääartikkeli: Luonnonvalinta
- Katso myös: Kelpoisuus
Luonnonvalinnan kautta tapahtuva evoluutio on prosessi jonka kautta lisääntymistä edistävistä piirteistä tulee yleisempiä ja pysyvämpiä populaation peräkkäisissä sukupolvissa. Sitä on usein kutsuttu "itsestään selväksi" mekanismiksi koska se johtuu kolmesta yksinkertaisesta tosiasasta:[11]
- Eliöpopulaatioissa on variaatiota morfologian, fysiologian ja käyttäytymisen suhteen (fenotyyppinen variaatio).
- Eri piirteet antavat eri suuruiset eloonjäämisen ja lisääntymisen todennäköisyydet (differentiaalinen kelpoisuus).
- Piirteet voivat periytyä sukupolvelta toiselle (kelpoisuuden heritabiliteetti).
Jokaisessa sukupolvessa on enemmän eliöitä kuin niiden on mahdollista selviytyä ja lisääntyä. Nämä olosuhteet tuottavat eliöiden välistä kilpailua selviytymisestä ja lisääntymisestä. Niinpä eliöt, joilla on etulyöntiasema kilpailijoihinsa nähden piirteiden suhteen, ovat todennäköisempiä siirtämään kyseiset piirteet seuraavaan sukupolveen.[107]
Luonnonvalinnan keskeinen käsite on eliön kelpoisuus.[108] Kelpoisuus kuvaa eliön kykyä selviytyä ja tuottaa jälkeläisiä, mikä määrittää kuinka suuri sen geneettinen vaikutus on seuraavaan sukupolveen.[108] Kelpoisuus ei kuitenkaan ole sama asia kuin jälkeläisten kokonaismäärä: sen sijaan, kelpoisuudesta kertoo se osuus seuraavista sukupolvista, jotka kantavat eliön geenejä.[109] Esimerkiksi jos eliö voisi selviytyä ja tuottaa jälkeläisiä nopeasti, mutta sen jälkeläiset olisivat liian pieniä ja heikkoja selviytyäkseen, silloin kyseisellä eliöllä olisi pieni geneettinen vaikutus tuleviin sukupolviin ja sillä olisi siten matala kelpoisuus.[108]
Jos tietty alleeli kasvattaa eliön kelpoisuutta enemmän kuin muut saman geenin alleelit, silloin tämän alleelin yleisyys kasvaa populaation jokaisessa peräkkäisessä sukupolvessa. Sanotaan, että valinta suosii näitä piirteitä. Esimerkkeinä piirteistä, jotka voivat kasvattaa kelpoisuutta ovat tehostunut selviytyminen ja lisääntynyt hedelmällisyys. Käänteisesti, vähemmän hyödyllisestä tai vahingollisesta alleelista johtuva matalampi kelpoisuus johtaa kyseisen alleelin harvenemiseen — valinta toimii piirrettä vastaan.[110] On tärkeää huomata, että alleelin kelpoisuus ei ole kiinteä ominaisuus; jos ympäristö muuttuu, aiemmin neutraaleista tai vahingollisista piirteistä voi tulla hyödyllisiä ja aiemmin hyödyllisistä piirteistä haitallisia.[63] Jos valinnan suunta kuitenkin muuttuu tällä tavoin, piirteet jotka olivat aiemmin kadonneet eivät voi kehittyä uudelleen identtisesti (katso Dollon laki).[111][112]

· Kaavio 1 esittää suuntaavaa valintaa, jossa yhden ääripään fenotyyppiä suositaan.
· Kaavio 2 esittää tasapainottavaa valintaa, jossa keskimmäistä fenotyyppiä suositaan enemmän kuin ääripään fenotyyppejä.
· Kaavio 3 esittää hajottavaa valintaa, jossa kummankin ääripään fenotyyppejä suositaan enemmän kuin keskimmäistä fenotyyppiä.
Populaation sisäinen luonnonvalinta korkeuden kaltaista jatkumolla vaihtelevaa piirrettä kohtaan voidaan luokitella kolmeen tyyppiin. Ensimmäinen on suuntaava valinta, joka on ajan myötä tapahtuva muutos piirteen keskimääräisessä arvossa — esimerkiksi, eliöistä tulee hitaasti korkeampia.[113] Toinen on hajottava valinta, joka suosii piirteen ääripäitä ja johtaa usein kahden eri arvon yleistymiseen – valinta toimii keskiarvoa vastaan. Tämä tapahtuisi jos matalilla tai korkeilla eliöillä olisi etu, mutta ei keskikorkeilla. Viimeisenä on tasapainottava (eli stabiloiva) valinta, joka karsii piirteen kummankin ääripään arvoja ja johtaa keskiarvon ympärillä olevan varianssin ja monimuotoisuuden pienenemiseen.[107][114] Tämä johtaisi esimerkiksi siihen, että eliöistä tulisi hitaasti saman korkuisia. Tasapainottava valinta usein vähentää populaation geneettistä monimuotoisuutta ja voi siten heikentää populaation mahdollisuuksia mukautua muuttuviin olosuhteisiin.
Seksuaalivalinta on erityinen luonnonvalinnan tapaus, jossa valinta kasvattaa eliön houkuttelevuutta mahdollisille parittelukumppaneille lisääntymisen onnistumisen kasvattamiseksi.[115] Seksuaalivalinnan kautta kehittyneet piirteet ovat erityisen esillä useiden eläinlajien urosten keskuudessa. Vaikka ne olisivat seksuaalisesti suosiossa, hankalien sarvien, parittelukutsujen, suurten ruumiinkokojen ja kirkkaiden värien kaltaiset piirteet vetävät usein puoleensa saalistajia, mikä vaarantaa yksittäisten urosten kyvyn selviytyä.[116][117] Tämä haitta selviytymiselle tasapainottuu näiden (seksuaalivalinnan kautta saavutettujen) vaikeasti väärennettävissä olevien piirteiden omaavien urosten suuremmalla lisääntymiskyvyllä.[118]
Luonnonvalinta yleisesti ottaen tekee luonnosta mitan sille, miten todennäköisesti tietyt yksilöt tai yksilölliset piirteet selviytyvät. "Luonnolla" viitataan tässä tapauksessa yksilön ympäristössä olevaan ekosysteemiin. Kukin tietyssä ekosysteemissä oleva populaatio täyttää tietyn ekologisen lokeron, tai aseman, jolla on tietyt vuorovaikutussuhteet muihin saman ekosysteemin osiin. Näihin suhteisiin vaikuttaa eliön elämänhistoria, asema ravintoketjussa ja maantieteellinen sijainti. Laaja ymmärrys luonnosta auttaa tieteilijöitä erottamaan tiettyjä tekijöitä, jotka yhdessä muodostavat luonnonvalinnan.
Luonnonvalinta voi toimia eri tasoilla, kuten geenien, solujen, yksittäisten eliöiden, eliöryhmien ja lajien tasolla.[119][120][121] Valinta voi toimia monella tasolla yhtäaikaisesti.[122] Esimerkki valinnasta jota tapahtuu yksittäisen eliön tason alapuolella on hyppiöksi (engl. transposon) kutsuttu geeni, joka voi replikoitua ja levitä koko perimään.[123] Yksittäisen eliön tason yläpuolella oleva valinta, kuten ryhmävalinta, voi johtaa yhteistyön kehittymiseen.[124]
Mutaatiotaipumus
Sen lisäksi, että mutaatiot ovat merkittävä lähde variaatiolle, ne voivat myös toimia evoluution mekanismina silloin kun molekyylitasolla on olemassa eri todennäköisyydet mutaatioiden tapahtumiselle – tätä ilmiötä kutsutaan mutaatiotaipumukseksi (engl. mutation bias).[125] Otetaan esimerkiksi kaksi genotyyppiä: yksi, jolla on nukleotidi G ja toinen, jolla on nukleotidi A samassa kohdassa. Jos näillä kahdella genotyypillä on sama kelpoisuus, mutta mutaatio G:stä A:han tapahtuu useammin kuin mutaatio A:sta G:hen, silloin on taipumus kehittyä genotyyppejä joilla on A.[126] Eri taipumukset liittymä- tai häviämämutaatioihin eri taksoneissa voivat johtaa erikokoisten perimien kehittymiseen.[127][128] Kehittymiseen ja mutaatioon liittyviä taipumuksia on havaittu myös morfologisessa evoluutiossa.[129][130] Esimerkiksi Baldwinin efektin mukaan mutaatiot voivat ajan myötä aiheuttaa ympäristön tuottamien piirteiden geneettisen assimilaation.[131][132]
Mutaatiotaipumuksen vaikutukset ovat päällekkäisiä muiden prosessien kanssa. Jos valinta suosii molempia mahdollisia mutaatioita samoissa määrin, mutta molempien mutaatioiden yhtäaikaisesta ilmenemisestä ei ole hyötyä, mutaatio joka ilmenee useimmin on se, joka tulee todennäköisimmin fiksoitumaan populaatiossa vallitsevaksi.[133][134] Geenin funktion menetykseen johtavat mutaatiot ovat paljon yleisempiä kuin mutaatiot, jotka tuottavat uuden, täysin toimivan geenin. Useimmat funktion menetykseen johtavat mutaatiot karsiutuvat valinnan kautta. Silloin kuitenkin kun valinta on heikkoa, taipumus funktion menetykseen voi vaikuttaa evoluutioon.[135] Esimerkiksi pigmenteistä ei enää ole hyötyä kun eläimet elävät luolien pimeydessä, joten ne yleensä katoavat.[136] Tämänkaltainen funktion menetys voi tapahtua mutaatiotaipumuksen takia ja/tai koska funktiolla oli hinta; kun funktiosta saatu hyöty katosi, luonnonvalinta johti sen menetykseen. Bacillus subtilis -nimisen bakteerin itiöintikyvyn menetys laboraatiossa tapahtuvan evoluution kautta johtuu ilmeisesti mutaatiotaipumuksesta eikä itiöintikyvyn ylläpitokustannusta vastaan tapahtuvasta luonnonvalinnasta.[137] Silloin kun ei ole funktion menetykseen johtavaa valintaa, menetyksen kehitysnopeus riippuu enemmän mutaationopeudesta kuin efektiivisestä populaatiokoosta.[138] Tämä viittaa siihen, että sitä ohjaa enemmän mutaatiotaipumus kuin geneettinen ajautuminen.
Geneettinen ajautuminen
- Pääartikkeli: Geneettinen ajautuminen

Geneettinen ajautuminen viittaa populaation alleelifrekvenssien muuttumiseen sukupolvesta toiseen alleelien otantavirhealttiuden takia.[139] Alleelifrekvensseillä on tämän seurauksena taipumus "ajautua" ylös tai alas sattumanvaraisesti (satunnaiskulkuna) silloin kun valintaa ohjaavia voimia ei ole tai ne ovat suhteellisen heikkoja. Tämä ajautuminen loppuu kun alleeli fiksoituu – joko katoamalla populaatiosta tai korvaamalla muut alleelit täysin. Geneettinen ajautuminen voi siten eliminoida joitain alleeleja populaatiosta pelkästään sattuman takia. Valintaa ohjaavien voimien puutteessakin, geneettinen ajautuminen voi aiheuttaa kaksi erillistä populaatiota (jotka aloittivat samalla geneettisellä koostumuksella) ajautumaan kahdeksi poikkeavaksi populaatioksi, joilla on eri alleeliryhmät.[140]
Valinnan ja luonnollisten prosessien (mukaan lukien geneettisen ajautumisen) suhteellista tärkeyttä on tavallisesti vaikeaa mitata.[141] Adaptiivisten ja ei-adaptiivisten voimien suhteelliset tärkeydet evoluutiomuutosten ajamisessa on nykyaikaisen tutkimuksen kohde.[142]
Molekyylievoluution neutraali teoria (engl. neutral theory of molecular evolution) ehdotti, että suurin osa evoluutiomuutoksista johtuvat neutraalien mutaatioiden fiksaatiosta geneettisen ajautumisen kautta.[13] Tästä seuraa, että kyseisen mallin mukaan suurin osa geneettisistä muutoksista johtuvat jatkuvasta mutaatiopaineesta ja geneettisestä ajautumisesta.[143] Tämä muoto neutraalista teoriasta on nyt suurimmaksi osaksi hylätty koska se ei vaikuta sopivan geneettiseen variaatioon, jota nähdään luonnossa.[144][145] On kuitenkin olemassa uudempi ja paremmin tukea saanut versio tästä mallista: Molekyylievoluution lähes neutraali teoria (engl. nearly neutral theory of molecular evolution) ehdottaa, että mutaatiot jotka ovat neutraaleja pienissä populaatioissa eivät välttämättä ole neutraaleja suurissa populaatioissa.[107] Muut vaihtoehtoiset teoriat ovat ehdottaneet, että geneettinen ajautuminen on mitätöntä verrattuna muihin stokastisiin evoluutiota ohjaaviin voimiin, kuten geneettiseen liftaamiseen (engl. genetic hitchhiking).[139][146][147]
Geneettisen ajautumisen kautta tapahtuvaan neutraalin alleelin fiksaatioon kuluva aika riippuu populaation koosta – fiksaatio tapahtuu nopeammin pienemmissä populaatioissa.[148] Populaatiossa olevien yksilöiden lukumäärä ei ole kriittinen, mutta efektiivisenä populaatiokokona (engl. effective population size) tunnettu suure on.[149] Efektiivinen populaatiokoko on tavallisesti pienempi kuin koko populaatio koska se ottaa huomioon tekijöitä kuten sisäsiittoisuuden yleisyyden ja populaation elinkaaren vaiheen, jossa sen koko on pienimmillään.[149] Efektiivinen populaatiokoko ei välttämättä ole sama kaikille samassa populaatiossa oleville geeneille.[150]
Geneettinen liftaaminen
Rekombinaatio mahdollistaa samalla DNA-juosteella olevien alleelien erottumisen. Rekombinaatioita tapahtuu kuitenkin harvoin (noin kaksi tapahtumaa per kromosomi per sukupolvi). Tämän vuoksi kromosomissa lähekkäin olevat geenit eivät aina sekoitu kauas toisistaan; geenit jotka ovat lähekkäin usein periytyvät yhdessä, koska juosteen katketessa ne jäävät todennäköisesti sen samaan osaan. Ilmiötä kutsutaan geneettiseksi kytkennäksi (engl. genetic linkage).[151] Taipumusta tähän mitataan löytämällä kuinka usein kaksi alleelia löydetään yhdessä yksittäiseltä kromosomilta verrattuna odotuksiin – tätä kutsutaan alleelien kytkentäepätasapainoksi. Ryhmänä periytyvää alleeliryhmää kutsutaan haplotyypiksi. Tämä voi olla tärkeätä silloin kun tietyssä haplotyypissä olevasta yksittäisestä alleelista on paljon hyötyä: luonnonvalinnan kautta tapahtuva valintapyyhkäisy (engl. selective sweep) voi johtaa muidenkin haplotyypissä olevien alleelien yleistymiseen populaatiossa; tätä ilmiötä kutsutaan geneettiseksi liftaamiseksi (engl. genetic hitchhiking).[152] Jotkut neutraalit geenit ovat geneettisesti kytkeytyneinä muihin valinnan alaisiin geeneihin. Tästä johtuva geneettinen liftaaminen voidaan osittain havaita sopivalla efektiivisellä populaatiokoolla.[146]
Geenivirta
Geenivirta viittaa populaatioiden ja lajien väliseen geenien siirtymiseen.[99] Geenivirran läsnäololla tai puuttumisella on merkittävä vaikutus evoluution kulkuun. Eliöiden monimutkaisuuden takia, missä tahansa kahdessa täysin eristetyssä populaatiossa tulee ajan myötä kehittymään geneettisiä yhteensopivuusongelmia neutraalien prosessien kautta (esim. Bateson-Dobzhansky-Muller -mallin mukaan[153]) vaikka molemmat populaatiot pysyisivät pohjimmiltaan identtisinä suhteessa niiden adaptaatioon ympäristön suhteen.
Jos geneettistä erilaisuutta kehittyy populaatioiden välille, populaatioiden välinen geenivirta voi tuottaa piirteitä tai alleeleja jotka ovat haitallisia paikallisessa populaatiossa. Tämä voi johtaa siihen, että näiden populaatioiden sisällä olevissa eliöissä kehittyy mekanismeja jotka estävät lisääntymisen geneettisesti etäisten populaatioiden kanssa – johtaen lopulta uusien lajien ilmestymiseen. Yksilöiden välillä tapahtuva geneettisen informaation vaihto on siten olennaisen tärkeää biologisen lajikäsitteen kehittämiselle.
Modernin synteesin kehityksen yhteydessä, Sewall Wright kehitti vaihtuvan balanssin teorian (engl. shifting balance theory), jonka mukaan geenivirta osittain eristettyjen populaatioiden välillä olisi tärkeä osa adaptiivista evoluutiota.[154] Viime aikoina on kuitenkin ollut merkittävää kritiikkiä sille kuinka tärkeä vaihtuvan balanssin teoria on.[155]
Lopputulokset
Evoluutio vaikuttaa eliöiden jokaiseen muodolliseen ja käytökselliseen osa-alueeseen. Näkyvimpiä ovat tietyt käytökselliset ja fyysiset adaptaatiot, jotka ovat luonnonvalinnan lopputuloksia. Nämä adaptaatiot kasvattavat kelpoisuutta auttamalla ruuan etsinnän, petoeläinten välttämisen tai parittelukumppanin houkuttelun kaltaisten toimintojen suorittamisessa. Eliöt voivat myös reagoida valintaan tekemällä yhteistyötä toistensa kanssa, yleensä auttamalla sukulaisia tai osallistumalla molempia osapuolia hyödyntävään symbioosiin. Pidemmällä aikavälillä evoluutio tuottaa uusia lajeja jakamalla vanhoja eliöpopulaatioita uusiin ryhmiin, jotka eivät voi tai eivät halua risteytyä.
Nämä evoluution lopputulokset luokitellaan aikaskaalan perusteella makroevoluutioon ja mikroevoluutioon. Makroevoluutio viittaa evoluutioon, joka tapahtuu lajien tasolla tai niiden yläpuolella olevilla tasoilla – erityisesti lajiutumiseen ja sukupuuttoon.[156] Mikroevoluutio viittaa pienempiin lajin tai populaation sisäisiin evoluutiomuutoksiin – erityisesti alleelifrekvenssien muutoksiin ja adaptaatioon.[156] Yleisesti ottaen makroevoluutiota pidetään pitkien mikroevoluution jaksojen lopputuloksena.[157] Koska mikroevolutiiviset muutokset kasautuvat makroevoluutioksi, ei mikro- ja makroevoluution välinen ero siten ole perustavanlaatuinen – ero on yksinkertaisesti aika, joka on kulunut.[158] On kuitenkin huomattava, että makroevoluutiossa koko lajin yhteiset piirteet voivat olla tärkeitä. Esimerkiksi, suuri määrä variaatiota yksilöiden välillä mahdollistaa sen, että laji voi nopeasti sopeutua uusiin elinympäristöihin – pienentäen sukupuuton todennäköisyyttä. Toisaalta, laaja maantieteellinen levinneisyys voi kasvattaa lajiutumisen todennäköisyyttä kasvattamalla todennäköisyyttä sille, että osa populaatiosta eristäytyy. Tässä mielessä, mikroevoluutioon ja makroevoluutioon voi sisältyä valintaa eri tasoilla. Mikroevoluutio vaikuttaa geeneihin ja eliöihin, kun taas makroevoluution prosessit (kuten lajivalinta) vaikuttavat kokonaisiin lajeihin vaikuttamalla niiden lajiutumisien ja sukupuuttojen nopeuksiin.[159][160][161]
Yleinen väärinkäsitys on, että evoluutiolla on tavoitteita tai pitkän aikavälin suunnitelmia. Realistisesti evoluutiolla ei ole pitkän aikavälin tavoitteita eikä välttämättä tuota suurempaa monimutkaisuutta.[162][163] Evoluution tuloksia ei tule sekoittaa tavoitteellisuuteen. Vaikka monimutkaisia lajeja on kehittynyt, ne ovat sivutuote siitä, että eliöiden kokonaismäärä kasvaa – yksinkertaiset elämänmuodot ovat edelleen yleisimmät biosfäärissä.[164] Valtaosa lajeista ovat mikroskooppisia esitumallisia, jotka muodostavat noin puolet maailman biomassasta niiden pienestä koosta riippumatta[165] ja muodostavat valtaosan maapallon biodiversiteetistä.[166] Yksinkertaiset eliöt ovat olleet dominoiva elämänmuoto maapallolla kautta sen historian ja ne jatkavat pääasiallisena elämänmuotona tänäkin päivänä.[167] Monimutkaiset elämänmuodot vaikuttavat monimuotoisemmilta vain koska ne ovat helpommin huomattavissa.[167] Tosiaan, mikrobien evoluutio on erityisen tärkeää modernille evoluutiotutkimukselle, koska niiden nopea lisääntyminen mahdollistaa kokeellisen evoluution tutkimisen ja evoluution ja adaptaation havainnoinnin reaaliaikaisesti.[168][169]
Adaptaatio
- Pääartikkeli: Adaptaatio

Adaptaatio on prosessi joka tekee eliöistä paremmin sopeutuneita elinympäristöihinsä.[170][171] Adaptaatio voi myös viitata piirteeseen, joka on tärkeä eliön selviytymiselle. Esimerkkinä hevosten hampaiden adaptaatio ruohon jauhontaan. Voimme erottaa sanan kaksi merkitystä käyttämällä adaptaatiota viittaamaan evoluutioprosessiin ja adaptiivista piirrettä prosessin tuotteeseen (ruumiinosaan tai funktioon). Adaptaatiot ovat luonnonvalinnan tuottamia.[172] Seuraavat määritelmät johtuvat Theodosius Dobzhanskyn työstä:
- Adaptaatio on evolutiivinen prosessi, jossa eliöstä tulee kyvykkäämpi elämään sen ympäristössä tai ympäristöissä.[173]
- Adaptoituneisuus on adaptaation tila: missä määrin eliö pystyy elämään ja lisääntymään tietyissä ympäristöissä.[174]
- Adaptiivinen piirre on eliön kehityskuvion ominaisuus, joka mahdollistaa tai parantaa todennäköisyyttä, että eliö säilyy hengissä ja lisääntyy.[175]
Adaptaatio voi johtaa joko uuden piirteen hankkimiseen tai esivanhemmilta perityn piirteen menetykseen. Esimerkki, joka havainnollistaa kummankin näistä muutoksista, on bakteerien adaptaatio antibioottivalintaan, jossa geneettiset muutokset aiheuttavat antibioottiresistenssin muuttamalla antibiootin kohdetta tai nostamalla antibioottia ulos solusta kuljettavien mekanismien tehokkuutta.[176] Muita merkittäviä esimerkkejä ovat kolibakteerien (Escherichia coli) kehittyminen käyttämään sitruunahappoa ravintoaineena pitkään kestäneessä laboratoriokokeessa;[177] Flavobacterium-nimisessä bakteerissa kehittynyt uudenlainen entsyymi, joka mahdollistaa bakteerin kasvamisen nailonin valmistuksessa syntyvien sivutuotteiden päällä;[178][179] ja maaperästä löytyvä Sphingobium -niminen bakteeri, jolla on kehittynyt täysin uudenlainen metaboliareitti, joka hajottaa torjunta-aineena käytettyä synteettistä pentakloorifenolia.[180][181] Mielenkiintoinen mutta edelleen kiistanalainen ajatus on, että jotkut adaptaatiot voisivat kasvattaa eliön kykyä tuottaa variaatiota ja adaptoitua luonnonvalinnan kautta, kasvattaen eliön kehittymiskykyä.[182][183][184][185][186]

Adaptaatio tapahtuu olemassa olevien rakenteiden asteittaisen muuttumisen kautta. Tästä seuraa, että sukua olevien lajien rakenteilla – joilla on samanlaiset sisäiset järjestäytymiset – voi olla eri funktiot. Tämä johtuu yksittäisten esivanhemmilta perittyjen rakenteiden adaptoimisesta eri tarkoituksiin. Esimerkiksi lepakoiden siivissä olevat luut ovat erittäin samanlaisia kuin hiirien jaloissa ja kädellisten käsissä olevat luut, johtuen siitä, että nämä ovat kaikki polveutuneet yhteisestä nisäkäsesivanhemmasta.[188] Koska kaikki elossa olevat lajit ovat kuitenkin jossain määrin sukua toisilleen,[189] elimet, joilla ei vaikuta olevan minkäänlaista rakenteellista samankaltaisuutta (kuten niveljalkaisten, kalmarien ja selkärankaisten silmät tai niveljalkaisten ja selkärankaisten raajat ja siivet) voivat olla riippuvaisia yhteisistä homologisista geeneistä, jotka säätelevät niiden rakenteita ja funktioita; tätä kutsutaan syväksi homologiaksi (engl. deep homology).[190][191]
Jotkut rakenteet voivat evoluution aikana menettää alkuperäisen funktionsa ja muuttua surkastumiksi.[192] Sellaisilla rakenteilla voi olla vähän jos lainkaan tehtävää nykyisissä lajeissa, mutta niillä voi olla selkeä funktio esivanhemmissa tai muissa läheistä sukua olevissa lajeissa. Esimerkkeinä ovat pseudogeenit,[193] ei-toimivat silmien jäänteet sokeissa luolakaloissa,[194] lentokyvyttömien lintujen siivet[195] ja lonkkaluiden olemassaolo valaissa ja käärmeissä.[187] Esimerkkejä ihmisten surkastumista ovat muun muassa viisaudenhampaat,[196] häntäluu,[192] umpilisäke[192] ja muita käyttäytymiseen liittyviä surkastumia kuten kananlihalle meneminen[197][198] ja primitiiviset refleksit.[199][200][201]
Kuitenkin, monet yksinkertaisilta adaptaatioilta vaikuttavat piirteet ovat itse asiassa eksaptaatioita: alun perin yhteen tarkoitukseen adaptoituja rakenteita, joista on sattumalta tullut jokseenkin käytännöllisiä muihin tarkoituksiin ajan myötä.[202] Esimerkkinä on Holaspis guentheri -niminen Afrikkalainen lisko, jossa kehittyi erittäin litteä pää, jotta se voisi piiloutua halkeamiin. Tämä voidaan päätellä tarkastelemalla kyseisen liskon lähisukulaisia. Holaspis guentherin päästä on kuitenkin tullut niin litteä, että siitä on hyötyä puusta puuhun liitämisessä – eksaptaatio.[202] Solujen sisällä, bakteeriflagellien[203] ja proteiinien lajittelukoneiden[204] kaltaiset molekyylikoneet kehittyivät rekrytoimalla useita jo olemassa olevia proteiineja, joilla oli aiemmin erilaiset tarkoitukset.[156] Toinen esimerkki on glykolyysissä ja vierasaineiden aineenvaihdunnassa käytettyjen entsyymien rekrytoiminen eliöiden silmälinsseihin toimimaan kristalliiniksi kutsuttuina rakenteellisina proteiineina.[205][206]
Eräs evolutiivisen kehitysbiologian nykyisistä tutkimuskohteista on adaptaatioiden ja eksaptaatioiden kehityksellinen perusta.[207] Tämä tutkimustyö käsittelee alkionkehityksen alkuperää ja evoluutiota ja miten muutokset kehityksessä ja kehitystä ohjaavissa prosesseissa tuottavat uusia ominaisuuksia.[208] Nämä tutkimukset ovat osoittaneet, että evoluutio voi muuttaa kehitystä tuottaakseen uusia rakennelmia. Esimerkkinä ovat alkion luuston rakenteet, jotka kehittyvät leuoiksi muissa eläimissä, muodostavat mieluummin osan välikorvaa nisäkkäissä.[209] On myös mahdollista, että rakenteet jotka ovat kadonneet evoluution myötä voivat ilmestyä uudelleen kehitystä ohjaavissa geeneissä tapahtuvien muutoksien takia. Esimerkkinä ovat mutaatiot kanoissa, jotka aiheuttavat, että niiden alkioissa kasvaa krokotiilien hampaita muistuttavia hampaita.[210] Nyt on tulossa selväksi, että useimmat eliöiden muodoissa tapahtuvat muutokset johtuvat muutoksista, jotka tapahtuvat pienessä säilyneiden geenien joukossa.[211]
Koevoluutio

- Pääartikkeli: Koevoluutio
Eliöiden väliset vuorovaikutukset voivat tuottaa sekä konfliktia että yhteistyötä. Kun vuorovaikutus on kahden lajin välillä (esimerkiksi patogeenin ja isäntäeliön tai saalistajan ja saaliin välillä), näissä lajeissa voi kehittyä toisiaan vastaavat adaptaatiot. Tässä tapauksessa, yhden lajin evoluutio aiheuttaa adaptaatioita toisessa lajissa. Nämä jälkimmäisessä lajissa tapahtuvat muutokset taas aiheuttavat adaptaatioita ensimmäisessä lajissa. Tätä valinnan ja reaktion kiertokulkua kutsutaan koevoluutioksi.[212] Esimerkkinä on tetrodotoksiinin tuotto länsirannikonsalamanterissa (Taricha granulosa) ja tetrodotoksiinin vastustuskyvyn kehittyminen sen saalistajassa, Thamnophis sirtalis -nimisessä käärmeessä. Tässä saalistaja-saalis -kaksikossa, evolutiivinen kilpavarustelu (engl. evolutionary arms race) on tuottanut suuren määrän myrkkyä salamanterissa ja vastaavasti korkean vastustuskyvyn käärmeessä.[213]
Yhteistyö
Kaikkiin lajien välisiin koevolutiivisiin vuorovaikutuksiin ei liity konfliktia.[214] Lukuisia tapauksia molempia osapuolia hyödyttävistä vuorovaikutussuhteista on kehittynyt. Esimerkkinä on äärimmäinen yhteistyö, joka on olemassa kasvien ja sienijuuren välillä. Sienijuuri kasvaa kasvin juuristossa ja auttaa sitä saamaan maaperästä paremmin ravinteita.[215] Tämä on vastavuoroinen suhde, jossa kasvit antavat sienille yhteyttämisen kautta tuotettuja sokereita. Tässä tapauksessa, sienet itse asiassa kasvavat kasvisolujen sisällä, antaen niiden vaihtaa ravinteita isäntäeliön kanssa samalla kun ne lähettävät signaaleja kasvin immuunijärjestelmän suppressoimiseksi.[216]
Saman lajin sisäisten eliöiden yhteenlyöttäytymisiä on myös kehittynyt. Äärimmäinen tapaus on mehiläisten, termiittien ja muurahaisten kaltaisissa sosiaalisissa hyönteisissä esiintyvä aitososiaalisuus, jossa lisääntymiskyvyttömät hyönteiset syöttävät ja vartioivat yhdyskunnan pienempää lisääntymiskykyistä jäsenkuntaa. Pienemmässä mittakaavassa, somaattiset solut (jotka muodostavat eläimen kehon) rajoittavat omaa lisääntymistään jotta ne voivat ylläpitää vakaata eliötä, joka vastaavasti ylläpitää pienempää lukumäärää sukusoluja jälkeläisten tuottamista varten. Tässä tapauksessa, somaattiset solut reagoivat tiettyihin signaaleihin, jotka ohjaavat niitä kasvamaan, pysymään samanlaisina tai kuolemaan. Jos solut eivät huomioi näitä signaaleja ja lisääntyvät sopimattomalla tavalla, niiden hallitsematon kasvu johtaa syövän kehittymiseen.[217]
Tämänkaltainen yhteistyö on voinut kehittyä sukulaisvalinnan prosessin kautta, jossa yksilö toimii sukulaisen jälkeläisten eduksi.[218] Valinta suosii tätä toimintaa koska jos auttava yksilö sisältää alleeleja, jotka korostavat auttamista, silloin on todennäköistä, että sen sukulaisella on myös nämä alleelit ja siten kyseiset alleelit kulkevat eteenpäin.[219] Muita prosesseja, jotka voivat kasvattaa yhteistyötä, ovat muun muassa ryhmävalinta, jossa yhteistyöstä on hyötyä eliöistä koostuvalle ryhmälle.[220]
Lajiutuminen
- Pääartikkeli: Lajiutuminen

Lajiutuminen on prosessi jonka myötä laji eroaa kahdeksi tai useammaksi lajiksi polveutumisen kautta.[221]
On lukuisia tapoja määritellä mikä on "laji". Määritelmän valinta riippuu tarkasteltavan lajin yksityiskohdista.[222] Esimerkiksi, jotkut lajikäsitteet ovat paremmin sovellettavissa suvullisen lisääntymisen kautta lisääntyviin eliöihin, kun taas toiset ovat paremmin sovellettavissa suvuttoman lisääntymisen kautta lisääntyviin eliöihin. Lajikäsitteet voidaan monipuolisuudestaan huolimatta luokitella kolmeen laajaan filosofiseen lähestymistapaan: keskenään lisääntyviin, ekologisiin ja fylogeneettisiin.[223] Biologinen lajikäsite on klassinen esimerkki lähestymistavasta, jonka mukaan eliöt kuuluvat samaan lajiin, jos ne pystyvät luonnonoloissa tuottamaan lisääntymiskykyisiä jälkeläisiä keskenään. Ernst Mayrin vuonna 1942 määrittelemä biologinen lajikäsite sanoo, että "lajit ovat luonnollisia populaatioryhmiä, jotka todellisuudessa tai potentiaalisesti lisääntyvät keskenään ja ovat lisääntymiskyvyllisesti eristettyinä toisista samankaltaisista ryhmistä".[224] Huolimatta sen pitkään jatkuneesta ja laajamittaisesta käytöstä, biologinen lajikäsite ei muiden lajikäsitteiden tavoin ole ongelmaton – esimerkkinä on se, että näitä käsitteitä ei voida soveltaa esitumallisiin[225] – ja tätä kutsutaan lajiongelmaksi (engl. species problem).[222] Jotkut tutkijat ovat yrittäneet monistisen lajikäsitteen kehittämistä – toiset taas ovat omaksuneet pluralistisen lähestymistavan ja ehdottaneet, että voi olla olemassa eri tapoja, joilla lajin määritelmää voidaan tulkita loogisesti.[222][223]
Kahden eriävän suvullisesti lisääntyvän populaation välillä on oltava isolaatiomekanismeja, jotta populaatioista voisi tulla kaksi uutta lajia. Geenivirta voi hidastaa tätä prosessia levittämällä uusia geneettisiä variaatioita muihin populaatioihin. Riippuen siitä kuinka paljon lajit ovat eriytyneet toisistaan viimeisen yhteisen esivanhemman jälkeen, on edelleen mahdollista, että ne voivat tuottaa jälkeläisiä – esimerkkinä muuliksi kutsuttu aasiorin ja hevostamman risteymä.[226] Sellaiset risteymät ovat pääsääntöisesti hedelmättömiä. Tässä tapauksessa, läheistä sukua olevat lajit voivat säännöllisesti risteytyä, mutta valinta toimii risteymiä vastaan ja lajit pysyvät erillisinä. On kuitenkin huomattava, että hedelmällisiä risteymiä voi silloin tällöin syntyä. Näillä uusilla lajeilla voi olla ominaisuuksia, jotka ovat niiden vanhempien fenotyyppien väliltä, tai niillä voi olla täysin uusi fenotyyppi.[227] Risteytymisen tärkeys uusien eläinlajien tuottamisessa on epäselvä, vaikka useita tapauksia on havaittu lukuisissa eläinlajeissa.[228] Hyla versicolor -niminen sammakko on erityisen paljon tutkittu esimerkki.[229]
Lajiutumista on havaittu lukuisia kertoja sekä hallituissa laboratorio-olosuhteissa että luonnossa.[230] Suvullisesti lisääntyvissä eliöissä, lajiutuminen johtuu lisääntymisesteen muodostumisesta ja sitä seuraavasta geneettisestä eriytymisestä. Lajiutumiselle on olemassa neljä mekanismia. Eläinten keskuudessa yleisin on allopatrinen lajiutuminen, jota tapahtuu kun populaatiot joutuvat maantieteellisesti eristyksiin, esimerkiksi elinympäristöjen pirstoutumisen (engl. habitat fragmentation) tai migraation kautta. Näissä olosuhteissa tapahtuva valinta voi tuottaa erittäin nopeita muutoksia eliöiden ulkoasuissa ja käyttäytymisissä.[231][232] Valinta ja ajautuminen toimivat itsenäisesti populaatioissa, jotka ovat eristettyinä muista saman lajin populaatioista. Eristyksissä oleminen voi sen takia tuottaa eliöitä, jotka eivät enää voi risteytyä.[233]
Toinen lajiutumisen mekanismi on peripatrinen lajiutuminen, joka tapahtuu kun pienet eliöpopulaatiot joutuvat eristyksiin uudessa ympäristössä. Tämä eroaa allopatrisesta lajiutumisesta siinä, että eristetyt populaatiot ovat luvullisesti paljon pienempiä kuin isäntäpopulaatio. Tässä tapauksessa perustajavaikutus aiheuttaa nopeaa lajiutumista sen jälkeen kun sisäsiittoisuuden kasvu kasvattaa homotsygootteihin kohdistuvaa valintaa, joka johtaa nopeisiin perinnöllisiin muutoksiin.[234]
Lajiutumisen kolmas mekanismi on parapatrinen lajiutuminen. Tämä on samanlainen kuin peripatrinen lajiutuminen siinä mielessä, että pieni populaatio siirtyy uuteen elinympäristöön, mutta eroaa siinä, että näiden kahden populaation välillä ei ole fyysistä estettä. Sen sijaan, lajiutuminen johtuu näiden kahden populaation välistä geenivirtaa vähentävien mekanismien kehittymisestä.[221] Yleensä tämä tapahtuu kun isäntäpopulaation elinympäristössä on tapahtunut raju muutos. Yksi esimerkki on heinäkasvi tuoksusimake (Anthoxanthum odoratum), joka voi lajiutua parapatrisen lajiutumisen kautta reaktiona kaivoksista johtuvalle paikalliselle metallisaastumiselle.[235] Tässä tapauksessa kehittyy kasveja, joilla on korkea sietokyky maaperässä olevia metalleja kohtaan. Valinta toimi metalleja sietämättömien kasvien kanssa risteytymistä vastaan, mikä johti siihen, että näiden kahden populaation kukinta-ajat eriytyivät asteittain. Tämä tuotti ajan myötä täydellisen lisääntymisisolaatioon. Kahden populaation risteymiä vastaan toimiva valinta voi johtaa lajin sisäistä parittelua tukevien piirteiden kehittymiseen ja lajien ulkonäölliseen erilaistumiseen.[236]

Viimeisenä on sympatrinen lajiutuminen, jossa lajit eriytyvät ilman maantieteellistä eristäytymistä tai elinympäristössä tapahtuvia muutoksia. Tämä lajiutumisen muoto on harvinainen, sillä pienikin geenivirtauksen määrä voi poistaa geneettiset erilaisuudet osasta populaatiota.[237] Eläimien sympatrinen lajiutuminen vaatii yleensä perinnöllisten erilaisuuksien ja valikoivan parinmuodostuksen kehittymisen, jotta lisääntymiseste voisi kehittyä.[238]
Yhteen sympatrisen lajiutumisen muodoista liittyy kahden läheistä sukua olevien lajien risteytyminen tuottaakseen uuden risteymälajin. Tämä ei ole yleistä eläimien keskuudessa koska eläinristeymät ovat yleensä lisääntymiskyvyttömiä. Tämä johtuu siitä, että meioosin aikana kummankin vanhemman homologiset kromosomit ovat eri lajeista eivätkä siksi voi pariutua onnistuneesti. Tästä huolimatta, se on yleisempää kasvien keskuudessa koska kasvit yleensä kaksinkertaistavat kromosomiensa lukumäärän, muodostaen polyploideja.[239] Tämä mahdollistaa sen, että kummankin vanhemman kromosomit voivat muodostaa vastaavat parit meioosin aikana.[240] Esimerkkinä tämänkaltaisesta lajiutumisesta on kun lituruoho (Arabidopsis thaliana) ja hietapitkäpalko (Arabidopsis arenosa) risteytyivät tuottaakseen uuden ruotsinpitkäpaloksi (Arabidopsis suecica) kutsutun lajin.[241] Tämä tapahtui noin 20 000 vuotta sitten.[242] Kyseinen lajiutuminen on toistettu laboratoriossa, mahdollistaen prosessissa mukana olleiden geneettisten mekanismien tutkimisen.[243] Kromosomien lukumäärän kaksinkertaistaminen tietyissä lajeissa voi siten olla yleinen lisääntymisisolaation aiheuttaja, sillä puolet kaksinkertaistuneista kromosomeista ovat pariutumattomia silloin kun laji risteytyy eliön kanssa, joka ei ole kaksinkertaistanut kromosomiensa lukumäärää.[244]
Lajiutumiset ovat tärkeitä jaksoittaisen tasapainon mallille, joka selittää fossiiliaineistossa olevat lyhyet evolutiiviset "pyrähdykset" suhteellisten pitkien suvantovaiheiden välissä, jolloin muutosta ei juurikaan tapahdu.[245] Tämän mallin mukaan, lajiutuminen ja nopea evoluutio ovat linkittyneitä – luonnonvalinta ja geneettinen ajautuminen vaikuttavat voimakkaimmin lajeihin, jotka ovat kesken uusissa elinympäristöissä tai pienissä populaatioissa tapahtuvaa lajiutumista. Tämän seurauksena, fossiiliaineiston suvantovaiheet vastaavat isäntäpopulaatioita ja eliöt, jotka ovat kesken lajiutumista ja nopeaa evoluutiota ovat pienissä populaatioissa tai rajoitetuissa elinympäristöissä, joten niistä syntyy harvoin fossiileja.[160]
Sukupuutto
- Pääartikkeli: Sukupuutto

Sukupuutto viittaa kokonaisen lajin katoamiseen. Sukupuutot eivät ole epätavallisia tapahtumia, sillä lajeja jatkuvasti ilmestyy lajiutumisien kautta ja katoaa sukupuuttojen kautta.[246] Lähes kaikki eläin- ja kasvilajit, jotka ovat eläneet Maapallolla ovat nyt kuolleet sukupuuttoon[247] ja sukupuutto vaikuttaa olevan kaikkien lajien lopullinen kohtalo.[248] Näitä sukupuuttoja on tapahtunut jatkuvasti kautta elämän historian, vaikka niiden tiheys kasvaa ajoittaisissa joukkosukupuutoissa.[249] Liitukauden joukkosukupuutto, jonka aikana dinosaurukset kuolivat sukupuuttoon, on parhaiten tunnettu, mutta aikaisempi permikauden joukkotuho oli vielä vakavampi: noin 96 % lajeista kuolivat silloin sukupuuttoon.[249] Holoseenikauden joukkosukupuutto on meneillään oleva joukkosukupuuttotapahtuma, joka liittyy viimeisen muutaman tuhannen vuoden aikana tapahtuneeseen ihmiskunnan levittäytymiseen ympäri Maapalloa. Nykypäivän sukupuuttotiheys on 100–1000 kertaa suurempi kuin taustatiheys, ja jopa 30 % nykyisistä lajeista voivat olla kuolleita vuoteen 2050 mennessä.[250] Ihmistoimet ovat nyt pääasiallinen syy meneillään olevalle joukkosukupuuttotapahtumalle;[251] ilmaston lämpeneminen voi edelleen kiihdyttää joukkosukupuuttotapahtumaa tulevaisuudessa.[252]
Sukupuuttojen roolia evoluutiossa ei ymmärretä kovin hyvin ja tämä voi olla riippuvainen tarkastelun kohteena olevan sukupuuton tyypistä.[249] Jatkuvasti tapahtuvien "matalan tason" sukupuuttojen aiheuttajat voivat olla rajoitetuista resursseista johtuvan lajien välisen kilpailun tulos (syrjäyttävän kilpailun periaate).[57] Jos yksi laji voittaisi toisen tässä kilpailussa, se voisi johtaa lajivalintaan, jossa kelvollisempi laji selviytyy ja toinen laji ajautuu sukupuuttoon.[120] Ajoittain tapahtuvat joukkosukupuutot ovat myös merkittäviä, mutta sen sijaan, että ne toimisivat valintaa ohjaavina voimina, ne rajusti vähentävät variaatiota epäspesifisellä tavalla ja tukevat selviytyjien keskuudessa tapahtuvia nopean evoluution ja lajiutumisien pyrähdyksiä.[253]
Elämän kehityshistoria
Elämän alkuperä
Erittäin korkeaenergiaisen kemian on ajateltu tuottaneen itsereplikoituvan molekyylin noin 4 miljardia vuotta sitten. Puoli miljardia vuotta myöhemmin eli kaikkien elossa olevien eliöiden yhteinen esivanhempi.[3] Nykypäivän tieteellinen konsensus on, että elämää muodostava monimutkainen biokemia kehittyi yksinkertaisemmista kemiallisista reaktioista.[254] Elämän alkuun on voinut osallistua RNA:n kaltaisia itsereplikoituvia molekyylejä[255] ja yksinkertaisten solujen kokoonpanoja.[256]
Yhteinen polveutuminen
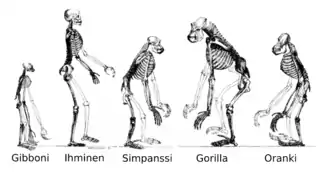
Kaikki maapallolla olevat eliöt ovat polveutuneet yhteisestä esivanhemmasta tai esivanhemmilta peritystä geenivarastosta.[189][257] Nykyiset lajit ovat vaihe evoluution prosessissa; lajien monimuotoisuus on pitkän lajiutumisien ja sukupuuttojen sarjan tuote.[258] Eliöiden yhteinen polveutuminen pääteltiin ensimmäisen kerran neljästä yksinkertaisesta tosiasiasta: Ensinnä, niillä on maantieteelliset jakaumat joita ei voida selittää paikallisella sopeutumisella. Toiseksi, elämän monimuotoisuus ei ole joukko täysin ainutlaatuisia eliöitä, mutta eliöitä joilla on morfologisia yhtäläisyyksiä. Kolmanneksi, surkastuneet piirteet ilman selkeää tarkoitusta muistuttavat toiminnallisia esivanhempien piirteitä. Viimeisenä, eliöt voidaan näiden yhtäläisyyksien perusteella luokitella (sukupuun kaltaiseen) sisäkkäisten ryhmien hierarkiaan.[259] Moderni tutkimus on kuitenkin ehdottanut, että tämä "elämän puu" voi olla monimutkaisempi kuin yksinkertainen haarautuva puu koska jotkut geenit ovat levinneet itsenäisesti etäistä sukua olevien lajien välillä horisontaalisten geeninsiirtojen kautta.[260][261]
Menneet lajit ovat myös jättäneet merkkejä omasta kehityshistoriastaan. Fossiilit ja nykypäivän eliöiden vertailtavissa olevat anatomiset piirteet luovat perustan morfologisen (eli anatomisen) kehityshistorian tutkimukselle.[262] Vertaamalla nykypäivän ja sukupuuttoon kuolleiden lajien anatomisia piirteitä, paleontologit voivat tehdä johtopäätöksiä näiden lajien sukulinjoista. Tämä lähestymistapa on kuitenkin menestyneintä eliöillä, joilla oli kovia ruumiinosia, kuten kuoria, luita ja hampaita. Lisäksi, koska esitumallisilla (kuten bakteereilla) ja arkeoneilla on rajoitettu määrä yhteisiä morfologisia piirteitä, niiden fossiilit eivät anna tietoa niiden sukujuurista.
Todisteita yhteisestä polveutumisesta on hiljattain tullut eliöiden biokemiallisten yhtäläisyyksien tutkimisesta. Esimerkiksi, kaikki elävät solut käyttävät samaa yksinkertaista nukleotidien ja aminohappojen joukkoa.[263] Molekyyligenetiikan kehittyminen on paljastanut merkit, jotka evoluutio on jättänyt eliöiden perimiin: mutaatioiden tuottaman molekyylikellon avulla voidaan määrittää milloin lajit erosivat toisistaan.[264] Esimerkiksi, DNA-jaksojen vertailun kautta tiedetään, että ihmiset ja simpanssit jakavat 98 % perimistään – eroavien osien tutkiminen antaa tietoa siitä, milloin näiden lajien yhteinen esivanhempi eli.[265]
Elämän kehittyminen

Esitumalliset elivät maapallolla noin 3–4 miljardia vuotta sitten.[267][268] Mitään merkittäviä muutoksia näiden eliöiden morfologiassa tai solujen järjestäytymisessä ei tapahtunut seuraavan muutaman miljardin vuoden aikana.[269] Aitotumaiset solut ilmestyivät 1,6–2,7 miljardia vuotta sitten. Seuraava merkittävä muutos solujen rakenteessa tuli kun aitotumaiset nielaisivat sisäänsä bakteereja, muodostaen niiden kanssa symbioottisen suhteen, jota kutsutaan endosymbioosiksi.[270][271] Nielaistut bakteerit ja isäntäsolut kehittyivät tämän jälkeen koevoluutiivisesti, ja bakteereista kehittyi joko mitokondrioita tai hydrogenosomeja.[272] Toinen nielaiseminen, joka kohdistui syanobakteerien kaltaisiin eliöihin, johti viherhiukkasten kehittymiseen levissä ja kasveissa.[273]
Elämän historia oli yksisoluisten aitotumaisten, esitumallisien ja arkeonien historiaa kunnes noin 610 miljoonaa vuotta sitten monisoluiset eliöt alkoivat ilmestyä Ediacarakauden merissä.[267][274] Monisoluisuuden evoluutio tapahtui lukuisten itsenäisten tapahtumien kautta eliöissä kuten sienieläimissä, ruskolevissä, syanobakteereissa, limasienissä ja limabakteereissa.[275]
Pian näiden monisoluisten eliöiden ilmestymisen jälkeen esiintyi huomattava määrä biologista monimuotoisuutta noin 10 miljoonaa vuotta kestäneen Kambrikauden lajiräjähdykseksi kutsutun tapahtuman aikana. Suurin osa nykyisten eläimien pääjaksoista ilmestyivät fossiilikerrostumiin tässä tapahtumassa.[276] Lukuisia syitä Kambrikauden lajiräjähdykselle on ehdotettu, mukaan lukien hapen kerääntyminen ilmakehään yhteyttämisen kautta.[277]
Noin 500 miljoonaa vuotta sitten, kasvit ja sienet asuttivat maan ja näitä seurasi pian niveljalkaiset ja muut eläimet.[278] Hyönteiset menestyivät erityisen hyvin ja vielä tänäkin päivänä muodostavat enemmistön eläinlajeista.[279] Sammakkoeläimet ilmestyivät ensimmäisen kerran noin 364 miljoonaa vuotta sitten. Näitä seurasi varhaiset vesikalvolliset ja linnut noin 155 miljoonaa vuotta sitten (kumpikin "matelijakaltaisista" suvuista), nisäkkäät noin 129 miljoonaa vuotta sitten, homininae noin 10 miljoonaa vuotta sitten ja nykyihminen noin 250 000 vuotta sitten.[280][281][282] On kuitenkin huomattava, että näiden suurten eläinten kehityksestä riippumatta, pienemmät eliöt ovat edelleen erittäin menestyksekkäitä ja vallitsevat Maassa – enemmistön sekä biomassasta että lajeista muodostavat esitumalliset.[166]
Sovellukset
Evoluutiobiologiassa käytetyillä käsitteillä ja malleilla, kuten luonnonvalinnalla, on useita sovelluksia.[283]
Jalostus viittaa elävien eliöiden, mikrobien, eläinten tai kasvien kehittämiseen haluttuun suuntaan keinotekoisesti. Tätä on käytetty tuhansien vuosien ajan kasvien ja eläinten domestikaatioon.[284] Tämänkaltaisesta valinnasta on hiljattain tullut merkittävä osa geenitekniikkaa, jossa käytetään antibioottiresistenttien geenien kaltaisia geenitekniikan tekijöitä manipuloimaan DNA:ta. Haluttuja ominaisuuksia omaavia proteiineja kehitetään toistuvien mutaatioiden ja valintojen kierrosten kautta prosessissa, jota kutsutaan ohjatuksi evoluutioksi.[285]
Useat sairaudet eivät ole pysyviä ilmiöitä, vaan kykenevät muuttumaan evoluution myötä. Virukset, bakteerit, sienet ja syövät kehittyvät resistenteiksi immuunijärjestelmille ja lääkeaineille.[286][287][288] Samat ongelmat ilmenevät maanviljelyssä kun käytetään hyönteismyrkkyjä[289] ja herbisideja[290].
Tietojenkäsittelytieteessä aloitettiin evoluution simulointi evoluutioalgoritmeja ja tekoelämää käyttäen 1960-luvulla.[291] Evoluutioalgoritmeista tuli laajalti tunnustettu optimointimenetelmä Ingo Rechenbergin 1960-luvulla tekemän työn johdosta. Hän käytti evoluution strategioita ratkaisemaan monimutkaisia teknisiä ongelmia.[292] Erityisesti geneettiset algoritmit nousivat suosioon John Hollandin kirjoitusten kautta.[293] Käytännön sovelluksiin lukeutuu myös tietokoneohjelmien automaattinen evoluutio.[294] Nykyään evoluutioalgoritmeja käytetään optimoimaan järjestelmiä ja ratkaisemaan moniulotteisia ongelmia tehokkaammin kuin ihmisten kehittämät ohjelmistot.[295]
Yhteiskunnalliset ja kulttuuriset vaikutukset
Uuden ajan aatehistoriassa on tapana puhua kahdesta suuresta vallankumouksesta. Ensimmäinen vallankumous oli maakeskisen todellisuuskäsityksen sortuminen. Tähtitieteilijä Kopernikuksen mukaan tätä ensimmäistä vallankumousta, joka tapahtui noin neljäsataa vuotta sitten, kutsutaan kopernikaaniseksi vallankumoukseksi. Toinen mullistus alkoi, kun Darwin osoitti, että ihmiskuntakin on osa luontoa eikä siitä erillinen ilmiö.[296][297]
Ajatus elämän kehittymisestä oli aktiivisen akateemisen keskustelun aiheena 1800-luvun aikana, erityisesti Lajien synnyn (1859) julkaisun jälkeen. Keskustelun keskipisteenä oli evoluutioteorian filosofiset, yhteiskunnalliset ja uskonnolliset vaikutukset. Valtaosa nykypäivän tieteilijöistä hyväksyy synteettisen evoluutioteorian.[57] Tästä huolimatta, evoluutio on edelleen kiistanalainen käsite eräille teisteille.[298]
Useat uskonnot ja kirkkokunnat ovat tulleet sovintoon evoluutioteorian kanssa teistisen evolutionismin kaltaisten käsitteiden kautta. Tästä huolimatta on kreationisteja jotka uskovat, että evoluutioteoria on ristiriidassa heidän uskonnossa olevan luomiskertomuksen kanssa ja vastustavat sitä.[156][299][300] Kuten tuli ilmi Vestiges of the Natural History of Creation -nimisen kirjan julkaisua seuranneessa keskustelussa vuonna 1844, evoluutiobiologian kiistellyin ominaisuus on ihmisen evoluution implisiitti, että ihmisillä on yhteinen esivanhempi apinoiden kanssa, ja että ihmiskunnan järjellisillä ja moraalisilla kyvyillä on samat luonnolliset syyt ja perityt ominaisuudet kuin eläimillä.[301] Joissain maissa, eritoten Yhdysvalloissa, nämä tieteen ja uskonnon väliset jännitteet ovat ruokkineet uskonnollista konfliktia, joka on keskittynyt julkista koulutusta ympäröivään politiikkaan.[302][303] Vaikka kosmologian[304] ja geotieteiden[305] kaltaiset tieteenhaarat ovat myös ristiriidassa usean pyhän kirjoituksen sananmukaisen tulkinnan kanssa, evoluutiobiologia on saanut osakseen huomattavasti enemmän vastustusta henkilöiltä, jotka tulkitsevat pyhiä kirjoituksia kirjaimellisesti.
Vuonna 2006 julkaistun tutkimuksen mukaan, 65 prosenttia suomalaisista piti kehitysoppia oikeana ja 30 prosenttia vääränä.[302] Tämä oli tiedelehti Sciencen mukaan epäilevämpää kuin monissa muissa Euroopan maissa ja Japanissa.[306] Suomalaisten vakuuttuneisuus evoluutiosta on kuitenkin kasvanut vähä vähältä niin, että vuonna 2019 teetetyn kyselyn mukaan 71 prosenttia piti ihmisten kehitystä varhaisemmista eläinlajeista totena ja 10 prosenttia ei.[307]
Lähteet
|
|
Viitteet
- Hall & Hallgrímsson: Strickberger's Evolution, s. 4–6.
- Hall & Hallgrímsson: Strickberger's Evolution, s. 3–5.
- Doolittle, W. Ford: Uprooting the Tree of Life. Scientific American, Helmikuu 2000, 282. vsk, nro 2, s. 90–95. London: Nature Publishing Group. PubMed:10710791. doi:10.1038/scientificamerican0200-90. ISSN 0036-8733. (englanniksi)
- Glansdorff, Nicolas; Ying Xu; Labedan, Bernard: The Last Universal Common Ancestor: emergence, constitution and genetic legacy of an elusive forerunner. Biology Direct, 9.7.2008, 3. vsk. London: BioMed Central. PubMed:18613974. doi:10.1186/1745-6150-3-29. ISSN 1745-6150. (englanniksi)
- Panno: The Cell: Evolution of the First Organism, s. xv-16.
- Futuyma: Assembling the Tree of Life, s. 33.
- Stearns & Stearns: Watching, from the Edge of Extinction, s. x.
- Novacek, Michael J.: Prehistory’s Brilliant Future The New York Times. 8. marraskuuta 2014. New York: The New York Times Company. Viitattu 16. elokuuta 2015. (englanniksi)
- Miller & Spoolman: Environmental Science, s. 62.
- Mora, Camilo; Tittensor, Derek P.; Adl, Sina; Simpson, Alastair G. B.; Worm, Boris: How Many Species Are There on Earth and in the Ocean?. PLOS Biology, 23. elokuuta 2011. San Francisco, CA: PLOS. PubMed:21886479. doi:10.1371/journal.pbio.1001127. ISSN 1544-9173. Artikkelin verkkoversio. Viitattu 16. elokuuta 2015. (englanniksi)
- Lewontin, R. C.: The Units of Selection. Annual Review of Ecology and Systematics, November 1970, 1. vsk, s. 1–18. Palo Alto, CA: Annual Reviews. doi:10.1146/annurev.es.01.110170.000245. ISSN 1545-2069. JSTOR 2096764. (englanniksi)
- Darwin: On the Origin of Species, s. 459.
- Kimura, Motoo: The neutral theory of molecular evolution: a review of recent evidence. The Japanese Journal of Human Genetics, 1991, 66. vsk, nro 4, s. 367–386. Mishima, Japan: Genetics Society of Japan. PubMed:1954033. doi:10.1266/jjg.66.367. ISSN 0021-504X. Artikkelin verkkoversio. (englanniksi)
- Provine: Evolutionary Progress, s. 49–79.
- National Academy of Sciences; Institute of Medicine: Science, Evolution, and Creationism, s. R11–R12.
- Ayala, Francisco J.; Avise, John C.: Essential Readings in Evolutionary Biology. Baltimore, MD: Johns Hopkins University Press, 2014. ISBN 978-1-4214-1305-1. OCLC 854285705. (englanniksi)lähde tarkemmin?
- National Academy of Sciences; Institute of Medicine: Science, Evolution, and Creationism, s. 17.
- Moore, Randy; Decker, Mark; Cotner, Sehoya: Chronology of the Evolution-Creationism Controversy., s. 454. Greenwood Press/ABC Clio, 2010. (englanniksi)
- Futuyama, Douglas J.: Evolution, Science, and Society: Evolutionary Biology and the National Research Agenda (PDF) 1999. Office of University Publications, Rutgers, The State University of New Jersey. Arkistoitu 31.1.2012. Viitattu 8.9.2015. (englanniksi)
- Darwin: The foundations of The origin of species, a sketch written in 1842, s. 53.
- The Presocratic Philosophers: A Critical History with a Selection of Texts, s. 100–142, 280–321. 2. painos. Cambridge; New York: Cambridge University Press, 1983. ISBN 0-521-27455-9. OCLC 9081712. (englanniksi)
- Lucretius: ”Kirja V, rivit 855–877”, De Rerum Natura. Kääntänyt William Ellery Leonard (1916). Medford/Somerville, MA: Tufts University. Teoksen verkkoversio (viitattu 15. elokuuta 2015). OCLC 33233743. (englanniksi)
- Sedley, David: Lucretius and the New Empedocles. Leeds International Classical Studies, 2003, 2. vsk, nro 4. Leeds, West Yorkshire, Englanti: Leeds International Classics Seminar. ISSN 1477-3643. Artikkelin verkkoversio (PDF). Viitattu 15. elokuuta 2015. (englanniksi)
- Torrey, Harry Beal; Felin, Frances: Was Aristotle an Evolutionist?. The Quarterly Review of Biology, maaliskuu 1937, 12. vsk, nro 1, s. 1–18. doi:10.1086/394520. ISSN 0033-5770. JSTOR 2808399. (englanniksi)
- Hull, David L.: The Metaphysics of Evolution. The British Journal for the History of Science, joulukuu 1967, 3. vsk, nro 4, s. 309–337. Cambridge: Cambridge University Press. doi:10.1017/S0007087400002892. JSTOR 4024958. (englanniksi)
- Mason: A History of the Sciences, s. 43–44.
- Mayr: The Growth of Biological Thought, s. 256–257.
- Ray: Historia Plantarum.
- Waggoner, Ben: Carl Linnaeus (1707-1778) 7.7.2000. Berkeley, CA: University of California Museum of Paleontology. Viitattu 15. elokuuta 2015. (englanniksi)
- Bowler: Evolution: The History of an Idea, s. 73–75.
- Erasmus Darwin (1731-1802) Evolution. 4. lokakuuta 1995. Berkeley, CA: University of California Museum of Paleontology. Viitattu 15. elokuuta 2015. (englanniksi)
- Lamarck: Philosophie Zoologique
- Nardon & Grenier: Symbiosis as a Source of Evolutionary Innovation: Speciation and Morphogenesis, s. 162.
- Gould: The Structure of Evolutionary Theory.
- Ghiselin, Michael T.: The Imaginary Lamarck: A Look at Bogus 'History' in Schoolbooks. The Textbook Letter, syyskuu–lokakuu 1994. Sausalito, CA: The Textbook League. Artikkelin verkkoversio. Viitattu 15. elokuuta 2015. (englanniksi) (Arkistoitu – Internet Archive)
- Magner: A History of the Life Sciences.
- Jablonka, Eva; Lamb, Marion J.: Précis of Evolution in Four Dimensions. Behavioural and Brain Sciences, elokuu 2007, 30. vsk, nro 4, s. 353–365. Cambridge: Cambridge University Press. doi:10.1017/S0140525X07002221. ISSN 0140-525X. (englanniksi)
- Burkhardt & Smith: The Correspondence of Charles Darwin.
- Darwin, C. R. to Lubbock, John (Kirje 2532, 22. marraskuuta 1859.) Darwin Correspondence Project. Cambridge, UK: University of Cambridge. Viitattu 15. elokuuta 2015. (englanniksi)
- Sulloway, Frank J.: Why Darwin rejected intelligent design. Journal of Biosciences, kesäkuu 2009, 34. vsk, nro 2, s. 173–183. Bangalore: Indian Academy of Sciences. PubMed:19550032. doi:10.1007/s12038-009-0020-8. ISSN 0250-5991. (englanniksi)
- Dawkins: Sokea kelloseppä.
- Sober, Elliott: Did Darwin write the Origin backwards?. Proc. Natl. Acad. Sci. U.S.A., 16. kesäkuuta 2009, 106. vsk, nro Suppl. 1, s. 10048–10055. Washington, D.C.: National Academy of Sciences. doi:10.1073/pnas.0901109106. ISSN 0027-8424. Bibcode:2009PNAS..10610048S. (englanniksi)
- Mayr: Evoluutio.
- Bowler: Evolution: The History of an Idea, s. 145–146.
- Sokal, Robert R.; Crovello, Theodore J.: The Biological Species Concept: A Critical Evaluation. The American Naturalist, maaliskuu–huhtikuu 1970, 104. vsk, nro 936, s. 127–153. Chicago, IL: University of Chicago Press; American Society of Naturalists. doi:10.1086/282646. ISSN 0003-0147. JSTOR 2459191. (englanniksi)
- Darwin, Charles; Wallace, Alfred: On the Tendency of Species to form Varieties; and on the Perpetuation of Varieties and Species by Natural Means of Selection. Journal of the Proceedings of the Linnean Society of London. Zoology, 20. elokuuta 1858, 3. vsk, nro 9, s. 45–62. doi:10.1111/j.1096-3642.1858.tb02500.x. ISSN 1096-3642. Artikkelin verkkoversio. Viitattu 15. elokuuta 2015. (englanniksi)
- Desmond, Adrian J.: Thomas Henry Huxley Encyclopædia Britannica Online. 17. heinäkuuta 2014. Chicago, IL: Encyclopædia Britannica, Inc.. Viitattu 15. elokuuta 2015. (englanniksi)
- Liu, Y. S.; Zhou, X. M.; Zhi, M. X.; Li, X. J.; Wang, Q. L.: Darwin's contributions to genetics. Journal of Applied Genetics, syyskuu 2009, 50. vsk, nro 3, s. 177–184. Poznań: Institute of Plant Genetics, Polish Academy of Sciences. PubMed:19638672. doi:10.1007/BF03195671. ISSN 1234-1983. (englanniksi)
- Weiling, Franz: Historical study: Johann Gregor Mendel 1822–1884. American Journal of Medical Genetics, heinäkuu 1991, 40. vsk, nro 1, s. 1–25; discussion 26. PubMed:1887835. doi:10.1002/ajmg.1320400103. (englanniksi)
- Wright: Genetic and Biometric Foundations, s. 480.
- Provine: The Origins of Theoretical Population Genetics.
- Stamhuis, Ida H.; Meijer, Onno G.; Zevenhuizen, Erik J. A.: Hugo de Vries on Heredity, 1889-1903: Statistics, Mendelian Laws, Pangenes, Mutations. Isis, kesäkuu 1999, 90. vsk, nro 2, s. 238–267. Chicago, IL: University of Chicago Press. PubMed:10439561. doi:10.1086/384323. ISSN 0021-1753. JSTOR 237050. (englanniksi)
- Quammen: The Reluctant Mr. Darwin.
- Bowler, Peter J.: The Mendelian Revolution: The Emergence of Hereditarian Concepts in Modern Science and Society. Baltimore: Johns Hopkins University Press, 1989. ISBN 0-8018-3888-6. OCLC 19322402. (englanniksi)lähde tarkemmin?
- Watson, J. D.; Crick, F. H. C.: Molecular Structure of Nucleic Acids: A Structure for Deoxyribose Nucleic Acid. Nature, 25. huhtikuuta 1953, 171. vsk, nro 4356, s. 737–738. Lontoo: Nature Publishing Group. PubMed:13054692. doi:10.1038/171737a0. ISSN 0028-0836. Bibcode:1953Natur.171..737W. Artikkelin verkkoversio (PDF). Viitattu 15. elokuuta 2015. (englanniksi)
- Hennig: Phylogenetic Systematics, s. 280.
- Wiley & Lieberman: Phylogenetics.
- Dobzhansky, Theodosius: Nothing in Biology Makes Sense Except in the Light of Evolution. The American Biology Teacher, maaliskuu 1973, 35. vsk, nro 3, s. 125–129. McLean, VA: National Association of Biology Teachers. doi:10.2307/4444260. (englanniksi)
- Kutschera, Ulrich; Niklas, Karl J.: The modern theory of biological evolution: an expanded synthesis. Naturwissenschaften, kesäkuu 2004, 91. vsk, nro 6, s. 255–276. Springer Science+Business Media. PubMed:15241603. doi:10.1007/s00114-004-0515-y. ISSN 1432-1904. Bibcode:2004NW.....91..255K. (englanniksi)
- Avise, John C.; Ayala, Francisco J.: In the light of evolution IV: The human condition. Proc. Natl. Acad. Sci. U.S.A., 11. toukokuuta 2010, 107. vsk, nro Suppl. 2, s. 8897–8901. Washington, D.C.: National Academy of Sciences. doi:10.1073/pnas.100321410. ISSN 0027-8424. Artikkelin verkkoversio (PDF). (englanniksi)
- Sturm, R.A.; Frudakis, T.N.: Eye colour: portals into pigmentation genes and ancestry. Trends Genet., 2004, 20. vsk, nro 8, s. 327–32. PubMed:15262401. doi:10.1016/j.tig.2004.06.010. Artikkelin verkkoversio. Viitattu 15.2.2016. (englanniksi)
- Pearson, H.: Genetics: what is a gene?. Nature, 2006, 441. vsk, nro 7092, s. 398–401. PubMed:16724031. doi:10.1038/441398a. Bibcode:2006Natur.441..398P. Artikkelin verkkoversio. Viitattu 15.2.2016. (englanniksi)
- Visscher, P.M.; Hill, W.G.; Wray, N.R.: Heritability in the genomics era—concepts and misconceptions. Nature Reviews Genetics, 2008, 9. vsk, nro 4, s. 255–66. PubMed:18319743. doi:10.1038/nrg2322. Artikkelin verkkoversio. Viitattu 15.2.2016. (englanniksi)
- Oetting, W.S.; Brilliant, M.H.; King, R.A.: The clinical spectrum of albinism in humans. Molecular medicine today, 1996, 2. vsk, nro 8, s. 330–5. PubMed:8796918. doi:10.1016/1357-4310(96)81798-9. (englanniksi)
- Futuyma, Douglas J.: Evolution. Sunderland, Massachusetts: Sinauer Associates, Inc, 2005. ISBN 0-87893-187-2. (englanniksi)
- Phillips, P.C.: Epistasis—the essential role of gene interactions in the structure and evolution of genetic systems. Nature Reviews Genetics, 2008, 9. vsk, nro 11, s. 855–67. PubMed:18852697. doi:10.1038/nrg2452. (englanniksi)
- Wu, R.; Lin, M.: Functional mapping – how to map and study the genetic architecture of dynamic complex traits. Nature Reviews Genetics, 2006, 7. vsk, nro 3, s. 229–37. PubMed:16485021. doi:10.1038/nrg1804. (englanniksi)
- Jablonka, E.; Raz, G.: Transgenerational epigenetic inheritance: Prevalence, mechanisms and implications for the study of heredity and evolution. The Quarterly Review of Biology, 2009, 84. vsk, nro 2, s. 131–176. PubMed:19606595. doi:10.1086/598822. (englanniksi)
- Bossdorf, O.; Arcuri, D.; Richards, C. L.; Pigliucci, M.: Experimental alteration of DNA methylation affects the phenotypic plasticity of ecologically relevant traits in Arabidopsis thaliana. Evolutionary Ecology, 2010, 24. vsk, nro 3, s. 541–553. doi:10.1007/s10682-010-9372-7. (englanniksi)
- Jablonka, E.; Lamb, M.: Evolution in four dimensions: Genetic, epigenetic, behavioural and symbolic. MIT Press, 2005. ISBN 0-262-10107-6. Teoksen verkkoversio. (englanniksi)
- Jablonka, E.; Lamb, M.J.: The changing concept of epigenetics. Annals of the New York Academy of Sciences, 2002, 981. vsk, nro 1, s. 82–96. PubMed:12547675. doi:10.1111/j.1749-6632.2002.tb04913.x. Bibcode:2002NYASA.981...82J. (englanniksi)
- Laland, Kevin N.; Sterelny, Kim: Perspective: Seven reasons (not) to neglect niche construction. Evolution, syyskuu 2006, 60. vsk, nro 8, s. 1751–1762. doi:10.1111/j.0014-3820.2006.tb00520.x. (englanniksi)
- Chapman, Michael J.; Margulis, Lynn: Morphogenesis by symbiogenesis. International Microbiology, joulukuu 1998, 1. vsk, nro 4, s. 319–326. PubMed:10943381. (englanniksi)
- Wilson, David Sloan; Wilson, Edward O.: Rethinking the theoretical foundation of sociobiology. The Quarterly Review of Biology, joulukuu 2007, 82. vsk, nro 4, s. 327–348. PubMed:18217526. doi:10.1086/522809. Artikkelin verkkoversio. (englanniksi) (Arkistoitu – Internet Archive)
- Harwood AJ: Factors affecting levels of genetic diversity in natural populations. Philosophical Transactions of the Royal Society B, 1998, 353. vsk, nro 1366, s. 177–86. PubMed:9533122. doi:10.1098/rstb.1998.0200. (englanniksi)
- Ewens W.J.: Mathematical Population Genetics. 2. painos. Springer-Verlag, New York, 2004. ISBN 0-387-20191-2. (englanniksi)
- Butlin RK, Tregenza T: Levels of genetic polymorphism: marker loci versus quantitative traits. Philosophical Transactions of the Royal Society B, 1998, 353. vsk, nro 1366, s. 187–98. PubMed:9533123. doi:10.1098/rstb.1998.0201. (englanniksi)
- Wetterbom A, Sevov M, Cavelier L, Bergström TF: Comparative genomic analysis of human and chimpanzee indicates a key role for indels in primate evolution. J. Mol. Evol., 2006, 63. vsk, nro 5, s. 682–90. PubMed:17075697. doi:10.1007/s00239-006-0045-7. (englanniksi)
- Sawyer SA, Parsch J, Zhang Z, Hartl DL: Prevalence of positive selection among nearly neutral amino acid replacements in Drosophila. Proc. Natl. Acad. Sci. U.S.A., 2007, 104. vsk, nro 16, s. 6504–10. PubMed:17409186. doi:10.1073/pnas.0701572104. Bibcode:2007PNAS..104.6504S. (englanniksi)
- Hastings, PJ; Lupski, JR; Rosenberg, SM; Ira, G: Mechanisms of change in gene copy number. Nature Reviews Genetics, 2009, 10. vsk, nro 8, s. 551–564. PubMed:19597530. doi:10.1038/nrg2593. (englanniksi)
- Sean B. Carroll; Jennifer K. Grenier; Scott D. Weatherbee: From DNA to Diversity: Molecular Genetics and the Evolution of Animal Design. 2. painos. Oxford: Blackwell Publishing, 2005. ISBN 1-4051-1950-0. (englanniksi)
- Harrison P, Gerstein M: Studying genomes through the aeons: protein families, pseudogenes and proteome evolution. J Mol Biol, 2002, 318. vsk, nro 5, s. 1155–74. PubMed:12083509. doi:10.1016/S0022-2836(02)00109-2. (englanniksi)
- Bowmaker JK: Evolution of colour vision in vertebrates. Eye (London, England), 1998, 12. vsk, nro Pt 3b, s. 541–7. PubMed:9775215. doi:10.1038/eye.1998.143. (englanniksi)
- Gregory TR; Hebert PD: The modulation of DNA content: proximate causes and ultimate consequences. Genome Res., 1999, 9. vsk, nro 4, s. 317–24. PubMed:10207154. doi:10.1101/gr.9.4.317. Artikkelin verkkoversio. (englanniksi)
- Hurles M: Gene duplication: the genomic trade in spare parts. PLoS Biol., 2004, 2. vsk, nro 7, s. E206. PubMed:15252449. doi:10.1371/journal.pbio.0020206. (englanniksi)
- Liu, N; Okamura, K; Tyler, DM: The evolution and functional diversification of animal microRNA genes. Cell Res., 2008, 18. vsk, nro 10, s. 985–96. PubMed:18711447. doi:10.1038/cr.2008.278. Artikkelin verkkoversio. (englanniksi)
- Siepel, A: Darwinian alchemy: Human genes from noncoding DNA. Genome Res., 2009, 19. vsk, nro 10, s. 1693–5. PubMed:19797681. doi:10.1101/gr.098376.109. Artikkelin verkkoversio. (englanniksi)
- Orengo, CA; Thornton, JM: Protein families and their evolution-a structural perspective. Annu. Rev. Biochem., 2005, 74. vsk, nro 1, s. 867–900. PubMed:15954844. doi:10.1146/annurev.biochem.74.082803.133029. (englanniksi)
- Long, M; Betrán, E; Thornton, K; Wang, W: The origin of new genes: glimpses from the young and old. Nature Reviews Genetics, 2003, 4. vsk, nro 11, s. 865–75. PubMed:14634634. doi:10.1038/nrg1204. (englanniksi)
- Wang, M; Caetano-Anollés, G: The evolutionary mechanics of domain organisation in proteomes and the rise of modularity in the protein world. Structure, 2009, 17. vsk, nro 1, s. 66–78. PubMed:19141283. doi:10.1016/j.str.2008.11.008. (englanniksi)
- Weissman, KJ; Müller, R: Protein-protein interactions in multienzyme megasynthetases. Chembiochem, 2008, 9. vsk, nro 6, s. 826–48. PubMed:18357594. doi:10.1002/cbic.200700751. (englanniksi)
- Radding, Charles M.: Homologous Pairing and Strand Exchange in Genetic Recombination. Annual Review of Genetics, joulukuu 1982, 16. vsk, s. 405–437. Palo Alto, CA: Annual Reviews. PubMed:6297377. doi:10.1146/annurev.ge.16.120182.002201. ISSN 0066-4197. (englanniksi)
- Agrawal, Aneil F.: Evolution of Sex: Why Do Organisms Shuffle Their Genotypes?. Current Biology, 5. syyskuuta 2006, 16. vsk, nro 17, s. R696–R704. Cambridge, MA: Cell Press. PubMed:16950096. doi:10.1016/j.cub.2006.07.063. ISSN 0960-9822. (englanniksi)
- Peters, Andrew D.; Otto, Sarah P.: Liberating genetic variance through sex. BioEssays, kesäkuu 2003, 25. vsk, nro 6, s. 533–537. Hoboken, NJ: John Wiley & Sons. PubMed:12766942. doi:10.1002/bies.10291. ISSN 0265-9247. (englanniksi)
- Goddard, Matthew R.; Godfray, H. Charles J.; Burt, Austin: Sex increases the efficacy of natural selection in experimental yeast populations. Nature, 31. maaliskuuta 2005, 434. vsk, nro 7033, s. 636–640. Lontoo: Nature Publishing Group. PubMed:15800622. doi:10.1038/nature03405. ISSN 0028-0836. Bibcode:2005Natur.434..636G. (englanniksi)
- Maynard Smith, John: The Evolution of Sex. Cambridge; New York: Cambridge University Press, 1978. ISBN 0-521-29302-2. OCLC 3413793. (englanniksi)
- Ridley, Matt: The Red Queen: Sex and the Evolution of Human Nature. New York: Viking, 1993. ISBN 0-670-84357-1. OCLC 636657988. (englanniksi)
- Van Valen, Leigh: A New Evolutionary Law. Evolutionary Theory, 1973, 1. vsk, s. 1–30. Chicago, IL: University of Chicago. ISSN 0093-4755. Artikkelin verkkoversio (PDF). (englanniksi) (Arkistoitu – Internet Archive)
- Hamilton, W. D.; Axelrod, Robert; Tanese, Reiko: Sexual reproduction as an adaptation to resist parasites (a review). Proc. Natl. Acad. Sci. U.S.A., 1. toukokuuta 1990, 87. vsk, nro 9, s. 3566–3573. Washington, D.C.: National Academy of Sciences. PubMed:2185476. doi:10.1073/pnas.87.9.3566. ISSN 0027-8424. (englanniksi)
- Birdsell, John A.; Wills, Christopher: ”The Evolutionary Origin and Maintenance of Sexual Recombination: A Review of Contemporary Models”, Evolutionary Biology. 33. osa Evolutionary Biology -sarjaa. New York: Springer Science+Business Media, 2003. ISBN 978-1-4419-3385-0. OCLC 751583918. (englanniksi)
- Morjan, C.; Rieseberg, L.: How species evolve collectively: implications of gene flow and selection for the spread of advantageous alleles. Mol. Ecol., 2004, 13. vsk, nro 6, s. 1341–56. PubMed:15140081. doi:10.1111/j.1365-294X.2004.02164.x. (englanniksi)
- Boucher, Yan; Douady, Christophe J.; Papke, R. Thane et al.: Lateral gene transfer and the origins of prokaryotic groups. Annu Rev Genet, joulukuu 2003, 37. vsk, nro 1, s. 283–328. PubMed:14616063. doi:10.1146/annurev.genet.37.050503.084247. (englanniksi)
- Walsh, Timothy R.: Combinatorial genetic evolution of multiresistance. Current Opinion in Microbiology, lokakuu 2006, 9. vsk, nro 5, s. 476–82. PubMed:16942901. doi:10.1016/j.mib.2006.08.009. (englanniksi)
- Kondo, Natsuko; Nikoh, Naruo; Ijichi, Nobuyuki et al.: Genome fragment of Wolbachia endosymbiont transferred to X chromosome of host insect. Proc. Natl. Acad. Sci. U.S.A., 29. lokakuuta 2002, 99. vsk, nro 22, s. 14280–5. PubMed:12386340. doi:10.1073/pnas.222228199. Bibcode:2002PNAS...9914280K. (englanniksi)
- Sprague, George F., Jr.: Genetic exchange between kingdoms. Current Opinion in Genetics & Development, joulukuu 1991, 1. vsk, nro 4, s. 530–3. PubMed:1822285. doi:10.1016/S0959-437X(05)80203-5. (englanniksi)
- Gladyshev, Eugene A.; Meselson, Matthew; Arkhipova, Irina R.: Massive horizontal gene transfer in bdelloid rotifers. Science, 30. toukokuuta 2008, 320. vsk, nro 5880, s. 1210–3. PubMed:18511688. doi:10.1126/science.1156407. Bibcode:2008Sci...320.1210G. (englanniksi)
- Baldo, Angela M.; McClure, Marcella A.: Evolution and horizontal transfer of dUTPase-encoding genes in viruses and their hosts. J. Virol., syyskuu 1999, 73. vsk, nro 9, s. 7710–21. PubMed:10438861. (englanniksi)
- River, M. C.; Lake, J. A.: The ring of life provides evidence for a genome fusion origin of eukaryotes. Nature, 2004, 431. vsk, nro 9, s. 152–5. PubMed:15356622. doi:10.1038/nature02848. Bibcode:2004Natur.431..152R. (englanniksi)
- Hurst, Laurence D.: Fundamental concepts in genetics: genetics and the understanding of selection. Nature Reviews Genetics, helmikuu 2009, 10. vsk, nro 2, s. 83–93. PubMed:19119264. doi:10.1038/nrg2506. (englanniksi)
- Orr, H. Allen: Fitness and its role in evolutionary genetics. Nature Reviews Genetics, elokuu 2009, 10. vsk, nro 8, s. 531–9. PubMed:19546856. doi:10.1038/nrg2603. (englanniksi)
- Haldane, J. B. S.: The theory of natural selection today. Nature, 14. maaliskuuta 1959, 183. vsk, nro 4663, s. 710–3. PubMed:13644170. doi:10.1038/183710a0. Bibcode:1959Natur.183..710H. (englanniksi)
- Lande, Russell; Arnold, Stevan J.: The measurement of selection on correlated characters. Evolution, marraskuu 1983, 37. vsk, nro 6, s. 1210–26. doi:10.2307/2408842. JSTOR 2408842. (englanniksi)
- Goldberg, Emma E.; Igić, Boris: On phylogenetic tests of irreversible evolution. Evolution, marraskuu 2008, 62. vsk, nro 11, s. 2727–2741. PubMed:18764918. doi:10.1111/j.1558-5646.2008.00505.x. (englanniksi)
- Collin, Rachel; Miglietta, Maria Pia: Reversing opinions on Dollo's Law. Trends in Ecology & Evolution, marraskuu 2008, 23. vsk, nro 11, s. 602–609. PubMed:18814933. doi:10.1016/j.tree.2008.06.013. (englanniksi)
- Hoekstra, Hopi E.; Hoekstra, Jonathan M.; Berrigan, David et al.: Strength and tempo of directional selection in the wild. Proc. Natl. Acad. Sci. U.S.A., 31. heinäkuuta 2001, 98. vsk, nro 16, s. 9157–60. PubMed:11470913. doi:10.1073/pnas.161281098. Bibcode:2001PNAS...98.9157H. (englanniksi)
- Felsenstein, Joseph: Excursions along the Interface between Disruptive and Stabilizing Selection. Genetics, marraskuu 1979, 93. vsk, nro 3, s. 773–95. PubMed:17248980. (englanniksi)
- Andersson, Malte; Simmons, Leigh W.: Sexual selection and mate choice. Trends Ecol. Evol. (Amst.), kesäkuu 2006, 21. vsk, nro 6, s. 296–302. PubMed:16769428. doi:10.1016/j.tree.2006.03.015. (englanniksi)
- Kokko, Hanna; Brooks, Robert; McNamara, John M.; Houston, Alasdair I.: The sexual selection continuum. Proc. Biol. Sci., 7. heinäkuuta 2002, 269. vsk, nro 1498, s. 1331–40. PubMed:12079655. doi:10.1098/rspb.2002.2020. (englanniksi)
- Quinn, Thomas P.; Hendry, Andrew P.; Buck, Gregory B.: Balancing natural and sexual selection in sockeye salmon: interactions between body size, reproductive opportunity and vulnerability to predation by bears. Evolutionary Ecology Research, 2001, 3. vsk, s. 917–937. (englanniksi)
- Hunt, John; Brooks, Robert; Jennions, Michael D. et al.: High-quality male field crickets invest heavily in sexual display but die young. Nature, 23. joulukuuta 2004, 432. vsk, nro 7020, s. 1024–7. PubMed:15616562. doi:10.1038/nature03084. Bibcode:2004Natur.432.1024H. (englanniksi)
- Okasha, S.: Evolution and the Levels of Selection. Oxford University Press, 2007. ISBN 0-19-926797-9. (englanniksi)
- Gould SJ: Gulliver's further travels: the necessity and difficulty of a hierarchical theory of selection. Philosophical Transactions of the Royal Society B, 1998, 353. vsk, nro 1366, s. 307–14. PubMed:9533127. doi:10.1098/rstb.1998.0211. (englanniksi)
- Mayr E: The objects of selection. Proc. Natl. Acad. Sci. U.S.A., 1997, 94. vsk, nro 6, s. 2091–4. PubMed:9122151. doi:10.1073/pnas.94.6.2091. Bibcode:1997PNAS...94.2091M. (englanniksi)
- Maynard Smith J: The units of selection. Novartis Found. Symp., 1998, 213. vsk, s. 203–11; discussion 211–7. PubMed:9653725. (englanniksi)
- Hickey DA: Evolutionary dynamics of transposable elements in prokaryotes and eukaryotes. Genetica, 1992, 86. vsk, nro 1–3, s. 269–74. PubMed:1334911. doi:10.1007/BF00133725. (englanniksi)
- Gould SJ, Lloyd EA: Individuality and adaptation across levels of selection: how shall we name and generalise the unit of Darwinism?. Proc. Natl. Acad. Sci. U.S.A., 1999, 96. vsk, nro 21, s. 11904–9. PubMed:10518549. doi:10.1073/pnas.96.21.11904. Bibcode:1999PNAS...9611904G. (englanniksi)
- Lynch, M.: The frailty of adaptive hypotheses for the origins of organismal complexity. PNAS, 2007, 104. vsk, nro suppl. 1, s. 8597–8604. PubMed:17494740. doi:10.1073/pnas.0702207104. Bibcode:2007PNAS..104.8597L. (englanniksi)
- Smith N.G.C., Webster M.T., Ellegren, H.: Deterministic Mutation Rate Variation in the Human Genome. Genome Research, 2002, 12. vsk, nro 9, s. 1350–1356. PubMed:12213772. doi:10.1101/gr.220502. Artikkelin verkkoversio. (englanniksi)
- Petrov DA, Sangster TA, Johnston JS, Hartl DL, Shaw KL: Evidence for DNA loss as a determinant of genome size. Science, 2000, 287. vsk, nro 5455, s. 1060–1062. PubMed:10669421. doi:10.1126/science.287.5455.1060. Bibcode:2000Sci...287.1060P. (englanniksi)
- Petrov DA: DNA loss and evolution of genome size in Drosophila. Genetica, 2002, 115. vsk, nro 1, s. 81–91. PubMed:12188050. doi:10.1023/A:1016076215168. (englanniksi)
- Kiontke K, Barriere A , Kolotuev I, Podbilewicz B , Sommer R, Fitch DHA , Felix MA: Trends, stasis, and drift in the evolution of nematode vulva development. Current Biology, 2007, 17. vsk, nro 22, s. 1925–1937. PubMed:18024125. doi:10.1016/j.cub.2007.10.061. (englanniksi)
- Braendle C, Baer CF, Felix MA: Bias and Evolution of the Mutationally Accessible Phenotypic Space in a Developmental System. PLoS Genetics, 2010, 6. vsk, nro 3, s. e1000877. PubMed:20300655. doi:10.1371/journal.pgen.1000877. (englanniksi)
- Palmer, RA: Symmetry breaking and the evolution of development. Science, 2004, 306. vsk, nro 5697, s. 828–833. PubMed:15514148. doi:10.1126/science.1103707. Bibcode:2004Sci...306..828P. (englanniksi)
- West-Eberhard, M-J.: Developmental plasticity and evolution. New York: Oxford University Press, 2003. ISBN 978-0-19-512235-0. (englanniksi)
- Stoltzfus, A; Yampolsky, L.Y.: Climbing Mount Probable: Mutation as a Cause of Nonrandomness in Evolution. J Hered, 2009, 100. vsk, nro 5, s. 637–647. PubMed:19625453. doi:10.1093/jhered/esp048. (englanniksi)
- Yampolsky, L.Y.; Stoltzfus, A: Bias in the introduction of variation as an orienting factor in evolution. Evol Dev, 2001, 3. vsk, nro 2, s. 73–83. PubMed:11341676. doi:10.1046/j.1525-142x.2001.003002073.x. (englanniksi)
- Haldane, JBS: The Part Played by Recurrent Mutation in Evolution. American Naturalist, 1933, 67. vsk, nro 708, s. 5–19. doi:10.1086/280465. JSTOR 2457127. (englanniksi)
- Protas, Meredith; Conrad, Melissa; Gross, Joshua B. et al.: Regressive evolution in the Mexican cave tetra, Astyanax mexicanus. Current Biology, 6. maaliskuuta 2007, 17. vsk, nro 5, s. 452–454. PubMed:17306543. doi:10.1016/j.cub.2007.01.051. (englanniksi)
- Maughan, Heather; Masel, Joanna; Birky, C. William, Jr.; Nicholson, Wayne L.: The roles of mutation accumulation and selection in loss of sporulation in experimental populations of Bacillus subtilis. Genetics, lokakuu 2007, 177. vsk, nro 2, s. 937–948. PubMed:17720926. doi:10.1534/genetics.107.075663. (englanniksi)
- Masel, Joanna; King, Oliver D.; Maughan, Heather: The loss of adaptive plasticity during long periods of environmental stasis. American Naturalist, tammikuu 2007, 169. vsk, nro 1, s. 38–46. PubMed:17206583. doi:10.1086/510212. (englanniksi)
- Masel J: Genetic drift. Current Biology, 2011, 21. vsk, nro 20, s. R837–R838. PubMed:22032182. doi:10.1016/j.cub.2011.08.007. (englanniksi)
- Lande R: Fisherian and Wrightian theories of speciation. Genome, 1989, 31. vsk, nro 1, s. 221–7. PubMed:2687093. doi:10.1139/g89-037. (englanniksi)
- Mitchell-Olds, Thomas; Willis, John H.; Goldstein, David B.: Which evolutionary processes influence natural genetic variation for phenotypic traits?. Nature Reviews Genetics, 2007, 8. vsk, nro 11, s. 845–856. PubMed:17943192. doi:10.1038/nrg2207. (englanniksi)
- Nei M: Selectionism and neutralism in molecular evolution. Mol. Biol. Evol., 2005, 22. vsk, nro 12, s. 2318–42. PubMed:16120807. doi:10.1093/molbev/msi242. (englanniksi)
- Kimura M: The neutral theory of molecular evolution and the world view of the neutralists. Genome, 1989, 31. vsk, nro 1, s. 24–31. PubMed:2687096. doi:10.1139/g89-009. (englanniksi)
- Kreitman M: The neutral theory is dead. Long live the neutral theory. BioEssays, 1996, 18. vsk, nro 8, s. 678–83; discussion 683. PubMed:8760341. doi:10.1002/bies.950180812. (englanniksi)
- Leigh E.G. (Jr): Neutral theory: a historical perspective. Journal of Evolutionary Biology, 2007, 20. vsk, nro 6, s. 2075–91. PubMed:17956380. doi:10.1111/j.1420-9101.2007.01410.x. (englanniksi)
- Gillespie, John H.: Is the population size of a species relevant to its evolution?. Evolution, 2001, 55. vsk, nro 11, s. 2161–2169. PubMed:11794777. doi:10.1111/j.0014-3820.2001.tb00732.x. (englanniksi)
- Neher, Richard A.; Shraiman, Boris I.: Genetic Draft and Quasi-Neutrality in Large Facultatively Sexual Populations. Genetics, 2011, 188. vsk, nro 4, s. 975–996. PubMed:21625002. doi:10.1534/genetics.111.128876. (englanniksi)
- Otto, Sarah P.; Whitlock, Michael C.: The probability of fixation in populations of changing size. Genetics, 1. kesäkuuta 1997, 146. vsk, nro 2, s. 723–33. PubMed:9178020. (englanniksi)
- Charlesworth, Brian: Fundamental concepts in genetics: Effective population size and patterns of molecular evolution and variation. Nature Reviews Genetics, maaliskuu 2009, 10. vsk, nro 3, s. 195–205. PubMed:19204717. doi:10.1038/nrg2526. (englanniksi)
- Cutter, Asher D.; Choi, Jae Young: Natural selection shapes nucleotide polymorphism across the genome of the nematode Caenorhabditis briggsae. Genome Research, elokuu 2010, 20. vsk, nro 8, s. 1103–1111. PubMed:20508143. doi:10.1101/gr.104331.109. (englanniksi)
- Lien, Sigbjørn; Szyda, Joanna; Schechinger, Birgit et al.: Evidence for heterogeneity in recombination in the human pseudoautosomal region: high resolution analysis by sperm typing and radiation-hybrid mapping. Am. J. Hum. Genet., helmikuu 2000, 66. vsk, nro 2, s. 557–66. PubMed:10677316. doi:10.1086/302754. (englanniksi)
- Barton, Nicholas H.: Genetic hitchhiking. Philosophical Transactions of the Royal Society B, 29. marraskuuta 2000, 355. vsk, nro 1403, s. 1553–1562. PubMed:11127900. doi:10.1098/rstb.2000.0716. (englanniksi)
- Orr, H. Allen: Dobzhansky, Bateson, and the Genetics of Speciation. Genetics, joulukuu 1996, 144. vsk, s. 1331–1335. Artikkelin verkkoversio (PDF). (englanniksi)
- Wright, Sewall: The roles of mutation, inbreeding, crossbreeding and selection in evolution. Proc. 6th Int. Cong. Genet, 1932, 1. vsk, s. 356–366. Artikkelin verkkoversio. (englanniksi)
- Coyne, Jerry A.; Barton, Nicholas H.; Turelli, Michael: Perspective: A Critique of Sewall Wright's Shifting Balance Theory of Evolution. Evolution, kesäkuu 1997, 51. vsk, nro 3, s. 643–671. doi:10.2307/2411143. (englanniksi)
- Scott EC, Matzke NJ: Biological design in science classrooms. Proc. Natl. Acad. Sci. U.S.A., 2007, 104. vsk, nro suppl_1, s. 8669–76. PubMed:17494747. doi:10.1073/pnas.0701505104. Bibcode:2007PNAS..104.8669S. (englanniksi)
- Hendry AP, Kinnison MT: An introduction to microevolution: rate, pattern, process. Genetica, 2001, 112–113. vsk, s. 1–8. PubMed:11838760. doi:10.1023/A:1013368628607. (englanniksi)
- Leroi AM: The scale independence of evolution. Evol. Dev., 2000, 2. vsk, nro 2, s. 67–77. PubMed:11258392. doi:10.1046/j.1525-142x.2000.00044.x. (englanniksi)
- Gould, Stephen Jay: The Structure of Evolutionary Theory, s. 657–658. Cambridge, MA: Belknap Press of Harvard University Press, 2002. ISBN 0-674-00613-5. OCLC 47869352. (englanniksi)
- Gould, Stephen Jay: Tempo and mode in the macroevolutionary reconstruction of Darwinism. Proc. Natl. Acad. Sci. U.S.A., 19. heinäkuuta 1994, 91. vsk, nro 15, s. 6764–71. Washington, D.C.: National Academy of Sciences. PubMed:8041695. doi:10.1073/pnas.91.15.6764. Bibcode:1994PNAS...91.6764G. (englanniksi)
- Jablonski, David: Micro- and macroevolution: scale and hierarchy in evolutionary biology and paleobiology. Paleobiology, 2000, 26. vsk, nro sp4, s. 15–52. Boulder, CO: doi:10.1666/0094-8373(2000)26[15:MAMSAH]2.0.CO;2. (englanniksi)
- Dougherty, Michael J.: Is the human race evolving or devolving?. Scientific American, 20. heinäkuuta 1998. Artikkelin verkkoversio. (englanniksi)
- TalkOrigins Archive vastaus kreationistien väittämään – Claim CB932: Evolution of degenerate forms (englanniksi)
- Carroll SB: Chance and necessity: the evolution of morphological complexity and diversity. Nature, 2001, 409. vsk, nro 6823, s. 1102–9. PubMed:11234024. doi:10.1038/35059227. (englanniksi)
- Whitman W, Coleman D, Wiebe W: Prokaryotes: the unseen majority. Proc Natl Acad Sci U S A, 1998, 95. vsk, nro 12, s. 6578–83. PubMed:9618454. doi:10.1073/pnas.95.12.6578. Bibcode:1998PNAS...95.6578W. (englanniksi)
- Schloss P, Handelsman J: Status of the microbial census. Microbiol Mol Biol Rev, 2004, 68. vsk, nro 4, s. 686–91. PubMed:15590780. doi:10.1128/MMBR.68.4.686-691.2004. (englanniksi)
- Nealson K: Post-Viking microbiology: new approaches, new data, new insights. Orig Life Evol Biosph, 1999, 29. vsk, nro 1, s. 73–93. PubMed:11536899. doi:10.1023/A:1006515817767. (englanniksi)
- Buckling A, Craig Maclean R, Brockhurst MA, Colegrave N: The Beagle in a bottle. Nature, 2009, 457. vsk, nro 7231, s. 824–9. PubMed:19212400. doi:10.1038/nature07892. Bibcode:2009Natur.457..824B. (englanniksi)
- Elena SF, Lenski RE: Evolution experiments with microorganisms: the dynamics and genetic bases of adaptation. Nature Reviews Genetics, 2003, 4. vsk, nro 6, s. 457–69. PubMed:12776215. doi:10.1038/nrg1088. (englanniksi)
- Mayr, Ernst: The Growth of Biological Thought: Diversity, Evolution, and Inheritance, s. 483. Cambridge, MA: Belknap Press, 1982. ISBN 0-674-36445-7. OCLC 7875904. (englanniksi): "Adaptation... could no longer be considered a static condition, a product of a creative past and became instead a continuing dynamic process."
- Oxford Dictionary of Science:n määritelmä sanalle adaptation: "Any change in the structure or functioning of an organism that makes it better suited to its environment".
- Orr, H. Allen: The genetic theory of adaptation: a brief history. Nature Reviews Genetics, helmikuu 2005, 6. vsk, nro 2, s. 119–27. PubMed:15716908. doi:10.1038/nrg1523. (englanniksi)
- Dobzhansky, Theodosius; Hecht, Max K.; Steere, William C.: ”On some fundamental concepts of evolutionary biology”, Evolutionary biology volume 2, s. 1–34. 1. painos. New York: Appleton-Century-Crofts, 1968. (englanniksi)
- Dobzhansky, Theodosius: Genetics of the evolutionary process, s. 4–6, 79–82, 84–87. N.Y.: Columbia University Press, 1970. ISBN 0-231-02837-7. (englanniksi)
- Dobzhansky, Theodosius: Genetics of natural populations XXV. Genetic changes in populations of Drosophila pseudoobscura and Drosphila persimilis in some locations in California. Evolution, maaliskuu 1956, 10. vsk, nro 1, s. 82–92. doi:10.2307/2406099. JSTOR 2406099. (englanniksi)
- Nakajima, Akira; Sugimoto, Yohko; Yoneyama, Hiroshi; Nakae, Taiji: High-level fluoroquinolone resistance in Pseudomonas aeruginosa due to interplay of the MexAB-OprM efflux pump and the DNA gyrase mutation. Microbiology and Immunology, kesäkuu 2002, 46. vsk, nro 6, s. 391–395. Tokio: PubMed:12153116. doi:10.1111/j.1348-0421.2002.tb02711.x. (englanniksi)
- Blount, Zachary D.; Borland, Christina Z.; Lenski, Richard E.: Inaugural Article: Historical contingency and the evolution of a key innovation in an experimental population of Escherichia coli. Proc. Natl. Acad. Sci. U.S.A., 10. kesäkuuta 2008, 105. vsk, nro 23, s. 7899–906. PubMed:18524956. doi:10.1073/pnas.0803151105. Bibcode:2008PNAS..105.7899B. (englanniksi)
- Okada, Hirosuke; Negoro, Seiji; Kimura, Hiroyuki; Nakamura, Shunichi: Evolutionary adaptation of plasmid-encoded enzymes for degrading nylon oligomers. Nature, 10. marraskuuta 1983, 306. vsk, nro 5939, s. 203–6. PubMed:6646204. doi:10.1038/306203a0. Bibcode:1983Natur.306..203O. (englanniksi)
- Ohno, Susumu: Birth of a unique enzyme from an alternative reading frame of the preexisted, internally repetitious coding sequence. Proc. Natl. Acad. Sci. U.S.A., huhtikuu 1984, 81. vsk, nro 8, s. 2421–5. PubMed:6585807. doi:10.1073/pnas.81.8.2421. Bibcode:1984PNAS...81.2421O. (englanniksi)
- Copley, Shelley D.: Evolution of a metabolic pathway for degradation of a toxic xenobiotic: the patchwork approach. Trends in Biochemical Sciences, kesäkuu 2000, 25. vsk, nro 6, s. 261–5. PubMed:10838562. doi:10.1016/S0968-0004(00)01562-0. (englanniksi)
- Crawford, Ronald L.; Jung, Carina M.; Strap, Janice L.: The recent evolution of pentachlorophenol (PCP)-4-monooxygenase (PcpB) and associated pathways for bacterial degradation of PCP. Biodegradation, lokakuu 2007, 18. vsk, nro 5, s. 525–39. PubMed:17123025. doi:10.1007/s10532-006-9090-6. (englanniksi)
- Eshel, Ilan: Clone-selection and optimal rates of mutation. Journal of Applied Probability, joulukuu 1973, 10. vsk, nro 4, s. 728–738. doi:10.2307/3212376. JSTOR 3212376.
- Altenberg, Lee: ”Genome growth and the evolution of the genotype-phenotype map”, Evolution and Biocomputation: Computational Models of Evolution, s. 205–259. Lecture Notes in Computer Science, vol. 899. Berlin; New York: Springer-Verlag Berlin Heidelberg, 1995. ISBN 3-540-59046-3. OCLC 32049812. doi:10.1007/3-540-59046-3_11. (englanniksi)
- Masel, Joanna; Bergman, Aviv: The evolution of the evolvability properties of the yeast prion [PSI+]. Evolution, heinäkuu 2003, 57. vsk, nro 7, s. 1498–1512. PubMed:12940355. doi:10.1111/j.0014-3820.2003.tb00358.x. (englanniksi)
- Lancaster, Alex K.; Bardill, J. Patrick; True, Heather L.; Masel, Joanna: The Spontaneous Appearance Rate of the Yeast Prion [PSI+] and Its Implications for the Evolution of the Evolvability Properties of the [PSI+] System. Genetics, helmikuu 2010, 184. vsk, nro 2, s. 393–400. PubMed:19917766. doi:10.1534/genetics.109.110213. (englanniksi)
- Draghi, Jeremy; Wagner, Günter P.: Evolution of evolvability in a developmental model. Theoretical Population Biology, helmikuu 2008, 62. vsk, s. 301–315. doi:10.1111/j.1558-5646.2007.00303.x. (englanniksi)
- Bejder, Lars; Hall, Brian K.: Limbs in whales and limblessness in other vertebrates: mechanisms of evolutionary and developmental transformation and loss. Evolution & Development, marraskuu 2002, 4. vsk, nro 6, s. 445–58. PubMed:12492145. doi:10.1046/j.1525-142X.2002.02033.x. (englanniksi)
- Young, Nathan M.; Hallgrímsson, Benedikt: Serial homology and the evolution of mammalian limb covariation structure. Evolution, 2005, 59. vsk, nro 12, s. 2691–704. PubMed:16526515. doi:10.1554/05-233.1. (englanniksi)
- Penny, David; Poole, Anthony: The nature of the last universal common ancestor. Current Opinion in Genetics & Development, joulukuu 1999, 9. vsk, nro 6, s. 672–77. PubMed:10607605. doi:10.1016/S0959-437X(99)00020-9. (englanniksi)
- Hall, Brian K.: Descent with modification: the unity underlying homology and homoplasy as seen through an analysis of development and evolution. Biological Reviews of the Cambridge Philosophical Society, 2003, 78. vsk, nro 3, s. 409–433. PubMed:14558591. doi:10.1017/S1464793102006097. (englanniksi)
- Shubin, Neil; Tabin, C.; Carroll, S.: Deep homology and the origins of evolutionary novelty. Nature, 2009, 457. vsk, nro 7231, s. 818–823. PubMed:19212399. doi:10.1038/nature07891. Bibcode:2009Natur.457..818S. (englanniksi)
- Fong, Daniel F.; Kane, Thomas C.; Culver, David C.: Vestigialisation and Loss of Nonfunctional Characters. Annual Review of Ecology and Systematics, marraskuu 1995, 26. vsk, nro 4, s. 249–68. doi:10.1146/annurev.es.26.110195.001341. (englanniksi)
- ZhaoLei Zhang; Gerstein, Mark: Large-scale analysis of pseudogenes in the human genome. Current Opinion in Genetics & Development, elokuu 2004, 14. vsk, nro 4, s. 328–35. PubMed:15261647. doi:10.1016/j.gde.2004.06.003. (englanniksi)
- Jeffery, William R.: Adaptive evolution of eye degeneration in the Mexican blind cavefish. Journal of Heredity, toukokuu–kesäkuu 2005, 96. vsk, nro 3, s. 185–96. PubMed:15653557. doi:10.1093/jhered/esi028. (englanniksi)
- Maxwell, Erin E.; Larsson, Hans C.E.: Osteology and myology of the wing of the Emu (Dromaius novaehollandiae) and its bearing on the evolution of vestigial structures. Journal of Morphology, toukokuu 2007, 268. vsk, nro 5, s. 423–41. PubMed:17390336. doi:10.1002/jmor.10527. (englanniksi)
- Silvestri, Anthony R., Jr.; Singh, Iqbal: The unresolved problem of the third molar: would people be better off without it?. Journal of the American Dental Association (1939), huhtikuu 2003, 134. vsk, nro 4, s. 450–5. PubMed:12733778. doi:10.14219/jada.archive.2003.0194. Artikkelin verkkoversio. (englanniksi) (Arkistoitu – Internet Archive)
- Coyne, Jerry A.: Why Evolution is True, s. 62. Penguin Group, 2009. ISBN 978-0-670-02053-9. (englanniksi)
- Darwin, Charles: Tunteiden ilmaisu ihmisissä ja eläimissä. (The expression of the emotions in man and animals, 1872.) Suomentanut Anto Leikola. Helsinki: Terra Cognita, 2009. ISBN 978-952-5697-32-2.
- Gray, Peter: Psychology, s. 66. 5. painos. Worth Publishers, 2007. ISBN 0-7167-0617-2. (englanniksi)
- Coyne, Jerry A.: Why Evolution Is True, s. 85–86. Penguin Group, 2009. ISBN 978-0-670-02053-9. (englanniksi)
- Stevens, Anthony: Archetype: A Natural History of the Self, s. 87. Routledge & Kegan Paul, 1982. ISBN 0-7100-0980-1. (englanniksi)
- Gould, Stephen Jay: The Structure of Evolutionary Theory, s. 1235–1236. Cambridge, MA: Harvard University Press, 2002. ISBN 0-674-00613-5. OCLC 47869352. (englanniksi)
- Pallen, Mark J.; Matzke, Nicholas J.: From The Origin of Species to the origin of bacterial flagella. Nature Reviews Microbiology, lokakuu 2006, 4. vsk, nro 10, s. 784–790. PubMed:16953248. doi:10.1038/nrmicro1493. Artikkelin verkkoversio (PDF). Viitattu 15. elokuuta 2015. (englanniksi)
- Clements, Abigail; Bursac, Dejan; Gatsos, Xenia et al.: The reducible complexity of a mitochondrial molecular machine. Proceedings of the National Academy of Sciences, 15. syyskuuta 2009, 106. vsk, nro 37, s. 15791–15795. PubMed:19717453. doi:10.1073/pnas.0908264106. Bibcode:2009PNAS..10615791C. (englanniksi)
- Piatigorsky, Joram; Kantorow, Marc; Gopal-Srivastava, Rashmi; Tomarev, Stanislav I.: Recruitment of enzymes and stress proteins as lens crystallins. Experientia, 1994, 71. vsk, s. 241–250. PubMed:8032155. doi:10.1007/978-3-0348-7330-7_24. (englanniksi)
- Wistow, Graeme: Lens crystallins: gene recruitment and evolutionary dynamism. Trends in Biochemical Sciences, elokuu 1993, 18. vsk, nro 8, s. 301–306. PubMed:8236445. doi:10.1016/0968-0004(93)90041-K. (englanniksi)
- Johnson, Norman A.; Porter, Adam H.: Toward a new synthesis: population genetics and evolutionary developmental biology. Genetica, marraskuu 2001, 112–113. vsk, s. 45–58. Dordrecht: PubMed:11838782. doi:10.1023/A:1013371201773. (englanniksi)
- Baguñà, Jaume; Garcia-Fernàndez, Jordi: Evo-Devo: the long and winding road. The International Journal of Developmental Biology, 2003, 47. vsk, nro 7–8, s. 705–713. PubMed:14756346. Artikkelin verkkoversio. (englanniksi)
- Love, Alan C.: Evolutionary Morphology, Innovation and the Synthesis of Evolutionary and Developmental Biology. Biology and Philosophy, maaliskuu 2003, 18. vsk, nro 2, s. 309–345. Dordrecht: doi:10.1023/A:1023940220348. (englanniksi)
- Allin, Edgar F.: Evolution of the mammalian middle ear. Journal of Morphology, joulukuu 1975, 147. vsk, nro 4, s. 403–437. PubMed:1202224. doi:10.1002/jmor.1051470404. (englanniksi)
- Harris, Matthew P.; Hasso, Sean M.; Ferguson, Mark W. J.; Fallon, John F.: The development of archosaurian first-generation teeth in a chicken mutant. Current Biology, 2006, 16. vsk, nro 4, s. 371–7. PubMed:16488870. doi:10.1016/j.cub.2005.12.047. (englanniksi)
- Carroll, Sean B.: Evo-Devo and an Expanding Evolutionary Synthesis: A Genetic Theory of Morphological Evolution. Cell, 11. heinäkuuta 2008, 134. vsk, nro 1, s. 25–36. PubMed:18614008. doi:10.1016/j.cell.2008.06.030. (englanniksi)
- Wade, Michael J.: The co-evolutionary genetics of ecological communities. Nature Reviews Genetics, maaliskuu 2007, 8. vsk, nro 3, s. 185–195. Lontoo: PubMed:17279094. doi:10.1038/nrg2031. (englanniksi)
- Geffeney, Shana; Brodie, Edmund D., Jr.; Ruben, Peter C.; Brodie, Edmund D., III: Mechanisms of adaptation in a predator-prey arms race: TTX-resistant sodium channels. Science, 23. elokuuta 2002, 297. vsk, nro 5585, s. 1336–1339. PubMed:12193784. doi:10.1126/science.1074310. Bibcode:2002Sci...297.1336G. (englanniksi)
- Brodie, Edmund D., Jr.; Ridenhour, Benjamin J.; Brodie, Edmund D., III: The evolutionary response of predators to dangerous prey: hotspots and coldspots in the geographic mosaic of coevolution between garter snakes and newts. Evolution, lokakuu 2002, 56. vsk, nro 10, s. 2067–2082. PubMed:12449493. doi:10.1554/0014-3820(2002)056[2067:TEROPT]2.0.CO;2. (englanniksi)
- Carroll, Sean B.: Remarkable Creatures – Clues to Toxins in Deadly Delicacies of the Animal Kingdom 21. joulukuuta 2009. New York Times. (englanniksi)
- Sachs, Joel L.: Cooperation within and among species. Journal of Evolutionary Biology, syyskyy 2006, 19. vsk, nro 5, s. 1415–1418; discussion 1426–1436. PubMed:16910971. doi:10.1111/j.1420-9101.2006.01152.x. (englanniksi)
- Nowak, Martin A.: Five rules for the evolution of cooperation. Science, 8. joulukuuta 2006, 314. vsk, nro 5805, s. 1560–1563. PubMed:17158317. doi:10.1126/science.1133755. Bibcode:2006Sci...314.1560N. (englanniksi)
- Paszkowski, Uta: Mutualism and parasitism: the yin and yang of plant symbioses. Current Opinion in Plant Biology, elokuu 2006, 9. vsk, nro 4, s. 364–370. PubMed:16713732. doi:10.1016/j.pbi.2006.05.008. (englanniksi)
- Hause, Bettina; Fester, Thomas: Molecular and cell biology of arbuscular mycorrhizal symbiosis. Planta, toukokuu 2005, 221. vsk, nro 2, s. 184–196. Berliini: PubMed:15871030. doi:10.1007/s00425-004-1436-x. (englanniksi)
- Bertram, John S.: The molecular biology of cancer. Molecular Aspects of Medicine, joulukuu 2000, 21. vsk, nro 6, s. 167–223. Amsterdam: PubMed:11173079. doi:10.1016/S0098-2997(00)00007-8. (englanniksi)
- Reeve, H. Kern; Hölldobler, Bert: The emergence of a superorganism through intergroup competition. Proc Natl Acad Sci U S A., 5. kesäkuuta 2007, 104. vsk, nro 23, s. 9736–40. PubMed:17517608. doi:10.1073/pnas.0703466104. Bibcode:2007PNAS..104.9736R. (englanniksi)
- Axelrod, Robert; Hamilton, W. D.: The evolution of cooperation. Science, 27. maaliskuuta 1981, 211. vsk, nro 4489, s. 1390–6. PubMed:7466396. doi:10.1126/science.7466396. Bibcode:1981Sci...211.1390A. (englanniksi)
- Wilson, Edward O.; Hölldobler, Bert: Eusociality: origin and consequences. Proc. Natl. Acad. Sci. U.S.A., 20. syyskuuta 2005, 102. vsk, nro 38, s. 13367–71. PubMed:16157878. doi:10.1073/pnas.0505858102. Bibcode:2005PNAS..10213367W. (englanniksi)
- Gavrilets, Sergey: Perspective: models of speciation: what have we learned in 40 years?. Evolution, lokakuu 2003, 57. vsk, nro 10, s. 2197–2215. PubMed:14628909. doi:10.1554/02-727. (englanniksi)
- de Queiroz, Kevin: Ernst Mayr and the modern concept of species. Proc. Natl. Acad. Sci. U.S.A., 3. toukokuuta 2005, 102. vsk, nro Suppl 1, s. 6600–6607. PubMed:15851674. doi:10.1073/pnas.0502030102. Bibcode:2005PNAS..102.6600D. (englanniksi)
- Ereshefsky, Marc: Eliminative pluralism. Philosophy of Science, joulukuu 1992, 59. vsk, nro 4, s. 671–690. doi:10.1086/289701. JSTOR 188136. (englanniksi)
- Mayr, E.: Systematics and the Origin of Species, s. 120. New York: Columbia Univ. Press, 1942. ISBN 978-0-231-05449-2. (englanniksi) Lainaus: "Species are groups of actually or potentially interbreeding natural populations, which are reproductively isolated from other such groups."
- Fraser, Christophe; Alm, Eric J.; Polz, Martin F. et al.: The bacterial species challenge: making sense of genetic and ecological diversity. Science, 6. helmikuuta 2009, 323. vsk, nro 5915, s. 741–746. PubMed:19197054. doi:10.1126/science.1159388. Bibcode:2009Sci...323..741F. (englanniksi)
- Short, Roger Valentine: The contribution of the mule to scientific thought. Journal of Reproduction and Fertility. Supplement, lokakuu 1975, nro 23, s. 359–364. PubMed:1107543. (englanniksi)
- Gross, Briana L.; Rieseberg, Loren H.: The Ecological Genetics of Homoploid Hybrid Speciation. Journal of Heredity, toukokuu–kesäkuu 2005, 96. vsk, nro 3, s. 241–52. PubMed:15618301. doi:10.1093/jhered/esi026. (englanniksi)
- Burke, John M.; Arnold, Michael L.: Genetics and the fitness of hybrids. Annu. Rev. Genet., joulukuu 2001, 35. vsk, nro 1, s. 31–52. PubMed:11700276. doi:10.1146/annurev.genet.35.102401.085719. (englanniksi)
- Vrijenhoek, Robert C.: Polyploid hybrids: multiple origins of a treefrog species. Current Biology, 4. huhtikuuta 2006, 16. vsk, nro 7, s. R245–7. PubMed:16581499. doi:10.1016/j.cub.2006.03.005. (englanniksi)
- Rice, William R.; Hostert, Ellen E.: Laboratory experiments on speciation: what have we learned in 40 years. Evolution, joulukuu 1993, 47. vsk, nro 6, s. 1637–1653. doi:10.2307/2410209. (englanniksi)
- Jiggins, Chris D.; Bridle, Jon R.: Speciation in the apple maggot fly: a blend of vintages?. rends in Ecology & Evolution, maaliskuu 2004, 19. vsk, nro 3, s. 111–114. PubMed:16701238. doi:10.1016/j.tree.2003.12.008. (englanniksi)
- Boxhorn, Joseph: Observed Instances of Speciation 1. syyskuuta 1995. TalkOrigins Archive. Viitattu 15. elokuuta 2015. (englanniksi)
- Weinberg, James R.; Starczak, Victoria R.; Jörg, Daniele: Evidence for Rapid Speciation Following a Founder Event in the Laboratory. Evolution, elokuu 1992, 46. vsk, nro 4, s. 1214–20. doi:10.2307/2409766. JSTOR 2409766. (englanniksi)
- Herrel, Anthony; Huyghe, Katleen; Vanhooydonck, Bieke et al.: Rapid large-scale evolutionary divergence in morphology and performance associated with exploitation of a different dietary resource. Proceedings of the National Academy of Sciences, 25. maaliskuuta 2008, 105. vsk, nro 12, s. 4792–5. PubMed:18344323. doi:10.1073/pnas.0711998105. Bibcode:2008PNAS..105.4792H. (englanniksi)
- Losos, Jonathan B.; Warhelt, Kenneth I.; Schoener, Thomas W.: Adaptive differentiation following experimental island colonization in Anolis lizards. Nature, 1. toukokuuta 1997, 387. vsk, nro 6628, s. 70–3. doi:10.1038/387070a0. Bibcode:1997Natur.387...70L. (englanniksi)
- Hoskin, Conrad J.; Higgle, Megan; McDonald, Keith R.; Moritz, Craig: Reinforcement drives rapid allopatric speciation. Nature, 27. lokakuuta 2005, 437. vsk, nro 7063, s. 1353–1356. PubMed:16251964. doi:10.1038/nature04004. Bibcode:2005Natur.437.1353H. (englanniksi)
- Templeton, Alan R.: The theory of speciation via the founder principle. Genetics, 1. huhtikuuta 1980, 94. vsk, nro 4, s. 1011–38. PubMed:6777243. Artikkelin verkkoversio. (englanniksi)
- Antonovics, Janis: Evolution in closely adjacent plant populations X: long-term persistence of prereproductive isolation at a mine boundary. Heredity, heinäkuu 2006, 97. vsk, nro 1, s. 33–7. PubMed:16639420. doi:10.1038/sj.hdy.6800835. Artikkelin verkkoversio. (englanniksi)
- Nosil, Patrik; Crespi, Bernard J.; Gries, Regine; Gries, Gerhard: Natural selection and divergence in mate preference during speciation. Genetica, maaliskuu 2007, 129. vsk, nro 3, s. 309–27. PubMed:16900317. doi:10.1007/s10709-006-0013-6. (englanniksi)
- Savolainen, Vincent; Anstett, Marie-Charlotte; Lexer, Christian et al.: Sympatric speciation in palms on an oceanic island. Nature, 11. toukokuuta 2006, 441. vsk, nro 7090, s. 210–3. PubMed:16467788. doi:10.1038/nature04566. Bibcode:2006Natur.441..210S. (englanniksi)
- Barluenga, Marta; Stölting, Kai N.; Salzburger, Walter et al.: Sympatric speciation in Nicaraguan crater lake cichlid fish. Nature, 9. helmikuuta 2006, 439. vsk, nro 7077, s. 719–23. PubMed:16467837. doi:10.1038/nature04325. Bibcode:2006Natur.439..719B. (englanniksi)
- Gavrilets, Sergey: The Maynard Smith model of sympatric speciation. Journal of Theoretical Biology, 21. maaliskuuta 2006, 239. vsk, nro 2, s. 172–82. Amsterdam: PubMed:16242727. doi:10.1016/j.jtbi.2005.08.041. (englanniksi)
- Wood, Troy E.; Takebayashi, Naoki; Barker, Michael S. et al.: The frequency of polyploid speciation in vascular plants. Proc. Natl. Acad. Sci. U.S.A., 18. elokuuta 2009, 106. vsk, nro 33, s. 13875–9. PubMed:19667210. doi:10.1073/pnas.0811575106. Bibcode:2009PNAS..10613875W. (englanniksi)
- Hegarty, Matthew J.; Hiscock, Simon J.: Genomic clues to the evolutionary success of polyploid plants. Current Biology, 20. toukokuuta 2008, 18. vsk, nro 10, s. 435–44. PubMed:18492478. doi:10.1016/j.cub.2008.03.043. (englanniksi)
- Jakobsson, Mattias; Hagenblad, Jenny; Tavaré, Simon et al.: A unique recent origin of the allotetraploid species Arabidopsis suecica: Evidence from nuclear DNA markers. Molecular Biology and Evolution, kesäkuu 2006, 23. vsk, nro 6, s. 1217–31. PubMed:16549398. doi:10.1093/molbev/msk006. (englanniksi)
- Säll, Torbjörn; Jakobsson, Mattias; Lind-Halldén, Christina; Halldén, Christer: Chloroplast DNA indicates a single origin of the allotetraploid Arabidopsis suecica. Journal of Evolutionary Biology, syyskuu 2003, 16. vsk, nro 5, s. 1019–29. PubMed:14635917. doi:10.1046/j.1420-9101.2003.00554.x. (englanniksi)
- Bomblies, Kirsten; Weigel, Detlef: Arabidopsis-a model genus for speciation. Current Opinion in Genetics & Development, joulukuu 2007, 17. vsk, nro 6, s. 500–4. PubMed:18006296. doi:10.1016/j.gde.2007.09.006. (englanniksi)
- Sémon, Marie; Wolfe, Kenneth H.: Consequences of genome duplication. Current Opinion in Genetics & Development, joulukuu 2007, 17. vsk, nro 6, s. 505–12. PubMed:18006297. doi:10.1016/j.gde.2007.09.007. (englanniksi)
- Eldredge, Niles; Gould, Stephen Jay: ”Punctuated equilibria: an alternative to phyletic gradualism”, Models in Paleobiology, s. 82–115. San Francisco, CA: Freeman, Cooper, 1972. ISBN 0-87735-325-5. OCLC 572084. (englanniksi)
- Benton, Michael J.: Diversification and extinction in the history of life. Science, 7. huhtikuuta 1995, 268. vsk, nro 5207, s. 52–8. PubMed:7701342. doi:10.1126/science.7701342. Bibcode:1995Sci...268...52B. (englanniksi)
- Raup, David M.: Biological extinction in Earth history. Science, 28. maaliskuuta 1986, 231. vsk, nro 4745, s. 1528–33. PubMed:11542058. doi:10.1126/science.11542058. Bibcode:1986Sci...231.1528R. (englanniksi)
- Avise, John C.; Hubbell, Stephen P.; Ayala, Francisco J.: In the light of evolution II: Biodiversity and extinction. Proc. Natl. Acad. Sci. U.S.A., 12. elokuuta 2008, 105. vsk, nro Suppl. 1, s. 11453–11457. PubMed:18695213. doi:10.1073/pnas.0802504105. ISSN 0027-8424. Bibcode:2008PNAS..10511453A. (englanniksi)
- Raup, David M.: The role of extinction in evolution. Proc. Natl. Acad. Sci. U.S.A., 19. heinäkuuta 1994, 91. vsk, nro 15, s. 6758–63. PubMed:8041694. doi:10.1073/pnas.91.15.6758. Bibcode:1994PNAS...91.6758R. (englanniksi)
- Novacek, Michael J.; Cleland, Elsa E.: The current biodiversity extinction event: scenarios for mitigation and recovery. Proc. Natl. Acad. Sci. U.S.A., 8. toukokuuta 2001, 98. vsk, nro 10, s. 5466–70. PubMed:11344295. doi:10.1073/pnas.091093698. Bibcode:2001PNAS...98.5466N. (englanniksi)
- Pimm, Stuart; Raven, Peter; Peterson, Alan et al.: Human impacts on the rates of recent, present and future bird extinctions. Proc. Natl. Acad. Sci. U.S.A., 18. heinäkuuta 2006, 103. vsk, nro 29, s. 10941–10946. PubMed:16829570. doi:10.1073/pnas.0604181103. Bibcode:2006PNAS..10310941P. (englanniksi)
- Barnosky, Anthony D.; Koch, Paul L.; Feranec, Robert S. et al.: Assessing the causes of late Pleistocene extinctions on the continents. Science, 2004, 306. vsk, nro 5693, s. 70–75. PubMed:15459379. doi:10.1126/science.1101476. Bibcode:2004Sci...306...70B. (englanniksi)
- Lewis, Owen T.: Climate change, species-area curves and the extinction crisis. Philosophical Transactions of the Royal Society B, 29. tammikuuta 2006, 361. vsk, nro 1465, s. 163–71. PubMed:16553315. doi:10.1098/rstb.2005.1712. (englanniksi)
- Jablonski, David: Lessons from the past: evolutionary impacts of mass extinctions. Proc. Natl. Acad. Sci. U.S.A., 8. toukokuuta 2001, 98. vsk, nro 10, s. 5393–8. PubMed:11344284. doi:10.1073/pnas.101092598. Bibcode:2001PNAS...98.5393J. (englanniksi)
- Peretó, Juli: Controversies on the origin of life. Int. Microbiol., maaliskuu 2005, 8. vsk, nro 1, s. 23–31. PubMed:15906258. Artikkelin verkkoversio. (englanniksi) (Arkistoitu – Internet Archive)
- Joyce, Gerald F.: The antiquity of RNA-based evolution. Nature, 11. heinäkuuta 2002, 418. vsk, nro 6894, s. 214–21. PubMed:12110897. doi:10.1038/418214a. Bibcode:2002Natur.418..214J. (englanniksi)
- Trevors, Jack T.; Psenner, Roland: From self-assembly of life to present-day bacteria: a possible role for nanocells. FEMS Microbiol. Rev., joulukuu 2001, 25. vsk, nro 5, s. 573–82. PubMed:11742692. doi:10.1111/j.1574-6976.2001.tb00592.x. (englanniksi)
- Theobald, Douglas L.: A formal test of the theory of universal common ancestry. Nature, 13. toukokuuta 2010, 465. vsk, nro 7295, s. 219–22. PubMed:20463738. doi:10.1038/nature09014. Bibcode:2010Natur.465..219T. (englanniksi)
- Bapteste, Eric; Walsh, David A.: Does the 'Ring of Life' ring true?. Trends Microbiol., kesäkuu 2005, 13. vsk, nro 6, s. 256–61. PubMed:15936656. doi:10.1016/j.tim.2005.03.012. (englanniksi)
- Darwin, Charles: On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life. 1. painos. London: John Murray, 1859. OCLC 741260650. (englanniksi)
- Doolittle, W. Ford; Bapteste, Eric: Pattern pluralism and the Tree of Life hypothesis. Proc. Natl. Acad. Sci. U.S.A., 13. helmikuuta 2007, 104. vsk, nro 7, s. 2043–9. PubMed:17261804. doi:10.1073/pnas.0610699104. Bibcode:2007PNAS..104.2043D. (englanniksi)
- Kunin, Victor; Goldovsky, Leon; Darzentas, Nikos; Ouzounis, Christos A.: The net of life: reconstructing the microbial phylogenetic network. Genome Res., heinäkuu 2005, 15. vsk, nro 7, s. 954–9. PubMed:15965028. doi:10.1101/gr.3666505. (englanniksi)
- Jablonski D: The future of the fossil record. Science, 1999, 284. vsk, nro 5423, s. 2114–16. PubMed:10381868. doi:10.1126/science.284.5423.2114. (englanniksi)
- Mason SF: Origins of biomolecular handedness. Nature, 1984, 311. vsk, nro 5981, s. 19–23. PubMed:6472461. doi:10.1038/311019a0. Bibcode:1984Natur.311...19M. (englanniksi)
- Wolf YI, Rogozin IB, Grishin NV, Koonin EV: Genome trees and the tree of life. Trends Genet., 2002, 18. vsk, nro 9, s. 472–79. PubMed:12175808. doi:10.1016/S0168-9525(02)02744-0. (englanniksi)
- Varki A, Altheide TK: Comparing the human and chimpanzee genomes: searching for needles in a haystack. Genome Res., 2005, 15. vsk, nro 12, s. 1746–58. PubMed:16339373. doi:10.1101/gr.3737405. (englanniksi)
- Ciccarelli FD, Doerks T, von Mering C, Creevey CJ, Snel B, Bork P: Toward automatic reconstruction of a highly resolved tree of life. Science, 2006, 311. vsk, nro 5765, s. 1283–87. PubMed:16513982. doi:10.1126/science.1123061. Bibcode:2006Sci...311.1283C. (englanniksi)
- Cavalier-Smith T: Cell evolution and Earth history: stasis and revolution. Philosophical Transactions of the Royal Society B, 2006, 361. vsk, nro 1470, s. 969–1006. PubMed:16754610. doi:10.1098/rstb.2006.1842. (englanniksi)
- Schopf J: Fossil evidence of Archaean life. Philosophical Transactions of the Royal Society B, 2006, 361. vsk, nro 1470, s. 869–85. PubMed:16754604. doi:10.1098/rstb.2006.1834. (englanniksi)
- Altermann W, Kazmierczak J: Archean microfossils: a reappraisal of early life on Earth. Res Microbiol, 2003, 154. vsk, nro 9, s. 611–17. PubMed:14596897. doi:10.1016/j.resmic.2003.08.006. (englanniksi)
- Schopf J: Disparate rates, differing fates: tempo and mode of evolution changed from the Precambrian to the Phanerozoic. Proc Natl Acad Sci U S A, 1994, 91. vsk, nro 15, s. 6735–42. PubMed:8041691. doi:10.1073/pnas.91.15.6735. Bibcode:1994PNAS...91.6735S. (englanniksi)
- Poole A, Penny D: Evaluating hypotheses for the origin of eukaryotes. BioEssays, 2007, 29. vsk, nro 1, s. 74–84. PubMed:17187354. doi:10.1002/bies.20516. (englanniksi)
- Dyall S, Brown M, Johnson P: Ancient invasions: from endosymbionts to organelles. Science, 2004, 304. vsk, nro 5668, s. 253–57. PubMed:15073369. doi:10.1126/science.1094884. Bibcode:2004Sci...304..253D. (englanniksi)
- Martin W: The missing link between hydrogenosomes and mitochondria. Trends Microbiol., 2005, 13. vsk, nro 10, s. 457–59. PubMed:16109488. doi:10.1016/j.tim.2005.08.005. (englanniksi)
- Lang B, Gray M, Burger G: Mitochondrial genome evolution and the origin of eukaryotes. Annu Rev Genet, 1999, 33. vsk, nro 1, s. 351–97. PubMed:10690412. doi:10.1146/annurev.genet.33.1.351. (englanniksi)
- McFadden G: Endosymbiosis and evolution of the plant cell. Current Opinion in Plant Biology, 1999, 2. vsk, nro 6, s. 513–19. PubMed:10607659. doi:10.1016/S1369-5266(99)00025-4. (englanniksi)
- DeLong E, Pace N: Environmental diversity of bacteria and archaea. Syst Biol, 2001, 50. vsk, nro 4, s. 470–8. PubMed:12116647. doi:10.1080/106351501750435040. (englanniksi)
- Kaiser D: Building a multicellular organism. Annu. Rev. Genet., 2001, 35. vsk, nro 1, s. 103–23. PubMed:11700279. doi:10.1146/annurev.genet.35.102401.090145. (englanniksi)
- Valentine JW, Jablonski D, Erwin DH: Fossils, molecules and embryos: new perspectives on the Cambrian explosion. Development, March 1, 1999, 126. vsk, nro 5, s. 851–9. PubMed:9927587. Artikkelin verkkoversio. (englanniksi)
- Ohno S: The reason for as well as the consequence of the Cambrian explosion in animal evolution. J. Mol. Evol., 1997, 1. vsk, nro S1, s. S23–7. PubMed:9071008. doi:10.1007/PL00000055. (englanniksi)
- Valentine J, Jablonski D: Morphological and developmental macroevolution: a paleontological perspective. Int. J. Dev. Biol., 2003, 47. vsk, nro 7–8, s. 517–22. PubMed:14756327. Artikkelin verkkoversio. (englanniksi)
- Waters ER: Molecular adaptation and the origin of land plants. Mol. Phylogenet. Evol., 2003, 29. vsk, nro 3, s. 456–63. PubMed:14615186. doi:10.1016/j.ympev.2003.07.018. (englanniksi)
- Mayhew PJ: Why are there so many insect species? Perspectives from fossils and phylogenies. Biol Rev Camb Philos Soc, 2007, 82. vsk, nro 3, s. 425–54. PubMed:17624962. doi:10.1111/j.1469-185X.2007.00018.x. (englanniksi)
- Carroll, Robert L.: The Palaeozoic Ancestry of Salamanders, Frogs and Caecilians. Zool J Linn Soc, May 2007, 150. vsk, nro s1, s. 1–140. PubMed:12752770. doi:10.1111/j.1096-3642.2007.00246.x. (englanniksi)
- Wible JR, Rougier GW, Novacek MJ, Asher RJ: Cretaceous eutherians and Laurasian origin for placental mammals near the K/T boundary. Nature, 2007, 447. vsk, nro 7147, s. 1003–6. PubMed:17581585. doi:10.1038/nature05854. Bibcode:2007Natur.447.1003W. (englanniksi)
- Witmer LM: Palaeontology: An icon knocked from its perch. Nature, 2011, 475. vsk, nro 7357, s. 458–459. PubMed:21796198. doi:10.1038/475458a. (englanniksi)
- Bull JJ, Wichman HA: Applied evolution. Annu Rev Ecol Syst, 2001, 32. vsk, nro 1, s. 183–217. doi:10.1146/annurev.ecolsys.32.081501.114020. (englanniksi)
- Doebley JF, Gaut BS, Smith BD: The molecular genetics of crop domestication. Cell, 2006, 127. vsk, nro 7, s. 1309–21. PubMed:17190597. doi:10.1016/j.cell.2006.12.006. (englanniksi)
- Jäckel C, Kast P, Hilvert D: Protein design by directed evolution. Annu Rev Biophys, 2008, 37. vsk, nro 1, s. 153–73. PubMed:18573077. doi:10.1146/annurev.biophys.37.032807.125832. (englanniksi)
- Merlo, LM; Pepper, JW; Reid, BJ; Maley, CC: Cancer as an evolutionary and ecological process.. Nature reviews. Cancer, Joulukuu 2006, 6. vsk, nro 12, s. 924–35. PubMed:17109012. doi:10.1038/nrc2013. (englanniksi)
- Pan, D; Xue, W; Zhang, W; Liu, H; Yao, X: Understanding the drug resistance mechanism of hepatitis C virus NS3/4A to ITMN-191 due to R155K, A156V, D168A/E mutations: a computational study.. Biochimica et Biophysica Acta, Lokakuu 2012, 1820. vsk, nro 10, s. 1526–34. PubMed:22698669. doi:10.1016/j.bbagen.2012.06.001. (englanniksi)
- Woodford, N.; Ellington, MJ: The emergence of antibiotic resistance by mutation.. Clinical microbiology and infection : the official publication of the European Society of Clinical Microbiology and Infectious Diseases, Tammikuu 2007, 13. vsk, nro 1, s. 5–18. PubMed:17184282. doi:10.1111/j.1469-0691.2006.01492.x. (englanniksi)
- Labbé, P; Berticat, C; Berthomieu, A; Unal, S; Bernard, C; Weill, M; Lenormand, T: Forty years of erratic insecticide resistance evolution in the mosquito Culex pipiens.. PLoS genetics, Marraskuu 2007, 3. vsk, nro 11, s. e205. PubMed:18020711. doi:10.1371/journal.pgen.0030205. (englanniksi)
- Neve, P: Challenges for herbicide resistance evolution and management: 50 years after Harper. Weed Research, Lokakuu 2007, 47. vsk, nro 5, s. 365–369. doi:10.1111/j.1365-3180.2007.00581.x. (englanniksi)
- Fraser AS: Monte Carlo analyses of genetic models. Nature, 1958, 181. vsk, nro 4603, s. 208–9. PubMed:13504138. doi:10.1038/181208a0. Bibcode:1958Natur.181..208F. (englanniksi)
- Rechenberg, Ingo: Evolutionsstrategie – Optimierung technischer Systeme nach Prinzipien der biologischen Evolution (PhD thesis). Fromman-Holzboog, 1973. (saksaksi)
- Holland, John H.: Adaptation in Natural and Artificial Systems. University of Michigan Press, 1975. ISBN 0-262-58111-6. (englanniksi)
- Koza, John R.: Genetic Programming: On the Programming of Computers by Means of Natural Selection. MIT Press, 1992. ISBN 0-262-11170-5. (englanniksi)
- Jamshidi M: Tools for intelligent control: fuzzy controllers, neural networks and genetic algorithms. Philosophical Transactions of the Royal Society A, 2003, 361. vsk, nro 1809, s. 1781–808. PubMed:12952685. doi:10.1098/rsta.2003.1225. Bibcode:2003RSPTA.361.1781J. (englanniksi)
- Jämsä, Tuomo: Erottaako kieli ihmisen? Semiotiikan perusteet. Internetix. Arkistoitu 15.12.2004. Viitattu 15. elokuuta 2015.
- Leikola, Anto: Ihminen on osa luontoa, sanoi Darwin. Tiede, 1/2009, s. 28–31. Artikkelin tekstissä vertauskohteena on Isaac Newton.
- Yleiskuva filosofisista, uskonnollisista ja kosmologisista kiistanalaisuuksista: Dennett, D.: Darwin's Dangerous Idea. Simon & Schuster, 1995. ISBN 978-0-684-82471-0. (englanniksi)
- Evoluutioteorian tieteellisestä ja yhteiskunnallisesta vastaanotosta 19. ja 20. vuosisadan aikana: Johnston, Ian C.: History of Science: Origins of Evolutionary Theory Liberal Studies Department, Malaspina University College. Arkistoitu 7.1.2017. Viitattu 15. elokuuta 2015. (englanniksi)
- Bowler, Peter J.: Evolution: The History of an Idea. 3. painos. Berkeley, CA: University of California Press, 2003. ISBN 0-520-23693-9. OCLC 49824702. (englanniksi)
- Zuckerkandl E: Intelligent design and biological complexity. Gene, 2006, 385. vsk, s. 2–18. PubMed:17011142. doi:10.1016/j.gene.2006.03.025. (englanniksi)
- Ross, M.R.: Who Believes What? Clearing up Confusion over Intelligent Design and Young-Earth Creationism. Journal of Geoscience Education, 2005, 53. vsk, nro 3. Artikkelin verkkoversio. Viitattu 15. elokuuta 2015. (englanniksi)
- Hameed, Salman: Science and Religion: Bracing for Islamic Creationism. Science, 12.12.2008, 322. vsk, nro 5908, s. 1637–1638. PubMed:19074331. doi:10.1126/science.1163672. Artikkelin verkkoversio. Viitattu 15. elokuuta 2015. (englanniksi) (Arkistoitu – Internet Archive)
- Bowler, Peter J.: Evolution: The History of an Idea. 3. painos. Berkeley, CA: University of California Press, 2003. ISBN 0-520-23693-9. OCLC 49824702. (englanniksi)
- Miller, JD; Scott, EC; Okamoto, S: Public Acceptance of Evolution. Science, 2006, 313. vsk, nro 5788, s. 765–66. PubMed:16902112. doi:10.1126/science.1126746. (englanniksi)
- Hakusana kreationismi teoksessa Selin, Risto: Ihmeellinen maailma: Skeptikon tietosanakirja. Ursan julkaisuja 81. Helsinki: Tähtitieteellinen yhdistys Ursa, 2001. ISBN 952-5329-19-4. Teoksen verkkoversio (viitattu 3.2.2011).
- Spergel, D. N.; Verde, L.; Peiris, H. V.; Komatsu, E.; Nolta, M. R.; Bennett, C. L.; Halpern, M.; Hinshaw, G. et al.: First-Year Wilkinson Microwave Anisotropy Probe (WMAP) Observations: Determination of Cosmological Parameters. The Astrophysical Journal Supplement Series, 2003, 148. vsk, nro 1, s. 175–94. doi:10.1086/377226. Bibcode:2003ApJS..148..175S. (englanniksi)
- Wilde SA, Valley JW, Peck WH, Graham CM: Evidence from detrital zircons for the existence of continental crust and oceans on the Earth 4.4 Gyr ago. Nature, 2001, 409. vsk, nro 6817, s. 175–78. PubMed:11196637. doi:10.1038/35051550. (englanniksi)
- 65 prosenttia suomalaisista uskoo evoluutioteoriaan 22.8.2006. Helsingin Sanomat. Arkistoitu 1.9.2014. Viitattu 5.2.2015.
- Kiljunen, Pentti: Tiedebarometri 2019 (PDF) 14.10.2019. Tieteen tiedotus ry. Viitattu 6.5.2020.
Kirjallisuutta
- Johdantoteoksia
- Bargum, Katja & Kokko, Hanna: Kutistuva turska ja muita evoluution ihmeitä. Helsinki: WSOY, 2008. ISBN 978-951-0-34020-2.
- Carroll, Sean B.: Loputtomat kauniit muodot: Evodevon uusi tiede. (Endless forms most beautiful: The new science of evo devo, 2005.). Suomentanut Kimmo Pietiläinen. Helsinki: Terra Cognita, 2007. ISBN 978-952-5697-03-2.
- Coyne, Jerry A.: Miksi evoluutio on totta. (Why evolution is true, 2009.). Suomentanut Risto Suikkanen. Tampere: Vastapaino, 2011. ISBN 978-951-768-376-0.
- Dawkins, Richard: Geenin itsekkyys. (The selfish gene, 1976, 1989.). Suomentanut Kimmo Pietiläinen. Helsinki: Art House, 1993. ISBN 951-884-107-1.
- Dawkins, Richard: Sokea kelloseppä. (The blind watchmaker, 1986.). Suomentanut Risto Varteva. Helsinki: WSOY, 1989. ISBN 951-0-15863-1.
- Dawkins, Richard: Maailman hienoin esitys: Evoluution todisteet. (The greatest show on Earth: The evidence for evolution, 2009.). Suomentanut Kimmo Pietiläinen. Helsinki: Terra cognita, 2009. ISBN 978-952-5697-27-8.
- Gould, Stephen Jay: Ihmeellinen elämä: Burgessin esiintymä ja historian kulku. (Wonderful life: The Burgess shale and the nature of history, 1989.). Suomentanut Outi Nummi ja Markus Nummi. Helsinki: Art House, 1991. ISBN 951-884-088-1.
- Hanski, Ilkka & Niniluoto, Ilkka & Hetemäki, Ilari (toim.): Kaikki evoluutiosta. Helsinki: Gaudeamus, 2009. ISBN 978-952-495-098-5.
- Portin, Petter & Vuorisalo, Timo (toim.): Evoluutio NYT! Charles Darwinin juhlaa. Turku: Kirja-Aurora, 2008. ISBN 978-951-29-3666-3.
- Weiner, Jonathan: Darwinin linnut. (The beak of the finch: A story of evolution in our time, 1994.). Suomentanut Tuula Hollmén. Helsinki: Otava, 1997. ISBN 951-1-13805-7.
- Juha Valste: Evoluutio – Miten lajit kehittyvät. SKS, 2018. ISBN 978-952-222-924-3.
- Historia
- Browne, Janet: Darwinin Lajien synty. (Darwin’s Origin of species, 2007.). Suomentanut Riitta Virkkunen. Helsinki: Ajatus, 2009. ISBN 978-951-20-7441-9.
- Darwin, Charles: Lajien synty. (Origin of Species, 1859.). Suomentanut Pertti Ranta. Tampere: Osuuskunta Vastapaino, 2009. ISBN 978-951-768-236-7.
- Oppikirjoja
- Björklund, Mats: Evoluutiobiologia. Helsinki: Gaudeamus, 2008. ISBN 978-952-495-076-3.
- Cracraft, Joel; Bybee, Rodger W.: Evolutionary Science and Society: Educating a New Generation. Colorado Springs, CO: Biological Sciences Curriculum Study, 2005. ISBN 1-929614-23-3. Teoksen verkkoversio (PDF) (viitattu 15.8.2015). OCLC 64228003. (englanniksi) (Arkistoitu – Internet Archive)
- Futuyma, Douglas J.: Evolutionary Biology. Third Edition. Sunderland, Massachusetts: Sinauer Associates, 1998. ISBN 0-87893-189-9. (englanniksi)
- Laihonen, Pasi & Salo, Jukka & Vuorisalo, Timo: Evoluutio: Miten elämä kehittyy. Helsinki: Otava, 1986. ISBN 951-1-08486-0.
Aiheesta muualla
- Mitä evoluutio on? (Opetus.tv videopalvelu)
- Evoluutio 2009 -juhlavuosi (Arkistoitu – Internet Archive). Luonnontieteellinen keskusmuseo.
- Evoluutio ja evoluutioteoria – Elämän kehitys nykytieteen valossa. Biomi.org.
- Evoluutio. Internetix.
- Evoluutio (Arkistoitu – Internet Archive). Etälukio.
- Sloan, Phillip: The Concept of Evolution to 1872 The Stanford Encyclopedia of Philosophy. The Metaphysics Research Lab. Stanford University. (englanniksi)