Niasiini
Niasiinilla eli B3-vitamiinilla tarkoitetaan sekä nikotiiniamidia että nikotiinihappoa. Nämä ovat vastaavasti pyridiinin meta-substituoitu karboksyylihappo ja amidi. Molemmat toimivat monissa eliöissä vitamiineina.[1][2] ihmisen elimistö pystyy hyödyntämään eläinkunnan tuotteiden sisältämää nikotiiniamidia huomattavasti paremmin kuin kasvikunnan tuotteiden sisältämää nikotiinihappoa[3].

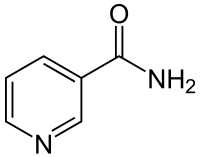
Joskus niasiiniksi luetaan myös NADH, NADPH ja muita B3-vitamiinitoimintoisia pyridiinien nukleotidejä.[2] Tässä artikkelissa niasiini tarkoittaa pääsääntöisesti kaikkia aineita, jotka ovat B3-vitamiinitoimintoisia ainakin ihmisillä.
Ihmisten ja muiden eliöiden tulee saada ravinnosta riittävästi niasiinia tai niasiinin esiasteena toimivaa tryptofaania[4]. Niasiinia tarvitaan esimerkiksi hormonien tuotantoon, keskushermoston toimintaan sekä verenkierron ja energiantuotannon ylläpitoon[5]. Niasiinipuutos aiheuttaa ihmisillä pellagra-puutostaudin, joka voi johtaa jopa kuolemaan[6].
Niasiinin biologiset reaktiot
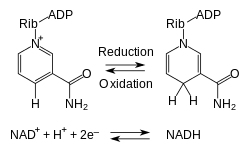
Niasiini muuntuu eliöissä NADH:ksi ja NADPH:ksi, jotka ovat kiinni tietyissä entsyymeissä niiden suorittamien reaktioiden osalta pakollisina "apumolekyyleinä" eli koentsyymeinä. Kuitenkin poly(ADP-riboosi)polymeraasi- ja ADP-ribosyylisyklaasi -entsyymien kohdalla NADH ei ole koentsyymi vaan näissä entsyymeissä reagoiva aine eli substraatti.[2] NADH- ja NADPH-riippuvaiset entsyymit suorittavat yli 200:aa eri reaktiotyyppiä. Koentsyymeinä pelkistyneet NADH ja NADPH tai näiden hapettuneet muodot NAD+ ja NADP+ muuntuvat toisikseen entsyymien hapetus-pelkistysreaktioissa. Reaktioissa pelkistyneeltä koentsyymiltä siirtyy hydridi (H–) entsyymissä reagoivalle aineelle ja samalla reagoivaan aineeseen liittyy liuoksesta protoni (H+). Edeltävä reaktiosarja voi tapahtua myös päinvastoin. NAD+ on NADH:ta yleisempi muoto eläinsoluissa. NAD+ on koentsyyminä tietyissä molekyylejä hajottavissa eli katabolisissa reaktioissa, joissa se muuntuu NADH:ksi. Reaktiot ovat osa reaktioreittejä, joissa muun muassa hiilihydraatteja, aminohappoja ja rasvahappoja hajotetaan pienemmiksi aineiksi, jolloin muodostuu energiaa eli ATP:tä. NADPH on NADP+:tä yleisempi muoto eläinsoluissa. NADPH on koentsyyminä tietyissä anabolisissa reaktioissa, joissa pienemmistä molekyyleistä tuotetaan suurempia. Reaktiot ovat osa esimerkiksi kolesterolin, rasvahappojen ja deoksiribonukleotidien tuottoa.[7]
Suositukset ja saanti
Niasiinin tarve riippuu muun muassa ihmisen iästä, sukupuolesta sekä fyysisen rasituksen määrästä. 20-39-vuotiaat raskasta työtä tekevät miehet tarvitsevat 23 milligrammaa niasiinia vuorokaudessa, kun taas kevyttä työtä tekeville 60-vuotiaille naisille riittää 9,6 milligrammaa.[8]
Suomen valtion ravitsemusneuvottelukunnan mukaan 6–65-vuotiaan tulisi saada niasiinia 13,4 milligrammaa niasiiniekvivalenttia (NE) vuorokaudessa, jos hänen energiatarpeensa on 2 000 kilokaloria vuorokaudessa.[9] Suosituksessa ei ole huomoitu sitä, että kasviperäinen niasiin imeytyy eläinperäistä heikommin, sillä niasiiniekvivalentilla tarkoitetaan ruoan sisältämän nikotiiniamidin ja nikotiinihapon yhteismäärää.[2]
Niasiinin vaihtoehtona voidaan nauttia sen esiasteena toimivaa tryptofaania, jota tarvitaan keskimäärin 60 milligrammaa yhden niasiinimilligramman syntetisoimiseen[4]. Tryptofaani muunnetaan niasiiniekvivalentiksi jakamalla se luvulla 60[2].
| Lapset | Miehet | Naiset | Raskaana
olevat | |||
|---|---|---|---|---|---|---|
| Ikä | mg | Ikä | mg | Ikä | mg | 17 mg |
| < 6 kk | – (a) | 10–13 v | 15 | 10–13 v | 14 | |
| 6–11 kk | 5 | 14–17 v | 19 | 14–17 v | 16 | |
| 12–23 kk | 7 | 18–30 v | 19 | 18–30 v | 15 | Imettävät |
| 2–5 v | 9 | 31–60 v | 18 | 31–60 v | 14 | 20 mg |
| 6–9 v | 12 | 61–74 v | 16 | 61–74 v | 13 | |
| ≥ 75 v | 15 | ≥ 75 v | 13 | |||
| (a): Äidinmaito tai äidinmaidonkorvike tyydyttää alle 6 kk ikäisten ravinnetarpeet | ||||||
Keskimääräinen niasiinin tarve tyydyttää ravinnetarpeen puolella väestöstä, mutta sitä hieman alhaisempi saanti ei silti välttämättä johda oireilevaan puutostilaan. Vuonna 2017 havaittiin, että 18–74-vuotiaat suomalaismiehet saivat niasiinia keskimäärin 39 milligrammaa vuorokaudessa ja 4,2 mg per ruuasta ravintoenergiana saatu megajoule (mg/MJ). Naisilla nämä arvot olivat 29 mg/vrk ja 4,1 mg/MJ. Kaikki tutkittavat saivat saantisuositusten mukaisen määrän niasiinia.[10] Vuonna 2013 julkaistussa ravitsemustieteen pro gradu opinnäytetyössä havaittiin kuitenkin, että kaikki tutkittavat 30-35-vuotiaat Helsingin seudulla asuvat saivat suositusta vähemmän niasiinia[11].
Niasiiniekvivalentti
Niasiinisuositukset ja ruokien niasiinipitoisuudet ilmoitetaan usein niasiiniekvivalentteina (NE), sillä ihmisillä ravintoproteiineista saadusta tryptofaanista noin 1/60-osa muuntuu kehossa NADH:ksi ja NADPH:ksi. NE on siksi summa ruuan niasiinin massasta ja 1/60-osa ruuan tryptofaanin massasta. Massat ovat usein milligrammoina (mg). Ihmisille NE on siis[2]
- NE (mg) = mg niasiinia + 1/60 mg tryptofaania
Suhde ei käytännössä ole vakio – jos tryptofaanisaanti on suuri ja niasiinisaanti pieni, on vitamiinin saanti usein pienempi. Jos niasiinin saanti on suuri ja tryptofaanin saanti pieni, on vitamiinin saanti usein suurempi. Myös muut ravinteet kuten aminohapot ja rasvahapot vaikuttavat imeytyvyyteen. Sairaudet voivat myös vaikuttaa. Lisäksi ihmisten tryptofaanimetabolian teho vaihtelee henkilöstä toiseen geneettisten erojen vuoksi.[2]
Muilla eläimillä niasiini:tryptofaani-suhde voi olla eri kuin 1:60. Esimerkiksi kananpojilla suhde on noin 1:45, rotilla 1:50 ja ankoilla 1:175.[7]
Liikasaanti
Kasviperäinen nikotiinihappo on eläinperäistä nikotiiniamidia myrkyllisempää. Suuret annokset nikotiinihappoa saattavat aiheuttaa muun muassa maksan toimintahäiriötä.
Nikotiiniamidi ei vaikuta veren kolesterolipitoisuuteen[7] eikä sen suuret annokset ole yhtä myrkyllisiä kuin nikotiinihapolla. Esimerkiksi noin 3 000 mg/vrk annokset nikotiiniamidia eivät ole aiheuttaneet ihmisillä merkittäviä haittavaikutuksia, mutta yli 9 000 mg/vrk annokset saattavat aiheuttaa maksan toimintahäiriöitä.[12]
Dyslipidemian hoito nikotiinihapolla
Suuret nikotiinihappoannokset alentavat veren kohonneita LDL-kolesterolipitoisuuksia, minkä vuoksi nikotiinihappoa käytetään joissain maissa dyslipidemian korjaamiseen.[13][12] Kyseisen hoidon tarkoituksena on ehkäistä valtimoahtaumatautia eli ateroskleroosia. Nikotiinihappo pienentää veren VLDL- ja LDL-kolesterolipitoisuuksia ja suurentaa veren HDL-kolesterolipitoisuuksia.[2] Suuret kerta-annokset nopeasti vapautuvaa nikotiinihappoa aiheuttavat yläruumiin hetkellistä punastumista ja muita sivuvaikutuksia. Pitkässä käytössä saattaa ilmetä maksan laajentumista (hepatomegaliaa) ja muita maksaongelmia.[2] Suuret kerta-annokset voivat myös aiheuttaa akuuttia maksan vajaatoimintaa.[14]
Saannin ylärajat
Suomen valtion ravitsemusneuvottelukunta on esittänyt 10 milligrammaa nikotiinihappoa per vuorokausi (mg/vrk) olevan turvallinen ravintolisistä saatu enimmäismäärä aikuisille. Yläraja nikotiiniamidille on 900 mg/vrk.[9] EFSA:n määrittämät turvalliset rajat ovat taulukossa alla.
| Ikä | 1–3 v | 4–6 v | 7–10 v | 11–14 v | 15–17 v | yli 18 v |
|---|---|---|---|---|---|---|
| Nikotiinihappo | 2 | 3 | 4 | 6 | 8 | 10 |
| Nikotiiniamidi | 150 | 220 | 350 | 500 | 700 | 900 |
Puutos
- Pääartikkeli: Pellagra
Niasiinin tai tryptofaanin liian vähäinen saanti ruuasta aiheuttaa pellagra-nimistä puutostautia, joka johtaa jatkuessaan kuolemaan. Ensioireina on symmetristä hilseilevää ja karheaa ihottumaa sekä ihon tummentumista (hyperpigmentaatio).[7] Ihottuma esiintyy auringonvalolle alttiilla alueilla kuten kämmenselissä, ja se voi olla kivuliasta ja muistuttaa auringon polttamaa. Lisäksi voi esiintyä ripulia. Pellagran keskushermostoperäisiä oireita ovat uupumus, psyykkinen tai emotionaalinen levottomuus, ahdistuneisuus, ärtyisyys ja masennus. Myöhemmin seuraa psykoottisia oireita, apatiaa, keskittymiskyvyttömyyttä ja muistihäiriöitä. Muita mahdollisia oireita ovat vapina, kävelyhäiriöt, ekstrapyramidaalinen jäykkyys, virtsa- ja ulosteinkontinenssi ja epileptiset kohtaukset. lopulta potilas kuolee.[6]
Interventiotutkimuksissa on havaittu, että pellagraa alkaa ilmetä, jos niasiinin saanti laskee keskimäärin 8,8 grammaan päivässä[3]. Pellagraa esiintyy etenkin kehitysmaissa[16], jossa se johtuu usein siitä, että vähälihainen ja -maitoinen ruokavalio perustuu maissiin tai valkoiseen riisiin[17]. Satatuhatta yhdysvaltalaista kuoli pellagraan 1900-luvun alussa, koska heidän ruokavaliossaan oli liian vähän lihaa ja maitoa[18].
Ruokapitoisuudet ja biologinen hyödynnettävyys
Kalkkuna, tonnikala ja naudanpaisti ovat erinomaisia niasiinin lähteitä. Myös maapahkinä ja kokojyvävilja olisivat hyviä niasiininlähteitä, mutta elimistö kykenee hyödyntämään vain pienen osan kasviperäisestä niasiinista.[19]
Valtaosa lihan niasiinista on entsyymeihin koentsyymeiksi sitoutuneena NADH:na ja NADPH:na, jotka imeytyvät verrattain helposti ihmiskehon käyttöön. Osa koentsyymeistä hajoaa kypsennyksen yhteydessä lisäksi helposti imeytyväksi nikotiiniamidiksi. Liha sisältää myös kasviravintoa runsaammin niasiinin esiastetta tryptofaania.[7]
Viljoissa ja maississa esiintyy lähinnä nikotiinihappoa, joka imeytyy huonosti ihmiskehoon, koska se on esteröitynyt polysakkarideihin ja glykopeptideihin. Esimerkiksi kypsän maissin niasiinin hyötyosuus on enintään noin 35 prosenttia, vaikka maissia olisi kuumennettu ruuanlaitossa. Niasiinia lisätään tämän vuoksi viljatuotteisiin lain vaatimana Yhdysvalloissa ja joissain muissa maissa. Joissain Väli-Amerikan kulttuureissa perinteisesti käytetty maissin emäskäsittely poltetulla kalkilla (eng. nixtamalization) vapauttaa nikotiinihappoa esterisidoksia hydrolysoimalla, mikä lisää merkittävästi sen hyödynnettävyyttä. Joissain kasveissa niasiinia on paljon trigonelliininä eli 1-metyylinikotiinihappona. Tämä on kasvihormoni, joka ei toimi vitamiinina eläimillä, mutta muuntuu vitamiinitoimintoiseksi nikotiinihapoksi kuumennettaessa ainetta kylliksi.[7]
| Heinäkasvit ja jauhot | Pavut ja pähkinät | Lihat | |||
|---|---|---|---|---|---|
| Kaurahiutale | 0,961 | Cashewpähkinä, kuivapaahdettu | 1,4 | Ankka, paahdettu | 5,1 |
| Maissijauho, täysjyvä | 1,9 | Härkäpapu, keitetty | 0,711 | Kalkkuna, iholla, paahdettu | 5,086 |
| Ohraryyni | 2,063 | Kidneypapu, keitetty | 0,578 | Kana, iholla, paahdettu | 11,13 |
| Pasta, täysjyvä, keitetty | 0,707 | Kikherne, keitetty | 0,526 | Kirjolohi (viljelemätön), paistettu | 5,77 |
| Riisi, tumma, keitetty | 1,528 | Linssi, keitetty | 1,06 | Makrilli, paistettu, kuivattu | 6,85 |
| Riisi, valkoinen, keitetty | 0,29 | Maapähkinä, kuivapaahdettu | 13,53 | Nauta (vasikka), maksa, käristetty[21] | 16,50 |
| Ruisjauho, puolikarkea | 1,727 | Manteli, kuivapaahdettu | 2,817 | Nauta, kylki, ¼ rasvaa, pariloitu | 3,25 |
| Soijajauho, rasvainen | 3,286 | Parapähkinä, kuivapaahdettu | 1,622 | Nauta, munuainen, pariloitu[21] | 10,70 |
| Vehnäjauho, täysjyvä | 6,365 | Pekaanipähkinä, kuivapaahdettu | 0,922 | Sika, jalka, vähärasvainen, paahdettu | 4,935 |
| Vehnälese | 13,58 | Pistaasi, kuivapaahdettu | 1,408 | Silli, säilyke | 3,3 |
| Vehnänalkio | 6,813 | Voipapu, keitetty | 0,096 | Turska, paistettu | 2,513 |
| Kasvikset ja ruokasienet | Hedelmät ja marjat | Lehmänmaitotuotteet | |||
| Bataatti, kuorimaton, uunipaistettu | 0,604 | Aitoviikuna | 0,4 | Briejuusto | 0,38 |
| Herne | 2,09 | Ananas | 0,42 | Camembertjuusto | 0,63 |
| Jääsalaatti | 0,187 | Appelsiini | 0,282 | Cheddarjuusto | 0,08 |
| Keltasipuli | 0,148 | Aprikoosi | 0,6 | Edamjuusto | 0,082 |
| Keräkaali, keitetty | 0,282 | Avokado | 1,921 | Maito, rasvaton | 0,088 |
| Kukkakaali, keitetty | 0,41 | Banaani | 0,54 | Raejuusto, 1 % rasvaa | 0,128 |
| Kurkku | 0,221 | Hunajameloni | 0,574 | Sinihomejuusto | 1,016 |
| Lanttu | 0,715 | Karviainen | 0,3 | Voi | 0,042 |
| Maissi, keltainen | 1,7 | Luumu | 0,5 | Kerma, 31,3 % rasvaa[21] | 0,04 |
| Parsa, keitetty | 1,082 | Mango | 0,584 | Herajauhe, makea[21] | 1,26 |
| Parsakaali, keitetty | 0,574 | Mansikka | 0,23 | Jugurtti, 3,3 % rasvaa[21] | 0,11 |
| Peruna, kuorimaton, uunipaistettu | 1,395 | Mustaherukka | 0,3 | Muut | |
| Pinaatti | 0,724 | Mustikka | 0,359 | Kananmuna, keitetty | 0,064 |
| Porkkana | 0,928 | Omena | 0,077 | Rintamaito (ihmisen) | 0,177 |
| Punajuuri, keitetty | 0,331 | Persikka | 0,99 | Rypsiöljy | 0 |
| Selleri | 0,323 | Päärynä | 0,1 | Oliiviöljy | 0 |
| Siitake, kuivattu | 14,1 | Vadelma | 0,9 | Olut, lager | 0,453 |
| Tomaatti | 0,628 | Viinirypäle | 0,25 | Leivinhiiva, tuore (puristehiiva)[21] | 11,20 |
| a: ruuat ovat raakoja eli valmistamattomia ellei toisin mainita. Pitoisuudet ovat keskimääräisiä.
b: lähteissä ei ole määritetty millaisena niasiinimuotona pitoisuuksien massat (mg) ovat. | |||||
Kemia
Nikotiinihappo
- Pääartikkeli: Nikotiinihappo#Kemia
Nikotiiniamidi
- Pääartikkeli: Nikotiiniamidi#Kemia
Nikotiiniamidiribosidi

Nikotiiniamidiribosidi (NR, CAS-numero 1341-23-7) on nikotiiniamidi, jonka pyridiinitypessä on kiinni riboosi. Se on rakenteeltaan sama kuin nikotiiniamidimononukleotidi (NMN), mutta fosforyloimaton. NR on niasiinivitamiini, sillä se muuntuu nikotiiniamidiribosidikinaasi 1:llä (NRK1, EC-numero 2.7.1.22) nikotiiniamidimononukleotidiksi. Tämä muuntuu NMN-adenyylitransferaasi 1:llä (NMAT1, EC 2.7.7.1) NAD+:ksi. NR:ää on luontaisesti esimerkiksi maidossa.[22][23] NR ei aiheuta punastumista esimerkiksi 2 000 mg/vrk annoksin, eivätkä tämän kokoluokan annokset tiettävästi ole vaarallisia.[24]
Inositoliheksanikotinaatti

Inositoliheksanikotinaatti (CAS 6556-11-2) eli inositolinikotinaatti on inositoli, jonka hydroksyyliryhmiin on esteröity nikotiinihappoa. Aine on niasiinivitamiini, sillä sen esterisidokset voivat hydrolysoitua ruuansulatuselimistössä vapauttaen nikotiinihappoa, joka imeytyy sitten normaalisti. Hydrolyysin teho vaihtelee suuresti ja vapautuvan nikotiinihapon hyötyosuus on keskimäärin noin 70 %. Hydrolyysi on hidasta: veren nikotiinihappopitoisuudet ovat veressä suurimmat 6–10 tunnin kuluttua aineen syönnistä. Vapaata nikotiinihappoa syötäessä veripitoisuus on suurin 0,5–1 tunnin kuluttua aineen syönnistä. Inositolinikotinaatti ei suurin annoksin hitaan vapautumisensa takia aiheuta muun muassa punastumista toisin kuin vapaa nikotiinihappo vastaavan kokolouokan annoksin.[25]
Valmistus
Nikotiinihappoa ja -amidia valmistetaan kemiallisella synteesillä. Lähtöaineina käytetään esimerkiksi 3-metyylipyridiiniä ja 5-etyyli-2-metyylipyridiiniä. Kuitenkin alla kuvattuihin reitteihin, joissa muodostuu välituotteena 3-syanopyridiiniä, saatetaan joissain tehtaissa yhdistää mikrobien entsyymien katalysoimia reaktioita. 3-syanopyridiini voidaan esimerkiksi hydrolysoida nitrilaasilla (EC-numero 3.5.5.1) nikotiinihapoksi tai hydratoida nitriilihydrataasilla (EC 4.2.1.84) nikotiiniamidiksi.[26]
3-metyylipyridiiniä tehdään reagoimalla kaasumaista asetaldehydiä, formaldehydiä ja ammoniakkia katalyyttisesti keskenään. Päätuote on pyridiini, mutta reaktioseoksesta 30–50 % on 3-metyylipyridiiniä. Toinen vaihtoehto on adiponitriilin valmistuksessa sivutuotteena muodostuvan 2-metyyliglutaronitriilin käyttö. Tämän syanoryhmät voidaan pelkistää – saadaan 2-metyyli-1,5-diaminopentaania. Tämä avoketjuisen aineen aminoryhmät kondensoidaan, jolloin saadaan rengasrakenteinen 3-metyylipiperidiini. Piperidiinirengas dehydrataan pyridiiniksi ja saadaan 3-metyylipyridiiniä. Edeltävin tavoin saatua 3-metyylipyridiiniä voidaan reagoida ammoniakin ja vedyn kanssa noin 350 °C:ssa ja korkeassa paineessa 3-syanopyridiiniksi. Reaktiossa käytetään antimonin, vanadiinin tai titaaniin oksidien heterogeenisiä katalyyttejä, jolloin 3-syanopyridiinin saanto voi olla jopa 96 %. Tuote hydrolysoidaan 130–150 °C:ssa nikotiiniamidiksi katalyyttisellä määrällä NaOH:ia tai muuta emästä.[27]
Reagoimalla esimerkiksi paraldehydiä ja ammoniakkia keskenään saadaan 5-etyyli-2-metyylipyridiiniä. Tämä hapetetaan vesiliuoksessa yksittäisessä reaktioastiassa nikotiinihapoksi käyttäen ylimäärää typpihappoa. Reaktio tapahtuu 230–270 °C:ssa 6–8 MPa paineessa. Välituotteena muodostuu epävakaata pyridiini-2,5-dikarboksyylihappoa, joka dekarboksyloituu nikotiinihapon nitraattisuolaksi. Suola neutraloidaan nikotiinihapoksi – tässä muodostuva typpioksidi hapetetaan ilmalla typpidioksidiksi. Tämä absorboidaan veteen ja saadaan typpihappoa, joka käytetään jälleen prosessissa.[27]
Toiminta eliöissä
Hapetus-pelkistysreaktiot
Niasiini muuntuu ihmisissä ja muissa eliöissä NADH:ksi ja NADPH:ksi. Nämä sitoutuvat tiettyihin oksidoreduktaasi-entsyymeihin koentsyymien roolissa. Entsyymit katalysoivat hapetus-pelkistysreaktioita, joiden yleisreaktio on[28]
- AH2 + NAD(P)+ ⇌ A + NAD(P)H + H+
NAD+ tai NADP+ ottaa hydridin (H–) entsyymillä reagoivalta aineelta A hapettaen sen. A:lta poistuu hydridinä 2 elektronia (2 e–). A:lta irtoaa reaktiossa myös protoni (H+), joka vapautuu entsyymiä ympäröivään liuokseen yhtyen esimerkiksi veteen (H2O) oksoniumioniksi (H3O+). Reaktio voi olla päinvastainen, jolloin NADH tai NADPH luovuttaa hydridin A:lle pelkistäen sen – hydridin mukana A saa 2 elektronia.[28]
Hydridin liitos on stereospesifinen reaktio sillä se voi kiinnittyä 2:lta eri puolelta NAD(P)+:hen. Tällä luokitellaan niasiinientsyymeitä niiden reaktiomekanismin mukaan A-tyypin eli pro-R-spesifisiin ja B-tyypin eli pro-S-spesifisiin (kuva), joita on likimain yhtä paljon.[29]
Eläimillä NAD+ on lähinnä osa ravintoaineiden hapettavia hajotusreaktioita, joissa se muuntuu NADH:ksi. Näitä reaktioita ovat muun muassa[7]
- glykolyysi
- pyruvaatin oksidatiivinen dekarboksylaatio
- asetaatin hapetus sitruunahappokierrossa
- etanolin hapetus
- rasvahappojen β-oksidaatio
Edeltävissä reaktioissa muodostuvat NADH:t siirtävät ravinteilta saamiaan elektroneja (hydrideitä) pääosin mitokondrioiden elektroninsiirtoketjuun hapettuen itse samalla takaisin NAD+:ksi. Ketju on osa solujen muiden entsyymien käyttämän kemiallisen energian eli ATP:n tuottoa oksidatiivisessa fosforylaatiossa.[2]

Eläimillä NADPH:ta muodostuu NADP+:stä pentoosifosfaattireitillä. NADPH osallistuu sitten pääosin pelkistäviin biosynteesireaktioihin (erotuksena NADH:n hajotusreaktiohin). NADPH on osa kolesterolin, rasvahappojen, deoksiribonukleotidien ja monien muiden molekyylien tuottoa sekä glutationidisulfidin pelkistystä jälleen glutationiksi.[2]
Tietyt entsyymit kuten glutamaattidehydrogenaasi voivat käyttää koentsyyminä NADH:ta ja NADPH:ta.[31] Useimpien entsyymien kohdalla kuitenkin NADPH:n fosfaattiryhmä tekee siitä sopivan vain tiettyihin entsyymeihin, joihin NADH taas ei sovi. Tämä sallii soluja rajaamaan hapetus- ja pelkistysreaktioreittien säätelyn ja ylläpidon toisistaan. Valtaosa solujen NADPH:sta on tässä muodossa NADP+:n sijaan. Taas NAD+ on pääosin tässä muodossa NADH:n sijaan.[2]
Muut roolit
Poly(ADP-riboosi)polymeraaseihin kuuluvissa entsyymeissä NAD+ ei toimi koentsyyminä vaan substraattina. Polymeraasit liittävät tiettyihin proteiineihin translaation jälkeisesti NAD+:n ADP-ribooseja ketjuiksi. Tätä kutsutaan ADP-ribosylaatioksi. Kussakin liitoksessa nikotiiniamidi lohkeaa vapaaksi. Polymeraasit osallistuvat epäsuorasti muun muassa DNA:n korjaukseen, solujen erikoistumiseen, solujakautumiseen ja apoptoosiin.[7]

NAD+ on syklisen ADP-riboosin (cADPR) esiaste, jota ADP-ribosyylisyklaasit tuottavat. cADPR on toisiolähetti, joka vapauttaa soluvarastoista kalsiumkationeja (Ca2+) solulimaan sitoutumalla solulimakalvoston ja sarkoplasmakalvoston ryanodiinireseptoreihin.[32]
Entsyymeitä
| Rooli | NAD(H) | NADP(H) | ||
|---|---|---|---|---|
| Nimi | EC | EC | ||
| Hiilihydraattien
metabolia |
Glyseraldehydi-3-fosfaattidehydrogenaasi | 1.2.1.12 | Glukoosi-6-fosfaattidehydrogenaasi | 1.1.1.49 |
| Laktaattidehydrogenaasi | 1.1.1.27 | 6-fosfoglukonaattidehydrogenaasi | 1.1.1.44 | |
| Alkoholidehydrogenaasi | 1.1.1.1 | |||
| Lipidien
metabolia |
Glyseroli-3-fosfaattidehydrogenaasi | 1.1.1.8 | 3-ketoasyyli-ACP-reduktaasi | 1.1.1.100 |
| 3-hydroksiasyyli-CoA-dehydrogenaasi | 1.1.1.35 | Enoyyli-ACP-reduktaasi | 1.3.1.9 | |
| HMG-CoA-reduktaasi | 1.1.1.34 | |||
| Aminohappojen
metabolia |
Glutamaattidehydrogenaasi | 1.4.1.2 | Glutamaattidehydrogenaasi | 1.4.1.4 |
| Muu | NADH-dehydrogenaasi | 1.6.99.3 | Glutationireduktaasi | 1.8.1.7 |
| Poly(ADP-riboosi)polymeraasi | 2.4.2.30 | Dihydrofolaattireduktaasi | 1.5.1.3 | |
| ADP-ribosyylisyklaasi[32] | 3.2.2.6 | Tioredoksiinireduktaasi | 1.8.1.9 | |
| 4-hydroksibentsoaatti-3-mono-oksigenaasi | 1.14.13.2 | |||
| NADPH-hemoproteiinireduktaasi | 1.6.2.4 | |||
Muu biokemia
Imeytyminen
Osa niasiinista irtoaa ravintoproteiineista vapaaksi NADH:ksi tai NADH:ksi ruuanlaiton aikana ja osa entsymaattisesti ruuansulatuselimistössä. Kasvien niasiiniglykosideista ja -peptideistä niasiinin vapautumista tapahtuu lähinnä ruuanlaiton aikana, muttei elimistössä.[33] Vapaa NADH ja NADPH pilkkoutuvat NAD(P)+-nukleosidaasilla (EC-numero 3.2.2.6) nikotiiniamidiksi ja ADP-riboosiksi. NADH:n pyrofosfaattisidos voi myös katketa pyrofosfataaseilla, jolloin muodostuu nikotiiniamidimononukleotidi (NMN) ja AMP. NMN defosforyloituu nikotiiniamidiribosidiksi. Nikotiiniamidiribosidia muodostuu myös jos NAD+ hajoaa fosfodiesteraaseilla, jolloin myös ADP:tä muodostuu. Nikotiiniamidiribosidista voi hydrolyysillä tai fosforylaatiolla vapautua nikotiiniamidi, jolloin muodostuu sivutuotteena vastaavasti riboosi tai riboosi-1-fosfaatti.[7]
Vapaa nikotiinihappo tai -amidi voi imeytyä mahalaukusta, mutta imeytyminen on nopeampaa ohutsuolessa.[2] Niasiinin imeytymismekanismi on osin vielä tuntematon. Imeytyminen ohutsuolisoluihin tapahtuu normaalipitoisuuksissa tiettävästi protonivälitteisen orgaanisten anionien kuljettajaproteiini 10[33] (lyhenne OAT10, geeni SLC22A13)[34] kautta, joka vastaa myös virtsahapon ja aminohippurihapon imeytymisestä. Suurissa niasiinipitoisuuksissa kuljetukseen osallistuu myös natriumvälitteinen monokarboksylaattien kuljettajaproteiini 1 (SMCT1, SLC5A8), joka vastaa myös maitohapon, pyruvaatin ja lyhyiden rasvahappojen imeytymisestä.[33]
Ohutsuolisoluihin päätynyt nikotiiniamidi voi muuntua NAD+:ksi tai päätyä porttilaskimon kautta verenkiertoon. Nikotiinihaposta osa vapautuu vereen sellaisenaan, mutta valtaosa muuntuu suolisoluissa entsymaattisesti NAD+:ksi, jota voidaan tarpeen mukaan hajottaa verenkiertoon vapautuvaksi nikotiiniamidiksi.[2]
Kuljetus
Porttilaskimossa ohutsuolesta imeytynyt nikotiinihappo- ja amidi päätyvät joko maksaan käsiteltäväksi tai sitten punasoluihin. Punasoluissa ne muuntuvat nikotiiniamidin nukleotideiksi.[2] Verenkierron vapaasta niasiinista pääosa on nikotiiniamidia ja pienempi osa nikotiinihappoa.[7]
Varastoituminen
Niasiini on ihmisillä kudoksissa pääosin hapettuneena NAD+:nä ja pelkistyneenä NADPH:na, jotka ovat entsyymeissä kiinni koentsyymeinä. Näitä entsyymeitä on jossain määrin enemmän maksassa kuin monissa muissa kehon osissa.[13] NAD+:tä on kehossa paljon enemmän kuin NADPH:ta.[7] Niasiinivarannot riittävät ihmisillä noin 2–6 viikkoa niasiinin (tai tryptofaanin) saannin loppumisesta.[35]
Hajotus ja erittyminen
NAD(P)+-nukleosidaasi (EC-numero 3.2.2.6) hajottaa NAD+:tä ja NADP+:tä nikotiiniamidiksi. Tämä voi muuntua taas NAD+:ksi tai metyloitua nikotiiniamidi-N-metyylitransferaasilla (EC 2.1.1.1) 1-metyylinikotiiniamidiksi. Metylaatio tapahtuu pääosin maksassa. Tämä erittyy virtsaan sellaisenaan tai hapettuu erinäisiksi metaboliiteiksi, joista ihmisillä pääasiallinen on 1-metyyli-6-pyridoni-3-karboksiamidi (CAS 701-44-0).[7]
Muita nisäkkäiden vähäisempiä virtsametaboliitteja ovat muun muassa nikotiiniamidi-1-oksidi, 1-metyyli-4-pyridoni-3-karboksiamidi, 1-metyyli-6-pyridoni-3-karboksiamidi, 6-hydroksinikotiiniamidi ja 6-hydroksinikotiinihappo. Saannin ollessa suurta, yli 65 % syödystä niasiinista erittyy sellaisenaan (nikotiinihappona tai -amidina).[7]
Tryptofaanin muunnos niasiiniksi

Nisäkkäillä tryptofaani muuntuu N-L-formyylikynureniiniksi maksassa tryptofaani-2,3-dioksigenaasilla (EC-numero 1.13.11.11) tai muissa kudoksissa kuin maksassa indoliamiini-2,3-dioksigenaasilla (EC 1.13.11.52). Aryyliformamidaasi (EC 3.5.1.9) hajottaa N-formyylikynureenin kynureniiniksi. Kynureeni muuntuu sitten kolmivaiheisesti 2-amino-3-karboksimukoni-6-semialdehydiksi (ACMS, CAS-numero 16597-58-3). Kolmea vaihetta katalysoivat järjestyksessä kynureniini-3-mono-oksigenaasi (EC 1.14.13.9), kynureninaasi (EC 3.7.1.3) ja 3-hydroksiantranilaatti-3,4-dioksigenaasi (EC 1.13.11.6). Avonainen ACMS kondensoituu spontaanisti rengasrakenteiseksi kinoliinihapoksi. Kinoliinihappo muuntuu nikotinaattinukleotididifosforylaasilla (EC 2.4.2.19) nikotiiniamidimononukleotidiksi (NMN).[33] Reaktio tapahtuu nisäkkäillä pääosin munuaisissa ja maksassa, sillä fosforylaasia on eniten näissä kudoksissa. NMN muuntuu NAD+ syntaasilla (EC 6.3.1.5) NAD+:ksi.[2]
Tämän reitin entsyymit vaativat toimintaansa B6-vitamiinia, riboflaviinia (B2) rautaa ja hemiä kofaktorien roolissa. Näiden puutos hidastaa siksi synteesireitin toimintaa.[33]
Antagonistit

Niasiinilla on antagonisteja, jotka estävät NADH:n ja/tai NADPH:n toimintaa tietyissä entsyymeissä. Näistä monet ovat pyridiinejä. Antagonisteja useimmille eliöille ovat muun muassa 3-asetyylipyridiini (CAS 350-03-8), isoniatsidi, pyridiini-3-sulfonihappo (CAS 636-73-7) ja 6-aminonikotiiniamidi (CAS 329-89-5).[36]
Historia ja nimet
Niasiinipuutoksesta johtuvaa pellagraa ei tiettävästi tunnettu Euroopassa ennen maissin tuontia Amerikasta ja sen kasvatuksen aloittamista myös Euroopassa.[37]
1735 Gaspar Casál Juliánin (elänyt 1680–1759) kirjaamat havainnot Espanjan Asturiassa ovat varhaisimpia lääketieteellisiä kuvauksia pellagrasta. Casál kuvaili sairailla olevan muun muassa ihottumaa auringonvalolle altistuneilla ihoalueilla. Hänen mukaansa paikalliset kutsuivat tilaa espanjaksi nimellä mal de la rosa eli ruusutauti. Casál myös havaitsi pellagran liittyvän köyhyyteen ja ruokavalioon, jossa ei juuri ollut eläinperäisiä ruokia. Casálin pellagraa koskevat kirjoitukset julkaistiin vasta 1762 hänen kuolemansa jälkeen.[38][37] Casálin kuvausten jälkeen pellagra alkoi yleistymään Euroopassa, eritoten Espanjassa ja Italiassa.[37] Pellagrassa ilmenevää kaulaa ympäröivää ihottumaa on jälkikäteen kutsuttu myös englanniksi nimellä Casal collar eli Casalin kaulus.[7]
1771 Franciscus Frapolli havaitsi Italiassa pellagran liittyvän ruokavalioon, joka pohjautui pääosin maissipohjaiseen polenta-ruokalajiin. Frapollin mukaan tilaa kutsuttiin paikallisesti nimellä pelle agra, joka on italiaksi karkea iho. Tästä myös sana pellagra juontuu.[39][37]
1867 Carl Huber valmisti nikotiinihappoa selvittäen sen empiirisen kaavan.[40][41] Vasta 1870 hän tunnisti aineen olevan pyridiini.[42][41] 1873 H. Weidel nimesi aineen nikotiinihapoksi valmistaen sitä nikotiinia hapettamalla.[43][41] Aineen toiminta pellagraa estävänä vitamiinina tunnistettiin kuitenkin vasta 1937.[37]
USA:ssa pellagra yleistyi myöhemmin kuin Euroopassa. Se yleistyi Yhdysvaltain sisällissodan jälkeen ja pääosin eteläisissä osavaltioissa. 1900-luvun alussa pellagra oli yleistynyt USA:ssa merkittävästi. Tuolloin oli vielä kiistanalaista se, että oliko pellagra tarttuva sairaus vai aiheutuiko se myrkkyjen, ravinnepuutosten vai perinnöllisten syiden takia.[37]

Pellagran yleistymisen takia 1914 Yhdysvaltain julkisen terveyden palvelulaitos (USPHS) palkkasi Joseph Goldbergerin (1874–1929) tutkimaan asiaa.[37] 1914 Goldberger päätteli muiden asiaa tutkineiden kirjoitusten pohjalta pellagran johtuvan yksipuolisesta ruokavaliosta ja suositteli vähentämään maissin, viljojen ja tölkitetyn ruuan syöntiä ja lisäämään tuoretta lihaa, maitoa ja munia ruokavalioon.[44] Todistaakseen ruokateoriansa 1915–1923 Goldberger ja kollegat suorittivat kokeita kahdessa orpokodissa ja eräässä mielisairaalassa, joissa oli pellagraepidemia. He osoittivat yksipuolisen ja maissipainotteisen ruokavalion monipuolistamisen tuoreella lihalla, maidolla ja munilla parantavan pellagran ja estävän uusien pellagratapauksien ilmenemisen.[45][46] 1915 Goldberger myös osoitti terveillä vangeilla monipuolisen ruuan korvaamisen pääosin maissilla saavan vangeilla aikaan pellagran.[47] Näyttääkseen ettei pellagra ole tarttuva sairaus, 1916 Golberger otti verta pellagrapotilailta ja pisti tätä terveiden verenkiertoon. Terveet eivät sairastuneet. Altistus muillekaan sairaiden eritteille ei aiheuttanut terveillä pellagraa.[48] 1920-luvulla Goldberger ja kollegat selvittivät ettei tuolloin tunnettujen vitamiinien tai muiden ravinteiden annostelu estänyt pellagraa. Siksi 1925 he esittivät olevan olemassa tuntematon pellagraa ehkäisevä tekijä tai joukko tekijöitä, joita he kutsuivat PP-tekijäksi tai -tekijöiksi (eng. pellagra-preventing factor).[49] 1922–1928 Goldberger myös näytti koirakokeissa ns. mustakielitaudin (eng. black tongue) olevan koirien vastine pellagralle. 1926 Goldberger havaitsi leivinhiivan syötön myös estävän koirien pellagraa. Hiivassa tiedettiin tuolloin olevan "antineuriittista vitamiinia", jota nykyään kutsutaan tiamiiniksi tai B1-vitamiiniksi. Siksi Goldberger arveli PP-tekijän olevan B-vitamiini.[50][51] 1928 Goldberger ja kollegat tunnistivat muun muassa sian maksassa olevan PP-tekijää.[52] Golberger ei tehnyt jatkotutkimuksia PP-tekijän tunnistamiseksi, sillä hän kuoli syöpään 1929.[51]

1937 Conrad Arnold Elvehjem (1901–1962) ja kollegat eristivät nikotiiniamidia maksasta ja selvittivät eristämänsä aineen rakenteen. He havaitsivat nikotiiniamidin ja myös aiemmin syntetisoidun nikotiinihapon olevan PP-tekijöitä eli vitamiineja, joiden puutos aiheutti koirilla pellagraa.[53] Pian tätä löytöä seuranneet monet muut jatkotutkimukset ihmisillä ja muilla eläimillä varmensivat näiden aineiden olevan vitamiineja.[51]
1939 C. A. Elvehjelm osoitti niasiinin toimivan NADH:n ja NADPH:n esiasteena.[54]

1942 C. A. Elvehjem, W. H. Sebrell ja Tom Douglas Spies nimesivät nikotiinihapon ja -amidin vastaavasti niasiiniksi ja niasiiniamidiksi. Niasiinin englanninkielinen sana niacin on yhdistelmä sanoista nicotinic acid ja vitamin. Uudet nimet keksittiin, jottei suurempi yleisö väärinymmärtäisi, että aineilla olisi jokin yhteys tupakan tai hyönteismyrkyissäkin käytettyyn nikotiiniin tai syövyttäviin happoihin.[55][56]
1940-luvulla ilmeni etteivät niasiiniköyhät ruokavaliot aina johtaneet pellagraan. Esimerkiksi maidoissa nikotiinihappoa tai -amidia on vähän. Silti maidon lisääminen ruokavalioon esti pellagraa.[51] Ristiriita selvisi 1945, kun rottien havaittiin voivan muodostaa tryptofaanista niasiinia.[57][58] Useissa jatkotutkimuksissa tryptofaania pistettiin koe-eläinten verenkiertoon ja käytettiin myös isotooppileimattua tryptofaania. Nämä kokeet osoittivat niasiinisynteesin tapahtuvan suoraan useiden eläinten soluissa niiden suolimikrobien sijaan. Myöhemmin selvisi myös solujen biokemiallinen reitti, jossa tryptofaani muuntuu NADH:ksi.[58]
Termin B3-vitamiini käyttö synonyyminä niasiinille alkoi yleistyä vasta 1970-luvun alussa osin William Griffith Wilsonin 1968 julkaiseman "The Vitamin B-3 Therapy"-kirjan takia.[59][60]
Katso myös
- FAD eli FADH
Lähteet
- GF Combs et al: The vitamins: fundamental aspects in nutrition and health. 3. painos. Elsevier Academic Press, 2008. ISBN 9780121834937.
Viitteet
- Ravintotekijä - Fineli fineli.fi. Viitattu 22.5.2019.
- J Zempleni et al: Handbook of vitamins, s. 192–223. 4. painos. Taylor & Francis, 2007. ISBN 9780849340222. (englanniksi)
- Elintarvikkeiden täydentäminen tiamiinilla, niasiinilla, foolihapolla, pyridoksiinilla ja B12-vitamiinilla Riskinarviointi työikäisessä väestössä. Kansanterveyslaitoksen julkaisuja 2005. https://www.julkari.fi/bitstream/handle/10024/78489/2005b25.pdf?sequence=1&isAllowed=y
- Elintarvikkeiden täydentäminen tiamiinilla, niasiinilla, foolihapolla, pyridoksiinilla ja B12-vitamiinilla Riskinarviointi työikäisessä väestössä. Kansanterveyslaitoksen julkaisuja 2005. https://www.julkari.fi/bitstream/handle/10024/78489/2005b25.pdf?sequence=1&isAllowed=y
- Vitamin B-3 and Beans Healthy Eating | SF Gate. Viitattu 16.3.2023. (englanniksi)
- Vitamiininpuutosten aiheuttamat enkefalopatiat www.duodecimlehti.fi. Viitattu 16.3.2023.
- Combs, s. 296–310
- Osmo Turpeinen: Ruoka-ainetaulukko. 29.-30. painos. Otava 1993.
- Suomalaiset ravitsemussuositukset 2014, s. 49, 51. 5. painos. Valtion ravitsemusneuvottelukunta, 2018. ISBN 9789524538015. Teoksen verkkoversio.
- L Valsta et al: Ravitsemus Suomessa - FinRavinto 2017 -tutkimus, s. 61, 102. Terveyden ja hyvinvoinnin laitos, 2018. ISBN 9789523432383. Teoksen verkkoversio.
- Hanna Kivimäki: POIKKILEIKKAUSTUTKIMUS VEGAANIEN RUOANKÄYTÖSTÄ JA RAVINTOAINEIDEN SAANNISTA. Pro Gradu -tutkielma, ravitsemustiede, lääketieteen laitos, terveystieteiden tiedekunta, Itä-Suomen yliopisto. https://erepo.uef.fi/bitstream/handle/123456789/11921/urn_nbn_fi_uef-20130281.pdf?sequence=-1&isAllowed=y
- M Knip et al: Safety of high-dose nicotinamide: a review. Diabetologia, marraskuu 2000, 43. vsk, nro 11, s. 1337–1345. PubMed:11126400. doi:10.1007/s001250051536. ISSN 0012-186X. Artikkelin verkkoversio. (englanniksi)
- B Caballero et al: Encyclopedia of human nutrition, s. 182–188. Academic Press, 2013. ISBN 9780123848857. Teoksen verkkoversio. (englanniksi)
- MA Ellsworth et al: Acute liver failure secondary to niacin toxicity. Case Reports in Pediatrics, 2014, 2014. vsk. PubMed:24711953. doi:10.1155/2014/692530. ISSN 2090-6803. Artikkelin verkkoversio. (englanniksi)
- Overview on tolerable upper intake levels as derived by the Scientific Committee on Food (SCF) and the EFSA panel on Dietetic Products, Nutrition and Allergies (NDA) (pdf) EFSA. 2018. Viitattu 6.3.2019. (englanniksi)
- J Zempleni et al: Handbook of vitamins, s. 192–223. 4. painos. Taylor & Francis, 2007. ISBN 9780849340222.
- B Caballero et al: ”Volume 4”, Encyclopedia of human nutrition, s. 183–188. 3. painos. Elsevier, 2013. ISBN 9780123750839.
- Mari Heikkilä: Ällöttävä testi paljasti ravinnepuutoksen. Tiede-lehti 3/2023, sivu 50.
- Elintarvikkeiden täydentäminen tiamiinilla, niasiinilla, foolihapolla, pyridoksiinilla ja B12-vitamiinilla Riskinarviointi työikäisessä väestössä. Kansanterveyslaitoksen julkaisuja 2005. https://www.julkari.fi/bitstream/handle/10024/78489/2005b25.pdf?sequence=1&isAllowed=y
- Combs, s. 528–560
- ME Ensminger et al: The concise encyclopedia of foods & nutrition, s. Ensminger, "Chapter 6 F". CRC Press, 1995. ISBN 9780849344558. (englanniksi)
- RS Fletcher, GG Lavery: The emergence of the nicotinamide riboside kinases in the regulation of NAD+ metabolism. Journal of Molecular Endocrinology, 30.5.2018, 61. vsk, nro 3, s. R107–R121. PubMed:30307159. doi:10.1530/JME-18-0085. ISSN 0952-5041. Artikkelin verkkoversio. (englanniksi)
- C Cantó et al: NRK1 controls nicotinamide mononucleotide and nicotinamide riboside metabolism in mammalian cells. Nature Communications, 11.10.2016, 11. vsk, nro 7, s. 13103. PubMed:27725675. doi:10.1038/ncomms13103. ISSN 2041-1723. Artikkelin verkkoversio. (englanniksi)
- CR Martens et al: Chronic nicotinamide riboside supplementation is well-tolerated and elevates NAD+ in healthy middle-aged and older adults. Nature Communications, 29.3.2018, 9. vsk, nro 1. PubMed:29599478. doi:10.1038/s41467-018-03421-7. ISSN 2041-1723. Artikkelin verkkoversio. (englanniksi)
- Inositol hexanicotinate (inositol hexaniacinate) as a source of niacin (vitamin B3) added for nutritional purposes in food supplements. EFSA Journal, 2009, 7. vsk, nro 2, s. 949. doi:10.2903/j.efsa.2009.949. ISSN 1831-4732. Artikkelin verkkoversio. (englanniksi)
- CG Acevedo-Rocha et al: Microbial cell factories for the sustainable manufacturing of B vitamins. Current Opinion in Biotechnology, 2019, 56. vsk, s. 18–29. PubMed:30138794. doi:10.1016/j.copbio.2018.07.006. Artikkelin verkkoversio. (englanniksi)
- M Eggersdorfer et al: ”Vitamins”, Ullmann's Encyclopedia of Industrial Chemistry, s. 146–152. American Cancer Society, 2000. ISBN 9783527306732. Teoksen verkkoversio. (englanniksi)
- DL Nelson, MM Cox: Lehninger principles of biochemistry, s. 517. 5. painos. W.H. Freeman, 2008. ISBN 9780716771081. Teoksen verkkoversio. (englanniksi)
- A Pennacchio et al: Insight into the stereospecificity of short-chain thermus thermophilus alcohol dehydrogenase showing pro-S hydride transfer and prelog enantioselectivity. Protein and Peptide Letters, 2010, 17. vsk, nro 4, s. 437–443. PubMed:19807673. doi:10.2174/092986610790963564. ISSN 1875-5305. Artikkelin verkkoversio. (englanniksi)
- CR Bellamacina: The nicotinamide dinucleotide binding motif: a comparison of nucleotide binding proteins. FASEB J, 1996, 10. vsk, nro 11, s. 1257–1269. PubMed:8836039. doi:10.1096/fasebj.10.11.8836039. ISSN 0892-6638. Artikkelin verkkoversio. (englanniksi)
- H Meng et al: Engineering a d-lactate dehydrogenase that can super-efficiently utilize NADPH and NADH as cofactors. Scientific Reports, 25.4.2016, 6. vsk, nro 24887. PubMed:27109778. doi:10.1038/srep24887. ISSN 2045-2322. Artikkelin verkkoversio. (englanniksi)
- SM Abdul-Awal et al: NO-Mediated [Ca2+]cyt increases depend on ADP-ribosyl cyclase activity in arabidopsis. Plant Physiology, 2016, 171. vsk, nro 1, s. 623–631. PubMed:26932235. doi:10.1104/pp.15.01965. ISSN 0032-0889. Artikkelin verkkoversio. (englanniksi)
- V Gasperi et al: Niacin in the central nervous system: an update of biological aspects and clinical applications. International Journal of Molecular Sciences, 23.2.2019, 20. vsk, nro 4. PubMed:30813414. doi:10.3390/ijms20040974. ISSN 1422-0067. Artikkelin verkkoversio. (englanniksi)
- SK Nigam et al: The organic anion transporter (OAT) family: a systems biology perspective. Physiological Reviews, tammikuu 2015, 95. vsk, nro 1, s. 83–123. PubMed:25540139. doi:10.1152/physrev.00025.2013. ISSN 0031-9333. Artikkelin verkkoversio. (englanniksi)
- Combs, s. 474
- Combs, s. 55
- DJ Lanska: Chapter 30: historical aspects of the major neurological vitamin deficiency disorders: the water-soluble B vitamins. Handbook of Clinical Neurology, 2010, 95. vsk, nro 95, s. 445–476. PubMed:19892133. doi:10.1016/S0072-9752(08)02130-1. ISSN 0072-9752. Artikkelin verkkoversio. (englanniksi)
- G Casal: Historia natural y médica del Principado de Asturias. en la Oficina de Manuel Martín, 1762. OCLC: 760668154. Teoksen verkkoversio. (englanniksi)
- Franciscus Frapolli: Physici Francisci Frapolli Animadversiones in morbum, vulgo pelagram. Galaetius, 1771. OCLC: 311638647. Teoksen verkkoversio.
- C Huber: Vorläufige notiz über einige derivate des nicotins. Justus Liebigs Annalen der Chemie, 1867, 141. vsk, nro 2, s. 271–271. doi:10.1002/jlac.18671410218. ISSN 1099-0690. Artikkelin verkkoversio. (englanniksi)
- CA Elvehjem, LJ Teply: The structure and estimation of natural products functionally related to nicotinic acid. Chemical Reviews, 1.12.1943, 33. vsk, nro 3, s. 185–208. doi:10.1021/cr60106a001. ISSN 0009-2665. Artikkelin verkkoversio. (englanniksi)
- C Huber: Vorläufige Mittheilung. Berichte der deutschen chemischen Gesellschaft, 1870, 3. vsk, nro 2, s. 849–849. doi:10.1002/cber.18700030254. ISSN 1099-0682. Artikkelin verkkoversio. (englanniksi)
- H Weidel: Zur kenntniss des nicotins. Justus Liebigs Annalen der Chemie, 1873, 165. vsk, nro 2, s. 328–349. doi:10.1002/jlac.18731650212. ISSN 1099-0690. Artikkelin verkkoversio. (saksaksi)
- J Goldberger: The Etiology of pellagra: the significance of certain epidemiological observations with respect thereto. Public Health Reports, 1914, 29. vsk, nro 26, s. 1683–1686. PubMed:808825. doi:10.2307/4570920. ISSN 0094-6214. Artikkelin verkkoversio. (englanniksi)
- J Goldberger, CH Waring, DG Willets: A test of diet in the prevention of pellagra. Southern Medical Journal, 1915, 8. vsk, nro 12, s. 1043–1044. ISSN 0038-4348. Artikkelin verkkoversio. (englanniksi)
- J Goldberger, CH Waring, WF Tanner: Pellagra prevention by diet among institutional inmates. Public Health Reports, 1923, 38. vsk, nro 41, s. 2361–2368. doi:10.2307/4576901. ISSN 0094-6214. Artikkelin verkkoversio. (englanniksi)
- J Goldberger, GA Wheeler: Experimental pellagra in the human subject brought about by a restricted diet. Public Health Reports, 1915, 30. vsk, nro 46, s. 3336–3339. doi:10.2307/4572984. ISSN 0094-6214. Artikkelin verkkoversio. (englanniksi)
- J Goldberger: The transmissibility of pellagra: experimental attempts at transmission to the human subject. Public Health Reports, 1916, 31. vsk, nro 46, s. 3159–3173. doi:10.2307/4574262. ISSN 0094-6214. Artikkelin verkkoversio. (englanniksi)
- J Goldberger, WF Tanner: A study of the pellagra-preventive action of dried beans, casein, dried milk, and brewers' yeast, with a consideration of the essential preventive factors involved. Public Health Reports, 1925, 40. vsk, nro 2, s. 54–80. doi:10.2307/4577425. ISSN 0094-6214. Artikkelin verkkoversio. (englanniksi)
- J Goldberger et al: A Further study of experimental blacktongue with special reference to the blacktongue preventive in yeast. Public Health Reports, 1928, 43. vsk, nro 12, s. 657–694. doi:10.2307/4578755. ISSN 0094-6214. Artikkelin verkkoversio. (englanniksi)
- DJ Lanska: The discovery of niacin, biotin, and pantothenic acid. Annals of Nutrition & Metabolism, 2012, 61. vsk, nro 3, s. 246–253. PubMed:23183297. doi:10.1159/000343115. ISSN 1421-9697. Artikkelin verkkoversio. (englanniksi)
- J Goldberger et al: A study of the blacktongue-preventive action of 16 foodstuffs, with special reference to the identity of blacktongue of dogs and pellagra of man. Public Health Reports, 1928, 43. vsk, nro 23, s. 1385–1454. doi:10.2307/4578856. ISSN 0094-6214. Artikkelin verkkoversio. (englanniksi)
- CA Elvehjem et al: The isolation and identification of the anti-black tongue factor. Journal of Biological Chemistry, 1.3.1938, 123. vsk, nro 1, s. 137–149. ISSN 0021-9258. Artikkelin verkkoversio. (englanniksi)
- CA Elvehjem, RJ Madden, AE Axelrod: The effect of a nicotinic acid deficiency upon the coenzyme I content of animal tissues. Journal of Biological Chemistry, 1.11.1939, 131. vsk, nro 1, s. 85–93. ISSN 0021-9258. Artikkelin verkkoversio. (englanniksi)
- Council on Foods and Nutrition. Journal of the American Medical Association, 7.3.1942, 118. vsk, nro 10, s. 819–819. doi:10.1001/jama.1942.02830100049011. ISSN 0002-9955. Artikkelin verkkoversio. (englanniksi)
- Niacin and nicotinic acid. Journal of the American Medical Association, 7.3.1942, 118. vsk, nro 10, s. 823–823. doi:10.1001/jama.1942.02830100053014. ISSN 0002-9955. Artikkelin verkkoversio. (englanniksi)
- C. A. Elvehjem et al: Growth-retarding effect of corn in nicotinic acid-low rations and its counteraction by tryptophane. Science, 11.5.1945, 101. vsk, nro 2628, s. 489–490. PubMed:17735529. doi:10.1126/science.101.2628.489. ISSN 0036-8075. Artikkelin verkkoversio. (englanniksi)
- G. A. Goldsmith: Niacin-tryptophan relationships in man and niacin requirement. The American Journal of Clinical Nutrition, 1.9.1958, 6. vsk, nro 5, s. 479–486. doi:10.1093/ajcn/6.5.479. ISSN 0002-9165. Artikkelin verkkoversio. (englanniksi)
- GM Briggs, K Carpenter: Letter: Niacin dervatives not "vitamin B3". The New England Journal of Medicine, 1.8.1974, 291. vsk, nro 5, s. 263. PubMed:4275865. doi:10.1056/NEJM197408012910526. ISSN 0028-4793. Artikkelin verkkoversio. (englanniksi)
- A Hoffer: "Vitamin B3". Canadian Medical Association Journal, 1.12.1986, 135. vsk, nro 11, s. 1250. PubMed:3779554. ISSN 0820-3946. Artikkelin verkkoversio. (englanniksi)
Aiheesta muualla
- Fineli: niasiinin lähteet ruoka-aineissa
- Linus Pauling Institute: Niacin (englanniksi)
- DSD: niasiinientsyymien stereospesifisyyden tietokanta (Arkistoitu – Internet Archive)
- PubChem: Nikotiinihappo (englanniksi)
- PubChem: Nikotiiniamidi (englanniksi)
- Dr. Duke's Phytochemical and Ethnobotanical Databases: Niacin (englanniksi)