Ugalketa asexual
Ugalketa asexuala gametoen batuketarik (ernalkuntzarik) gabeko ugalketa-mota da eta normalean kromosoma kopuruak (ploidia) ez du aldaketarik pairatzen. Ugalketa mota honen bidez lortzen diren ondorengoek gurasoaren geneak heredatzen dituzte. Ugalketa mota hau organismo zelulabakarren (Archaea eta Bacteria) ohiko ugalketa da; hala ere, hainbat animalia zelulaniztun, landare eta onddo asexualki ugal daitezke.[1]

Prokarioto gehienak geneen transferentzia horizontala egiten dute, eta hain zuzen ere, konjugazio, transformazio eta transdukzio deritze. Nahiz eta gametoen eraketarik eta fusiorik gabe ugaltzen diren, geneen transferentzia horizontala ugalketa sexualarekin erlazionatuta dago, meisosian gertatzen den birkonbinazio genetikoa gertatzen delako.[2] Beraz, ugalketa asexual hertsiaz mintzo garenean, birkonbinazio genetikorik gabeko ugalketaz ari gara.
Motak
Ugalketa asexuala hainbat organismotan gertatzen da, baina organismo bakoitzak mota desberdina eramaten du aurrera. Atal honetan ugalketa asexual mota esangarrienak azalduko dira.
Zelula-zatiketa
Prokariotoak (Archaea eta Bacteria) asexualki ugaltzen dira erdibitze baten bidez. Prozesu horretan organismo bat bi zatitan banatzen da, eta genetikoki berdinak diren bi organismo alaba lortzen dira. Eukariotoek (protistoek eta onddo zelulabakarrek) antzeko prozesua jarraitzen dute, mitosia deritzon prozesua hain zuzen. Hainbat protistotan (esporozoo eta algetan) erdibitze anizkoitza gertatzen da: mitosiaren bidez organismo amaren nukleoa hainbat nukleo txikiagotan banatzen da eta, ondoren, zitoplasma banandu, eta hainbat zelula alaba lortzen dira. [3][4][5] Hala ere, Eukariota domeinuko zelula gehienak sexualki ugaltzeko gai dira.
Gemazioa
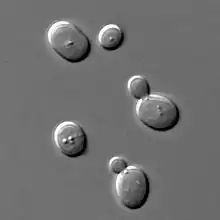
Zelulabakar batzuk (legamiak) eta zelulaniztun batzuk (Hydra)[6] gemazio bidez ugaltzen dira. Gemazioaren hasieran, ama-zelulan, zelula alaba izango den protuberantzia moduko bat hazten da. Hasieran, zelula alaba tamainaz nahikoa garatu ez den bitartean, zelula amari itsatsita geratzen da, eta bertan hazten da. Behin jada tamaina nahikoa lortzen duenean eta zelula alaba guztiz heldutzat har daitekeenean, zelula amarekiko lotura apurtu, eta bi organismo independente garatzen dira.
Parasitoetan, barne-gemazioa gertatzen da. Adibidez, Toxoplasma gondii protozooa barne-gemazioaren bidez ugaltzen da. Zelula amaren barnean bi zelula alaba (endodygeny) edo gehiago (endopolygeny) sortzen dira, eta horiek zelula ama elikagai modura erabiltzen dute hazteko eta ondoren banatzeko.[7] Kanpoko zein barneko gemazioa Tenia edo Echinococcus bezalako arretan ere ikusi izan da. Organismo horiek kiste moduko batzuk garatzen dituzte, eta kiste horietan ugaltzen dira gemazio bidez.
Propagazio begetatiboa

Propagazio begetatiboa landareetan gertatzen den ugalketa asexual mota bat da. Prozesu horretan indibiduo berriak hazi edo espora gabe garatzen dira, hau da, ez da meiosirik edo singamiarik [8] gertatzen. Ugalketa mota hau jarraitzen duten landareek, adibidez Kalanchoe landareak (Bryophyllum daigremontianum), hosto espezializatu batzuetan plantula deritzen landare txikiak sortzen dituzte. Askok landare berriak sortzen dituzte errizometatik edo estoloietatik, adibidez, marrubiko landareek. Beste landare batzuek erraboilak edo tuberkuluak erabiliz ugaltzen dira, hala nola, Tulipak erraboilaren bidez eta Daliak tuberkuluaren bidez. Landare batzuek ernamuinak leku arrotzetan sortuz garatzen dira, eta koloniak sortzen dituzte. Adibide hauek guztiak indibiduo klonalei dagozkie. Gainera, propagazio begetatiboak eremu zabala estaltzeko aukera eskaintzen die. [9]
Espora-eraketa
Organismo zelulaniztun askok esporak garatzen dituzte haien ziklo biologikoan zehar. Esporak sortzeko prozesuari esporogenesi deritzo. Animalia eta protistak dira prozesu hau egiten ez duten salbuespenak; izan ere, horiek meiosia egin eta ernalkuntza jarraian egiten dute. Bestalde, landareek eta hainbat algak meiosia egiten dute espora haploideak sortzeko eta ez gametoak sortzeko, aurretik aipatu dugun moduan. Espora haploide horiek indbiduo zelulaniztunetan garatzen dira, gametofito deritzenetan hain zuzen ere. Gametofitoa eratzeko prozesuan, ez da ernalkuntzarik gertatzen. Indibiduo zelulaniztun haploideak, hau da, gametofitoak gametoak sortzen ditu mitosi bidez. Dena den, gametoen eraketa eta meiosia bizi-zikloaren estadio desberdinetan gertatzen dira, eta horri belaunaldi-txandaketa esaten zaio. Prozesu horretan ploidiaren murrizketa gertatzen da, eta hori ugalketa sexualaren ezaugarrietako bat da. Kasu hori ordea, ugalketa asexual gisa sailkatzen da, gametoen fusiorik gertatzen ez delako. [10]
Onddo eta alga batzuetan esporen eraketa, ugalketa asexual hertsiaren bidez gertatzen da. Prozesu horretan, mitosia gertatzen da eta mitosporak deritzogun zelula ugalkorrak sortzen dira. Mitosporak indibiduo berrietan garatzen dira dispertsatu eta gero. Ugalketa mota hori konidioek [11] sortzen dituzten onddoetan eta alga gorrietan (Polysiphonia) gertatzen da, eta meiosi gabeko esporogenesia da. Kasu honetan, esporen kromosoma kopuruak ez du aldaketarik pairatzen. Hala ere, esporogenesi mitotikoa salbuespena da, espora gehienak meiosiaren bidez sortzen baitira.
Fragmentazioa
Fragmentazioan, organismo baten zatiki batetik guztiz garatutako indibiduo berri bat lortzen da. Ugalketa mota hori hainbat organismotan ageri da: asexualki ugaltzen diren animalietan, hala nola, planaria taldeko animalietan, anelido askotan (poliketoetan [12] eta oligoketo [12] batzuetan), turbelarioetan eta itsas izarretan. Hainbat onddo eta landare asexualki ugaltzen dira eta landare batzuk (Marchantiophyta dibisioko batzuk) egitura espezializatuak dituzte fragmentazio bidez ugaltzeko. Likenak sortzen dira onddoen eta alga fotosintetikoen edo zianobakterioen arteko elkarketa sinbiotikoari esker. Likenek fragmentazioaren bidez ziurtatzen dute hurrengo belaunaldiko indibiduoek elkarketa sinbiotikoa mantenduko dutela. Sortutako zatikiak soredio formakoak izan daitezke: hauts formako partikulak non onddoen hifa zelula fotobiotikoak inguratzen dituen.
Organismo kolonialetan edo zelulaniztunetan, organismo bat erdibitu egiten da eta zatiki bakoitzetik indibiduo berriak garatzen dira. Sortzen diren indibiduoak genetikoki berdinak dira, bai haien artean, baita gurasoekiko ere. Kontuan izan behar da ingurumenak eta epigenetikak organismoetan duten eraginaren ondorioz, nahiz eta bi indibiduo arbaso bakarretik garatu eta klonak izan, genetikoki desberdinak izan daitezkeela. [13]
Agametogenesia
Agametogenesian, gameto arraren beharra ez duen ugalketa mota da. Bi motatakoa izan daiteke, partenogenesia edo apomixia.
Partenogenesia
Partenogenesian, ernaldu gabeko obulutik organismo berri bat garatu daiteke. Ugalketa mota hau gertatzen da landare askotan, ornogabe batzuetan (errotifero, inurri, erle eta liztor parasitikoetan), eta ornodun batzuetan (narrasti batzuetan, anfibioetan, marrazo gutxitan eta hegazti batzuetan).
Meiosiaren ostean sortzen diren obuluak haploideak dira. Hau da, amaren zelula somatikoek duten kromosoma kopuruaren erdia dute. Animalien kasuan, indibiduoak haploideak izatea ez da bideragarria izaten. Partenogenesiaren bidez sortzen diren organismoak diploideak direnez, hainbat mekanismo garatu dira hori gertatzeko.
Alde batetik, obuluak meiosia gertatu gabe garatu daitezke. Horrela, kromosomen kopurua ez dute berreskuratu behar, eta gainera, sortuko diren ondorengoak amarekiko guztiz klonikoak izango dira, hau da, haren material genetiko osoa eta berdina edukiko dute.
Bestalde, meiosia gertatuz gero, diploidia berreskuratzeko mekanismoak garatu behar dira. Gainera, lortzen diren ondorengoak guztiz klonikoak ez direnez, klon partzial esaten zaie. Izan ere, gametoak sortzen direnean, meiosia gertatzeaz gain, birkonbinazio genetikoa ere gertatzen da. Hortaz, ondorengoek amaren material genetikoa edukitzen dute, baina ez dira berdinak izaten.
Apomixia
Landareetan gertatzen den ugalketa mota bat apomixia da, ernalketa gabeko esporofito berri baten sorkuntzan datzana. Oso ugalketa mota garrantzitsua da iratzeetan eta angiospermoetan, baina azken talde horretan soilik kasu batzuetan gertatzen da. Angiospermoetan, “apomixis” terminoak ernalketa gabeko hazien garapena adierazteko erabiltzen da, nahiz eta behinola ugalketa begetatiboa adierazteko ere erabiltzen zen. Landare apomitiko baten adibidea Taraxacum familiako landare triploideak dira. Apomixia bi modutan gertatzen da. Lehen modua, apomixia gametofitikoa da: enbrioia ernaldu gabeko arrautza batetik garatzen da, meiosia jasan ez duen enbrioi-zaku diploide baten barruan dagoena hain zuzen. Beste modua enbrioi nuzeletikoarena da: enbrioia enbrioi-zakua inguratzen duen ehun diploidetik (nuzela) garatzen da. Azken prozesu hori zitrikoen hazi batzuetan ageri da. Apomixia indibiduo arretan gertatzea nahiko arraroa da; hala ere, Saharako zipresetan (Cupressus dupreziana) ikusi da indibiduo horietan enbrioiaren material genetiko guztia polenetik datorrela.
Ugalketa sexualaren eta asexualaren arteko txandaketa
Espezie batzuk egoera desberdinen arabera, ugalketa sexualaren eta asexualaren arteko txandaketa egin dezakete: zenbait protisto eta onddok ugalketa sexuala eta asexuala txandakatzen dituzte; anfibio, narrasti eta hegazti batzuek ere antzeko gaitasunak dituzte. Gaitasun horri heterogamia deritzo. Txandaketa hori zenbait errotifero-espezietan ikus daiteke (partenogenesi ziklokoa e.g. Brachionus espezieetan), eta baita intsektu mota batzuetan ere, hala nola, afidoetan. Afidoek, baldintza jakin batzuetan, meiositik igaro ez diren arrautzak ekoizten dituzte, beraien burua klonatuz. El Cabo-ko Apis mellifera subsp. capensis erlea asexualki ugal daiteke Telitoki prozesuaren bidez. Ur gezetako Daphnia krustazeoa udaberrian partenogenesi bidez ugaltzen da urmaelak azkar populatzeko. Ondoren, harrapaketa eta lehia handitzen doazen neurrian, ugalketa sexualera aldatzen da. Brachionus generoko errotiferoak partenogenesi zikliko bidez ugaltzen dira: dentsitate baxuko populazioetan emeak asexualki ugaltzen dira eta, dentsitate altukoetan, ordea, seinale kimiko bat metatzen da eta ugalketa sexualerako trantsizio bat induzitzen da.
Dictyostelium onddo lingirdatsua baldintza faboragarrietan, mitosi bidez ugaltzen da ameba zelulabakarra balitz bezala. Hala ere, baldintzak desfaboragarri bihurtzen direnean, zelulak agregatu eta garapenerako bi bide desberdin jarraitzen dituzte. Lehenengoa, bide soziala da eta bertan, bare zelulanitz bat eratzen dute, gero gorputz fruitu-emailea eratuko duena asexualki sortutako esporekin. Bigarrena, bide sexuala da, eta bi zelula fusionatu, eta zelula erraldoi bat eratzen dute, ondoren kiste handi bihurtuko dena. Kiste handi hori hozitutakoan, ehunka ameba-zelula askatzen ditu. Horiek jatorrizko bi zelulen birkonbinazio meiotikoaren emaitza dira. Onddoak zein bide hautatuko duen ingurune-baldintzen araberakoa da.
Onddo arruntaren (Rhizopus) hifak espora mitotiko zein meiotikoak sortzeko gai dira. Alga askok modu berdintsuan txandakatzen dituzte ugalketa sexuala eta asexuala. Zenbait landarek bi ugalketa-motak, sexualak eta asexualak, erabiltzen dituzte landare berriak sortzeko, espezie batzuek txandakatu egiten dituzte beraien ugalketa modu primarioak sexualetik asexualera baldintza aldakorrenpean.
Adibideak organismo zelulanitzetan
Landareetan[14]
Ugalketa sexualean ez bezala, ugalketa asexualean ez da gameto arren eta emeen arteko elkarketarik gertatzen. Oro har, landareetan bi ugalketa asexual mota gerta daitezke: ugalketa begetatiboa eta apomixia. Edozein izanda ere, lortuko diren organismo berriak aurrekoen klonak izaten dira. Ugalketa begetatiboaren kasuan, beharrezkoa da jatorrizko landarearen zati begetatibo bat. Apomixia, aldiz, ugalketa sexuala ordezkatzen duen mekanismoa da, batzuetan hazien sorrera suposatzen duena. Azken mekanismo hori, landare espezie askotan gertatzen da, baita landareak ez diren bestelako organismo batzuetan ere.
Ugalketa begetatiboa gemazioz edota kimuen bidez gertatzen da batez ere landareetan, normalean zurtoinean edo sustraietan egiturazko aldaketak dakartzana. Hala ere, espezie batzuetan aldaketak hostoetan ere gerta daitezke. Gehien bat zurezko landaretan eta landare iraunkor belarkarretan gertatzen den prozesua da eta, landare horien kolonia klonikoak (ramet-ak) eratzea dakar. Hala ere, ugalketa mota horren bidezko hedapen ahalmena murritza da, landare batzuek errizoma adarkatuen bidez ramet nahiko hedatuak eratzeko ahalmena badute ere. Izan ere, ugalketa begetatiboa ez da horren garrantzitsua ugalketarako. Organismoen biziraupenerako eta biomasaren emendiorako, ordea, oso garrantzitsua da. Gainera, landareen ugalketa begetatiboa garrantzitsua da gizakiontzat nekazaritzan eta loregintzan erabiltzen delako: horren bidez intereseko landare baten ezaugarri berdinak lo ditzakegu indibiduo berri batean.
Apomixia prozesuan haziak sortzen dira, baina ez da enbrioiaren ernalketarik gertatzen horretarako. Hazi apomiktikoak dituzten landare batzuetan, pseudogamia gertatzen da. Horretarako, beharrezkoa da polinizazioa gertatzea enbrioiaren garapena sustatzeko, nahiz eta polenaren material genetikoa ez den ondorengoetara pasako[15]. Apomixia gertatzeko beste modu batzuk daude: plantulen sorrera gerta daiteke haziak ordezteko edota bulbiloak garatu daitezke loreen ordez.
Animalietan[16]
Animalietan ugalketa asexual ohikoena partenogenesia da. Horren bidez, amaren berdina den indibiduo bat sortzen da. Horren adibide dira mailu-arrainak[17] (Sphyrnidae familia) eta Carcharhinus limbatus espeziea[18]. Marrazo-talde horien emeak gatibutasunean eta arren presentziarik gabe hazita ere, ondorengoak izateko ahalmena erakutsi zuten: amarekiko klonikoak ziren ondorengoak.
Narrasti batzuetan sexuaren determinaziorako ZW sistema deskribatu da. Horren arabera, arrek ZZ kromosoma sexualak dituzte eta emeek, aldiz, ZW edo WW kromosoma sexualen pareak eduki ditzakete. 2010. urtera arte, WW zuten indibiduoak garatzea ezinezkoa zela pentsatzen zen. Urte horretan bertan ikusi zen ZW genotipoa zuen boa hertsatzaile eme batek WW genotipoa zuten ondorengo emeak izan zituela[19]. WW genotipodun ondorengoek ugalketa asexualaren bitartez sortu ziren.
Poliembrionia animalien artean hedatuta dagoen ugalketa asexualerako prozesua da. Prozesu honetan obulu ernaldua edo enbrioiaren zelulak bikoiztu, eta genetikoki identikoak diren klonak sortzen dituzte. Prozesu hori asko ikertu da Hymenoptera taldeko kide bizkarroietan.
Ugalketa asexualaren ezaugarri genetikoak eta ikuspegi ebolutiboa
Atal honetan, organismo eukariotoen ezaugarrietan sakonduko dugu gehien bat. Zehazkiago, organismo horien ugalketa asexualaren ezaugarri genetikoetan eta ugalketa asexualak eboluzioan duen ondoriotan sakonduko dugu. Ugalketa asexuala gertatzen da gurasoekiko ondorengo genetikoki identikoak sortzen direnean. Genetikoki identikoak dira loci mailan baita genoma mailan ere. Gertatzen diren aldaketa genetiko bakarrak mutazio somatikoen bidezkoak izaten dira. Beraz, ugalketa asexual terminoa ugalketa klonak terminoaren sinonimo gisa erabil genezake. [20]
Ugalketa asexual hertsia egiten duten goi mailako organismo eukariotoak % 0,1-1 direla estimatzen da. Dena den, estimazio horretan, goi mailako organismo eukariotoak eta ugalketa sexuala % 0an egiten duten organismoak hartzen dira kontuan. Horrekin esan nahi duguna da sailkapenak egiteko jartzen ditugun baldintzek erabat aldatuko dutela. Beraz, emaitzak interpretatzeko orduan, kontuan eduki beharko ditugu hasieratik ezarritako baldintza horiek. [20]
Ikuspegi genetikoa
Ugalketa asexual hertsiaren ondorio interesgarrietako bat Meselson efektua da. Lehenengo aldiz eta bakarraz deskribatu izan zen errotiferoetan. Organismo diploideen lerroak hainbat belaunalditan zehar ugalketa sexualik gabe ugaltzen direnean, espero dugu aleloek locus bakoitzean mutazio somatikoen bidezko aldakortasun genetikoa metatzea. Ondorioz, aleloen arteko dibergentzia genetiko maximoa lortuko da. Hori fenomeno arraroa da ordea, eta lehen aipatu dugun moduan, soilik errotifero bdelloideetan deskribatu da. Izan ere, Meselson-efektua gerta dadin, ez da geneen konbertsiorik gertatu behar, konbertsioak populazioak homogeneizatzen baititu.[20] Epe luzeko inplikazioak dira horiek, belaunaldi askoren zatiketak behar baitira ondorio horiek kontuan hartzeko modukoak izateko.
Bestalde, epe laburrean ere baditu inplikazioak ugalketa asexualak. Izan ere, hurrengo belaunaldirako (F1) espero diren genotipoen maiztasunak aldatzen ditu. Izan ere, ugalketa sexualerako badago Hardy-Wrinberg orekaren teoria. Teoria hori baliagarria da hurrengo belaunaldian aterako diren genotipoen maiztasunak estimatzeko. Ugalketa asexuala gertatzen den kasuetan, ondorengo heterozigotoak ateratzeko joera handiagoa deskribatu da. [20]
Ikuspegi ebolutiboa
Goi mailako organismoetan gehien deskribatu den ugalketa-estrategia ohikoena ugalketa sexuala da. Dena den, biologia ebolutiboan ikertzen jarraitzen den gaia da, eta oraindik zergatia ez dago guztiz argi. [20] Hainbat hipotesi daude ugalketa-estrategia bakoitzaren alde eta kontra.
Lehenik eta behin, ugalketa-estrategia bakoitzak, asexualak eta sexualak, abantaila desberdinak dituela hartu behar dugu kontuan. Horregatik, organismoaren araberakoa da ugalketa estrategia arrakastatsuena. Ugalketa sexualaren ageriko onura da geneen konbinazio berrien sorreraren emendioa, organismoari etengabe aldatzen ari den ingurune bati moldatzeko aukera anitzagoak eskaintzen dizkiona. Dena den, kontuan eduki behar dugu ugalketa sexualak sortzen dituen konbinazio genetikoak ez direla zertan onuragarriak izan, hau da, gertatzen diren konbinazioak ez dira direkzionalak eta ez daude diseinatuta. Ausaz gertatzen diren konbinazioak dira, eta ausazkoak izatearen arazoa da txarrerako konbinazioak ere sor daitezkeela. Beraz, beti ez da onuragarria izango ugalketa sexuala.[20] Baina aurretik aipatutakoaren harira, ugalketa asexualaren onura gehienak epe motzekoak izaten dira, besteak beste, kolonizatzeko aukera handiagoa eta ugaltzeko abiadura handiagoa. [21]Baina, ingurunera moldatzeko aukera gehiago eskaintzen ditu ugalketa sexualak, eta beraz, hori ugalketa sexuala gailendu izatearen erantzuna izan daiteke. Dena den, hasieran aipatu bezala, gailentze horren zergatia oraindik ez dago argi.
Erreferentziak
- Artikulu honen edukiaren zati bat Lur hiztegi entziklopedikotik edo Lur entziklopedia tematikotik txertatu zen 2011/12/27 egunean. Egile-eskubideen jabeak, Eusko Jaurlaritzak, hiztegi horiek CC-BY 3.0 lizentziarekin argitaratu ditu, Open Data Euskadi webgunean.
- (Ingelesez) Engelstädter, Jan. (2017-06-01). «Asexual but Not Clonal: Evolutionary Processes in Automictic Populations» Genetics 206 (2): 993–1009. doi:. ISSN 0016-6731. PMID 28381586. (Noiz kontsultatua: 2020-11-16).
- (Ingelesez) Narra, Hema Prasad; Ochman, Howard. (2006-09). «Of What Use Is Sex to Bacteria?» Current Biology 16 (17): R705–R710. doi:. (Noiz kontsultatua: 2020-11-16).
- (Ingelesez) «reproduction | Definition, Examples, Types, Importance, & Facts» Encyclopedia Britannica (Noiz kontsultatua: 2020-11-15).
- Fungi, algae, and protists. (1st ed. argitaraldia) Britannica Educational Pub. in association with Rosen Educational Services 2011 ISBN 978-1-61530-463-9. PMC 681412451. (Noiz kontsultatua: 2020-11-15).
- Puranik, Prakash.. (2007). Animal forms & functions : invertebrata. Sarup & Sons ISBN 81-7625-791-5. PMC 294988919. (Noiz kontsultatua: 2020-11-15).
- (Ingelesez) «IV. Part of a letter from Mr Antony van Leeuwenhoek, F. R. S. concerning green weeds growing in water, and some animalcula found about them.» Philosophical Transactions of the Royal Society of London 23 (283): 1304–1311. 1703-12-31 doi:. ISSN 0261-0523. (Noiz kontsultatua: 2020-11-15).
- Smyth, J. D. (James Desmond), 1917-1999.. (1994). Introduction to animal parasitology. (3rd ed. argitaraldia) Cambridge University Press ISBN 0-521-41770-8. PMC 28798892. (Noiz kontsultatua: 2020-11-15).
- «Asexual Reproduction» ucmp.berkeley.edu (Noiz kontsultatua: 2020-11-16).
- «Celebrating Wildflowers - Fading Gold - How Aspens Grow» web.archive.org 2010-09-23 (Noiz kontsultatua: 2020-11-16).
- «Landare-ugalketa - hiru» www.hiru.eus (Noiz kontsultatua: 2020-11-16).
- «Plantas y Hongos» www.plantasyhongos.es (Noiz kontsultatua: 2020-11-16).
- Ruppert, Edward E.. (2004). Invertebrate zoology : a functional evolutionary approach. (7th ed. argitaraldia) Thomson-Brooks/Cole ISBN 0-03-025982-7. PMC 53021401. (Noiz kontsultatua: 2020-11-16).
- (Ingelesez) Neuhof, Moran; Levin, Michael; Rechavi, Oded. (2016-09-15). «Vertically- and horizontally-transmitted memories – the fading boundaries between regeneration and inheritance in planaria» Biology Open 5 (9): 1177–1188. doi:. ISSN 2046-6390. PMID 27565761. PMC PMC5051648. (Noiz kontsultatua: 2020-11-16).
- (Ingelesez) Plant reproduction. 2020-11-20 (Noiz kontsultatua: 2020-11-21).
- «Why apomixis is genetic gold - Australian Life Scientist» web.archive.org 2009-10-26 (Noiz kontsultatua: 2020-11-21).
- (Ingelesez) Asexual reproduction. 2020-11-25 (Noiz kontsultatua: 2020-11-26).
- (Ingelesez) Eilperin, Juliet. (2007-05-23). Female Sharks Can Reproduce Alone, Researchers Find. ISSN 0190-8286. (Noiz kontsultatua: 2020-11-26).
- (Ingelesez) Chapman, D. D.; Firchau, B.; Shivji, M. S.. (2008). «Parthenogenesis in a large-bodied requiem shark, the blacktip Carcharhinus limbatus» Journal of Fish Biology 73 (6): 1473–1477. doi:. ISSN 1095-8649. (Noiz kontsultatua: 2020-11-26).
- (Ingelesez) «Boa constrictor produces fatherless babies | CBC News» CBC (Noiz kontsultatua: 2020-11-26).
- (Ingelesez) de Meeûs, T.; Prugnolle, F.; Agnew, P.. (2007-06). «Asexual reproduction: Genetics and evolutionary aspects» Cellular and Molecular Life Sciences 64 (11): 1355–1372. doi:. ISSN 1420-682X. (Noiz kontsultatua: 2020-11-20).
- (Ingelesez) Dawson, Kevin J.. (1995-10-07). «The Advantage of Asexual Reproduction: When is it Two-fold?» Journal of Theoretical Biology 176 (3): 341–347. doi:. ISSN 0022-5193. (Noiz kontsultatua: 2020-11-21).
Kanpo estekak
| Autoritate kontrola |
|---|
 Datuak: Q173432
Datuak: Q173432 Multimedia: Asexual reproduction / Q173432
Multimedia: Asexual reproduction / Q173432