Nerbio-sistema
Biologian, nerbio sistema animalia baten zati oso konplexu bat da, organismoen ekintza eta informazio sentsoriala koordinatzen dituena seinaleak transmitituz gorputzeko zati desberdinetatik. Nerbio sistemak gorputzari eragiten dioten inguruneko aldaketak antzematen ditu, gero tandemean egiten du lan sistema endokrinoarekin gertakizun horiei aurre egiteko[1]. Nerbio ehuna lehenengoz zizare moduko organismoetan agertu zen duela 550 eta 600 milioi urte artean. Ornodunetan nerbio sistema bi zati nagusiez dago osatuta, nerbio sistema zentrala (NSZ) eta nerbio sistema periferikoa (NSP). NSZa garuna eta bizkarrezur-muinaz dago osatuta. NSP gehienbat nerbioez dago osatuta, axoien edo zuntz luzeen sorta itxiak direnak, NSZ gorputzeko beste zati guztiekin lotzen dituztenak. Garunetik seinaleak transmititzen dituztenei nerbio motor edo eferente deritze; informazioa gorputzetik NSZra transmititzen dituztenak, aldiz, nerbio sentsorial edo aferenteak deritze. Bizkarrezurreko nerbioak bi funtzioak dituzte eta nerbio mistoak deritze. Nerbio sistema periferikoa hiru azpisistemetan dago banatuta: nerbio sistema somatikoa, autonomoa eta enterikoa. Nerbio somatikoek borondatezko mugimenduak bideratzen dituzte. Nerbio sistema autonomoa, nerbio sistema sinpatiko eta parasinpatikoan dago banatuta. Nerbio sistema sinpatikoa larrialdi egoeretan aktibatzen da energia mobilizatzeko, nerbio sistema parasinpatikoa, ordea, organismoak erlaxazio egoeran daudenean aktibatzen da. Nerbio sistema enterikoak sistema gastrointestinala kontrolatzen du. Bai nerbio sistema autonomikoak, bai enterikoak nahi gabeko funtzionamendua dute.
| Nerbio-sistema | |
|---|---|
 Giza nerbio-sistema: horiz: nerbio-sistema zentrala; urdinez: nerbio-sistema periferikoa | |
| Xehetasunak | |
| Honen parte | sistema neurobaskularra |
| Konponenteak | Nerbio-sistema zentrala Nerbio-sistema periferikoa Neurona glia |
| Identifikadoreak | |
| Latinez | systema nervosum |
| MeSH | A08 |
| TA | A14.0.00.000 |
| FMA | 7157 |
| Terminologia anatomikoa | |
Maila zelularrean, nerbio sistema zelula mota berezi baten presentziaz dago definituta, neurona deritzona[2], nerbio zelula ere deitua. Neuronek egitura berezi bat dute beste zelulei seinaleak azkar eta modu zehatzean bidaltzea ahalbidetzen dietena. Seinaleak uhin elektrokimiko moduan bidaltzen dituzte axoi izeneko zuntz finetan barrena, neurotransmisore izeneko substantzia kimikoak isurtzea eragiten dutenak sinapsi izeneko lotunetan. Neurona batetik seinale sinaptiko bat jasotzen duen zelula bat kitzikatua, inhibitua edo modulatua izan daiteke. Neuronen arteko konexioek bide neuronalak, zirkuitu neuronalak eta sare handiagoak eratzen dituzte organismoen munduaren pertzepzioa sortu eta haien portaera zehazten dutenak. Neuronez gain, nerbio sistemak beste zelula espezializatu batzuez dago osatuta glia zelulak (edo soilik glia) deiturikoak euskarri estruktural eta metabolikoa ematen diena.
Nerbio sistemak animalia zelulaniztun gehienetan aurkitu daitezke, baina nabarmenki aldakorrak dira konplexutasunari dagokionez. Belakiak, plakozooak eta mesozooak dira nerbio sistemarik ez duten animalia zelulaniztun bakarrak, oso gorputz sinpleak dituztenak. Erradialki simetrikoak diren organismoen nerbio sistemak, ktenoforoak (orrazi marmokak) eta knidarioak (anemonak, hidrak, koralak eta marmokak) nerbio sare hedatu batez daude osatuta. Beste animalia espezie guztiak, zizare mota batzuk izan ezik, garun bat, muin zentral bat (edo 2 muin paraleloan ezarriak) eta garunetik eta muin zentraletik ateratzen diren nerbioz osaturiko nerbio sistema bat dute. Nerbio sistemaren tamaina aldakorra da, ehun bat zelulatik zizare sinpleenetan, 300.000 milioi zeluletara elefante afrikarretan[3].
Nerbio sistema zentralak zelula batetik bestera edo gorputzaren zati batetik bestera seinaleak bidali eta erantzuna jasotzen du. Nerbio sistemaren funtzionamendu txarra gerta daiteke akats genetikoen ondorioz, traumatismoen ondorioz edo toxikotasunaren ondorioz, baita infekzioen edo zahartze biologikoaren ondorioz ere. Neurologia da nerbio sistemaren gaixotasunak aztertzen dituen zientzia eta hauek saihesteko asmakizun edo tratamenduak bilatzen dituena. Nerbio sistema periferikoan, arazo ohikoena nerbio kondukzioaren akatsa da, arrazoi desberdinengatik gerta daiteke neuropatia diabetikoa eta nahaste desmielizatzailea, adibidez, esklerosi anizkoitza eta alboko esklerosi amiotrofikoarekin batera. Neurozientzia nerbio sistema aztertzen duen zientziaren adarra da.
Egitura
Nerbio sistemaren izena nerbioetatik eratorria da, garunetik bizkarrezur muinera doazen zuntz zilindriko sorta bat direnak (neuronen axoiak), behin eta berriz adarkatuz gorputzeko zati guztiak inerbatzeko[4]. Nerbioak antzinako egiptoar, greko eta erromatarrek[5], antzemateko nahiko handiak dira, baina horien barne egitura ez zen ezaguna izan mikroskopioarekin aztertzea posible izan zen arte. Michael Nikoletseas autoreak idatzi zuen[6]:
| « | Zaila da sinistea gutxi gorabehera 1900. urtea arte ez zela ezagutzen neuronak garunaren oinarrizko unitatea zirenik (Santiago Ramón y Cajal). Berdinki harrigarria, garunean substantzia kimikoen transmisioaren kontzeptua ez zela 1930 arte ezagutu (Henry Hallet Dale eta Otto Loewi). Beraien artean komunikatzeko neuronek erabiltzen duten oinarrizko fenomeno elektrikoa, ekintza potentziala, 1950eko hamarkadan hasi ginen ulertzen (Alan Lloyd Hodgkin, Andrew Huxley eta John Eccles). 1960eko hamarkadan konturatu ginen sare neuronalak estimulatzen diren sinpletasunaz eta oinarrizko kontzeptu horiek nola diren posible (David H. Hubel eta Torsten Wiesel). Iraultza molekularra AEB-ko unibertsitateetatik 1980ko hamarkadan zabaldu zen. 1990eko hamarkadan hedatu zen portaera fenomenoen mekanismo molekularren ezagutza (Eric Richard Kandel). | » |
Azterketa mikroskopiko batek nerbioak nagusiki axoiez osatuta daudela erakutsi zuen, horiek estali eta faszikuluetara jariatzen dituzten ehun desberdinekin batera. Nerbioak sortzen dituzten neuronak ez daude guztiz nerbioen barruan, haien zelula-gorputzak garunean, bizkarrezurrean edo ganglio periferikoan bizi dira[7].
Belakiak baino garatuagoak diren animalia guztiak nerbio sistema dute. Hala ere, belakiek, animalia zelulabakarrek eta animaliak ez diren organismoek, esaterako, onddo lirdingatsuek ere neuronen aitzindariak diren zelula-zelula seinaleztapen mekanismoak dituzte[8]. Erradialki simetrikoak diren animalien, adibidez, marmoka edo hidren nerbio sistema nerbio sare batez dago osatuta, zelula isolatuen sare zabaldu bat[9]. Animalia bilateraletan, existitzen diren espezieen gehiengoa osatzen dutenak, haien nerbio sistemek egitura komun bat dute Ediacarar garai goiztiarrean duena jatorria, duela 550 milioi urte baino gehiago[10][11].
Zelulak
Nerbio sistemak bi zelula mota nagusiz osatuta dago: neuronak eta glia zelulak.
Neuronak
| Neurona |
|---|
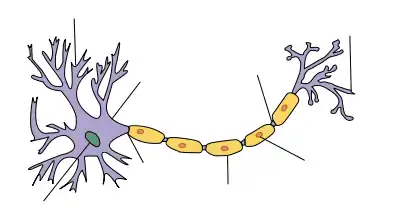
Nerbio sistema zelula berezi baten presentziaz dago definiturik, neurona (batzuetan nerbio zelula ere deitua). Neuronak beste zeluletatik hainbat eratara bereiz daitezke, baina beste zelulekin sinapsi bidez komunikatzea da hauen propietaterik ohikoena, seinale molekularren transmisio azkarra, bai elektriko eta bai kimikoak, ahalbidetzen duten tresneria molekularra duten mintz-mintz bidezko lotailuak direnak[4]. Neurona mota askok axoia dute, protrusio protoplasmiko bat da, gorputzaren alde urrunetara hedatu eta milaka kontaktu sinaptiko egin ditzakeena[12]; axoiak gorputz osoan zehar hedatzen dira nerbioak izeneko fardeletan.
Gizakiok bezalako espezie bakarraren nerbio sisteman ere, ehunka neurona mota ezberdin daude, hainbat morfologia eta funtziorekin[12]. Hauek estimulu fisikoak eraldatzen dituzten neurona sentsorialak barne hartzen dituzte, hala nola, argia eta soinua seinale neuronaletara eraldatzen dituztenak, baita neurona motorrak ere, seinale neuronalak muskulu edo guruinen aktibaziora eraldatzen dituztenak. Hala ere, espezie askotan neurona gehienek egitura zentralizatuak osatzean parte hartzen dute (garuna eta ganglioak), eta beste neuronetatik jasotzen dute beraien sarrera mezu guztiak zein neuronetara bidaltzen dituzte irteerak[4].
Glia zelulak

Glia zelulak (“kola” hitzerako grezierako hitzetik izendatua) zelula ez-neuronalak dira. Euren funtzioen artean daude neuronei euskarri eta elikadura ematea, homeostasia mantentzea, mielina eratzea eta nerbio sisteman seinaleak transmititzerakoan parte hartzea[13]. Gizakion burmuinean, glia zelula kopurua neuronenaren berdina dela estimatzen da gutxi gorabehera, nahiz eta burmuinaren eremu ezberdinetan proportzioak aldatu[14]. Glia zelulen funtzio garrantzitsuenen artean hauek daude: neuronei eutsi eta bertan eustea, neuronei elikagaiak ematea, neuronak elektrikoki isolatzea, patogenoak suntsitu eta hildako neuronak deuseztatzea eta neuronen axoiak beraien helburura gidatzea. Glia zelula mota oso garrantzitsu batek (oligodendrozitoak nerbio sistema zentralean eta Schwann zelulak nerbio sistema periferikoan) mielina izeneko substantzia koipetsu baten geruzak sortzen ditu, axoiak inguratzen dituena eta isolamendu elektrikoa ematen dietena, ekintza potentziala askoz azkarrago eta eraginkorrago transmititzea ahalbidetzen diena. Aurkikuntza berriek, glia zelulek, hala nola, mikrogliak eta astrozitoak, nerbio sistema zentralean zelula immunologiko garrantzitsu gisa balio dutela adierazten dute[15][16].
Anatomia ornodunetan
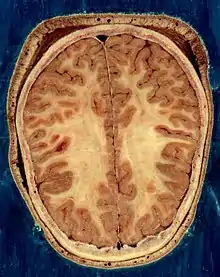
Ornodunen nerbio sistema (gizakiena barne), nerbio-sistema zentral (NSZ) eta nerbio-sistema periferikoan (NSP) banatzen da[4].
Nerbio-sistema zentrala burmuina eta bizkarrezur muinean dago banatuta[4]. Bizkarrezurraren kanalean bizkarrezur muina dago, garezurrak, aldiz, burmuina du. Nerbio-sistema zentrala meningeez itxita eta babestuta dago, hiru geruzako mintz sistema bat, kanpo geruza gogor bat barne, dura mater izenekoa. Burmuina ere garezurraren bidez dago babestuta, eta bizkarrezur muina ornoez.
Nerbio-sistema periferikoa (NSP) nerbio sistema zentralaren barnean ez dauden nerbio sistemako egiturei dagokien termino kolektibo bat da[17]. Nerbio deritzen axoi fardel gehiengoa nerbio sistema periferikoaren parte direla kontsideratzen da, baita hauei dagozkien neuronen zelula-gorputzak garunean edo bizkarrezur muinean daudenean. NSP zati somatiko eta erraietako-zatietan dago banatuta. Zati somatikoa larruazal, artikulazio eta muskuluetatik inerbatzen duten nerbioek osatzen duten. Neurona somatikoen zelula-gorputzak bizkarrezur muineko bizkarrezurreko gongoilean daude. Erraiei dagokion zatiak, nerbio-sistema autonomo gisa ere ezagutzen dena, barne organo, odol hodi edo guruinak inerbatzen dituen neuronez dago osatuta. Nerbio sistema autonomoak bi zati ditu: nerbio-sistema sinpatikoa eta nerbio-sistema parasinpatikoa. Autore batzuek neurona sentsorialak ere hartzen dituzte, hauen zelula gorputzak periferian daudelarik (entzumena bezalako zentzuentzat) NSPren zati gisa; beste batzuek, ordea, alde batera uzten dituzte[18].
Ornodunen nerbio sistema materia grisa eta materia zuria izeneko eremuetan banatu daiteke[19]. Materia grisa (ehun kontserbatuetan bakarrik da grisa, eta ehun bizietan hobeto deskribatzen da arrosa edo marroi arin gisa) neuronen zelula-gorputzen proportzio handia du. Materia zuria batez ere mielinaz inguratutako axoiez osatuta dago, eta bere kolorea mielinatik hartzen du. Materia zuriak nerbio guztiak hartzen ditu barne, eta garun eta bizkarrezur min barneko asko. Materia grisa garunean eta bizkarrezur muinean dauden neurona multzoetan aurkitzen da, eta euren gainazaleko geruza kortikaletan. Konbentzioz, neurona multzo bat burmuinean edo bizkarrezurrean badago nukleo deritzo eta periferian badago, ganglio. Hala ere, arau horren salbuespen batzuk daude, batez ere garunean dauden ganglio basaletan[20].
Anatomia konparatiboa eta eboluzioa
Neuronen aitzindariak belakietan
Belakiek ez dute sinapsi bidez konektaturiko zelularik, hau da, ez dute neuronarik eta, beraz, ez dute nerbio sistemarik. Hala ere, funtzio sinaptikoetan funtsezko paperak betetzen dituzten gene askoren homologoak dituzte. Ikerketa berriek frogatu dute belakien zelulek elkarrekin biltzen diren proteina multzo bat adierazten dutela, dentsitate postsinaptiko (sinapsi baten zati seinale-hartzailea) bat gogorarazten duen egitura bat osatuz. Hala eta guztiz ere, egitura horren funtzioa ez dago argi gaur egun[8]. Nahiz eta belakien zelulek transmisio sinaptikorik ez erakutsi, kaltzio uhin eta beste bulkada batzuen bidez elkarrekin komunikatzen dira, gorputz osoaren kontrakzioak bezalako ekintza sinpleak burutzen dituztenak[21].
Radiata
Marmokek, ktenoforoak, eta erlazionatutako animaliek nerbio sare zabalduak dituzte nerbio sistema baten ordez. Marmoka gehienetan nerbio sarea gutxi gorabehera berdintsu hedatzen da gorputzean zehar, ktenoforoetan ahotik gertu dago kontzentratuta. Nerbio sareak hiru neurona motaz daude osatuak: neurona sentsorialek seinale kimiko, taktiko eta bisualak igortzen dituzte; neurona motorrek, aldiz, gorputz osoaren kontrakzioa aktibatzen dute; eta interneuronak, neurona sentsorialen jarduera ereduak detektatzen dituztenak eta neurona motorren multzoei seinaleak bidaltzen dituztenak erantzun gisa. Kasu batzuetan, interneurona taldeak ganglio diskretuan biltzen dira[9].
Radiatan, nerbio sistemaren bilakaera ez dago erlatiboki egituratuta. Bilateralek ez bezala, radiatek bi funtsezko zelula bakarrik ditu: endodermoa eta ektodermoa. Neuronak ektodermoko zelula aitzindari multzo berezi batetik sortzen dira, beste zelula guztien aitzindari bezala ere balio dutenak[22].
Bilateria

Gaur egungo animalia gehienak bilateralak dira, hots, esker eta eskuin aldeko eremuak dituzten animaliak. Bilateral guztiak Ediacarar garaian (duela 550-600 milioi urte) bizi izan zen arbaso komun batetik eratorriak direla uste da[10]. Bilateralen oinarrizko gorputzaren forma hodi bat da barruko zulo batekin, ahotik uzkiraino, eta nerbio-soka bat du gorputzaren zati bakoitzean handitzen dena, ganglio bat. Aurrealdean, bereziki luzea den ganglio bat du, burmuin izenekoa.
Ugaztunek ere, gizakiak barne, gorputz bilateralaren segmentazioa dute nerbio sistema milan. Bizkarrezur-muinak ganglio segmentario batzuk ditu, eta horietako bakoitzak nerbio motorrak eta sentsorialak sortzen ditu, gorputz-azaleraren eta azpiko muskulaturaren zati bat inerbatzen dutenak. Gorputz-adarretan, zaila da inerbazio-patroiaren kokapena jarraitzea, baina enborrean zerrenda estu batzuk sortzen ditu. Goiko hiru segmentuak burmuinari dagozkio, aurreko garuna, erdiko garuna eta ondorengo garuna sortuz[23].
Bilateralak, enbrioi-garapenean oso goiz gertatzen diren gertakarietan oinarrituta, protostomio eta deuterostomio izeneko bi taldetan bana daitezke. Deuterostomioak ornodunak, ekinodermatuak, hemikordatuak eta xenoturbellak hartzen ditu barne[24]. Protostomioak, talderik handienak, artropodoak, moluskuak eta har mota ugari hartzen ditu. Oinarrizko desberdintasun bat dago bi taldeen artean, gorputz barneko nerbio-sistemaren posizioan hain zuzen ere: protostomioek nerbio-soka bat dute gorputzaren alde bakoitzean (normalean beheko aldean); deuterostomiotan, berriz, nerbio-soka dortsalean dago (normalean goiko aldean). Izan ere, gorputzaren alde asko alderantzikatu egiten dira bi taldeen artean, dortsaletik abdomenerako gradienteak erakusten dituzten zenbait generen adierazpen-ereduak barne. Anatomista gehienek, gaur egun, protostomio eta deuterostomioen gorputzak elkar "alderantzizkatuak" direla kontsideratzen dute, Geoffroy Saint-Hilairek hasieran intsektuentzat proposatutako hipotesia ornodunetan aplikaturik. Intsektuek, adibidez, nerbio-sokak izaten dituzte gorputzaren erdiko lerroan zehar, ornodun guztiek, berriz, bizkarrezur muina dute, dortsal-erdiko lerroan zehar[25].
Zizareak eta nematodoak

Anelidoak animalia bilateralik sinpleenak dira, eta bilateralen nerbio-sistemaren oinarrizko egitura adierazten dute modurik zuzenenean. Adibide gisa, lurreko zizareek nerbio-soka dualak dituzte gorputzaren luzeran zehar, isatsarekin eta ahoarekin bat eginez. Nerbio-soka hauek nerbio transbertsalez lotuta daude, eskailera baten errenkadak bezala. Nerbio transbertsal horiek animaliaren bi aldeak koordinatzen laguntzen dute. Buruan dauden bi ganglioek ("nerbio eraztuna") garun sinple baten antzeko funtzioa betetzen du. Animaliaren begien posizioko fotoerrezeptoreek argiaren eta iluntasunaren inguruko informazioa sentsoriala ematen dute[26].
Har txiki biribil baten nerbio-sistema, Caenorhabditis elegans nematodoa, erabat deskribatua izan da konektoma batean, bere sinapsiak barne. Neurona bakoitza eta bere zelula-lerroa eta neuronen konexio gehienak, ez badira guztiak, ezagutzen dira. Espezie honetan, nerbio-sistema sexualki dimorfikoa da; bi sexuetako nerbio-sistemek, arrek eta eme hermafroditak, neurona eta neurona talde kopuru ezberdinak dituzte, funtzio sexuarekiko espezifikoak diren funtzioak betetzen dituztenak. C. elegansetan, arrek 383 neurona dituzte, hermafroditek, berriz, 302 neurona[27].
Artropodoak
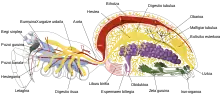
Artropodoek, hala nola intsektuek eta krustazeoek, sabelaren luzeran zehar doazen nerbio-soka paraleloz konektatutako ganglioez osatutako nerbio-sistema bat dute[28]. Egiturarik ohikoenean, gorputz segmentu bakoitzak ganglio bat du alde bakoitzean, nahiz eta ganglio batzuek garuna edo beste ganglio handi batzuk osatzeko bat egiten duten. Burualdeko segmentua garuna da, eta ganglio supraesofageala bezala ere ezagutzen da. Intsektuen nerbio-sisteman, garuna anatomikoki banatzen da protogaruna, deutogaruna eta tritogarunaren artean. Garunaren atzean ganglio subesofageoa dago, hiru ganglio pare fusionatuz osatua dagoena. Ahoa, listu-guruinak eta muskulu jakin batzuk kontrolatzen ditu. Artropodo askok ongi garaturiko organo sentsorialak dituzte, ikusmenerako begi konposatuak, usaimenerako antenak eta sentsazioetarako feromonak barne. Organo hauetako informazio sentsoriala garunak prozesatzen du.
Intsektuetan, neurona askok garunaren ertzean kokatuta dauden eta elektrikoki pasiboak diren zelula-gorputzak dituzte. Zelula-gorputzek laguntza metabolikoa emateko bakarrik balio dute, eta ez dute seinaleztapenean parte hartzen. Zuntz protoplasmiko bat doa oparotasunez zelula gorputz eta adarretatik, zati batzuk seinaleak igortzen eta beste batzuk seinaleak jasotzen dituztelarik. Honela, intsektu-garunaren zati gehienek zelula-gorputz pasiboak dituzte periferian antolatuta, eta seinale neuronalen prozesamendua, berriz, neuropilo izeneko zuntz protoplasmatikoen multzo batean gertatzen da, barnealdean kokatua[28].
Neurona "identifikatuak"
Neurona bat identifikatua deritze animalia bereko beste neurona guztietatik bereizten duten propietateak baldin baditu, — hala nola, kokalekua, neurotransmisorea, gene adierazpen patroia eta konexioa — eta espezie bereko banako bakoitzak propietate multzo bereko neurona bat eta bakarra baldin badu[29]. Ornodunetan, nerbio-sistema neurona gutxi dira "identifikatuak" —gizakietan ez dagoela pentsatzen da— nerbio sistema sinpleagoetan, ordea, neurona batzuk edo guztiak dira mota horretakoak. C. elegens nematodoan, zeinaren nerbio-sistema beste edozein animaliena baino deskribatuagoa dagoen, gorputzeko neurona guztiak dira bereizgarri, kokapen eta lotura berberekin zizare banako guztietan. Honen ondorio nabarmenetako bat, C.elegansen nerbio-sistemaren forma erabat determinatuta dagoela da, genomaren bidez, esperientzietan oinarritutako plastikotasunik gabe[27].
Molusku eta intsektu askoren garunek identifikatutako neurona kopuru handiak ere badituzte[29]. Ornodunetan, gehien ezagutzen diren neuronak Mauthner arrain zelula erraldoiak dira[30]. Arrain guztiek bi Mauthner zelula dituzte, garunaren behealdean, bat ezkerraldean eta bestea eskuinaldean. Mauthner zelula bakoitzak zeharkatzen duen axoi bat du, garuneko maila berean neuronak inerbatzen, eta gero bizkarrezur muinetik behera jaisten dena, lotura ugari eginez. Mauthner zelulek sortzen dituen sinapsiak hain dira indartsuak, non potentzial bakar batek portaera erantzun nagusi bat sortzen duen: milisegundoetan arrainak bere gorputza C forman kurbatzen du, gero zuzen jartzen da, eta horrela azkar egiten du aurrera[31][32]. Hau, funtzionalki, ihesaldi azkarreko erantzun bat da, arrainaren albo-organoari eragiten dion soinu uhin edo presio-uhin indartsu batek erraz kitzikatzen duena. Mauthner zelulak ez dira arrainen artean identifikatutako neurona bakarrak — 20 bat mota gehiago daude, bizkarrezurraren nukleo segmental bakoitzean "Mauthner zelula analogo" pare batzuk barne. Nahiz eta Mauthner zelula batek ihes-erantzun indibiduala emateko ahalmena duen, portaera arruntaren esparruan beste zelula mota batzuek erantzunaren anplitudea eta norabidea eratzen laguntzen dute.
Mauthner zelulak agindu neurona gisa izan dira deskribatuak[30]. Agindu neurona bat, neurona identifikatu mota berezi bat da, jokabide zehatz bat indibidualki gidatzeko gai den neurona gisa definitua. Neurona horiek espezie ezberdinen ihes azkarreko sistemetan agertzen dira gehienetan: txipiroiaren axoi erraldoia eta sinapsi erraldoia, neurofisiologia esperimentu aitzindariak egiteko erabiltzen dena euren tamaina izugarriagatik, biek txipiroiaren ihesaldi azkarreko zirkuituan parte hartzen dutelarik. Hala ere, agindu neurona kontzeptua eztabaidagarri bihurtu da, hasieran deskribapenari egokitzen zitzaizkiola ematen zuten neurona batzuk egoera multzo mugatu batean bakarrik erantzun zezaketela erakusten duten ikerketengatik[33].
Funtzioa
Oinarrizko mailan, nerbio-sistemaren funtzioa zelula batetik bestera edo gorputzaren zati batetik bestera seinaleak bidaltzea da. Zelula batek beste zelula batzuetara seinaleak bidaltzeko modu asko daude. Horietako bat, hormonak deitzen diren substantzia kimikoak barne zirkulazioan jartzean datza, urruneko lekuetara zabal daitezen. Nerbio-sistemak "puntutik-punturako" seinaleak ditu "seinaleztatzeko" modu horren aurrean — neuronek beren axoiak proiektatzen dituzte itu-eremu zehatzetara eta lotura sinaptikoak egiten dituzte itu-zelula zehatzekin[34]. Nerbio seinaleztapen hori hormona seinaleztapen normala baino askoz espezifikoagoa da. Askoz bizkorragoa ere bada: nerbio-seinalerik azkarrenek segundoko 100 metro gainditzen dituzten abiaduretan bidaiatzen dute.
Maila inklusiboago batean, nerbio-sistemaren funtzio nagusia gorputza kontrolatzea da[4]. Inguruneko informazioa biltzen du hartzaile sentsorialak erabiliz, informazio hori kodetuz nerbio-sistema zentralera seinaleak bidaliz, informazioa prozesatuz erantzun egoki bat zehazteko, eta kanpoko seinaleak gihar edo guruinetara bidaliz erantzuna aktibatzeko. Nerbio sistema konplexu baten eboluzioak animali espezie ezberdinek pertzepzio gaitasun aurreratuak izatea ahalbidetu du, hala nola ikusmena, elkarrekintza sozial konplexuak, organo sistemen koordinazio azkarra eta seinale bateratuen prozesu integratua. Gizakietan, nerbio-sistemaren sofistikazioak hizkuntza izatea ahalbidetzen du eta, kontzeptuen irudikapen abstraktua, kulturaren transmisioa, eta gizartearen beste ezaugarri asko, giza garunik gabe existituko ez liratekeenak.
Neuronak eta sinapsia
| Sinapsi kimiko baten ohiko egiturak. |
|---|
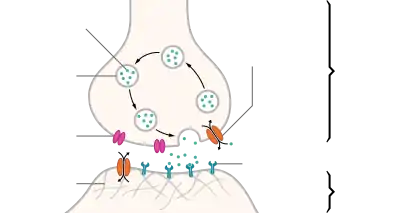
Dentsitate
postsinaptikoa Potentzial-
diferentzia Ca++ kanala Besikula
sinaptiko Neurotransmisoreen
garraiatzailea Errezeptorea
Axoi terminal
Arrail sinaptiko
|
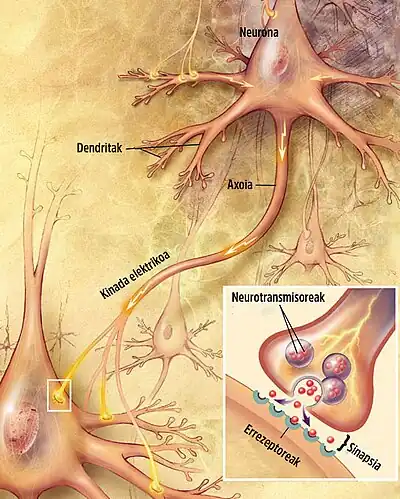
Neurona gehienek axoiaren bidez seinaleak bidaltzen dituzte, nahiz eta zenbait motatako neuronak dentrita-dentrita motako komunikaziorako gai diren. (Izan ere, zelula amakrina deituriko neurona motek ez dute axoirik, eta beren dendriten bidez bakarrik komunikatu daitezke). Seinale-neuronalak axoi batean zehar barreiatzen dira ekintza-potentzial deituriko uhin elektrokimiko forman, zeinak zelula-zelula seinaleak sortzen dituzten, axoi terminalek beste zelula batzuekin kontaktu sinaptikoa egiten duten puntuetan[4]. Sinapsiak elektrikoak edo kimikoak izan daitezke. Sinapsi elektrikoek neuronen arteko zuzeneko konexio elektrikoak egiten dituzte[35], baina sinapsi kimikoak askoz ohikoagoak dira, eta funtzio ugariagoak dituzte[36]. Sinapsi kimiko batean, seinaleak igortzen dituen zelulari presinaptikoa deitzen zaio, eta seinaleak jasotzen dituen zelulari postsinaptikoa. Bai presinaptikoak, bai postsinaptikoak, makineria molekularrez beteta daude seinaleztapen prozesua burutzen duena. Eremu presinaptikoan odol-hodi esferiko txiki ugari daude, besikula sinaptiko deritzenak, neurotransmisore kimikoz beteak.[4] Terminal presinaptikoa elektrikoki estimulatzen denean, mintzean txertatutako molekula multzo bat aktibatu egiten da, eta besikulen edukia askatzea eragiten du presinaptiko eta postsinaptiko arteko espazio estuan, arrail sinaptikoa deiturikoa. Neurotransmisorea, orduan, mintz postsinaptikoan txertatutako hartzaileekin lotzen da, hauek aktibatuz[4]. Hartzaile motaren arabera, zelula postsinaptikoan eragindako efektua hau kitzikatzea, inhibitzea edo modulatze izan daiteke modu konplexuagoan. Adibidez, azetilkolina neurotransmisorearen askapenak neurona motor baten eta muskulu zelula baten arteko kontaktu sinaptikoa, eta muskulu zelularen kontrakzio azkarra eragiten du[4]. Transmisio sinaptiko prozesu osoak milisegundo baten frakzio bat baino ez du irauten, nahiz eta zelula postsinaptikoaren efektuek askoz gehiago iraun dezaketen (baita era mugagabe batean ere, seinale sinaptikoak oroimenera eramaten duen kasuetan)[4].
Ehunka sinapsi mota daude. Izan ere, ehun neurotransmisore ezagun baino gehiago daude, eta horietako askok hainbat hartzaile mota dituzte[4]. Sinapsi askok neurotransmisore bat baino gehiago erabiltzen dute — ohikoa da sinapsi batek eragin azkarreko neurotransmisore molekula bat erabiltzea, adibidez, glutamatoa edo GABA, eragin mantsoko neurotransmisore peptido bat edo gehiagorekin batera—. Neurozientzialari molekularrek, normalean, hartzaileak bi multzo zabaletan banatzen dituzte: kimikoki itxitako ioi-kanalak eta bigarren mezularitzako sistemak. Kimikoki itxitako ioi-kanal bat aktibatzen denean, mintzean zehar ioi-mota zehatz batzuk isurtzeko aukera ematen duen pasabidea irekitzen da. Ioi motaren arabera, itu-zelularen eragina hau kitzikatzea edo inhibitzea izango da. Bigarren mezularitza-sistema bat aktibatzen denean, itu-zelularen barruan elkarrekintza molekularren ur-jauzi bat abiarazten du, azkenean efektu konplexu anitz asko eragin ditzakeena, hala nola zelulak estimulatzeko duen sentikortasuna handitu edo murriztu, edo gene transkripzioa ere aldatzen duena.
Daleren printzipioa deritzon arauaren arabera, salbuespen ezagun batzuk besterik ez dituena, neurotransmisore batek bere sinapsi guztietan neurotransmisore bera askatzen du[37]. Honek ez du esan nahi neurona batek bere helburu guztietan eragin bera duenik, eragina ez baitago neurotransmisorearen menpe, bere hartzaileen menpe baizik[4]. Itu desberdinek hartzaile mota desberdinak erabili ditzaketelako (askotan gertatzen da), gerta daiteke neurona bat kitzikatua izatea zati batean, inhibitua beste batean eta konplexuki modulatua izatea beste zatietan. Hala ere gertatzen da, bi neurotransmisore erabilienek, glutamatoa eta GABA, ondorio oso sendoak edukitzea. Glutamatoak zenbait hartzaile mota ditu, baina guztiak kitzikakorrak edo modularrak dira. Era berean, GABAk zenbait hartzaile mota ditu, baina guztiak inhibitzaileak dira[38]. Hori dela eta zelula glutamatergikoei neurona kitzikatzaileak eta GABAergikoei neurona inhibitzaileak deritze. Hertsiki hitz eginez, terminologia hori gehiegikeria bat da — hartzaileak dira kitzikatzaileak eta inhibitzaileak, eta ez neuronak—, baina argitalpen akademikoetan ere ikusi ohi da.
Sinapsien azpimultzo oso garrantzitsu bat oroimenaren aztarnak sortzeko gai da, iraupen luzeko indar sinaptikoaren aldaketa batzuen ondorioz[39]. Oroimen neuronal forma ezagunena epe luzerako potentziazio (LTP laburtua) deritzon prozesu bat da, glutamato neurotransmisorea erabiltzen duten sinapsietan funtzionatzen duena, NMDA izeneko hartzaile berezi batean eragiten duena[40]. NMDAren hartzaileak propietate "asoziatzaile" bat du: sinapsian sartuta dauden bi zelulak aldi berean aktibatzen badira, kanal bat irekitzen da kaltzioa itu-zeluletara isurtzea baimentzen duena[41]. Indar aldaketa honek asteak edo gehiago iraun ditzake. 1973an LTP aurkitu zenetik, oroimen sinaptikoaren mota asko aurkitu dira, baldintza ezberdinek eragiten duten indar sinaptikoa gehitzea edo murriztea ekartzen duena, eta denbora aldakorrez irauten duena[40]. Sari sistema, adibidez, desiratutako portaera sendotzen duena, LPTaren aldaera baten menpe dago, dopamina neurotransmisorea erabiltzen duen sari-seinaleztapen baten sarrera estra batek baldintzatzen duena[42]. Aldakortasun sinaptikoaren forma guztiek, kolektiboki begiratuta, plastikotasun neuronala sortzen dute, hau da, nerbio-sistemak ingurumenean dituen aldaketetara egokitzeko gaitasuna.
Zirkuitu eta sistema neuralak
Beste zelula batzuetara seinaleak bidaltzearen oinarrizko funtzio neuronalak elkarren artean seinaleak trukatzeko gaitasun bat barne hartzen du. Neurona-talde interkonektatuek sortutako sareak funtzio ugari izan ditzakete, besteak beste, ezaugarriak detektatzea, ereduak sortzea eta sinkronizatzea[43], eta hainbat informazio prozesamendu mota posible daudela ikusi da. Warren McCullochek eta Walter Pittsek 1943an erakutsi zuten neurona baten abstrakzio matematiko oso sinplifikatu batetik sortutako neurona-sare artifizialak ere konputazio unibertsalerako gai direla[44].
Historikoki, urte askotan, nerbio-sistemaren funtzioaren ikuspegi nagusia estimulu-erantzunarekin lotu da[45]. Diseinu honetan, neurona sentsorialak aktibatzen dituzten estimuluekin hasten da prozesamendu neuronala, bizkarrezurraren eta garunaren konexio-kateen bidez ugaltzen diren seinaleak sortuz, azkenean neurona motorrak aktibatu eta muskuluen kontrakzioa sortuz. Descartesek uste zuen animalien portaera guztiak, eta gizakien portaera gehienak, estimulu-erantzun zirkuituen bidez azal zitezkeela, nahiz eta uste zuen hizkuntza bezalako funtzio kognitibo altuagoak ezin zirela mekanikoki azaldu[46]. Charles Sherringtonek, 1906ko bere liburuan, "The Integrative Action of the Nervous System"[45], estimulu-erantzun mekanismoen kontzeptua garatu zuen, eta konduktismoa, XX. mendean psikologia menperatzen zuen eskola, giza portaera guztia estimulu-erantzun mekanismoen bidez azaltzen saiatu zen[47].
Hala ere, elektrofisiologiaren ikerketa esperimentalek, XX. mendearen hasieran hasi eta 1940ko hamarkadan produktibitate altuenera iritsiz, nerbio-sistemak zelulen kitzikapena mantentzeko mekanismo asko dituela erakutsi zuten, kanpoko estimulurik behar izan gabe[48]. Neuronak ekintza-potentzialen sekuentzia erregularrak sortzeko gai direla ikusi zen, edo eztanda sekuentziak, guztiz isolatuta egonda ere[49]. Berez aktiboak diren neuronak zirkuitu konplexuetan elkarri lotuta daudenean, aldi baterako berezko eredu konplexuak sortzeko aukerak askoz ere zabalagoak bihurtzen dira[43]. Kontzepzio moderno batek nerbio-sistemaren funtzioa partzialki estimulu-erantzun kateetan ikusten du— bi jarduera-motek elkar eragiten dute portaeraren errepertorio osoa sortzeko[50].
Erreflexuak eta beste estimulu-erantzun mekanismoak
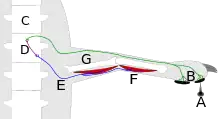
Zirkuitu neuronal mota sinpleena arku erreflexu bat da, sentsore sarrera batekin hasten dena eta motorraren irteera batekin amaitzen dena, seriean konektatutako neurona sekuentzia bat zeharkatuz[4]. Hau "erretiratzeko erreflexuetan" ikusi daiteke, esku batek berogailu bat ukitu ondoren atzerantz egitea, adibidez. Zirkuitua larruazalaren hartzaile sentsorialekin hasten da, bero-maila kaltegarriek aktibatzen dutena: mintz barruan txertatutako egitura molekular mota berezi batek beroa eragiten du mintzean zeharreko eremu elektrikoa aldatzeko. Potentzial elektrikoaren aldaketa emandako atalasea pasatzeko bezain handia bada, ekintza potentzial bat sortzen du, axoiaren ardatzean zehar bizkarrezur muinera igortzen dena. Bertan axoiak kontaktu sinaptiko deigarriak egiten ditu beste zelula batzuekin, horietako batzuk bizkarrezur muinaren alde berera, eta beste batzuk garunera. Helburu bat bizkarrezurreko interneurona multzo bat da, besoko giharrak kontrolatzen dituzten neurona motorretarako proiektatzen dena. Interneuronek neurona motorrak kitzikatzen dituzte, eta kitzikapena nahiko indartsua bada, zenbait neurona motorrek akzio-potentzialak sortzen dituzte, beren ardatzetan behera doazenak muskulu-zelulekin kontaktu sinaptiko kitzikakorrak egiten dituzten punturaino. Seinale kitzikatzaileek muskulu-zelulen kontrakzioa eragiten dute, eta horrek besoko angelu bateratuak aldatzea eragiten du.
Errealitatean, eskema hau konplikazio ugariren menpe dago[4]. Erreflexu sinpleenentzat neurona sentsorialetatik neurona motorrerako bide laburrak dauden arren, zirkuituan parte hartzen duten inguruko beste neurona batzuk ere badaude eta erantzuna modulatzen dute. Gainera, garunetik bizkarrezur muinera erreflexua hobetu edo oztopatzeko gai diren proiekzioak daude.
Nahiz eta erreflexurik sinpleenak bizkarrezur muinaren barruan erabat dauden zirkuituekin egin daitezkeen, erantzun konplexuagoak garuneko seinale-prozesamenduan oinarritzen dira[4]. Adibidez, ikusmen eremuaren periferiako objektu bat mugitzen denean eta pertsona batek hari begiratzen dionean seinale prozesuko fase asko hasten dira. Hasierako erantzun sentsoriala, begiaren erretinan, eta azken erantzun motorra, garun-enborraren nukleo okulomotorrean, ez dira erreflexu sinple batean daudenetatik ezberdinak, baizik eta bitarteko faseak erabat ezberdinak dira. Prozesatze kate bat edo biren ordez, ikusizko seinaleak dozena bat integrazio fasetatik igarotzen dira, eta horien artean talamoa, garunaren kortexa, ganglio basala, goiko kolikulua, zerebeloa eta zenbait enbor entzefalikoko nukleo daude. Gune hauek seinale prozesamendu funtzioak betetzen dituzte, besteak beste, ezaugarri detekzioa, pertzepzioaren analisia, oroitzapena aktibatzea, erabakiak hartzea eta planifikazio motorra[4].
Ezaugarri detekzioa seinale sentsorialen konbinazioetatik biologikoki esanguratsua den informazioa ateratzeko gaitasuna da[4]. Ikusmen-sisteman, adibidez, begiaren erretinan dauden hartzaile sentsorialak indibidualki "argi-puntuak" detektatzeko gai dira. Bigarren mailako ikusmen-neuronak lehen mailako hartzaileen multzoen sarrera jasotzen dituzte, maila goreneko neuronak bigarren mailako neuronen sarrerak eta horrela, prozesu hierarkiko bat sortzen da. Une bakoitzean, informazio garrantzitsua ateratzen da seinaleetatik eta garrantzirik gabeko informazioa baztertzen da. Prozesuaren amaierarako, "argi-puntuak" irudikatzen dituzten seinaleak, inguruko objektuen eta horien ezaugarrien irudikapen neuronal bihurtu dira. Zentzumenen prozesatze sofistikatuena garunaren barruan gertatzen da, baina ezaugarri konplexuak ere gertatzen dira bizkarrezur muinean eta erretina bezalako organo sentsorial periferikoetan[4].
Eredu intrintsekoen sorrera
Nahiz eta estimuluei erantzuteko mekanismoak errazenak izan ulertzeko, nerbio-sistema ere gai da gorputza kontrolatu ahal izateko kanpoko estimuluen beharrik gabe, barneko aktibitate erritmoen bidez. Neurona baten mintzean txertatu daitezken boltajearekiko sentikorrak diren ioi-kanalen barietatea dela eta, neurona mota asko gai dira, baita isolamenduan ere, akzio potentzialen sekuentzia erritmikoak sortzeko, edo kargarik handiena eta kieszentziaren arteko txandakatze erritmikoak egiteko. Erritmo intrintsekoa duten neuronak elkarri konektaturik daude sinapsi kitzikatzaile edo inhibitzaile bidez, sortzen diren sareak hainbat elementu dinamiko sortzeko gai dira, besteak beste, dinamika atraktorea, aldizkakotasuna eta kaosa. Neurona-sare bat, bere barne egitura aldi baterako egiturak sortzeko erabiltzen duena, aldi baterako egitura estimulu bat behar izan gabe, eredu zentraleko sorgailu bat deritze.
Barne-ereduko sortzaileak denbora-eskala zabal batean eragiten du, milisegundoetatik orduetara edo luzeagotara. Ordu-patroirik garrantzitsuenetako bat erritmo zirkadianoa da, hau da, 24 ordu inguruko erritmoak. Aztertutako animalia guztiek fluktuazio zirkadianoak erakusten dituzte aktibitate neuronalean, alterazio zirkadianoak kontrolatzen dituztenak lo-esnatu zikloaren antzera. 1990eko hamarkadatik datozen ikerketa esperimentalek frogatu dute erritmo zirkadianoak "erloju genetiko" batek sortuak direla, gene multzo berezi batek osatuak, zeinen espresio maila gora eta behera egiten duen egunean zehar. Intsektuak eta ornodunak bezalako animaliek antzeko erloju genetikoa partekatzen dute. Erloju zirkadianoetan argiak eragina du, baina funtzionatzen jarraitzen du, baita argi maila konstantea izan eta beste ordu-tarterik ez dagoenean ere. Erloju geneak nerbio-sistemako atal askotan adierazten dira, baita organo periferiko askotan ere, baina ugaztunetan, "ehunezko erloju" horiek guztiak sinkronian mantentzen dira, erlojugile nagusi batengandik nukleo suprakiasmatiko deritzon garunaren zati txiki batean sortzen diren seinaleen bidez.
Ispilu neuronak
Ispilu neuronak animalia batek zerbait egin eta ekintza bera beste animalia batek egiten ikusten duenean pizten den neurona da[51][52][53]. Horrela, neuronak bestearen portaera islatzen du, behatzaileak berak balu bezala. Primate espezieetan behatu dira zuzenean neurona hauek[54]. Hegaztiek portaera imitatzailea dutela frogatu da, eta ebidentzia neurologikoak ispilu-sistemaren nolabaiteko presentzia iradokitzen du[55]. Gizakietan, ispilu neuronekin bat datorren burmuinaren aktibitatea aurkitu da kortex premotorrean, eremu osagarri motorrean, kortex somatosentsorial primarioan eta behe kortex parietalean[56]. Ispilu sistemaren funtzioa espekulazio askoko gaia da. Neurozientzia kognitibo eta psikologia kognitiboko ikertzaile askok uste dute sistema honek mekanismo fisiologikoa eskaintzen duela pertzepzioa/ekintza erlazionatzeko. Ispilu-neuronak garrantzitsuak izan daitezkeela eztabaidatzen dute beste pertsonen ekintzak ulertzeko eta imitazioaren bidez trebetasun berriak ikasteko. Ikertzaile batzuek, gainera, ispilu sistemek ikusitako ekintzak simulatu ditzaketela uste dute, eta, horrela, adimen-trebetasunen teoriari laguntzen diote; eta beste batzuek, aldiz, ispilu neuronak hizkuntza-gaitasunekin erlazionatzen dituzte[57]. Hala ere, gaur egun ez da oso onartutako konputazio eredu edo eredu neuronalik aurkeztu ispilu neuronak imitazioa bezalako funtzio kognitiboei nola erlazionatzen diren deskribatzeko[58]. Badaude, neurona ispiluei eman zaien papera ikerketa egokietan oinarritzen ez dela esaten duten neurozientzialariak[59].
Garapena

Ornodunetan, enbrioi-garapen neuronalaren ezaugarri nagusiak hauek dira: neuronen jaiotza eta desberdintzapena ama zeluletatik, neurona heldugabeen migrazioa jaiotzatik enbrioiaren azken posiziorantz, axoiak neuronetatik irtetea eta hazkunde motorraren gidaritza enbrioiaren bidez bikote postsinaptikorantz, axoien eta haien bikote postsinaptikoen arteko sinapsiaren sorrera eta, azkenik, sinapsietan behin-betiko aldaketak uste dena oroimena eta ikasteko ahalmena ematen dutela[4].
Garapen fase goiztiarrean dauden animalia bilateral guztiek gastrula bat osatzen dute, polarizatua dagoena, mutur bati animalia-poloa eta besteari polo-begetala deritzona. Gastrulak hiru zelula-geruza dituen disko baten forma du, endodermoa izeneko barne geruza, barne organo gehienen lerrokatzea eragiten duena, mesodermoa izeneko erdiko geruza, hezur eta muskuluak sortzen dituena, eta ektodermoa izeneko kanpoko geruza, azala eta nerbio-sistema sortzen dituena[60].
Ornodunetan, nerbio-sistemaren lehen ezaugarria bizkar erdian nerbio-xafla izeneko zelula xingola mehe bat agertzea da, plaka neuronala deiturikoa. Plaka neuronalaren barruko zatia (erdialdetik) nerbio-sistema zentrala (NSZ) izatera bideratua dago, kanpoaldea, berriz, nerbio-sistema periferikoa (NSP). Garapena aurrera doan heinean, neurona ildoa deritzon tolestura bat agertzen da erdialdean. Tolesdura sakonagoa egiten da, eta gero goian itxi egiten da. Une honetan, NSZ izango dena agertzen da, hodi neuronala deritzon egitura zilindriko bat agertzean eratzen da, eta NSP izango dena, aldiz, gailur neuronala deritzon ehunezko bi xingola agertzen sortzen da, hodi neuronalaren gainetik luzetara doana. Plaka neuronaletik hodi neuronal eta gailur neuronalerako faseen sekuentzia, neurulazioa bezala ezagutzen da[61].
XX. mende hasieran, Hans Spemann eta Hilde Mangoldek egindako esperimentu famatu batzuek erakutsi zuten nerbio-ehunaren formazioa gune antolatzailea izeneko zelula mesodermal talde baten seinaleen bidez "induzitua" dela[4]. Hala ere, hamarkada askotan, indukzio neuronalaren izaerak hura asmatzen saiatu ziren guztiak garaitu zituen, azkenean 1990eko hamarkadako hurbilketa genetikoek ebatzi zuten arte. Ehun neuronalak induzitzeak genea inhibitzea eskatzen du hezur proteina morfogenetikoa edo BMP izeneko batentzat. Bereziki BMP4 proteina tartean dagoela dirudi. Noggin eta Chordin izeneko bi proteinak, biak mesodermoak jariatuak, BMP4 detektatzeko gai dira, eta, beraz, ektodermoa ehun neuronal bihurtzeko gai dira. Antza denez, antzeko mekanismo molekular bat dago animalia mota oso desberdinetan, artropodoak nahiz ornodunak barne. Animalia batzuetan, hala ere, Fibroblastoaren Hazkuntza Faktore edo FGF izeneko molekula mota batek ere rol garrantzitsua joka dezake indukzioan[62].
Nerbio ehunen indukzioak nerbio zelulen aitzindarien eraketa eragiten du, neuroblasto deiturikoa[4]. Drosophiletan, neuroblastoak asimetrikoki banatzen dira, produktu bat ganglio ama zelula bat (GMC), eta bestea neuroblasto bat izanik. GMC bat behin bakarrik banatzen da, neurona pare bat edo glia zelula pare bat sortzeko. Azken finean, neuroblasto bat neurona edo glia kopuru zehaztugabe bat sortzeko gai da.
2008ko ikerketa batean azaltzen denez, organismo bilateral guztien (gizakiak barne) artean dagoen faktore bat jariatuak diren seinaleztapen molekula familia bat da, neutrofilo izenekoak, neuronen hazkuntza eta biziraupena arautzen dutenak[63]. Ikerketa horretan DNT1 identifikatu zuten, eulietan aurkitutako lehenengo neutrofiloa. DNT 1-ek neutrofilo ezagun guztiekin ezaugarri estrukturalak partekatzen ditu eta Drosophilaren neuronen patuaren giltzarri diren faktoreetako bat da. Gaur egun ornodunetan zein ornogabeetan neurotrofiloak identifikatu dira, ebidentzia honek adierazten du neurotrofiloak organismo bilateralen arbaso komun batean egon zirela eta nerbio-sistema eratzeko mekanismo komun bat izan daitekeela.
Patologia

Nerbio-sistema zentrala hesi fisiko eta kimiko garrantzitsuek babesten dute. Fisikoki, garuna eta bizkarrezur muina mintz meningeo gogorrez inguratuta daude, eta garezurreko eta bizkarrezurreko ornoetako hezurretan sartuta, babes fisiko indartsu bat osatzeko elkartzen direnak. Kimikoki, garuna eta bizkarrezurra isolatuta daude muga hematoentzefalikoaren bidez, eta horrek eragotzi egiten ditu substantzia kimiko mota gehienak odol-fluxutik NSZ-ren barnealdera mugitzea. Babes hauek NSZak NSP-ak baino sentikortasun gutxiago izatea eragiten dute, baina NSZko kalteek ondorio larriagoak izateko joera dute.
Nahiz eta nerbioak larruazalaren azpian sakon egoteko joera duten, nerbio kubitala bezalako leku batzuetan izan ezik, kalte fisikoa jasateko aukera duten, eta horrek mina, sentsazio galera edo muskuluen kontrola galtzea ekar dezake. Nerbioetako kalteak hantura edo ubeldurek ere eragin ditzakete, nerbio bat ubide hezurtsu estu batetik pasatzen den lekuetan, tunel karpianoaren sindromean gertatzen den bezala. Nerbio bat erabat moztuta badago, askotan erregeneratu egingo da, baina nerbio luzeetarako hilabeteak beharko dira prozesu hau osatzeko. Kalte fisikoez gain, beste arazo mediko askok ere eragin dezakete neuropatia periferikoa, besteak beste, diabetesa bezalako baldintza genetikoek, baldintza metabolikoek, Guillain-Barréren sindromeak, bitamina faltak, legenarrak edo zoster herpesa bezalako gaixotasun infekziosoek, edo toxinen pozoitzeak, eta metal astunek. Kasu askok ez dute identifikatu daitekeen kausarik, eta idiopatia deitzen zaie. Nerbioek ere anestesiarekin galdu dezakete euren funtzioa, adibidez lidokainarekin.
Bizkarrezur muinaren kalte fisikoak sentsazio edo mugimendu galera eragin dezake. Bizkarrezurrean lesio batek hantura baino okerragorik sortzen ez badu, sintomak transistoreak izan daitezke, baina bizkarrezurraren nerbio zuntzak benetan suntsitzen badira, funtzioaren galera behin betikoa izan ohi da. Ikerketa esperimentalek frogatu dute bizkarrezurreko nerbio zuntzak berriz hazten saiatzen direla, baina bizkarrezur muinaren ehunen suntsipenak orbainak sortu ohi ditu, berriz hazten ari diren nerbioek zeharkatu ezin dutena.
Gaixotasun neurologikoak
Gaixotasun neurologikoak nerbio-sistema zentralaren gaixotasunak deskribatzeko erabilitako kontzeptua da, garunak, bizkarrezur muinak edo nerbioek jasandako kalte fisikoak eraginda eta, NSZ edo NSP-ri eragin diezaioketenak. Beste kausa batzuk alderdi biomedikoetan zenbait aldaketak izan daitezke edo kausa ezezaguna ere izan dezakete, baina, hala ere, nerbio-sistema zentralean eraginak ikusgai izan. Gaixotasun neurologikoek hurrengo hauek barne hartzen dituzte: dementzia, adibidez, alzheimerra, parkinsona, Huntington...; gaixotasun zerebrobaskularrak, esaterako, migraina edo antzeko buruko minak; kondizio degeneratiboak, esate baterako, esklerosi anizkoitza... Gaixotasun hauek oso hedatuta daude eta gehienbat adineko jendeari eragiten dizkiete ondorio larrienak. Heredatutako mutazioak nerbio sistemaren garapen anormal bat, disfuntzio edo gaixotasunak eragin ditzakete. Gainera, inguruko faktoreek mutazio genetikoak eragin ditzakete, alzheimerra bezalako gaixotasunak esaterako.
Gaixotasun neurodegeneratiboek aldaketak eragiten dituzte nerbio-sistema zentraleko hainbat mekanismo biologiko eta biokimikoetan. Hauek erradikal askeen sintesia handitzen du zauri oxidatiboak eraginez, edo proteina multzo anormalak eta, azkenik hantura eragiten du. Are gehiago, neurotransmisoreak agortu, degradatu edo ez nahikoak sintetizatu daitezke entzimen gehiegizko aktibitatearengatik.
Erreferentziak
- Principles of Anatomy and Physiology Wileyplus Registration Card + Print Companion.. John Wiley & Sons Inc 2016 ISBN 978-1-119-34373-8. PMC 1000581460. (Noiz kontsultatua: 2021-09-08).
- «ZT Hiztegi Berria» zthiztegia.elhuyar.eus (Noiz kontsultatua: 2021-09-08).
- (Ingelesez) Herculano-Houzel, Suzana; Avelino-de-Souza, Kamilla; Neves, Kleber; Porfírio, Jairo; Messeder, Débora; Mattos Feijó, Larissa; Maldonado, José; Manger, Paul R.. (2014). «The elephant brain in numbers» Frontiers in Neuroanatomy 0 doi:. ISSN 1662-5129. PMID 24971054. PMC PMC4053853. (Noiz kontsultatua: 2021-09-08).
- Principles of neural science. (4th ed. argitaraldia) McGraw-Hill, Health Professions Division 2000 ISBN 0-8385-7701-6. PMC 42073108. (Noiz kontsultatua: 2021-09-08).
- Finger, Stanley. (1994). Origins of neuroscience : a history of explorations into brain function. Oxford University Press ISBN 0-19-506503-4. PMC 27151391. (Noiz kontsultatua: 2021-09-08).
- Nikoletseas, Michael M.. (2010). Behavioral and Neural Plasticity : a Learning Textbook. [CreateSpace] ISBN 978-1-4537-8945-2. PMC 756822934. (Noiz kontsultatua: 2021-09-08).
- Principles of neural science. (4th ed. argitaraldia) McGraw-Hill, Health Professions Division 2000 ISBN 0-8385-7701-6. PMC 42073108. (Noiz kontsultatua: 2021-09-08).
- (Ingelesez) Sakarya, Onur; Armstrong, Kathryn A.; Adamska, Maja; Adamski, Marcin; Wang, I.-Fan; Tidor, Bruce; Degnan, Bernard M.; Oakley, Todd H. et al.. (2007-06-06). «A Post-Synaptic Scaffold at the Origin of the Animal Kingdom» PLOS ONE 2 (6): e506. doi:. ISSN 1932-6203. PMID 17551586. PMC PMC1876816. (Noiz kontsultatua: 2021-09-08).
- Ruppert, Edward E.. (2004). Invertebrate zoology : a functional evolutionary approach. (7th ed. argitaraldia) Thomson-Brooks/Cole ISBN 0-03-025982-7. PMC 53021401. (Noiz kontsultatua: 2021-09-08).
- Balavoine, G.. (2003-02-01). «The Segmented Urbilateria: A Testable Scenario» Integrative and Comparative Biology 43 (1): 137–147. doi:. ISSN 1540-7063. (Noiz kontsultatua: 2021-09-08).
- (Ingelesez) Ortega-Hernandez, Javier. «Our 500 million-year-old nervous system fossil shines a light on animal evolution» The Conversation (Noiz kontsultatua: 2021-09-08).
- Principles of neural science. (4th ed. argitaraldia) McGraw-Hill, Health Professions Division 2000 ISBN 0-8385-7701-6. PMC 42073108. (Noiz kontsultatua: 2021-09-09).
- (Ingelesez) Allen, Nicola J.; Barres, Ben A.. (2009-02). «Glia — more than just brain glue» Nature 457 (7230): 675–677. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2021-09-09).
- (Ingelesez) Azevedo, Frederico A. C.; Carvalho, Ludmila R. B.; Grinberg, Lea T.; Farfel, José Marcelo; Ferretti, Renata E. L.; Leite, Renata E. P.; Filho, Wilson Jacob; Lent, Roberto et al.. (2009). «Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain» Journal of Comparative Neurology 513 (5): 532–541. doi:. ISSN 1096-9861. (Noiz kontsultatua: 2021-09-09).
- Freeman, Marc R.; Rowitch, David H.. (2013-10). «Evolving Concepts of Gliogenesis: A Look Way Back and Ahead to the Next 25 Years» Neuron 80 (3): 613–623. doi:. ISSN 0896-6273. PMID 24183014. PMC PMC5221505. (Noiz kontsultatua: 2021-09-09).
- (Ingelesez) «Interactions of innate and adaptive immunity in brain development and function» Brain Research 1617: 18–27. 2015-08-18 doi:. ISSN 0006-8993. PMID 25110235. PMC PMC4320678. (Noiz kontsultatua: 2021-09-09).
- Gray's anatomy : the anatomical basis of clinical practice.. (Thirty-ninth edition. argitaraldia) 2005 ISBN 0-443-07168-3. PMC 57143745. (Noiz kontsultatua: 2021-09-09).
- Hubbard, John I.. (1974). The peripheral nervous system.. Plenum Press ISBN 0-306-30764-2. PMC 874138. (Noiz kontsultatua: 2021-09-09).
- Neuroscience. (4th ed. argitaraldia) Sinauer 2008 ISBN 978-0-87893-697-7. PMC 144771764. (Noiz kontsultatua: 2021-09-09).
- (Ingelesez) Afifi, Adel K.. (1994-07-01). «Topical Review: Basal Ganglia: Functional Anatomy and Physiology. Part 1» Journal of Child Neurology 9 (3): 249–260. doi:. ISSN 0883-0738. (Noiz kontsultatua: 2021-09-09).
- Jacobs, D. K.; Nakanishi, N.; Yuan, D.; Camara, A.; Nichols, S. A.; Hartenstein, V.. (2007-06-01). «Evolution of sensory structures in basal metazoa» Integrative and Comparative Biology 47 (5): 712–723. doi:. ISSN 1540-7063. (Noiz kontsultatua: 2021-09-09).
- Sanes, Dan Harvey. (2006). Development of the nervous system. (Second edition. argitaraldia) ISBN 978-0-08-047249-2. PMC 123418470. (Noiz kontsultatua: 2021-09-09).
- Ghysen, Alain. (2003). The origin and evolution of the nervous system. (Noiz kontsultatua: 2021-09-09).
- (Ingelesez) Bourlat, Sarah J.; Juliusdottir, Thorhildur; Lowe, Christopher J.; Freeman, Robert; Aronowicz, Jochanan; Kirschner, Mark; Lander, Eric S.; Thorndyke, Michael et al.. (2006-11). «Deuterostome phylogeny reveals monophyletic chordates and the new phylum Xenoturbellida» Nature 444 (7115): 85–88. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2021-09-09).
- Lichtneckert, R.; Reichert, H.. (2005-05). «Insights into the urbilaterian brain: conserved genetic patterning mechanisms in insect and vertebrate brain development» Heredity 94 (5): 465–477. doi:. ISSN 0018-067X. PMID 15770230. (Noiz kontsultatua: 2021-09-09).
- (Ingelesez) Adey, W. R.. (1951). «The nervous system of the earthworm megascolex» Journal of Comparative Neurology 94 (1): 57–103. doi:. ISSN 1096-9861. (Noiz kontsultatua: 2021-09-09).
- «Specification of the nervous system» www.wormbook.org (Noiz kontsultatua: 2021-09-09).
- Chapman, R. F.. (1998). The insects : structure and function. (4th ed. argitaraldia) Cambridge University Press ISBN 0-521-57048-4. PMC 37682660. (Noiz kontsultatua: 2021-09-09).
- Identified neurons and behavior of arthropods. Plenum Press 1977 ISBN 0-306-31001-5. PMC 3205551. (Noiz kontsultatua: 2021-09-09).
- Neurons, networks, and motor behavior. MIT Press 1997 ISBN 0-262-19390-6. PMC 36219348. (Noiz kontsultatua: 2021-09-09).
- Korn, Henri; Faber, Donald S.. (2005-07). «The Mauthner Cell Half a Century Later: A Neurobiological Model for Decision-Making?» Neuron 47 (1): 13–28. doi:. ISSN 0896-6273. (Noiz kontsultatua: 2021-09-09).
- Hale, Me. (2000-10-01). «Startle responses of fish without Mauthner neurons: escape behavior of the lumpfish (Cyclopterus lumpus)» The Biological Bulletin 199 (2): 180–182. doi:. ISSN 0006-3185. (Noiz kontsultatua: 2021-09-09).
- Simmons, Peter J.. (1999). Nerve cells and animal behaviour. (2nd ed. argitaraldia) Cambridge University Press ISBN 0-511-01961-0. PMC 51283866. (Noiz kontsultatua: 2021-09-09).
- Gray, Peter. (2007). Psychology. (5th ed. argitaraldia) Worth Publishers ISBN 0-7167-0617-2. PMC 76872504. (Noiz kontsultatua: 2021-09-09).
- (Ingelesez) «Electrical synapses: a dynamic signaling system that shapes the activity of neuronal networks» Biochimica et Biophysica Acta (BBA) - Biomembranes 1662 (1-2): 113–137. 2004-03-23 doi:. ISSN 0005-2736. (Noiz kontsultatua: 2021-09-10).
- Principles of neural science. (4th ed. argitaraldia) McGraw-Hill, Health Professions Division 2000 ISBN 0-8385-7701-6. PMC 42073108. (Noiz kontsultatua: 2021-09-10).
- (Ingelesez) «Dale’s principle» Brain Research Bulletin 50 (5-6): 349–350. 1999-11-01 doi:. ISSN 0361-9230. (Noiz kontsultatua: 2021-09-10).
- Marty, Alain; Llano, Isabel. (2005-06). «Excitatory effects of GABA in established brain networks» Trends in Neurosciences 28 (6): 284–289. doi:. ISSN 0166-2236. (Noiz kontsultatua: 2021-09-10).
- Bear, Mark F.. (2007). Neuroscience : exploring the brain. (3rd ed. argitaraldia) Lippincott Williams & Wilkins ISBN 0-7817-7607-4. PMC 62509134. (Noiz kontsultatua: 2021-09-10).
- Cooke, S. F.. (2006-07-01). «Plasticity in the human central nervous system» Brain 129 (7): 1659–1673. doi:. ISSN 0006-8950. (Noiz kontsultatua: 2021-09-10).
- (Ingelesez) Bliss, T. V. P.; Collingridge, G. L.. (1993-01). «A synaptic model of memory: long-term potentiation in the hippocampus» Nature 361 (6407): 31–39. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2021-09-10).
- (Ingelesez) Kauer, Julie A.; Malenka, Robert C.. (2007-11). «Synaptic plasticity and addiction» Nature Reviews Neuroscience 8 (11): 844–858. doi:. ISSN 1471-0048. (Noiz kontsultatua: 2021-09-10).
- Dayan, Peter. (2001). Theoretical neuroscience : computational and mathematical modeling of neural systems. Massachusetts Institute of Technology Press ISBN 978-0-262-27130-1. PMC 51991287. (Noiz kontsultatua: 2021-09-10).
- (Ingelesez) McCulloch, Warren S.; Pitts, Walter. (1943-12-01). «A logical calculus of the ideas immanent in nervous activity» The bulletin of mathematical biophysics 5 (4): 115–133. doi:. ISSN 1522-9602. (Noiz kontsultatua: 2021-09-10).
- (Ingelesez) Sherrington, Sir Charles Scott. (1906). The Integrative Action of the Nervous System. Scribner (Noiz kontsultatua: 2021-09-10).
- Kant, Immanuel. (1987). Critique of judgment. Hackett Pub. Co ISBN 0-87220-025-6. PMC 13796153. (Noiz kontsultatua: 2021-09-10).
- Baum, William M.. (2005). Understanding behaviorism : behavior, culture, and evolution. (2nd ed. argitaraldia) Blackwell Pub ISBN 1-4051-1261-1. PMC 54082327. (Noiz kontsultatua: 2021-09-10).
- Piccolino, Marco. (2002-11). «Fifty years of the Hodgkin-Huxley era» Trends in Neurosciences 25 (11): 552–553. doi:. ISSN 0166-2236. (Noiz kontsultatua: 2021-09-10).
- Johnston, Daniel. (1995). Foundations of cellular neurophysiology. MIT Press ISBN 0-262-10053-3. PMC 29702508. (Noiz kontsultatua: 2021-09-10).
- Simmons, Peter J.. (1999). Nerve cells and animal behaviour. (2nd ed. argitaraldia) Cambridge University Press ISBN 0-511-01961-0. PMC 51283866. (Noiz kontsultatua: 2021-09-10).
- Rizzolatti, Giacomo; Craighero, Laila. (2004-06-24). «The mirror-neuron system» Annual Review of Neuroscience 27 (1): 169–192. doi:. ISSN 0147-006X. (Noiz kontsultatua: 2021-09-10).
- Keysers, Christian. (2009-11). «Mirror neurons» Current Biology 19 (21): R971–R973. doi:. ISSN 0960-9822. (Noiz kontsultatua: 2021-09-10).
- (Ingelesez) «Log In or Sign Up to View» www.facebook.com (Noiz kontsultatua: 2021-09-10).
- Rizzolatti, G.; Fadiga, L.; Fogassi, L.; Gallese, V.. (1999-05). «Resonance behaviors and mirror neurons» Archives Italiennes De Biologie 137 (2-3): 85–100. ISSN 0003-9829. PMID 10349488. (Noiz kontsultatua: 2021-09-10).
- (Ingelesez) Akins, Chana K.; Klein, Emily D.; Zentall, Thomas R.. (2002-08-01). «Imitative learning in Japanese quail (Coturnix japonica) using the bidirectional control procedure» Animal Learning & Behavior 30 (3): 275–281. doi:. ISSN 1532-5830. (Noiz kontsultatua: 2021-09-10).
- (Ingelesez) «Is the mirror neuron system involved in imitation? A short review and meta-analysis» Neuroscience & Biobehavioral Reviews 33 (7): 975–980. 2009-07-01 doi:. ISSN 0149-7634. (Noiz kontsultatua: 2021-09-10).
- Théoret, Hugo; Pascual-Leone, Alvaro. (2002-10). «Language Acquisition: Do as You Hear» Current Biology 12 (21): R736–R737. doi:. ISSN 0960-9822. (Noiz kontsultatua: 2021-09-10).
- Dinstein, Ilan; Thomas, Cibu; Behrmann, Marlene; Heeger, David J.. (2008-01-08). «A mirror up to nature» Current biology: CB 18 (1): R13–18. doi:. ISSN 0960-9822. PMID 18177704. PMC 2517574. (Noiz kontsultatua: 2021-09-10).
- Hickok, Gregory. (2009-07-01). «Eight Problems for the Mirror Neuron Theory of Action Understanding in Monkeys and Humans» Journal of Cognitive Neuroscience 21 (7): 1229–1243. doi:. ISSN 0898-929X. PMID 19199415. PMC PMC2773693. (Noiz kontsultatua: 2021-09-10).
- Sanes, Dan Harvey. (2006). Development of the nervous system. (Second edition. argitaraldia) ISBN 978-0-08-047249-2. PMC 123418470. (Noiz kontsultatua: 2021-09-10).
- Nielsen, Bjarke Frost; Nissen, Silas Boye; Sneppen, Kim; Mathiesen, Joachim; Trusina, Ala. (2020-02). «Model to Link Cell Shape and Polarity with Organogenesis» iScience 23 (2): 100830. doi:. ISSN 2589-0042. PMID 31986479. PMC PMC6994644. (Noiz kontsultatua: 2021-09-10).
- Rash, Brian G.; Lim, H. David; Breunig, Joshua J.; Vaccarino, Flora M.. (2011-10-26). «FGF Signaling Expands Embryonic Cortical Surface Area by Regulating Notch-Dependent Neurogenesis» Journal of Neuroscience 31 (43): 15604–15617. doi:. PMID 22031906. PMC PMC3235689. (Noiz kontsultatua: 2021-09-10).
- (Ingelesez) Zhu, Bangfu; Pennack, Jenny A.; McQuilton, Peter; Forero, Manuel G.; Mizuguchi, Kenji; Sutcliffe, Ben; Gu, Chun-Jing; Fenton, Janine C. et al.. (2008-11-18). «Drosophila Neurotrophins Reveal a Common Mechanism for Nervous System Formation» PLOS Biology 6 (11): e284. doi:. ISSN 1545-7885. PMID 19018662. PMC PMC2586362. (Noiz kontsultatua: 2021-09-10).
Kanpo estekak
| Autoritate kontrola |
|---|
 Datuak: Q9404
Datuak: Q9404 Multimedia: Nervous system / Q9404
Multimedia: Nervous system / Q9404