Giza eboluzioa
Giza eboluzioa prozesu ebolutiboa da primateen historiaren barruan. Homo sapiensa hominidoen familiatik bereizitako espezie gisa agertzea deskribatzen du, tximino handi guztiak barne hartzen dituen prozesua. Prozesu horrek giza bipedismoaren, trebetasunaren eta hizkuntza konplexuaren ezaugarriak mailaz maila garatzea ekarri zuen[1], baita beste hominino batzuekiko mestizajea ere[2]. Giza eboluzioa ez zen lineala izan, korapilatua baizik[3][4][5]. Giza eboluzioaren azterketak hainbat diziplina zientifiko hartzen ditu, hala nola antropologia fisikoa eta ebolutiboa, paleontologia eta genetika[6][7].
Primateak beste ugaztun batzuengandik bereizi ziren duela 85 milioi urte inguru, Goi Kretazeoan, eta haien lehen fosilak duela 55 Ma baino gehiago agertu ziren, Paleozenoan[8]. Denboraren poderioz, primateen barruan hainbat klado sortu ziren, tximinoen superfamiliara iritsi zirenak, bertatik hominidoen eta giboien familiak sortu zirelarik, 15 eta 20 milioi urte artean banatu zirenak. Hominido afrikarrak eta asiarrak (orangutanak barne) 14 Ma inguru banatu ziren. Hominidoak (australopitecina eta panina subtribuak barne) gorillini tributik (gorilak) 8-9 Ma artean bereizi ziren; australopitezinoak (gizakien arbaso bipedo desagertuak barne) 4-7 mya dituen Pan generotik (txinpantzeak eta bonoboak) bereizi ziren[9]. Homo generoaren lehen arbaso ezaguna H. habilis izan zen, duela 2 Ma baino gehiago agertu zena[10][11]; anatomikoki modernoak diren gizakiak, berriz, duela 300.000 urte inguru sortu ziren Afrikan.
Denbora-lerroa
Homininoen eboluzioan hainbat espezie sortu dira. Hona hemen eurak laburbiltzen dituen taula bat:

Homo baino lehen
Primateen eboluzio goiztiarra

Primateen eboluzio-historiak 65 milioi urte hartzen ditu, gutxienez[12][13][14][15][16]. Ezagutzen diren primate zaharrenen antzeko ugaztun espezieetako bat, Plesiadapis, Ipar Amerikatik zetorren[17][18][19][20]; beste bat, Archicebus, Txinatik[21]. Antzeko beste primate basal batzuk Eurasian eta Afrikan zehar hedatu ziren Paleozenoaren eta Eozenoaren baldintza tropikaletan.
David R. Begunek ondorioztatu zuen primate primitiboak Eurasian loratu zirela[22] eta tximino eta gizaki afrikarrak sortu zituen leinu batek, Dryopithecus barne, hegoalderantz emigratu zuela Europatik edo Asia mendebaldetik Afrikaraino. Biziraun zuten primateen populazio tropikalak, Kairoko hego-mendebaldean dagoen Faiyum sakonuneko Goi Eozenoko eta Behe Oligozenoko aztarnategi fosiletan ikusten direnak, gaur egungo primate espezie guztiak sortu zituen, Madagaskarko lemurrak, Asiako hego-ekialdeko loriak, Afrikako galagoak eta antropoideak barne. Antropodieen barnean daude Mundu Berriko Platirrinoak eta Mundu Zaharreko katarrinoak, tximino handi guztiak, gizakiak eta beste hominido batzuk barne dituen taldea.
Lehen katarrino ezaguna Kamoyapithecus da, Eragaleiteko Goi Oligozenokoa, Rift Haran Handiaren iparraldean (Kenya), duela 24 milioi urtekoa[23]. Bere jatorria Aegyptopithecus, Propliopithecus eta Parapithecus espezieekin lotuta dagoela uste da, duela 35 milioi urte ingurukoak[24]. 2010ean, Saadanius katarrinoen koroa-taldearen azken arbaso komunaren hurbileko ahaide gisa deskribatu zen, eta behin-behinean 29 eta 28 mya artean datatu zen, erregistro fosilean 11 milioi urteko hutsunea betetzen lagunduz[25].
.JPG.webp)
Miozeno goiztiarrean, duela 22 milioi urte inguru, Afrika ekialdeko zuhaitzetako katarrino mota ugariek aldez aurreko dibertsifikazio-historia luzea iradokitzen dute. Duela 20 milioi urteko talde honetako fosil oso ezagun bat Victoriapithecus da. Duela 13 milioi urte arte tximinoen leinukoak zirela uste den generoen artean, Proconsul, Rangwapithecus, Dendropithecus, Limnopithecus, Nacholapithecus, Equatorius, Nyanzapithecus, Afropithecus, Heliopithecus eta Kenyapithecus daude, guztiak ekialdeko Afrikatik etorriak.
Oso urruti dauden aztarnategietatik (Tavipithecus, Namibiako labar-aztarnategietatik, eta Pierolapithecus eta Dryopithecus, Frantziatik, Espainiatik eta Austriatik) datozen Miozeno ertaineko beste forma orokor batzuen presentzia Afrika osoan eta Mediterraneo arroan Miozeno goiztiar eta ertaineko erregimen klimatiko nahiko beroetan forma ugari izatearen froga da. Miozenoko hominoideoen artean gazteena, Oreopithecus izenekoa, duela 9 milioi urte datatutako Italiako ikatz aztarnategietatik dator.
Proba molekularren arabera, giboien leinua gainontzeko tximino handiko leinutik bereizi zen orain 18-12 Ma inguru, eta orangutanena (Ponginae subfamilia) beste tximino handiengandik 12 Ma inguru. Ez dago giboien jatorria argi dokumentatzen duen fosilik, orain arte ezagutzen ez zen Asiako hego-ekialdeko hominoideoen populazio batean sortuak izan zitezkeenak, baina proto-orangutan fosilak Indiako Sivapithecus eta Turkiako Griphopithecus generoak dira, orain 10 milioi urte ingurukoak[26].
Modu orokorrean tximino deitzen ditugun giza arbasoen eboluzioa Afrikan gertatu zen. Gizaki modernoen arestiko afrikar jatorria onarpen handiko teoria da, nahiz eta eztabaidak existitu[27], eta bere arbasoak kontinente horretan bizi izan zirela are eta onartuago[28]. Miozenoaren amaieran, orain dela 10 milioi urte inguru, Rift Haran Handia sortzen hasi zen, Afrikako ekialdeko guneak altxatu zituen prozesua. Belarrez beteriko sabanak sortzen hasi ziren, ugaztun belarjale handiagoen eboluzioa eragin zuena. Bertan bizi ziren primateek zuhaitz gutxiago eta belardi handiagoak zituzten. Eremu horietan zutik ibiltzea eraginkorragoa zen[29]. Hominidoak (gizakiak eta Australopithecina eta Panina subtribuak barne) 8-9 Ma artean bereizi ziren Gorillini tributik (gorilak); Australopithecina tribuak (gizakien arbaso bipedo desagertuak barne) Pan generotik (txinpantzeak eta bonoboak[9]) 4-7 Ma. Homo generoaren lehen arbaso ezaguna H. habilis izan zen, duela 2 Ma baino gehiago agertu zena[10][11]; anatomikoki modernoak diren gizakiak, berriz, duela 300.000 urte inguru sortu ziren Afrikan.
Gizakien kladoa beste tximinoengandik bereizi zen
Gorilen, txinpantzeen eta gizakien arbaso komunaren inguruko espezieak Kenyan aurkitutako Nakalipithecus eta Grezian aurkitutako Ouranopithecus fosilekin ordezka daitezke. Proba molekularrek iradokitzen dutenez, duela 8 eta 4 milioi urte bitartean, lehenik gorilak eta gero txinpantzeak (Pan generoa) gizakiak sortu zituen lerrotik bereizi ziren. Giza DNA % 98,4 inguru txinpantzeen berdina da nukleotido bakar baten polimorfismoak alderatuz gero. Hala ere, gorilen eta txinpantzeen erregistro fosila mugatua da; kontserbazio txarrak (oihan tropikaleko lurzoruak azidoak izaten dira eta hezurrak disolbatzen dituzte) eta laginketaren alborapenak lagundu egiten dute, ziurrenik, arazo horretan.

Beste hominido batzuk, seguru asko, ekuatore gerrikotik kanpoko inguru lehorrenetara egokitu ziren, eta han antilopeak, hienak, txakurrak, txerriak, elefanteak, zaldiak eta beste aurkitu zituzten. Ekuatore gerrikoa duela 8 milioi urte inguru uzkurtu zen, eta gorila eta txinpantze leinuen hominidoen leinuaren zatiketaren froga fosil oso gutxi daude. Batzuen arabera, giza leinuko fosilik zaharrenak Sahelanthropus tchadensis (7 Ma) eta Orrorin tugenensis (6 Ma) dira, eta ondoren Ardipithecus (5,5-4,4 Ma), Ar. kadabba eta Ar. ramidus espezieekin. Proposatu da Ar. ramidus-ek zituen hainbat adaptazio oso ezberdinak direla beste ahaide batzuekin alderatuta. Horietatik nabarmenena da burezurraren forma, heldu batean txinpantze gazte baten itxura duena. Fenomeno honi neotenia deitzen zaio, eta gizaki modernoak duen ezaugarrietako bat da, beste primateetan gazteen ezaugarriak direnak mantentzen baititu[30]. Ar. ramidusek ez zuen, gainera, letagin handirik eta dimorfismo sexuala txikiagoa zen, bonoboetan ere ikusten den tendentzia bat, baina are nabarmenago[31]. Baliteke eboluzio elementu nagusia hautespen sexualaren ondorio izatea, kumeen zaintza handituz[32].
Australopithecus
.jpg.webp)
Oihanen gutxitzearekin batera klima lehorragoa bilakatu zen, Australopithecus generoko lehen izakiak dieta berritzera eraman zituen gertakaria[33][34]. Generoa duela 4 milioi urte inguru garatu zen Afrika ekialdean, kontinente osoan zabaldu eta duela 2 milioi urte desagertu baino lehen. Aldi horretan hainbat australopiteko mota egon ziren, hala nola Australopithecus anamensis, Au. afarensis, Au. sediba eta Au. africanus. Australopithecina azpileinuaren barruan beste genero bat ere sortu zen, Homo generoaren aurrekoa ez dena, Paranthropus izenekoa. Hauek barail eta hortz oso handiak zituzten. Tradizioz esan da oskoldun fruituak eta elementu gogorrak prozesatzen zuen dieta baten ondorio zela[29], baina baliteke izatea nutriente oso gutxiko belar kopuru handiak jateko egokitzapena izatea[35]. Egile batzuek genero hau Australopithecusen barruan sartzen dute; hala denean, australopiteko sendo eta australopiteko lirain bereizketa egiten da. Kenyanthropus generoa leinu berberekoa dela ere uste da.
- Australopithecus (4–1.8 Ma), Au. anamensis, Au. afarensis, Au. africanus, Au. bahrelghazali, Au. garhi eta Au. sediba espezieekin.;
- Kenyanthropus (3–2.7 Ma), K. platyops espeziearekin;
- Paranthropus (3–1.2 Ma), P. aethiopicus, P. boisei, and P. robustus espezieekin.
Proposatutako espezie berri bat, Australopithecus deyiremeda, Au. afarensis espeziearen garai berean aurkitu zen. Au. deyiremeda espezie berri bat den edo Au. afarensis den eztabaidatzen da[36]. Australopithecus prometheus, Oin Txiki bezala ere ezaguna, duela gutxi datatu da 3,67 milioi urtetan datazio teknika berri baten bidez, eta, horren ondorioz, Australopithecus generoa bezain zaharra[37]. Oin Txikiren hatz lodi kontrajarria ikusita, badirudi eskalatzaile ona zela. Uste denez, eskualdeko gaueko harrapariak ikusita, plataforma bat eraikitzen zuten gauez zuhaitzetan habiak egiteko, txinpantze eta gorilen antzera[38].
Homo generoa

Homo generoko lehen ordezkari ezaguna Homo habilis da, duela 2,8 milioi urte inguru eboluzionatu zuena[39], eta, ziurrenik, harrizko tresnen erabileraren froga positiboak dituen espezierik zaharrena da. Lehen hominido horien garunek txinpantze batenaren tamaina bera zuten, gutxi gorabehera, baina garai hartan bikoiztu zen SRGAP2 giza genea, kortex prefrontalaren konexio azkarragoa sortuz. Hurrengo milioi urteetan entzefalizazio azkarreko prozesu bat gertatu zen, eta Homo erectus eta Homo ergasterra erregistro fosilera iritsi zirenean, garezur-ahalmena bikoiztu egin zen 850 cm3-ra arte[40]. Giza garunaren tamaina handitze hori belaunaldi bakoitzak gurasoek baino 125.000 neurona gehiago izatearen parekoa da. Uste denez, H. erectus eta H. ergaster izan ziren sua eta tresna konplexuak erabiltzen lehenak, eta hominidoen lerroko lehenak izan ziren Afrikatik irteten, Afrikan, Asian eta Europan hedatuz 1,3 ― 1,8 milioi urte bitartean.
Gizaki modernoen jatorri afrikarraren teoria berriaren arabera, hauek Afrikan eboluzionatu zuten H. heidelbergensis, H. rhodesiensis edo H. antecessor delakoetatik abiatuta, eta kontinentetik kanpora emigratu zuten duela 50.000 eta 100.000 urte bitartean, pixkanaka H. erectus, Denisovako gizakia, H. floresiensis, H. luzonensis eta H. neanderthalensis tokiko populazioak ordezkatuz[41][42][43][44]. Homo sapiens arkaikoa, anatomikoki modernoak diren gizakien aitzindaria, Erdi Paleolitoan eboluzionatu zuen duela 400.000 eta 250.000 urte bitartean[45]. Berriki egindako DNA probek iradokitzen dute jatorri neandertaleko haplotipo batzuk afrikarrak ez diren populazio guztietan daudela, eta neandertalek eta beste hominino batzuek, denisovakoek kasu, euren genomaren % 6 eman ahal izan dietela egungo gizakiei, eta horrek espezie horien arteko mestizaje mugatua iradokitzen du[46][47][48]. Kultura sinbolikoaren, hizkuntzaren eta teknologia litiko espezializatuaren garapenarekin jokabide-modernitaterako trantsizioa duela 50.000 urte inguru gertatu zen, antropologo batzuen arabera[49], baina beste proba batzuen arabera, portaeran aldaketa mailakatua gertatu zen denbora-tarte luzeago batean[50].
Homo sapiens bere generoko espezie bakarra da, Homo. Homo espezie batzuk (desagertuak) Homo sapiensen arbasoak izan zitezkeen arren, asko, agian gehienak, ziurrenik "lehengusuak" ziren, homininoen antzinako lerrotik kanpo espezializatu zirenak[51][52]. Oraindik ez dago adostasunik talde hauetatik zein den espezie banandua eta zein azpiespeziea; hau fosil eskasiagatik edo Homo generoko espezieak sailkatzeko erabilitako ezberdintasun arinengatik izan daiteke. Saharako ponparen teoriak (Saharako basamortu "heze" bat deskribatzen du, noiz edo noiz ibiltzeko modukoa) Homo generoko aldakuntza goiztiarraren azalpen posible bat eskaintzen du.
Proba arkeologiko eta paleontologikoetan oinarrituta, zenbait Homo espezieren antzinako praktika dietetikoak ondorioztatu ahal izan dira neurri bateraino[53], eta dietak Homoren barruko bilakaera fisikoan eta jokabidean duen eragina aztertu ahal izan da[54][55][56][57][58].
Antropologo eta arkeologo batzuek Tobako hondamendiaren teoria proposatzen dute. Teoria horren arabera, duela 70.000 urte inguru Sumatrako uhartean (Indonesia) Toba aintziraren supererupzioak ondorio globalak izan zituen[59], gizaki gehienak hilez eta gaur egungo gizaki guztien herentzia genetikoari eragin zion populazio-lepo bat sortuz[60]. Hala ere, gertaera horren froga genetikoak eta arkeologikoak zalantzagarriak dira oraindik[61].
H. habilis eta ingurukoak
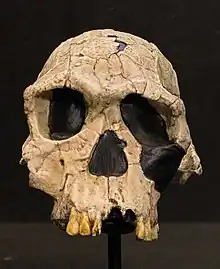
Homo habilis 2,8 eta 1,4 Ma artean bizi izan zen, gutxi gorabehera. Espeziea Pliozeno berantiarrean edo Pleistozeno goiztiarrean (2,5-2 Ma) eboluzionatu zen hegoaldeko eta ekialdeko Afrikan, australopitezinoetatik molar txikiagoen eta garun handiagoen garapenarekin aldendu zenean. Lehen hominido ezagunetako bat da, harrizko erremintak eta, agian, animalien hezurrak lantzen zituen. Horri esker, Louis Leakey aurkitzaileak Homo habilis (latinez, "gizon trebea") izena eman zion. Zientzialari batzuek Homo espezie hau Australopithecusera eramatea proposatu dute, bere eskeletoaren morfologia zuhaitzetan bizitzera egokituagoa dagoelako, ondorengo hominidoak bezala bi hanken gainean ibiltzera baino[62].
2010eko maiatzean espezie berri bat aurkitu zen Hegoafrikan: Homo gautengensis. Garezur bakar batean oinarrituta, Homo rudolfensis espeziea deskribatu zen. Baliteke H. habilis izatea, baina ez da ziurra[63]. Georgian Homo georgicus gisa deskribatutako beste hezur batzuk aurkitu dira, akaso H. habilis eta H. erectus arteko espezie bat izan daitekeena[64], edo azken honen azpiespezie bat[65].
Homo erectus eta ingurukoak

Homo erectusen lehen fosilak Eugene Dubois mediku holandarrak aurkitu zituen 1891n Indonesiako Java uhartean. Hasiera batean materialari Anthropopithecus erectus (1892-1893, garai hartan txinpantze baten antzeko primate fosiltzat hartzen zena) eta Pithecanthropus erectus (1893-1894, ordutik aurrera bere morfologian oinarrituz iritziz aldatzen zena, gizakiena eta tximinoena bitarteko gisa hartzen zituena) deitu zion[66]. Urte batzuk geroago, XX. mendean, Franz Weidenreich (1873-1948) mediku eta paleoantropologo alemaniarrak, Duboisen Javako Gizakiaren karaktereak, orduan Pithecanthropus erectus deitua, Pekingo Gizakiarenekin, orduan Sinanthropus pekinensis deitua, zehatz-mehatz alderatu zituen. Weidenreichek 1940an ondorioztatu zuen, gizaki modernoekin duten antzekotasun anatomikoa dela eta, beharrezkoa zela Java eta Txinako espezimen horiek guztiak Homo generoko espezie bakar batean biltzea, H. erectus espeziean[67][68].
Homo erectus duela gutxi gorabehera 1,8 Ma sortu zen eta orain dela 70.000 urte desagertu, eta horrek esan nahiko luke Tobako hondamendiak suntsitu zuela ziurrenik; hala ere, gertuko H. floresiensis bizirik atera zen handik. Batzuen ustez, H. erectusen hasierako fasea, 1,8 eta 1,25 Ma artekoa, espezie banandua da, H. ergaster, edo azpiespezie gisa ere har liteke, H. erectus ergaster. Paleoantropologo askok orain Homo ergaster terminoa erabiltzen dute talde honetako forma ez-asiarrentzat, eta H. erectus Asian dauden fosilentzat bakarrik gordetzen dute, H. ergasterrekiko apur bat ezberdinak diren eskeleto- eta hortzeria-baldintza jakin batzuk betetzen dituztenentzat.
Afrikan, Pleistozeno goiztiarrean, 1,5-1 Ma, uste da Homo habilisen populazio batzuek garun handiagoak garatu zituztela eta harrizko tresna landuagoak egin zituztela; desberdintasun horiek eta beste batzuk nahikoak dira antropologoek espezie berri gisa sailkatzeko[69]. Uste da belaunak blokeatzeko gaitasuna eta foramen magnumaren mugimendua biztanleria-aldaketa handien kausa izan daitezkeela. Baliteke, halaber, espezie horrek sua erabiltzea haragia prestatzeko. Richard Wranghamek dioenez, badirudi Homo erectus lurzoruan bizi zela, heste-luzera txikiagoarekin, hortzeria txikiagoa eta garunak gaur egun energia-behar handiarekin puztuta[70]; honela, hipotesi hau planteatzen du: suaren kontrola eta egostea, nutrizio-balio handiagoa askatzen zuena, Homo zuhaitzetan lo egiten zuten australopitezinoetatik bereizi zuen funtsezko egokitzapena izan zen.
Homo erectus eta Homo heidelbergensis artean bi espezie proposatu dira. Homo antecessor, Ingalaterran zein Espainiako Atapuercan aurkitu dena, orain dela 1,2 milioi eta 500.000 urte artekoa[71][72]. Italian aurkitu zen burezur bati Homo cepranensis izena eman zitzaion, orain dela 800.000 urte ingurukoa[73].
Homo heidelbergensis
Homo heidelbergensis (baita H. sapiens heidelbergensis ere[74]) Pleistozeno ertainean existitu zen gizaki arkaikoaren espezie edo azpiespezie desagertu bat da. H. erectus azpiespezie gisa aurkeztu zen 1950ean, H. e. heidelbergensis izenarekin, baina mendearen amaiera aldera espezie propio gisa gehiagotan sailkatu zen. H. heidelbergensis espeziea Europara bakarrik mugatu behar ote den edo Afrikako eta Asiako espezimenak ere sartu behar ote diren eztabaidatzen da, eta hori are gehiago zailtzen da, espezie-tipoa (Mauer 1) masailezur bat delako, masailezurrek ezaugarri diagnostiko gutxi dituztelako eta Pleistozeno ertaineko espezimenen artean nabarmen falta direlako. Horrela, bada, fosil horietako batzuk beren espezie propioan edo H. erectus azpiespezie batean bana daitezkeen eztabaidatzen da.

H. heidelbergensis kronoespezietzat hartzen da, H. erectus (batzuetan H. ergaster) forma afrikarretik abiatuta eboluzionatu zuena. Konbentzioz, H. heidelbergensis da gizaki modernoen (H. sapiens edo H. s. sapiens) eta neandertalen (H. neanderthalensis edo H. s. neanderthalensis) azken arbaso komuna. Litekeena da H. heidelbergensis-i esleitutako fosil asko gizaki modernoen eta neandertalen arteko banaketaren ondoren existitzea. Pleistozeno ertainean, burmuinaren batez besteko tamaina 1.200 zentimetro kubikokoa (cc) zen, gizaki modernoen tamainaren parekoa. Erdi Pleistozenoko garaiera hiru fosilekin soilik kalkula daiteke: Sima de los Huesos, Espainia, 169,5 cm arren kasuan eta 157,7 cm emeen kasuan; 165 cm Jinniushango (Txina) eme baten kasuan; eta 181,2 cm Kabweko (Zambia) fosil baten kasuan. Neandertalek bezala, bularra zabala zuten eta sendoak ziren oro har.
Afrika eta Europako Erdi Pleistozenoaren ezaugarria teknologia Acheul aldi berantiarra agertzea da, lehenago H. erectusek erabiltzen zuenaren ezberdina, eta ziurrenik adimen handiagoaren emaitza dena. Litekeena da sua eguneroko bizitzaren parte bihurtzea duela 400.000 urtetik aurrera, eta hori bat dator, gutxi gorabehera, Europako okupazio iraunkor eta orokorragoarekin (45° N-tik gora) eta lantzak sortzeko helduleku-teknologiaren agerpenarekin. Litekeena da H. heidelbergensisek ehiza-estrategia koordinatuak gauzatzeko gai izatea, eta, ondorioz, haragiarekiko mendekotasun handiagoa izan dutela dirudi.
H. rhodesiensis, orain dela 300.000-125.000 urte artean bizi izan zen espezie proposamen bat da. Gaur egungo ikerlari gehienek Homo heidelbergensisen taldean kokatzen dute, baina beste izen batzuk ere erabili dira, eta bada Homo sapiens rhodesiensis bezala sailkatzen duenik ere.
Lehengusu hurbilak
Homo sapiensa Homo heidelbergensisaren ondorengoa bada, badaude beste espezie batzuk ere H. heidelbergensisen ondorengotzat hartzen direnak. Horien artean ezagunenak neandertalak dira, baina azken urteotan denisovarrak ere agertu dira. Espezie hauek ez dira, sensu stricto, gizaki modernoaren arbasoak, eta beraz lehengusutzat har daitezke. Azken urteotan egindako analisi genetikoek erakutsi dute hibridazioa existitu zela, nahiz eta hau ez den globala.
Neandertalak
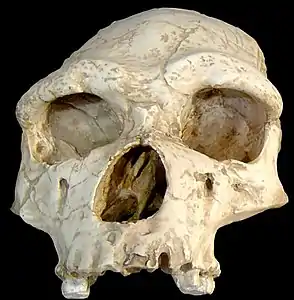 1. estaia: aurre-neandertal goiztiarra, agian Homo erectus (Tautavelko Gizakia, orain 450.000 urte).
1. estaia: aurre-neandertal goiztiarra, agian Homo erectus (Tautavelko Gizakia, orain 450.000 urte). 2. estaia: Neandertal goiztiarra, agian Homo heidelbergensis (Miguelón, orain dela 430.000 urte).
2. estaia: Neandertal goiztiarra, agian Homo heidelbergensis (Miguelón, orain dela 430.000 urte)._CROP.jpg.webp) 3. estaia: Neandertal goiztiarra (Saccopastore I, orain dela 130.000 urte).
3. estaia: Neandertal goiztiarra (Saccopastore I, orain dela 130.000 urte). 4. estaia: Neandertal europar klasikoa (La Chapelle-aux-Saints 1, orain dela 50.000 urte).
4. estaia: Neandertal europar klasikoa (La Chapelle-aux-Saints 1, orain dela 50.000 urte).
Neandertalak[75] (halaber, Neandertalgo gizakia, Homo neanderthalensis edo Homo sapiens neandertalensis[76]) Eurasian orain dela 40.000 urte arte bizi izan zen giza espezie edo azpiespezie arkaiko bat izan zen[77][78][79][80]. Ziurrenik aldaketa klimatikoaren ondorioz[81][82][83], gaixotasunen ondorioz[84][85] edo bi horien arteko konbinazio baten ondorioz desagertu ziren[83].
Ez dago argi neandertalak gizaki modernoengandik noiz banandu ziren; ikerketek duela 315.000[86] eta 800.000 urte bitarteko tarteak proposatu dituzte[87]. H. heidelbergensis arbasoaren eta neanderthalen arteko dibergentziaren garaia ere ez da argia. Neanderthal hezurrik zaharrenak duela 430.000 urte datatuak daude, baina sailkapena zalantzazkoa da[88]. Neandertalak fosil ugariengatik ezagutzen dira, batez ere duela 130.000 urtetik aurrera[89]. Espeziaren tipoa, Neanderthal 1, 1856an aurkitu zen Alemaniako Neander haranean. Fosil horren baliagarritasunaz eztabaida asko izan ondoren, XX. mendearen hasieran primitiboak, inozoak eta basatiak zirela pentsatzen zen. Nahiz eta horien ezagutza eta pertzepzioa aldatu egin den, geroztik komunitate zientifikoan, eboluziorik gabeko kobazuloetako gizaki arketipoaren irudiak lehentasuna izaten jarraitzen du herri-kulturan[90][91].

Neandertal teknologia nahiko sofistikatua izan zela uste da. Mousteriar harrizko lanabes industria zuten[92][93], eta litekeena da sua sortzeko[94][95] eta haitzuloetako sutondoak eraikitzeko trebetasunak ere[96][97], urki azalezko bikea[98], gutxienez burusien eta pontxoen antzeko arropa sinpleak[99], ehungintza[100], Mediterraneoan zehar nabigatzea[101][101], sendabelarrak erabiltzea[102][103][104], baita lesio larriak artatzeko ere[105], janaria biltegiratzea[106] eta sukaldaritzako teknika ezberdinak erabiltzea ere, hala nola, erretzea, irakitea[107] eta ketzea[108]. Neandertalek janari asko erabiltzen zuten, batez ere apatxak dituzten animaliak[109], baina baita beste megafauna[110], landare[111][112][113], ugaztun txiki, txori[114] eta uretako eta itsasoko baliabideak ere[115]. Nahiz eta puntako harrapariak izan, hartza, lehoiak, kobazuloko hiena eta beste harrapari handi batzuekin lehiatzen ziren[116]. Pentsaera sinboliko eta Paleolitikoko labar artearen hainbat adibide egotzi zaizkie Neandertalei, baina ez da ziurra[117]. Adibideetako batzuk hegazti hezurretatik[118] edo oskoletatik[119] egindako apaingarri posibleak, objektu ezohikoen bildumak, hala nola kristalak eta fosilak[120], grabatuak, musika ekoizpena[121], Divje Babe flautak adierazten duen bezala[122], eta duela 65.000 urte baino lehen eztabaidarekin[123] dataturiko Espainiako kobazuloko pinturak[124][125]. Sinesmen erlijiosoen proposamen batzuk ere egin dira[126]. Neandertalak mintzatzeko gai bide ziren, agian hizkuntza artikulatuekin, nahiz eta haien hizkuntzaren konplexutasuna ez den ezagutzen[127][128].
Gizaki modernoen aldean, neandertalek gorpuzkera sendoagoa eta gorputz-adar laburragoak zituzten. Ezaugarriok klima hotzean beroa mantentzeko egokitzapen gisa azaltzen dira sarri, baina neandertalak sarri bizi izan ziren paisaia epel eta egokitzapenak ere izan litezke[129]. Hala ere, hoztasun espezifikoko egokitzapenak zituzten, hala nola, gorputz gantz-biltegi espezializatua[130] eta sudur handia, aireaz berotzeko[131] (nahiz eta sudurra isuri genetikoek eragin izan[132]). Gizon neandertalen batezbestekoa 165 cm ingurukoa zen eta emakumezkoena, 153 cm, industrializazioaren aurreko gizaki modernoen antzekoa[133]. Neandertal gizonen eta emakumeen burmuinek 1.600 cm3 eta 1.300 cm3 inguru zituzten hurrenez hurren, gizaki modernoentzako balioen eremuan dagoena[134][135][136].
Biztanleria oso txikia zen, gene ahul kaltegarriak nabarmen hedatu ziren[137] eta distantzia luzeko sare eraginkorrak ez-ohikoak ziren. Dena den, eskualdeko kulturen eta komunitateen arteko komunikazio erregularraren frogak daude[138][139]. Kobazuloetan bizi ziren, eta leize artean mugitzen ziren urtaroen arabera[140]. Neandertalak tentsio handiko giro batean bizi ziren, traumatismo tasa altuekin, eta % 80 inguru 40 urte baino lehen hiltzen ziren[141]. Neanderthalen eta gizaki anatomikoki modernoen arteko hibridazioaren frogak 2010eko Neanderthal Genoma Proiektuaren zirriborroan aurkeztu ziren[142][143][144]. Baliteke duela 316-219 mila urte gertatu izana[145], baina litekeena da duela 100.000 urte gertatu izan eta, berriro ere, duela 65.000 urte[146]. Badirudi, halaber, Siberiako gizaki arkaikoa zen Denisovarrekin ere hibridatu izana[147][148]. Eurasiar, Australo-Melanesiar, natibo amerikar eta iparrafrikar genomen % 4 inguru gene neandertalak dira, Afrikako Saharaz azpiko eremuan azpialdeko biztanleek, aldiz, ez dute bat bera ere edo agian % 0,3ko gehienez ere. Guztira, neandertalren genomaren % 20 bizirik dirau gaur egun[149]. Neandertalengandik heredaturiko gene asko kaltegarriak edo hautespen naturalaren eragina izan lezaketen arren, neanderthal genetikak giza immunitate sistema modernoan eragina duela dirudi[150][151], eta beste hainbat funtzio biologikotan eta egituratan ere inplikatuta dagoela[152], baina zati handi batek DNA ez kodifikatzailea dela dirudi[153].
Denisovarrak

Denisovarrak edo Denisovako gizakiak gizaki arkaikoen espezie edo azpiespezie desagertu bat dira, behe eta erdi Paleolitoan Asian zehar hedatu zirenak. Denisovarren hondakin fisiko gutxi ezagutzen dira eta, ondorioz, horiei buruz dakigun gehiena DNA probetatik dator[154][155]. Material fosil osoagoa eduki arte, ez da espeziearen izen ofizialik ezarri.
Denisovarren baten lehen identifikazioa 2010ean egin zen, Altai mendietako Siberiako Denisova haitzuloan 2008an induskatutako eme gazte baten hatz-hezur batetik ateratako DNA mitokondrialetik (DNAmt) abiatuta[156]. DNA nuklearrak neandertalekiko kidetasun estuak adierazten ditu. Kobazuloan neandertalak ere bizi izan ziren aldian-aldian, baina ez dago argi neandertalak eta denisovarrak inoiz bizi izan ziren bertan. Ondoren, Denisova haitzuloko beste espezimen batzuk identifikatu ziren, baita Baishiya koba karstikoko (Tibeteko ordokian[157]) eta Cobra haitzuloko (Laoseko Annamite mendietan) ale bakar bat ere. DNA probek iradokitzen dutenez, larruazala, begiak eta ilea ilunak zituzten, eta neandertalen antzeko gorpuzkera eta aurpegi-ezaugarriak. Hala ere, Pleistozeno ertain eta berantiarreko gizaki arkaikoak eta australopitezinoak gogorarazten dituzten molar handiagoak zituzten.
Dirudienez, denisovarrek gizaki modernoekin gurutzatu ziren, eta portzentajerik handienak (%5 inguru) melanesiar, aborigen australiar eta negrito filipinarrak izan ziren. Banaketa horrek iradokitzen du populazio denisovarrak egon zirela Eurasia osoan, Filipinetan eta Ginea Berrian eta/edo Australian, baina hori ezin da baieztatu. Gizaki modernoen introgresioa duela 30.000 urte bakarrik gerta zitekeen Ginea Berrian, eta horrek, egia bada, populazio horrek duela 14.500 urte arte iraun zuela adieraz lezake. Neandertal populazioarekin mestizajearen zantzuak ere badaude, Denisova kobazuloko genoma denisovarren % 17 inguru bertatik baitator[158][159][160]. Lehen belaunaldiko "Denny" izeneko hibrido bat aurkitu zen, aita denisovarra eta ama neandertala zuena[161][162]. Gainera, genoma denisovarraren % 4 duela milioi bat urte baino gehiago gizaki modernoengandik aldendu zen giza espezie arkaiko ezezagun batetik dator[163].
H. floresiensis

Homo floresiensis orain dela 190.000 eta 50.000 urte bizi izan zen hominino bat izan zen. Hobbit ezizena jaso du bere tamaina txikiagatik, uharte-enanismoaren ondorio izan daitekeena[164]. H. floresiensis intrigagarria da, bai tamainagatik, bai antzinatasunagatik, gizaki modernoekin partekatzen ez dituen ezaugarri deribatuak dituen Homo generoko espezie berriaren adibidea baita. Beste era batera esanda, H. floresiensisek gizaki modernoekin partekatzen du arbaso komun bat, baina giza leinu modernotik bereizi zen eta eboluzio-bide desberdin bati jarraitu zion. Aurkikuntza nagusia 30 urte inguruko emakume baten hezurdura izan zen. 2003an aurkitua, 18.000 urte ingurukoa da. Kalkuluen arabera, emakumeak metro bateko altuera eta 380 cm3-ko garun-bolumena zuen (txikitzat jotzen da txinpantze batentzat, eta H. sapiensaren batez bestekoaren heren bat baino gutxiago, 1.400 cm3-koa).
Hala ere, H. floresiensis benetan espezie banandua ote den eztabaidatzen ari dira[165]. Zientzialari batzuek diote H. floresiensis H. sapiens modernoa zela, eta enanismo patologikoa zuela[166]. Hipotesi hau, neurri batean, Floresen bizi diren gizaki moderno batzuk, eskeletoa aurkitu zen Indonesiako uhartean, pigmeoak direnean oinarritzen da. Honek, enanismo patologikoarekin batera, gizaki nabarmen ñimiño bat sor zezakeen. H. floresiensis espeziearen aurkako beste eraso handi bat da H. sapiensekin soilik lotutako tresnak aurkitu zirela bere inguruan.
Hala ere, enanismo patologikoaren hipotesiak ez ditu beste ezaugarri anatomiko batzuk azaltzen, gizaki modernoen (gaixoak izan ala ez) antzekoak ez direnak, baina bai gure generoko kide ohien antzekoak. Garezurreko ezaugarriez gain, ezaugarri horiek eskumuturreko hezurren forma, besaurrea, sorbalda, belaunak eta oinak barne hartzen dituzte. Gainera, hipotesi honek ez du azaltzen ezaugarri horiek berak dituzten gizabanakoen adibide asko aurkitu izana, eta horrek adierazten du populazio handi batenak zirela eta ez zirela gizabanako bakar batera mugatzen.
2016an, Mata Mengen, Liang Boatik 74 bat kilometrora, hortz fosilak eta ustez H. floresiensisen arbasoak ziren hominidoen masailezur partziala aurkitu ziren[167]. Duela 700.000 urte ingurukoak dira eta[168], Gerrit van den Bergh arkeologo australiarraren arabera, ondorengo fosilak baino are txikiagoak dira[169].
Homo sapiens

Homo sapiens (sapiens adjektiboak "jakintsua" edo "adimentsua" esan nahi du latinez) duela 300.000 urte inguru sortu zen Afrikan, ziurrenik H. heidelbergensis edo lotutako leinu batetik eratorria[170][171]. 2019ko irailean, zientzialariek konputerizatutako determinazioaren berri eman zuten, 260 tomografia konputerizatutan oinarritua, gizaki modernoek partekatzen duten azken giza arbasoaren garezur birtual baten bidez, eta iradoki zuten gizaki modernoak duela 260.000 eta 350.000 urte sortu zirela, Afrikako ekialdeko eta hegoaldeko populazioen fusio baten bidez[172][173].
Duela 400.000 urte eta Pleistozeno ertaineko glaziazio arteko bigarren aldiaren artean, duela 250.000 urte inguru, garezur barneko bolumena hedatzeko eta harrizko erreminten teknologiak egiteko joera garatu zen, eta horrek H. erectus eta H. sapiens arteko trantsizioaren frogak ematen ditu. Zuzeneko probek iradokitzen dutenez, H. erectus espeziearen migrazio bat gertatu zen Afrikatik kanpo, eta, ondoren, H. sapiens espeziazio berri bat Afrikako H. erectusetik aurrera. Geroagoko migrazio batek (Afrika barruan zein kanpoan) ordezkatu egin zuen lehen sakabanatutako H. erectus. Migrazioaren eta jatorriaren teoria horri "oraintsuko jatorri bakarraren hipotesia" edo "Afrikatik kanpo" deitu ohi zaio. H. sapiensa gizaki arkaikoekin gurutzatu zen, bai Afrikan, bai Eurasian, batez ere neandertalekin eta denisovarrekin.
Tobako hondamendiaren teoria, duela 70.000 urte inguru H. sapiensentzat populazio-lepo bat aldarrikatzen duena[174], eztabaidagarria izan zen 1990eko hamarkadan lehenengo proposamena egin zenetik, eta 2010eko hamarkadan oso babes txikia zuen[175]. Giza aldakortasun genetiko bereizgarria efektu fundatzailearen ondorioz sortu da, Eva mitokondrial batek proposatzen duena[176], nahasketa arkaikoaren eta azkenaldiko presio ebolutiboen ondorioz.
Aldaketa ebolutiboak
Homo sapiensa txinpantzeekin partekatutako azken arbaso komunetik banandu zenetik, giza eboluzioaren ezaugarri dira aldaketa morfologikoak, garapenekoak, fisiologikoak, jokabidezkoak eta ingurumenekoak. Geroago, Pleistozenoan zehar, aurkitu zen ingurumenaren (kulturaren) bilakaerak garrantzi handia izan zuen biziraupen-sistemen arteko giza trantsizioen bidez ikusitako giza bilakaeran[177]. Egokitzapen horien artean esanguratsuenak dira bipedismoa, garunaren tamaina handitzea, ontogenia luzatzea (haurdunaldia eta haurtzaroa) eta dimorfismo sexuala murriztea. Aldaketa horien arteko lotura eztabaidagai da[178]. Beste aldaketa morfologiko esanguratsu batzuk indar- eta zehaztasun-prentsio baten bilakaera dira, H. erectusen lehen aldiz gertatu zen aldaketa[179].
Bipedismoa

bipedismoa hominidoen oinarrizko egokitzapena da, eta hominido hankabiko guztiek partekatzen dituzten aldaketa eskeletikoen kausa nagusitzat jotzen da. Lehen hominidoa, bipedismo ustez primitibokoa, Sahelanthropus[180] edo Orrorin izan zen, biak duela 6-7 milioi urte sortuak. Gorilak eta txinpantze ez-hankabikoak hominidoen lerrotik aldendu ziren garai berean, eta, beraz, Sahelanthropus edo Orrorin izan zitezkeen gure azken arbaso komunak. Ardipithecus, hankabiko osoa, duela 5,6 milioi urte inguru sortu zen[181].
Lehen hankabikoak uneren batean Australopithecus generoan bihurtu ziren, eta, geroago, Homo generokoak. Hainbat teoria daude bipedismoaren balio ebolutiboari buruz. Litekeena da bipedismoa bultatzeko arrazoietako bat, eskuak askatzen zituela izatea, elikagaiak lortzeko eta garraiatzeko, energia aurrezten zuelako lokomozioan, distantzia luzeak egiteko eta ehizatzeko aukera ematen zuelako, ikus-eremu hobea eskaintzen zuelako eta hipertermia saihesten laguntzen zuelako, eguzki zuzenaren eraginpean zegoen azalera murrizten zuelako. Ezaugarri horiek guztiak onuragarriak dira Afrikako ekialdeko Rift Haranaren altxamenduaren ondorioz sortutako sabana eta baso ingurune berrian aurrera egiteko, aurreko baso itxiaren habitataren aldean[182][183]. 2007ko ikerketa baten arabera, bipedismoak eboluzionatu egin zuen lauoinekoek baino energia gutxiago kontsumitzen zuelako[184][185]. Hala ere, berriki egindako ikerketek iradokitzen dutenez, sua erabiltzeko gaitasunik gabeko bipedismoak ez zuen ahalbidetuko sakabanatze globala[186]. Ibileraren aldaketa horrek hankak luzatu zituen, besoen luzeraren proportzioan, eta besook laburtu egin ziren zuhaitzetan eskegitzeko beharra kendu zenean. Beste aldaketa bat oineko behatz lodiaren forma da. Berriki egindako ikerketek iradokitzen dutenez, australopitezinoek denboraren zati bat zuhaitzetan bizitzen jarraitzen zuten, oineko erpuru lodiak eusteko gaitasuna baitzuen. Hau pixkanaka galtzen joan zen H. habilisen.
Anatomikoki, bipedismoaren bilakaerarekin batera aldaketa eskeletiko ugari egin dira, ez soilik hanketan eta pelbisean, baita bizkarrezurrean, oinetan eta orkatiletan eta garezurrean ere[187]. Femurra pixka bat angeluarrago bihurtu zen grabitate-zentroa gorputzaren erdigune geometrikora eramateko. Belauneko eta orkatilako artikulazioak gero eta sendoagoak egin ziren, pisua handitzea hobeto jasateko. Orno bakoitzaren pisu handiagoa posizio tentean jasateko, giza bizkarrezurrak S forma hartu zuen eta gerrialdeko ornoak laburragoak eta zabalagoak egin ziren. Oinetan, behatz lodia beste hatzekin lerrokatu zen, aurreranzko lokomozioa errazteko. Besoak eta besaurreak murriztu egin ziren hankekin alderatuta, eta horrek erraztu egin zuen lasterketa. Foramen magnuma burezurraren azpitik eta aurrerantz mugitu zen[188]
Aldaketarik nabarmenenak pelbis-eremuan gertatu ziren, beherantz begira zegoen ilion luzea laburtu eta zabaldu egin baitzen, ibiltzean grabitate-zentroa egonkor mantentzeko baldintza gisa; horregatik, hominido bipedoek pelbisa laburragoa baina zabalagoa dute, katilu itxurakoa. Eragozpen bat da tximino bipedoen erditzearen kanala txikiagoa dela hatz-koskorrekin ibiltzen diren tximinoena baino, nahiz eta zabaldu egin den australopiteko eta gizaki modernoekin alderatuta, eta horrek jaioberrien igarotzea ahalbidetzen du, garezurraren tamaina handitzearen ondorioz. Hau goiko zatira mugatzen da, igoera handiago batek hankabiko mugimendu normala zaildu dezakeelako[189].
Pelbisaren laburtzeak eta erditze kanalaren txikitzean eboluzionatu egin zuten bipedismorako baldintza gisa, eta eragin esanguratsuak izan zituzten giza erditzearen prozesuan, askoz ere zailagoa baita gizaki modernoengan beste primate batzuetan baino. Giza erditzean, pelbis-eremuaren tamaina aldatzen denez, fetuaren buruak zeharkako posizioan egon behar du (amarekin alderatuta) erditzearen kanalean sartzean, eta 90 gradu inguru biratu behar du irtetean[190]. Erditze kanala txikitzea faktore mugatzailea bihurtu zen lehen gizakietan garunaren tamaina handitzeko, eta haurdunaldi laburragoa eragin zuen, giza kumeen heldutasun eza erlatibora eraman zuena, 12 hilabete baino askoz lehenago ibiltzeko gai ez zirenak eta neotenia handiagoarekin, beste primate batzuekin alderatuta, askoz ere gazteagoak baitira[191]. Jaio ondoren garuna gehiago hazteak eta seme-alabek amarekiko mendekotasun handiagoa izateak eragin garrantzitsua izan zuten emakumeen ugalketa-zikloan eta, beste hominido batzuekin alderatuta, aloparentalitatearen agerpen ohikoagoan[192]. Giza heldutasun sexualaren atzerapenak menopausiaren bilakaerara ere eraman zuen, azalpen batekin, amonaren hipotesia, zeinaren arabera emakume zaharrek beren geneak hobeto transmititu baitzitzaketen beren alaben seme-alabak zainduz, beren seme-alaba gehiago izatearekin alderatuta[193][194].
Entzefalizazioa

Denborarekin, giza espezieak beste primate batzuena baino garun askoz handiagoa garatu zuen, 1.330 cm3 ingurukoa gizaki modernoengan, txinpantze edo gorila baten garunaren tamaina baino ia hiru aldiz handiagoa[197]. Australopithecus anamensis eta Ardipithecus espezieekin geldialdi bat izan ondoren, beren lokomozio bipedoaren ondorioz garun txikiagoak zituzten espezieak[198], entzefalizazio-eredua Homo habilis espeziearekin hasi zen, zeinaren garuna 600 cm3-koa txinpantzeena baino pixka bat handiagoa baitzen. Bilakaera horrek Homo erectusean jarraitu zuen 800-1.100 cm3-rekin, eta maximoa lortu zuen neandertaletan 1.200-1.900 cm3-rekin, Homo sapiens modernoa baino handiagoa. Garunaren hazkunde hori jaio ostean batez ere garatzen zen, beste tximinoena (heterokronia) aise gaindituz. Halaber, gizarte-ikaskuntzako eta hizkuntza eskuratzeko aldi luzeak ahalbidetu zituen gazte-gizakiengan, duela 2 milioi urte hasi zirenak. Entzefalizazioa kaloria askoko eta lortzeko zailak diren elikagaien mendekotasunaren ondorio izan daiteke[199]. Gainera, giza garunaren egituraren aldaketak tamaina handitzea baino are esanguratsuagoak izan daitezke[200][201][202].
Lobulu tenporalak, hizkuntza prozesatzeko zentroak dituztenak, neurriz kanpo handitu ziren, baita kortex prefrontala ere, erabaki konplexuak hartzearekin eta portaera soziala moderatzearekin lotu dena[197]. Entzefalizazioa dietan almidoi[203] eta haragi gehiago izatearekin lotu da[204][205], baina 2022ko meta-azterketa batek zalantzan jarri zuen haragiaren papera[206]. Beste faktore batzuk sukaldaritzaren garapena dira[207], eta proposatu da adimenak gora egin zuela gizarte-arazoak konpontzeko premia handiagoari erantzuteko, gizartea konplexuago bihurtu ahala[208]. Garezurraren morfologian izandako aldaketek, hala nola masailezurren eta masailezurretako gihar txertaketen tamaina txikiagoak, garunak hazteko leku gehiago izatea ahalbidetu zuten[209].
Neokortexaren bolumena handitzeak zerebeloaren tamaina ere azkar handitzea ekarri zuen. Haren funtzioa oreka eta kontrol motor finarekin lotu izan da tradizionalki, baina berrikiago mintzoarekin eta kognizioarekin. Tximino handiek, hominidoak barne, beste primate batzuek baino zerebelo nabarmenagoa zuten neokortexari dagokionez. Iradoki da zerebeloak giza egokitzapen teknologikoei eutsi ahal izan diela, mintzamenaren aurretiazko baldintzak barne, sentsoriomotorra kontrolatzeko eta muskulu-ekintza konplexuak ikasteko duen funtzioa dela eta[210][211][212][213]. Ikerketa batek erakutsi du gizakiek beste tximino batzuek egin ohi dituzten ohiko keinuak interpreta ditzaketela[214].
Entzefalizazioaren berehalako biziraupenaren abantaila bereiztea zaila da, Homo erectusetik Homo heidelbergensisera garuneko aldaketa nagusiak ez baitziren aldaketa teknologiko handiekin batera gertatu. Aldaketak batez ere sozialak eta jokabidezkoak izan zirela iradoki da, hala nola enpatia-gaitasunak handitzea[215][216], gizarte-taldeen tamaina handitzea[217][218] eta jokabide-plastikotasuna handitzea[219]. Gizakiak bakarrak dira gizarte-transmisioaren bidez informazioa eskuratzeko eta informazio hori egokitzeko gaitasunean. Kulturaren eboluzioaren alorrak giza aldaketa soziokulturala ikuspegi ebolutibotik aztertzen du[220].

Dimorfismo sexuala
Gizakien dimorfismo sexualaren maila txikia agerian geratzen da, batez ere, gizonezkoen letaginen murrizketan, beste tximino-espezie batzuekin alderatuta (giboiak izan ezik), eta gandor supraorbitalaren murrizketan eta arren sendotasun orokorrean. Gizakien sexualitatearekin lotutako beste aldaketa fisiologiko garrantzitsu bat obulazio ezkutuaren bilakaera izan zen. Gizakiak dira emea urte osoan ugalkorra den eta gorputzak ugalkortasun-seinale berezirik eragiten ez duen hominoide bakarrak (hala nola genitalen hantura edo araldian zehar sexua jasotzeko aukeretan aldaketa nabarmenak egitea)[223].
Hala ere, gizakiek nolabaiteko dimorfismo sexuala dute gorputzeko ilearen eta larruazalpeko koipearen banaketan, bai eta tamaina orokorrean ere; arrak emeak baino % 15 handiagoak dira, gutxi gorabehera[224]. Aldaketa horiek guztiak bikote-loturan enfasi handiagoa egitearen ondorio gisa interpretatu dira, kumeen haurtzaro luzearen ondorioz guraso-inbertsio handiagoa izateko premiaren balizko konponbide gisa[225].
Kubituaren oposizioa

Kubituaren oposizioa (hatz lodiaren eta esku bereko hatz txikiaren puntaren arteko kontaktua) Homo generokoa da[226] soilik, neandertalak, Hezurren Leizeko hominininoak eta anatomikoki modernoak diren gizakiak barne. Beste primate batzuetan, hatz lodia motza da, eta ezin du hatz txikia ukitu[227][228]. Kubituaren aurkakotasunak doitasunezko gaitasuna ematen dio giza eskuari, hala nola indar berezia objektuak heltzeko, manipulazioaren trebezia baimentzen duena.
 Giza eskua eta oina, mugimendu gaitasun handikoa.
Giza eskua eta oina, mugimendu gaitasun handikoa. Orangutan bat prezisiozko heltzea egiten.
Orangutan bat prezisiozko heltzea egiten. Gizakiak soilik uki dezake hatz txikia erpuruarekin.
Gizakiak soilik uki dezake hatz txikia erpuruarekin.
Beste aldaketa batzuk
Gizakiaren eboluzioak beste aldaketa batzuk ere izan ditu, hala nola ikusmenarekiko mendekotasun handiagoa usaimenarekiko baino (usaimen-erraboil oso txikia); gazteen garapen-aldi luzeagoa eta haurren mendekotasun handiagoa[229]; heste txikiagoa eta hortz txiki eta deslerrokatuak; metabolismo basal azkarragoa[230]; gorputz-ilearen galera[231]; izerdi-guruinen bilakaera; hortz-arkuaren formaren aldaketa, U formatik paraboliko izatera igaro zena; kokots baten garapena (Homo sapiensen baino ez dago); apofisi estiloideak; eta laringe jaitsi bat.
Portaeraren eboluzioa
Portaera modernorako aldaketa

Antropologoek giza portaera modernoa deskribatzen dute, kultura- eta jokabide-ezaugarriak barne hartzeko, hala nola tresnen espezializazioa, bitxien eta irudien erabilera (hala nola labar artea), bizi-espazioaren antolamendua, erritualak (hileta-eskaintzak, esaterako), ehiza-teknika espezializatuak, biziraupen hain erraza ez duten eremu geografikoen esplorazioa eta truke-merkataritzako sareak, baita ezaugarri orokorragoak ere, hala nola hizkuntza eta pentsamendu sinboliko konplexua. "Iraultza" batek gizaki modernoak gidatu ote zituen ("giza kontzientziaren big bang") edo eboluzioa mailakatuagoa izan ote zen eztabaidatzen jarraitzen da[232].
Duela 50.000-40.000 urte arte, badirudi harrizko erreminten erabilerak aurrera egin duela etapaka. Fase bakoitzak (H. habilis, H. ergaster, H. neanderthalensis) teknologia berri bat markatu zuen, eta hurrengo fasera arte oso astiro garatu zen. Gaur egun, paleoantropologoek eztabaidatzen dute Homo espezie horiek giza portaera moderno batzuk edo asko ote zituzten. Badirudi kulturalki kontserbadoreak izan direla, eta teknologia eta elikadura-jarraibide berberak mantendu dituztela aldi oso luzeetan.
Orain dela 50.000 urte inguru, giza kultura azkarrago eboluzionatzen hasi zen. Jokabide-modernitaterako trantsizioaren ezaugarri nagusiak "Aurrerak Jauzi Handia"[233] edo "Goi Paleolitoko Iraultza"[234] izan dira, arkeologia-erregistroan portaera modernoaren eta ehiza larriaren zeinu bereizgarriak bat-batean agertu direlako. Afrikan ere badira portaera-modernotasun nabarmen lehenagoko baten frogak, irudi abstraktuen froga zaharragoekin, biziraupen-estrategia zabalagoekin, tresna eta arma sofistikatuagoekin eta beste jokabide "moderno" batzuekin, eta aditu askok berriki argudiatu dute uste baino lehenago gertatu zela modernitaterako trantsizioa[235][236][237]. Beste batzuen ustez, trantsizioa mailakatuagoa izan zen, eta ezaugarri batzuk Homo sapiens arkaiko afrikarren artean agertu zirela diote, duela 300.000-200.000 urte[238][239][240][241]. Duela gutxi egindako proben arabera, Australiako aborigenen populazioa Afrikakotik banandu zen duela 75.000 urte, eta 160 km-ko itsas bidaia egin zuen duela 60.000 urte. Horrek garrantzia kendu diezaioke Goi Paleolitoko Iraultzari[242].
Gizaki modernoak hildakoak lurperatzen hasi ziren, animalien larruekin jantziak egiten, teknika sofistikatuagoekin ehizatzen (lubaki-tranpak erabiltzea edo itsaslabarretara animaliak bideratzea) eta kobazuloak margotzen[243]. Giza kulturak aurrera egin ahala, populazio desberdinek lehendik zeuden teknologiak berritu zituzten: amuak, botoiak eta hezurrezko orratzak bezalako tresnek aldaketa kulturalaren zantzuak erakusten dituzte, 50.000 BP baino lehen ikusi ez zirenak. Oro har, H. neanderthalensis espeziearen populazio zaharrenek ez zituzten beren teknologiak aldatu, baina ikusi da chatelperroniar multzoak H. sapiens espeziearen teknologia aurignaziarren imitazio neandertalak direla[244].
Psikologia ebolutiboa
Psikologia ebolutiboaren arabera, garunaren funtzioak behar bezala ulertzeko, garunak eboluzionatu zuen ingurunearen propietateak ulertu behar dira. Sarritan, ingurune hori "eboluzio-egokitzapeneko ingurune" gisa aipatzen da.[245]
Egokitzapen ebolutiboko ingurune baten ideia John Bowlbyren atxikimendu-teoriaren parte gisa esploratu zen lehen aldiz.[246] Mekanismo eboluzionatu berezi bat ingurune horretara egokitzen da. Zehazkiago, moldagarritasun ebolutiboaren ingurunea egokitzapen jakin bat osatu zuten historikoki behin eta berriz egin diren hautespen-presioen multzoa da, bai eta egokitzapenaren garapen eta funtzionamendu egokirako beharrezkoak izan diren inguruneko alderdiak ere.
Gizakiak, Homo generoa barne hartzen dutenak, duela 1,5 eta 2,5 milioi urte artean agertu ziren, duela 2,6 milioi urte Pleistozenoaren hasierarekin batera gutxi gorabehera. Pleistozenoa duela 12.000 urte amaitu zenez, giza egokitzapen gehienak duela gutxi eboluzionatu ziren Pleistozenoan, edo Pleistozenoan hautaketa egonkortzaile baten bidez mantendu ziren. Psikologia ebolutiboak, beraz, giza mekanismo psikologiko gehienak Pleistozenoko inguruneetan maiz aurkitzen diren ugalketa-arazoetara egokituta daudela proposatzen du. Oro har, arazo horiek hazkuntza, garapena, desberdintzea, bizirautea, sexu-harremanak izatea, haztea eta gizarte-harremanak barne hartzen dituzte.
Egokitzapen ebolutiboko giroa gizarte modernoaren oso bestelakoa da.[247] Gizaki modernoen arbasoak talde txikiagoetan bizi ziren, kultura kohesiboagoak zituzten eta identitaterako eta esanahirako testuinguru egonkorragoak eta aberatsagoak zituzten. Ikertzaileak dauden ehiztari-biltzaileen gizarteetan bila dabiltza, ehiztari-biltzaileak egokitzapen ebolutiboko ingurunean nola bizi ziren jakiteko. Nitxoaren ezaugarriak, neurri handi batean, duela 30 milioi urte baino gehiago eboluzionatu zuten ugaztun sozialen ezaugarri berak dira: jaiotza inguruko esperientzia lasaigarria, zenbait urtez eskatutako edoskitzea, afektua edo hurbiltasun fisiko ia etengabea, beharrei erantzuteko gaitasuna (ondorengoen larritasuna arintzea), joko autogidatua eta, gizakientzat, zaintzaile hartzaile ugari. Hasierako azterketek osagai horiek bizitzako lehen urteetan duten garrantzia erakusten dute, haurrengan emaitza positiboak lortzeko[248][249].
Psikologo ebolutiboek batzuetan txinpantzeei, bonoboei eta beste tximino handi batzuei begiratzen diete gizakiaren antzinako portaera ulertzeko.
Desoreka ebolutiboak
Organismo baten egokitzapenak bere antzinako ingurunera egokitu zirenez, ingurune berri eta ezberdin batek desoreka ebolutiboa sor dezake. Gizakiak batez ere Pleistozenoko inguruneetara egokituta daudenez, mekanismo psikologikoek batzuetan "desorekak" erakusten dituzte ingurune modernoarekin. Adibidez, AEBetan 10.000 pertsona inguru hiltzen dira su-armekin. Urtero, armiarmek eta sugeek eskukada bakar bat hiltzen duten bitartean, jendeak armiarmen eta sugeen beldur izaten ikasten du, arma zorrotz baten beldur bezain erraz, eta zuri apuntatzen ari ez den arma bati, untxi bati edo lore bati baino errazago[250]. Azalpen posiblea da armiarmak eta sugeak mehatxu bat zirela gizakien arbasoentzat Pleistozenoan zehar, suzko armak, untxiak edo loreak ez ziren bitartean. Beraz, gizakien beldur eboluzionatuaren ikaskuntza-psikologia eta ingurune modernoa ez datoz bat[251][252].
Desoreka hori estimulu supranormalaren fenomenoetan ere agertzen da; estimulu horrek erantzun bat eragiten du, erantzuna eboluzionatu zuen estimulua baino indar handiagoz. Niko Tinbergenek sortu zuen hitza, animalien portaera ez-gizatiarrari erreferentzia egiteko, baina Deirdre Barrett psikologoak esan zuen naturaz gaindiko estimulazioak beste animaliek bezain indartsu gobernatzen duela gizakien portaera. Azaldu zuenez, janari lasterra gehiegizko estimulua da gatz, azukre eta gantzezko[253] antojuetarako, eta dio telebistan barrea, irribarrea eta atentzioa ematen duten aurpegien gehiegikeria bat dela. Aldizkarien erdiko orrialdeek eta gaztarekin egindako hanburgesa bikoitzek erakarri egiten dituzte egokitzapen ebolutiboko ingurune batera bideratutako instintuak, non gaztetasunaren eta emankortasunaren garapena oso handia zen, eta gantza, azukrea edo gatza gutxitan eskuratzen zen sari bat zen. Mark van Vugt psikologoak ere proposatzen du gizarte-antolaketan desorekak nagusi direla.[254] Giza adimenak lidergo karismatiko eta pertsonalizatuari erantzuten dio oraindik, batez ere ingurune informal eta berdinzaleen testuinguruan. Hortik dator langile askok sentitzen duten asegabetasuna eta alienazioa. Soldatek, hobariek eta bestelako pribilegioek estatus erlatiboaren instintuak ustiatzen dituzte, batez ere gizonak goragoko postu exekutiboetara erakartzen dituztenak[255].
Tresnen erabilera


Tresnen erabilera adimen-zeinu gisa interpretatu da, eta teorizatu da baliteke giza bilakaeraren zenbait alderdi estimulatu izana, bereziki giza garunaren hedapen jarraitua. Paleontologiak oraindik azaldu behar du organo horrek milioika urtetan zehar izan duen hedapena, energia-kontsumoari dagokionez oso zorrotza izan arren. Gizaki moderno baten garunak, batez beste, 13 watt kontsumitzen ditu (260 kilokaloria egunean), atsedenean dagoen gorputzaren energia-kontsumoaren bosten bat[256]. Tresnak gehiago erabiliz gero, energia ugariko haragi-produktuak ehizatu ahal izango lirateke, eta energia askoko landare-produktu gehiago prozesatu. Ikertzaileek iradoki dutenez, lehen hominidoek presio ebolutiboa jasan zuten tresnak sortzeko eta erabiltzeko gaitasuna handitzeko[257].
Zaila da zehaztea noiz hasi ziren tresnak erabiltzen lehen gizakiak, zenbat eta primitiboagoak izan tresna horiek (adibidez, ertz zorrotzeko harriak) orduan eta zailagoa baita erabakitzea objektu naturalak edo giza tresnak diren. Zantzu batzuen arabera, australopitezinoek (4 Ma) hezur hautsiak erabil zitzaketen tresna gisa, baina hori eztabaidagarria da[258].
Espezie askok fabrikatzen eta erabiltzen dituzte tresnak, baina gizakiak dira nagusi fabrikazio-eremuetan eta tresna konplexuagoen erabileran. Ezagutzen diren tresna zaharrenak Mendebaldeko Turkanako (Kenya) lauzak dira, duela 3,3 milioi urtekoak. Hurrengo harrizko tresnarik zaharrenak Gonatik (Etiopia) datoz, eta Olduvai teknologiaren hasieratzat jotzen dira. Tresna horiek duela 2,6 milioi urtekoak dira[259]. Homoren fosil bat aurkitu zuten Olduvai tresna batzuetatik gertu, eta 2,3 milioi urte zituela adierazi zuten. Horrek iradokitzen du Homo espezieak tresna horiek sortu eta erabili zituela. Aukera bat da, baina oraindik ez da froga sendorik[260]. Hirugarren metakarpianoaren apofisi estiloideari esker, eskuko hezurra eskumuturreko hezurretan sartzen da, eta, horri esker, presio handiagoa egin daiteke eskumuturrean eta eskuan, hatz lodiarekin eta hatzekin heltzean. Horri esker, gizakiak tresna konplexuak egiteko eta erabiltzeko beharrezko trebetasuna eta indarra ditu. Ezaugarri anatomiko bakar horrek gizakiak tximinoetatik eta gizakiak ez diren beste primateetatik bereizten ditu, eta ez da ikusten 1,8 milioi urte baino gehiagoko giza fosiletan[261].
Bernard Woodek adierazi zuen Paranthropus espezieak Homoren lehen espezieekin batera bizi zirela "Olduvaiko industria-gunean", gutxi gorabehera aldi berean. Paranthropusak erreminta-fabrikatzaile gisa identifikatzen dituen zuzeneko frogarik ez dagoen arren, haien anatomia arlo horretan dituzten gaitasunen zeharkako probak egiteko aukera ematen zaie. Paleoantropologo gehienak ados daude Homoren lehen espezieak izan zirela aurkitutako Olduvai tresna gehienen arduradunak. Argudiatzen dutenez, Olduvai aldiko tresna gehienak giza fosilekin elkartuta aurkitu zirenean, Homo beti zegoen bertan, baina Paranthropus ez[260].
1994an, Randall Susmanek hatz lodi aurkarien anatomia erabili zuen Homo eta Paranthropus espezieak erreminta-fabrikatzaileak zirela argudiatzeko. Gizakien eta txinpantzeen hatz lodien hezurrak eta muskuluak alderatu zituen, eta gizakiek txinpantzeek ez dituzten 3 muskulu dituztela ikusi zuen. Gizakiek ere metakarpo lodiagoak dituzte, buru zabalagoekin, eta horrek txinpantzearen eskuak egin dezakeena baino helduleku zehatzagoa ahalbidetzen du. Giza hatz lodi aurkariaren anatomia modernoa erreminten fabrikazioari eta manipulazioari lotutako eskakizunei emandako erantzun ebolutiboa dela esan zuen Susmanek, eta bi espezieak, izatez, erreminten fabrikatzaileak zirela[260].
Giza eboluzio garaikidea
Anatomikoki modernoak diren giza populazioek eboluzionatzen jarraitzen dute, hautespen naturalak zein jito genetikoak eragiten baitie. Aro modernoan baztangaren aurkako erresistentzia bezalako ezaugarri batzuen hautespen-presioa murriztu bada ere, gizakiek hautespen naturalaren mende jarraitzen dute beste ezaugarri askotarako. Horietako batzuk ingurumen-presio espezifikoen ondorio dira; beste batzuk, berriz, bizimoduaren aldaketekin lotuta daude, nekazaritzaren garapenaz geroztik (duela 10.000 urte), urbanizazioaz geroztik (duela 5.000) eta industrializazioaz geroztik (duela 250 urte). Duela 10.000 urte nekazaritza eta duela 5.000 urte zibilizazioa garatu zirenetik giza eboluzioa bizkortu egin dela baieztatu da, eta horrek, dakarrenez, funtsezko desberdintasun genetikoak eragin ditu egungo giza populazioen artean[262], eta ikerketa berrienek adierazten dute, ezaugarri batzuetarako, giza kulturaren aurrerapenek eta berrikuntzek hautespen naturalarekin batera existitzen den hautespen modu berri bat bultzatu dutela, eta, kasu batzuetan, hura neurri handi batean ordezkatu duela[263].
Bereziki deigarria da azaleko ezaugarrien aldaketa, hala nola testura afroko ilea, edo zenbait populaziotan larruazal argiaren eta ile horiaren azken bilakaera, desberdintasun klimatikoei egozten baitzaie. Presio selektibo bereziki gogorrek garaierara egokitzea ekarri dute gizakiengan, eta desberdintasunak daude populazio isolatuetan. Oinarri genetikoen azterketek erakusten dute batzuk duela gutxi eboluzionatu zutela, tibetarren kasuan bezala, zeinak 3.000 urtean zehar eboluzionatu baitzuten, altuera handietara egokitzen den EPAS1eko alelo baten proportzio handiak izan arte[264]. Alelo hau denisovarretan ere aurkitzen da, eta pentsatu da gizakietan hibridazio bidez sartu zela[265].
Beste eboluzio batzuk gaixotasun endemikoekin lotuta daude: malariaren presentziak ezaugarri faltziformea hautatzen du (gene faltziformearen forma heterozigotikoa), eta malariarik ezean, anemia faltziformeak osasunean dituen efektuak ezaugarri horren aurkakoak dira. Beste adibide bat: gaixotasun ahulgarri kuru larria izateko arriskua duten biztanleek G127V proteina prionikoaren genearen aldaera immunitario baten gehiegizko presentzia nabarmena dute, immunitarioak ez diren aleloen aldean. Aldaera genetiko horren maiztasuna pertsona immuneen biziraupenaren ondorio da[266][267]. Obulutegi polikistikoaren sindromeak (SOP) ugalkortasuna murrizten du, eta, beraz, hautespen negatibo oso indartsu baten mende egotea espero da, baina giza populazioetan duen maiztasun erlatiboak hautespen-presioa iradokitzen du, eta horrek indargabetu egiten du. Presio horren identitatea eztabaidagai da oraindik[268].

Giza eboluzio berriak, nekazaritzarekin lotutakoak, barne hartzen du gizakien populazioetan agertu den gaixotasun infekziosoen aurkako erresistentzia genetikoa, espezieen arteko hesia animalia etxekotuetatik zeharkatzean[269], bai eta metabolismoan izandako aldaketak ere, dietan izandako aldaketen ondorioz, hala nola laktasaren iraunkortasuna.
Kulturak bultzatutako bilakaerak hautespen naturala bera desafiatu dezake: giza populazioek nolabaiteko presioa jasaten duten bitartean, adin txikiagoetan seme-alabak izateko hautaketa bultzatzen duena, antisorgailu eraginkorren etorrerak, goi-mailako hezkuntzak eta gizarte-arauen aldaketak kontrako norabidean ikusitako hautaketa bultzatu dute[270]. Hala ere, kulturak bultzatutako hautespenak ez du nahitaez hautespen naturalaren aurka egin behar: giza garunaren azkenaldiko hedapen-tasa handia azaltzeko proposamen batzuek atzeraelikadura moduko bat adierazten dute, non garunaren ikaskuntza sozialaren eraginkortasun handiagoak garapen kulturalak sustatzen dituen, aldi berean efizientzia handiagoa sustatuz, eta horrek efizientzia handiagoa eskatzen duten garapen kultural konplexuagoak bultzatzen ditu, eta horrela hurrenez hurren[271]. Kulturak bultzatutako bilakaerak badu abantaila bat: ondorio genetikoez gain, erregistro arkeologikoan ere ikus daiteke: Paleolitoan zehar harrizko tresnak garatu izanak bat egiten du kulturak bultzatutako garapen kognitiboarekin, kulturak eta gero eta konplexuagoak diren teknologien garapenak eta horiek egiteko gaitasun kognitiboak lagunduta[272].
Gaur egun, industrializaziotik, joera batzuk ikusi dira: adibidez, menopausia eboluzionatzen ari da, beranduago gertatzeko. Beste joera batzuk gizakien ugalketa-aldia luzatzea eta populazio batzuetan kolesterola, odoleko glukosa eta arteria-presioa murriztea dira[273].
Ebidentziak
Giza eboluzioari buruzko kontakizun zientifikoen oinarri diren ebidentziak natur zientzien alor askotatik datoz. Eboluzio-prozesuari buruzko ezagutza-iturri nagusia erregistro fosila izan da tradizionalki, baina 1970eko hamarkadatik aurrera genetika garatu zenetik, DNAren analisiak antzeko garrantzia izan du. Ontogenia, filogenia eta, batez ere, garapenaren biologia ebolutiboko ikerketek, ornodunenak zein ornogabeenak, bizitza osoko bilakaeraren ikuspegi nabarmena eskaintzen dute, giza eboluzioa barne. Gizakien jatorriaren eta bizitzaren azterketa espezifikoa antropologia da, bereziki paleoantropologia, giza historiaurrea aztertzen duena[274].
Ebidentzia genetikoak
Gizakien ahaide bizi hurbilenak bonoboak eta txinpantzeak (biak Pan generokoak) eta gorilak (Gorilla generokoak) dira. Giza genomaren eta txinpantzeen sekuentziazioarekin, 2012an DNA sekuentzien arteko antzekotasunaren estimazioak % 95 eta % 99 artekoak dira[275][276][277]. Bi leinuren arteko mutazio dibergenteen kopurua metatzeko behar den denbora kalkulatzen duen erloju molekularra izeneko teknikaren bidez, leinuen arteko zatiketaren gutxi gorabeherako data kalkula daiteke.
Giboiak (Hylobatidae familia) eta ondoren orangutanak (Pongo generoa) izan ziren homininoetara daraman lerrotik banandu ziren lehen taldeak, gizakiak barne, ondoren gorilak (Gorilla generoa) eta, azkenik, txinpantzeak eta bonoboak (Pan generoa). Batzuek hominidoen leinuen eta txinpantzeen arteko banaketa data duela 4 eta 8 milioi urte bitartean kokatzen dute, hau da, Miozeno berantiarrean[278][279][280]. Hala ere, badirudi espeziazioa ohiz kanpo luzea izan dela. Hasierako dibergentzia duela 7 eta 13 milioi urte bitartean gertatu zen uneren batean, baina hibridazioak banaketa lausotu zuen eta banaketa osoa atzeratu zuen milioika urtez. Pattersonek (2006) duela 5 eta 6 milioi urte arteko azken dibergentziaren data jarri zuen[281].
Homo generoko espezieak alderatzeko, lehen gizaki modernoen eta neandertalen arteko fluxu genetikoa ikertzeko eta giza migrazioaren lehen ereduak eta bereizte-datak hobeto ulertzeko ere erabili dira proba genetikoak. Hautespen naturalaren mende ez dauden eta, beraz, mutazioak erritmo nahiko konstantean metatzen dituzten genomaren zatiak konparatuz gero, posible da arbaso partekatu batetik giza espezie osoa barne hartuko duen zuhaitz genetiko bat berreraikitzea.
Mutazio jakin bat (nukleotido bakar baten polimorfismoa) banako batean agertu eta bere ondorengoei transmititzen zaien bakoitzean, haplotalde bat eratzen da, mutazio horren eramaile ere izango diren gizabanakoaren ondorengo guztiak barne hartzen dituena. DNA mitokondriala, amarengandik bakarrik heredatzen dena, alderatuz gero, genetistek ondorioztatu dute markatzaile genetikoa gizaki moderno guztietan duen azken emakumezko arbaso komuna, Eva mitokondriala izenekoa, duela 200.000 urte inguru bizi izan zela.
Giza genetika ebolutiboak giza genomak gizabanakoen artean nola bereizten diren, horiek sorrarazi zituen eboluzio-iragana eta gaur egungo ondorioak aztertzen ditu. Genomen arteko desberdintasunek ondorioak eta aplikazio antropologikoak, medikoak eta forentseak dituzte. Datu genetikoek giza eboluzioari buruzko ezagutza garrantzitsuak eman ditzakete.
Ebidentzia fosilak
%252C_3%252C2_milioni_di_anni_fa_02.jpg.webp)
Gorila, txinpantze eta hominidoen leinuen dibergentziaren froga fosil gutxi dago[282]. Hominidoen leinuko kide izateko proposatu diren lehen fosilak Sahelanthropus tchadensis (duela 7 milioi urte), Orrorin tugenensis (duela 5,7 milioi urte) eta Ardipithecus kadabba (duela 5,6 milioi urte) dira. Horietako bakoitza ondorengo hominidoen hankabiko arbaso bat dela adierazi da, baina kasu guztietan ezeztatu egin dira baieztapenak. Era berean, litekeena da espezie horietako bat edo batzuk Afrikako tximinoen beste adar baten arbasoak izatea, edo hominidoen eta beste tximinoen artean partekatutako arbaso bat ordezkatzea.
Beraz, lehen espezie fosil hauen eta hominidoen leinuaren arteko erlazioaren auzia oraindik konpontzeke dago. Lehen espezie horietatik abiatuta, australopitezinoak duela 4 milioi urte inguru sortu ziren, eta adar sendoa (Paranthropus ere deitua) eta arinetan dibergitu ziren; adar horietako bat (beharbada A. garhi), ziurrenik, Homo generoaren arbaso bihurtu zen. Australopitezino espezieak lehen aldiz agertzen dira erregistro fosilean Australopithecus afarensisekin, 100 ale fosil baino gehiagorekin, Etiopia iparraldetik ("Lucy" ospetsua bezala) Kenya eta Hegoafrikaraino aurkituak. Au. robustus (edo Paranthropus robustus) eta Au./P. boisei bezalako australopitezino sendoen fosilak bereziki ugariak dira Hegoafrikan, Kromdraai eta Swartkrans bezalako aztarnategietan, eta Turkana lakuaren inguruan Kenyan.

Homo generoko kiderik zaharrena Homo habilis da, duela 2,8 milioi urte inguru eboluzionatu zuena. H. habilis da harrizko erreminten erabileraren froga positiboak ditugun lehen espeziea. Olduvai teknologia litikoa garatu zuten, horrela deitua Olduvai lekuagatik, non lehen espezimenak aurkitu ziren. Zientzialari batzuen ustez, Homo rudolfensis, H. habilis espeziearen jatorrizko fosilen antzeko tamaina eta morfologia handiagoko fosil-taldea, aparteko espezie bat da; beste batzuen ustez, berriz, H. habilis espeziearen parte dira, eta aldaketa intraespezifikoa besterik ez da, edo, agian, baita dimorfismo sexuala ere. Lehen hominido hauen garuna txinpantze baten tamaina berekoa zen gutxi gorabehera, eta bere egokitzapen nagusia bipedismoa zen, lurreko bizitzara egokitzeko.

Hurrengo milioi urteetan entzefalizazio-prozesu bat hasi zen, eta H. erectus erregistro fosilera iristeko (duela 1,9 milioi urte inguru) garezur-ahalmena bikoiztu egin zen. H. erectusak izan ziren Afrikatik emigratu zuten lehen hominidoak, eta, duela 1,8 eta 1,3 milioi urte artean, espezie hori Afrikan, Asian eta Europan zabaldu zen. H. erectus espezieko populazio bat, batzuetan H. ergaster espezie bereizi gisa ere sailkatua, Afrikan geratu zen eta H. sapiens bihurtu zen. Uste da H. erectus eta H. ergaster izan zirela sua eta tresna konplexuak erabiltzen lehenak. Eurasian, H. erectusek eboluzionatu egin zuen H. antecessor, H. heidelbergensis eta H. neanderthalensis espezieetan. Anatomikoki modernoak diren gizakien lehen fosilak Erdi Paleolitokoak dira, duela 300-200.000 urte ingurukoak, hala nola Etiopiako Herto eta Omoko hondakinak, Marokoko Jebel Irhoudekoak eta Hegoafrikako Florisbadekoak; Israelgo eta Europa hegoaldeko Skhul haitzuloaren ondorengo fosilak duela 90.000 urte inguru hasi ziren (0,09 milioi urte).
Gizaki modernoak Afrikatik hedatu ahala, beste hominido batzuekin topo egin zuten, hala nola H. neanderthalensis eta denisovanoekin, duela 2 milioi urte Afrikatik alde egindako H. erectus populazioetatik eboluzionatu zutenak. Lehen gizakien eta espezie ahizpa hauen arteko elkarrekintzaren izaera eztabaida iturri izan da luzaroan, izan ere, kontua da ea gizakiek aurreko espezie horiek ordezkatu zituzten ala, izatez, gurutzatzeko bezain antzekoak ziren, eta kasu horretan aurreko populazio hauek material genetikoa ekar ziezaieketen gizaki modernoei.
Kalkuluen arabera, Afrikatik egindako migrazio hori duela 70-50.000 urte hasi zen. Eta gizaki modernoak mundu osora hedatu zirela, aurreko hominidoen ordez konpetentzia edo hibridazioa ezarriz. Eurasian eta Ozeanian orain dela 40.000 urte baino lehenago bizi ziren, eta Amerikara orain dela 14.500 urte lehenago gutxienez[283].
Ikerketaren historia
Darwinen aurretik
Homo hitzak, gizakia kide duen genero biologikoaren izenak, "gizaki" esan nahi du latinez. Jatorriz Lineok aukeratu zuen bere sailkapen-sisteman. Latineko "homo" erroa indoeuroperazko *dhghem edo "lurra" hitzetik dator[284]. Lineok eta bere garaiko beste zientzialari batzuek ere uste zuten tximino handiak gizakien ahaide hurbilenak zirela, antzekotasun morfologiko eta anatomikoetan oinarrituta, eta hala sailkatu zituzten[285]. Horregatik, gizakien senideei Primates izena jarri zien, lehenengoak[286].
Darwin
.jpg.webp)
Gizakiak aurreko tximinoekin ondorengotza bidez lotzeko aukera 1859tik aurrera soilik geratu zen argi Charles Darwinen Espezieen jatorriaz liburuaren argitalpenarekin, zeinean espezie berrien eboluzioaren ideia defendatzen baitzuen aurreko beste batzuetatik abiatuta. Darwinen liburuak ez zuen giza eboluzioaren gaia jorratzen, "gizakiaren jatorriari eta historiari buruzko argia emango dela" esatera mugatuta[287].
Giza eboluzioaren izaerari buruzko lehen eztabaidak Thomas Henry Huxley eta Richard Owenen artean sortu ziren. Huxleyk gizakien eboluzioa defendatu zuen tximinoetatik abiatuta, gizakien eta beste tximinoen arteko antzekotasun eta desberdintasun asko ilustratuz, eta bereziki 1863ko Evidence as to Man's Place in Nature liburuan. Darwinen lehen aldekoetako asko (Alfred Russel Wallace eta Charles Lyell kasu) hasieran ez zeuden ados adimen-gaitasunen jatorria eta gizakien sentiberatasun morala hautespen naturalaren bidez azal zitekeenarekin, nahiz eta hau geroago aldatu zen. Darwinek eboluzioaren eta hautespen sexualaren teoria aplikatu zien gizakiei 1871ko The Descent of Man, and Selection in Relation to Sex liburuan[288].
Lehen fosilak

XIX. mendeko arazo garrantzitsu bat trantsiziozko fosilen falta izan zen. 1856an neandertalen aztarnak aurkitu ziren kareharrizko harrobi batean, Espezieen jatorriaz argitaratu baino hiru urte lehenago, eta Gibraltarren neandertalen fosilak aurkitu ziren, baita lehenago ere, baina, hasiera batean, gaixotasunen bat izan zuen gizaki moderno baten hondakinak zirela baieztatu zen. 1891n Eugène Duboisek Trinilen (Java) gaur egun Homo erectus bezala ezagutzen dena aurkitu zuen arren, 1920ko hamarkadara arte, Afrikan mota honetako fosilak aurkitu ziren arte, ez ziren bitarteko espezieak pilatzen hasi[289]. 1925ean, Raymond Dartek Australopithecus africanus deskribatu zuen[290]. Taungeko Haurra zen espezie tipoa, leize batean aurkitu zuten australopitezino ume bat. Haurraren hondarrak oso ongi kontserbatutako burezur ñimiño bat eta burmuineko endokasto bat ziren.
Garuna txikia zen arren (410 cm3), forma borobila zuen, txinpantze eta gorilena ez bezala, eta giza garun modernoaren antzekoagoa. Gainera, letagin motzak zituen, eta foramen magnumaren posizioak (bizkarrezurra sartzen den garezurraren zuloa) lokomozio bipedoa erakusten zuen. Ezaugarri horiek guztiek Dart konbentzitu zuten Taungen Haurra giza arbaso hankabikoa zela, tximinoen eta gizakien arteko iragaitzazko modu bat.
Afrikako Ekialdeko fosilak

1960ko eta 1970eko hamarkadetan ehunka fosil aurkitu ziren Ekialdeko Afrikan, Olduvai zintzurreko eta Turkana lakuko eskualdeetan. Bilaketa hauek Leakey familiak egin zituen, Louis Leakey eta bere emazte Mary Leakeyrekin, eta beranduago bere seme Richard eta bere errain Meave fosil ehiztari eta paleoantropologoekin. Olduvai eta Turkana aintzirako aztarnategi fosiletan lehen hominidoen espezimenak bildu zituzten: Australopithecus eta Homo espezieak, eta baita H. erectus ere.
Aurkikuntza horiek gizadiaren sorleku bihurtu zuten Afrika. Hirurogeita hamarreko hamarkadaren amaieran eta laurogeiko hamarkadan, Etiopia paleoantropologiaren puntu bero berria bihurtu zen "Lucy" aurkitu ondoren. Australopithecus afarensis espeziearen fosilik osatuena da Lucy, eta 1974an Donald Johansonek aurkitu zuen, Hadarretik gertu, Afarreko triangeluaren basamortuko eskualdean, Etiopiako iparraldean. Espezimenak burmuin txikia bazuen ere, pelbisa eta hanketako hezurrak gizaki modernoen ia berdinak ziren, eta horrek argi erakusten zuen hominido horiek tente zebiltzala. Lucy espezie berri bezala sailkatua izan zen, Australopithecus afarensis, uste dena Homo generoarekin lotura estuagoa duela zuzeneko arbaso bezala, edo ezagutzen ez den arbaso baten hurbileko ahaide bezala, denbora tarte goiztiar honetako beste edozein hominido edo hominino ezagun baino[291]. ("Lucy" ezizena jarri zioten Beatlesen "Lucy in the Sky with Diamonds" abestiagatik, bolumen osoz eta behin eta berriz entzuten baitzen kanpamenduan indusketetan)[292]. Afar triangeluaren inguruan hominidoen fosil askoz gehiago aurkitu ziren gerora, bereziki 1990eko hamarkadan Tim D. Whitek zuzendutako taldeek aurkitutakoak edo deskribatutakoak, horien artean Ardipithecus ramidus eta Ar. kadabba.
2013an, Homo naledi espeziearen hezurdura fosilak aurkitu ziren, Homo generoari (behin-behinean) esleitutako hominido espezie desagertua, Rising Star koba-sisteman, Gizateriaren Sehaska Hegoafrikako eskualdeko aztarnategi bat, Gauteng probintzian, Johannesburgetik gertu[293]. 2015eko irailetik, gutxienez hamabost banako fosil induskatu dira kobazuloan, guztira 1.550 zati[294]. Espeziearen ezaugarriak hauek dira: gorputz txikiko giza populazioen antzeko gorputz-masa eta garaiera, Australopithecusen antzeko bolumen endokraneal txikiagoa eta Homoren lehen espezieen antzeko garezur-morfologia (garezurraren forma). Anatomia eskeletikoak australopitezinoen jatorrizko ezaugarri ezagunak eta lehen hominidoen ezaugarri ezagunak konbinatzen ditu. Gizabanakoek, hil ziren unetik gertu, kobazulo barrura nahita bota izanaren zantzuak erakusten dituzte. Fosilak duela 250.000 urte datatuak izan ziren, beraz, ez dira zuzeneko arbaso bat[295], baizik eta anatomikoki modernoak diren garun handiagoko gizakien lehen agerpenaren garaikide bat[296].
Iraultza genetikoa
Iraultza genetikoa giza eboluzioari buruzko ikerketetan, Vincent Sarichek eta Allan Wilsonek odol-serum albuminaren erreakzio immunologiko gurutzatuen indarra neurtu zutenean hasi zen, izaki pareen artean, gizakiak eta tximu afrikarrak barne (txinpantzeak eta gorilak)[297]. Erreakzioaren indarra zenbakiz adieraz zitekeen distantzia immunologiko gisa, zeina, aldi berean, espezie ezberdinetako proteina homologoen arteko aminoazidoen diferentzia kopuruarekiko proportzionala zen. Erregistro fosilean dibergentzia-denbora ezagunak dituzten espezieen bikoteen IDaren kalibrazio-kurba bat eraikitzean, datuak erloju molekular gisa erabil zitezkeen erregistro fosil pobreagoak edo ezezagunagoak dituzten pareen dibergentzia-denborak balioesteko.
1967an Sciencen argitaratutako Sarich eta Wilsonen funtsezko artikuluan gizakien eta tximinoen dibergentzia-denbora lauzpabost milioi urtekoa zela kalkulatu zuten, erregistro fosilaren interpretazio estandarrek 10 eta 30 milioi urte arteko dibergentzia hori ematen zuten unean. Geroagoko aurkikuntzek, "Lucy" k kasu, eta fosil zaharragoen berrinterpretazioak, Ramapithecusena kasu, azken zenbatespenak zuzenak zirela frogatu zuten eta albuminaren metodoa baliozkotu zuten.
DNAren sekuentziazioan izandako aurrerapenek, zehazki DNA mitokondrialarenak (DNAmt) eta ondoren Y kromosomaren DNArenak (DNA-Y), giza jatorria hobeto ulertzea ahalbidetu zuten[298][299]. Erloju molekularraren printzipioaren aplikazioak eboluzio molekularraren azterketa irauli zuen.
Orangutana duela 10 eta 20 milioi urte artean banandu zenean oinarrituta, erloju molekularraren aurreko azterketek iradokitzen zuten belaunaldi bakoitzeko 76 mutazio inguru zeudela, eta horiek ez zituztela gurasoen giza seme-alabek heredatzen; ebidentzia horrek aurretik aipatutako homininoen eta txinpantzeen arteko dibergentzia-denbora babesten zuen. Hala ere, 2012an Islandian 78 haurrekin eta haien gurasoekin egindako ikerketa baten arabera, mutazio-tasa 36 mutazio baino ez dira belaunaldi bakoitzeko; datu horrek gizakien eta txinpantzeen arteko bereizketa duela 7 milioi urte baino lehenagoko aldi batera zabaltzen du (Ma). Txinpantze basatien populazioen 226 kumerekin zortzi lekutan egindako ikerketa osagarriek iradokitzen dute txinpantzeak 26,5 urteko batez besteko adinean ugaltzen direla; horrek iradokitzen du txinpantzeen giza dibergentzia 7 eta 13 mya artean gertatu zela. Eta datu horiek iradokitzen dute Ardipithecusa (4,5 Ma), Orrorina (6 Ma) eta Sahelanthropusa (7 Ma) hominidoen leinukoak izan zitezkeela, baita banaketa Ekialdeko Afrikako Rift eskualdetik kanpo ere.
Lehen homininoaren bila
1990eko hamarkadan, paleoantropologo talde batzuk lanean ari ziren Afrika osoan zehar, tximino handietatik aurrera hominidoen leinuaren dibergentzia goiztiarrenaren frogak bilatzen. 1994an, Meave Leakeyk Australopithecus anamensis aurkitu zuen. Tim D. Whitek 1995ean aurkitu zuen Ardipithecus ramidus delakoaren aurkikuntzak eklipsatu egin zuen aurkikuntza, eta duela 4,2 milioi urtera arte atzeratu zuen erregistro fosila.
2000. urtean, Martin Pickfordek eta Brigitte Senutek, Tugengo muinoetan (Kenya), 6 milioi urteko hominido hankabiko bat aurkitu zuten, Orrorin tugenensis izenekoa. Eta 2001ean, Michel Brunetek zuzendutako talde batek Sahelanthropus tchadensisen garezurra aurkitu zuen, 7,2 milioi urtekoa, Bruneten arabera hankabikoa zena eta, beraz, homininoa zen.
Giza migrazio goiztiarren ikerketa

Giza migrazio goiztiarrakkontinenteen arteko gizaki moderno eta arkaikoen lehen migrazio eta hedapeneideritze. Ustez, gutxi gorabehera duela bi milioi urte hasi ziren, Homo erectusaren Afrikatik kanpoko hedapen goiztiarrarekin. Hasierako migrazio hau bestelako gizaki arkaikoen migrazioa baino lehenago gertatu zen; hala nola, H. heidelbergensis, duela 500,000 urte inguru bizi zena eta, ustez, Denisovanoen, Neanderthalen eta gizaki modernoen arbasoa.[301]
Afrikan zehar, Homo sapiens bere espeziazio-denboraren inguruan hedatu zen, duela 300,000 urte inguru[302], 350,000–260,000 urteko tartean.[91][303] Arestiko Afrika-jatorriaren paradigmaren arabera, Afrika-kanpoko anatomikoki modernoa den gizakia duela 70,000–50,000 urte Ekialdeko Afrikatik migraturiko H. sapiensen populazio baten ondorengoa da, Asiako eta Ozeaniako kostaldetik migratuz hedatzen jarraitu zuena, duela 50,000 urte inguru. Europan zehar, era berean, duela 40,000 urte inguru hedatzen hasi zen gizaki modernoa.
Eurasiako Homo sapiens goiztiarraren fosilak Palestina eta Grezian aurkitu dira, duela 194,000–177,000 urtekoak eta duela 210,000 urtekoak, hurrenez hurren. Fosil hauek, ustez, Homo sapiens goiztiarraren huts egindako sakabanatze-prozesuak lirateke, seguruenik populazio Neanderthal lokalek ordezkaturikoak.[302][304][305][306][90]
Migratu zuten giza populazio modernoek zaharragoak ziren populazio lokalekin gurutzatzu zirela aski ezaguna da egun; hala, gaur egungo giza populazioak, zati batean bada ere –%10ekoa baino gutxiago, non baliorik altuenak Melanesian aurkitzen diren, eta baxuenak, Eurasia eta Afrikan–, giza arkaikoen eskualde-barietateen ondorengo dira.[307][308][309]
Azken Glaziazioaren Maximoaren ostean, Ipar Eurasiako populazioak Ameriketara migratu zuten duela 20,000 urte inguru, Ipar Eurasiako iparraldeeneko lurraldeak Holozenoaren hasierara arte –duela 12,000 urte inguru– populatua izan ez zen arren. Kanada Artikoa eta Groenlandia duela 4,000 urte izan zen giza hedaduraren helburu, Paleo-Eskimoak bertara ailegatu zirenean. Amaitzeko, Polinesia duela 2,000 urte inguru populatu zen ere, Austronesiarren hedapenaren azken fasean.
Oharrak
- Ez dago akordio orokorrik H. sapiens eta H. erectus espezieen arteko lerroaren inguruan. Espezieetako batzuk akaso ez dira H. sapiensen arbaso zuzenak, eta baliteke bata bestearen ondorengo ere ez izatea, batez ere:
- H. heidelbergensis ziurrenik ez zen izan H. antecessorren ondorengoa[195].
- Agian H. heidelbergensis ere ez da H. sapiensen arbaso zuzena, ezta H. antecessor ere[195].
- H. ergaster agertzen da maiz H. erectusen ondorengo eta H. sapiensen arbaso gisa, baina ez dago argi H. ergaster espezie bat ote den, edo H. erectusen azpiespezie bat[196].
Erreferentziak
- (Ingelesez) Hall, Brian K.; Hallgrímsson, Benedikt. (2011-06-07). Strickberger's Evolution. Jones & Bartlett Publishers ISBN 978-1-4496-6390-2. (Noiz kontsultatua: 2023-01-26).
- (Ingelesez) Mondal, Mayukh; Bertranpetit, Jaume; Lao, Oscar. (2019-01-16). «Approximate Bayesian computation with deep learning supports a third archaic introgression in Asia and Oceania» Nature Communications 10 (1): 246. doi:. ISSN 2041-1723. PMID 30651539. PMC PMC6335398. (Noiz kontsultatua: 2023-01-26).
- (Ingelesez) Ackermann, Rebecca Rogers; Mackay, Alex; Arnold, Michael L.. (2016-03-01). «The Hybrid Origin of “Modern” Humans» Evolutionary Biology 43 (1): 1–11. doi:. ISSN 1934-2845. (Noiz kontsultatua: 2023-01-26).
- (Ingelesez) «Denisovans & Neandertals: Rethinking Species Boundaries» Living Anthropologically 2011-06-02 (Noiz kontsultatua: 2023-01-26).
- (Ingelesez) Yong, Ed. (2011-07-30). «Mosaic humans, the hybrid species» New Scientist 211 (2823): 34–38. doi:. ISSN 0262-4079. (Noiz kontsultatua: 2023-01-26).
- (Ingelesez) Heng, Henry H. Q.. (2009-05). «The genome-centric concept: resynthesis of evolutionary theory» BioEssays 31 (5): 512–525. doi:. (Noiz kontsultatua: 2023-01-26).
- (Ingelesez) Marlowe, Frank W.. (2005-04-13). «Hunter-gatherers and human evolution» Evolutionary Anthropology: Issues, News, and Reviews 14 (2): 54–67. doi:. (Noiz kontsultatua: 2023-01-26).
- «Meet Your Ancestors — NOVA | PBS» www.pbs.org (Noiz kontsultatua: 2023-01-26).
- (Ingelesez) «Bonobos Join Chimps as Closest Human Relatives» www.science.org (Noiz kontsultatua: 2023-01-26).
- The Cambridge encyclopedia of human evolution. Cambridge University Press 1992 ISBN 0-521-32370-3. PMC 26096467. (Noiz kontsultatua: 2023-01-26).
- (Ingelesez) Schrenk, Friedemann; Kullmer, Ottmar; Bromage, Timothy. (2007). Henke, Winfried ed. «9 The Earliest Putative Homo Fossils» Handbook of Paleoanthropology (Springer): 1611–1631. doi:. ISBN 978-3-540-33761-4. (Noiz kontsultatua: 2023-01-26).
- Maxwell 1984.
- Zhang, Rui; Wang, Yin-Qiu; Su, Bing. (2008-07). «Molecular evolution of a primate-specific microRNA family» Molecular Biology and Evolution 25 (7): 1493–1502. doi:. ISSN 1537-1719. PMID 18417486. (Noiz kontsultatua: 2023-01-26).
- (Ingelesez) Willoughby, Pamela R.. (2005-12-31). «Palaeoanthropology and the Evolutionary Place of Humans in Nature» International Journal of Comparative Psychology 18 (1) doi:. ISSN 0889-3675. (Noiz kontsultatua: 2023-01-26).
- Martin 2001, 12032–12038 orr. .
- Tavaré, Simon; Marshall, Charles R.; Will, Oliver; Soligo, Christophe; Martin, Robert D.. (2002-04-01). «Using the fossil record to estimate the age of the last common ancestor of extant primates» Nature 416: 726–729. doi:. ISSN 0028-0836. (Noiz kontsultatua: 2023-01-26).
- (Ingelesez) Rose, Kenneth D.. (2005-06-02). «The earliest primates» Evolutionary Anthropology: Issues, News, and Reviews 3 (5): 159–173. doi:. (Noiz kontsultatua: 2023-01-26).
- «John Fleagle Primate Evolution» alltheworldsprimates.org (Noiz kontsultatua: 2023-01-26).
- (Ingelesez) «National Geographic» National Geographic (Noiz kontsultatua: 2023-01-26).
- «Found in Wyoming: New fossils of oldest American primate : Gazette Archives» gazette.jhu.edu (Noiz kontsultatua: 2023-01-26).
- «Palm-Size Fossil Resets Primates’ Clock, Scientists Say - The New York Times | Ghostarchive» ghostarchive.org (Noiz kontsultatua: 2023-01-26).
- (Ingelesez) Kordos, László; Begun, David R.. (2001-01-01). «Primates from Rudabánya: allocation of specimens to individuals, sex and age categories» Journal of Human Evolution 40 (1): 17–39. doi:. ISSN 0047-2484. (Noiz kontsultatua: 2023-01-26).
- Cameron 2004, 76 orr. .
- Wallace 2004, 240 orr. .
- (Ingelesez) Zalmout, Iyad S.; Sanders, William J.; MacLatchy, Laura M.; Gunnell, Gregg F.; Al-Mufarreh, Yahya A.; Ali, Mohammad A.; Nasser, Abdul-Azziz H.; Al-Masari, Abdu M. et al.. (2010-07). «New Oligocene primate from Saudi Arabia and the divergence of apes and Old World monkeys» Nature 466 (7304): 360–364. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2023-01-26).
- Srivastava 2009.
- «Out-of-Africa versus the multiregional hypothesis | Learn Science at Scitable» www.nature.com (Noiz kontsultatua: 2023-01-26).
- (Ingelesez) Wilshaw, Alex. (2018-10-04). Trevathan, Wenda ed. «Out of Africa hypothesis» The International Encyclopedia of Biological Anthropology (John Wiley & Sons, Inc.): 1–7. doi:. ISBN 978-1-118-58442-2. (Noiz kontsultatua: 2023-01-26).
- (Ingelesez) Owen-Smith, Norman. «Large mammals shaped the evolution of humans: here’s why it happened in Africa» The Conversation (Noiz kontsultatua: 2023-01-26).
- Dean, M.C.; Wood, B.A.. (1984-02-14). «Phylogeny, Neoteny and Growth of the Cranial Base in Hominoids» Folia Primatologica 43 (2-3): 157–180. doi:. ISSN 0015-5713. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Sayers, Ken; Raghanti, Mary Ann; Lovejoy, C. Owen. (2012-10-21). «Human Evolution and the Chimpanzee Referential Doctrine» Annual Review of Anthropology 41 (1): 119–138. doi:. ISSN 0084-6570. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Clark, Gary; Henneberg, Maciej. (2015-06-30). «The life history of “Ardipithecus ramidus”: a heterochronic model of sexual and social maturation» Anthropological Review 78 (2): 109–132. doi:. ISSN 2083-4594. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Bonnefille, R.; Potts, R.; Chalié, F.; Jolly, D.; Peyron, O.. (2004-08-17). «High-resolution vegetation and climate change associated with Pliocene Australopithecus afarensis» Proceedings of the National Academy of Sciences 101 (33): 12125–12129. doi:. ISSN 0027-8424. PMID 15304655. PMC PMC514445. (Noiz kontsultatua: 2023-01-26).
- (Ingelesez) Sponheimer, Matt; Lee-Thorp, Julia A.. (1999-01-15). «Isotopic Evidence for the Diet of an Early Hominid, Australopithecus africanus» Science 283 (5400): 368–370. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2023-01-26).
- (Ingelesez) Cerling, Thure E.; Mbua, Emma; Kirera, Francis M.; Manthi, Fredrick Kyalo; Grine, Frederick E.; Leakey, Meave G.; Sponheimer, Matt; Uno, Kevin T.. (2011-06-07). «Diet of Paranthropus boisei in the early Pleistocene of East Africa» Proceedings of the National Academy of Sciences 108 (23): 9337–9341. doi:. ISSN 0027-8424. PMID 21536914. PMC PMC3111323. (Noiz kontsultatua: 2023-01-26).
- «The Human Family Tree Bristles With New Branches - The New York Times | Ghostarchive» ghostarchive.org (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Crompton, Robin Huw; McClymont, Juliet; Elton, Sarah; Thorpe, Susannah; Sellers, William; Heaton, Jason; Pickering, Travis Rayne; Pataky, Todd et al.. (2021). «StW 573 Australopithecus prometheus: Its Significance for an Australopith Bauplan» Folia Primatologica 92 (5-6): 243–275. doi:. ISSN 0015-5713. PMID 34583353. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Senut, Brigitte. (2014-09-01). «When the ancestors were arboreal» Antiquity 88 (341): 921–922. doi:. ISSN 0003-598X. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) «'First human' discovered in Ethiopia» BBC News 2015-03-04 (Noiz kontsultatua: 2023-01-27).
- Swisher, Curtis & Lewin 2001.
- Stringer 1994, 242 orr. .
- McHenry 2009, 265 orr. .
- (Ingelesez) «This Week in Science» Science 308 (5724): 921–921. 2005-05-13 doi:. ISSN 0036-8075. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Stringer, Chris. (2003-06). «Out of Ethiopia» Nature 423 (6941): 693–695. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) «Evolution's Past Is Modern Human's Present» www.nsf.gov (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Reich, David; Green, Richard E.; Kircher, Martin; Krause, Johannes; Patterson, Nick; Durand, Eric Y.; Viola, Bence; Briggs, Adrian W. et al.. (2010-12). «Genetic history of an archaic hominin group from Denisova Cave in Siberia» Nature 468 (7327): 1053–1060. doi:. ISSN 1476-4687. PMID 21179161. PMC PMC4306417. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Noonan, James P.. (2010-05-01). «Neanderthal genomics and the evolution of modern humans» Genome Research 20 (5): 547–553. doi:. ISSN 1088-9051. PMID 20439435. PMC PMC2860157. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Abi-Rached, Laurent; Jobin, Matthew J.; Kulkarni, Subhash; McWhinnie, Alasdair; Dalva, Klara; Gragert, Loren; Babrzadeh, Farbod; Gharizadeh, Baback et al.. (2011-10-07). «The Shaping of Modern Human Immune Systems by Multiregional Admixture with Archaic Humans» Science 334 (6052): 89–94. doi:. ISSN 0036-8075. PMID 21868630. PMC PMC3677943. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Mellars, Paul. (2006-06-20). «Why did modern human populations disperse from Africa ca. 60,000 years ago? A new model» Proceedings of the National Academy of Sciences 103 (25): 9381–9386. doi:. ISSN 0027-8424. PMID 16772383. PMC PMC1480416. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Mcbrearty, Sally; Brooks, Alison S.. (2000-11-01). «The revolution that wasn't: a new interpretation of the origin of modern human behavior» Journal of Human Evolution 39 (5): 453–563. doi:. ISSN 0047-2484. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Strait, David S.; Grine, Frederick E.; Moniz, Marc A.. (1997-01-01). «A reappraisal of early hominid phylogeny» Journal of Human Evolution 32 (1): 17–82. doi:. ISSN 0047-2484. (Noiz kontsultatua: 2023-01-27).
- Bryson 2004, 522–543 orr. .
- «For Evolving Brains, a ‘Paleo’ Diet of Carbs - The New York Times | Ghostarchive» ghostarchive.org (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Leonard, William R.; Snodgrass, J. Josh; Robertson, Marcia L.. (2007-08-01). «Effects of Brain Evolution on Human Nutrition and Metabolism» Annual Review of Nutrition 27 (1): 311–327. doi:. ISSN 0199-9885. (Noiz kontsultatua: 2023-01-27).
- Walker 2007, 3–10 orr. .
- Ungar & Teaford 2002.
- Bogin 1997, 96–142 orr. .
- (Ingelesez) Barnicot, N. A.. (2005-04-01). «Human nutrition: Evolutionary perspectives» Integrative Physiological & Behavioral Science 40 (2): 114–117. doi:. ISSN 1936-3567. (Noiz kontsultatua: 2023-01-27).
- «BBC - Science & Nature - The evolution of man» web.archive.org 2006-01-18 (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) When humans faced extinction. 2003-06-09 (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) «Modern humans flourished through ancient supervolcano eruption 74,000 years ago: Modern humans flourished through ancient supervolcano eruption» ScienceDaily (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Wood, B.; Collard, M.. (1999). «The changing face of genus Homo» Evolutionary Anthropology: Issues (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Wood, Bernard. (1999-01-01). «‘Homo rudolfensis’ Alexeev, 1986—fact or phantom?» Journal of Human Evolution 36 (1): 115–118. doi:. ISSN 0047-2484. (Noiz kontsultatua: 2023-01-27).
- (Frantsesez) Gabounia, Léo; de Lumley, Marie-Antoinette; Vekua, Abesalom; Lordkipanidze, David; de Lumley, Henry. (2002-09-01). «Découverte d’un nouvel hominidé à Dmanissi (Transcaucasie, Géorgie)» Comptes Rendus Palevol 1 (4): 243–253. doi:. ISSN 1631-0683. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Lordkipanidze, David; Vekua, Abesalom; Ferring, Reid; Rightmire, G. Philip; Zollikofer, Christoph P.E.; Ponce de León, Marcia S.; Agusti, Jordi; Kiladze, Gocha et al.. (2006-11). «A fourth hominin skull from Dmanisi, Georgia» The Anatomical Record Part A: Discoveries in Molecular, Cellular, and Evolutionary Biology 288A (11): 1146–1157. doi:. (Noiz kontsultatua: 2023-01-27).
- Turner, W.. (1895-04). «On M. Dubois' Description of Remains recently found in Java, named by him Pithecanthropus erectus: With Remarks on so-called Transitional Forms between Apes and Man» Journal of Anatomy and Physiology 29 (Pt 3): 424–445. PMID 17232143. PMC 1328414. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Weidenreich, Franz. (1940-07). «Some Problems Dealing with Ancient Man» American Anthropologist 42 (3): 375–383. doi:. ISSN 0002-7294. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Grine, Frederick E.; Fleagle, John G.. (2009). Grine, Frederick E. ed. «The First Humans: A Summary Perspective on the Origin and Early Evolution of the Genus Homo» The First Humans – Origin and Early Evolution of the Genus Homo: Contributions from the Third Stony Brook Human Evolution Symposium and Workshop October 3 – October 7, 2006 (Springer Netherlands): 197–207. doi:. ISBN 978-1-4020-9980-9. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Spoor, Fred; Wood, Bernard; Zonneveld, Frans. (1994-06). «Implications of early hominid labyrinthine morphology for evolution of human bipedal locomotion» Nature 369 (6482): 645–648. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2023-01-27).
- «Catching Fire: How Cooking Made Us Human by Richard Wrangham: review» www.telegraph.co.uk (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Bermúdez de Castro, J. M.; Arsuaga, J. L.; Carbonell, E.; Rosas, A.; Martı́nez, I.; Mosquera, M.. (1997-05-30). «A Hominid from the Lower Pleistocene of Atapuerca, Spain: Possible Ancestor to Neandertals and Modern Humans» Science 276 (5317): 1392–1395. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Carbonell, Eudald; Bermúdez de Castro, José M.; Parés, Josep M.; Pérez-González, Alfredo; Cuenca-Bescós, Gloria; Ollé, Andreu; Mosquera, Marina; Huguet, Rosa et al.. (2008-03). «The first hominin of Europe» Nature 452 (7186): 465–469. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Manzi, G.; Mallegni, F.; Ascenzi, A.. (2001-08-14). «A cranium for the earliest Europeans: Phylogenetic position of the hominid from Ceprano, Italy» Proceedings of the National Academy of Sciences 98 (17): 10011–10016. doi:. ISSN 0027-8424. PMID 11504953. PMC PMC55569. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Czarnetzki, A.; Jakob, T.; Pusch, C. M.. (2003-04-01). «Palaeopathological and variant conditions of the Homo heidelbergensis type specimen (Mauer, Germany)» Journal of Human Evolution 44 (4): 479–495. doi:. ISSN 0047-2484. (Noiz kontsultatua: 2023-01-27).
- «ZT Hiztegi Berria» zthiztegia.elhuyar.eus (Noiz kontsultatua: 2021-02-18).
- Pääbo, Svante. (2014). Neanderthal man : in search of lost genomes. ISBN 978-0-465-02083-6. PMC 862400377. (Noiz kontsultatua: 2021-02-16).
- (Ingelesez) Higham, Tom; Douka, Katerina; Wood, Rachel; Ramsey, Christopher Bronk; Brock, Fiona; Basell, Laura; Camps, Marta; Arrizabalaga, Alvaro et al.. (2014-08). «The timing and spatiotemporal patterning of Neanderthal disappearance» Nature 512 (7514): 306–309. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Higham, Thomas. (2011/03). «European Middle and Upper Palaeolithic radiocarbon dates are often older than they look: problems with previous dates and some remedies» Antiquity 85 (327): 235–249. doi:. ISSN 0003-598X. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Pinhasi, Ron; Higham, Thomas F. G.; Golovanova, Liubov V.; Doronichev, Vladimir B.. (2011-05-24). «Revised age of late Neanderthal occupation and the end of the Middle Paleolithic in the northern Caucasus» Proceedings of the National Academy of Sciences 108 (21): 8611–8616. doi:. ISSN 0027-8424. PMID 21555570. PMC PMC3102382. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Galván, Bertila; Hernández, Cristo M.; Mallol, Carolina; Mercier, Norbert; Sistiaga, Ainara; Soler, Vicente. (2014-10-01). «New evidence of early Neanderthal disappearance in the Iberian Peninsula» Journal of Human Evolution 75: 16–27. doi:. ISSN 0047-2484. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Bradtmöller, Marcel; Pastoors, Andreas; Weninger, Bernhard; Weniger, Gerd-Christian. (2012-01-09). «The repeated replacement model – Rapid climate change and population dynamics in Late Pleistocene Europe» Quaternary International 247: 38–49. doi:. ISSN 1040-6182. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Wolf, D.; Kolb, T.; Alcaraz-Castaño, M.; Heinrich, S.; Baumgart, P.; Calvo, R.; Sánchez, J.; Ryborz, K. et al.. (2018-05-04). «Climate deteriorations and Neanderthal demise in interior Iberia» Scientific Reports 8 (1): 7048. doi:. ISSN 2045-2322. PMID 29728579. PMC PMC5935692. (Noiz kontsultatua: 2021-02-15).
- Black, Benjamin A.; Neely, Ryan R.; Manga, Michael. (2015-05). «Campanian Ignimbrite volcanism, climate, and the final decline of the Neanderthals» Geology 43 (5): 411–414. doi:. ISSN 1943-2682. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Underdown, Simon. (2008-01-01). «A potential role for Transmissible Spongiform Encephalopathies in Neanderthal extinction» Medical Hypotheses 71 (1): 4–7. doi:. ISSN 0306-9877. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Sullivan, Alexis P.; de Manuel, Marc; Marques-Bonet, Tomas; Perry, George H.. (2017-07-01). «An evolutionary medicine perspective on Neandertal extinction» Journal of Human Evolution 108: 62–71. doi:. ISSN 0047-2484. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Stringer, Chris. (2012). «The status of Homo heidelbergensis (Schoetensack 1908)» Evolutionary Anthropology: Issues, News, and Reviews 21 (3): 101–107. doi:. ISSN 1520-6505. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Gómez-Robles, Aida. (2019-05-01). «Dental evolutionary rates and its implications for the Neanderthal–modern human divergence» Science Advances 5 (5): eaaw1268. doi:. ISSN 2375-2548. PMID 31106274. PMC PMC6520022. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Meyer, Matthias; Arsuaga, Juan-Luis; de Filippo, Cesare; Nagel, Sarah; Aximu-Petri, Ayinuer; Nickel, Birgit; Martínez, Ignacio; Gracia, Ana et al.. (2016-03). «Nuclear DNA sequences from the Middle Pleistocene Sima de los Huesos hominins» Nature 531 (7595): 504–507. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2021-02-15).
- KLEIN, RICHARD G.. (1983). «ANTHROPOLOGY: What Do We Know About Neanderthals and Cro-Magnon Man?» The American Scholar 52 (3): 386–392. ISSN 0003-0937. (Noiz kontsultatua: 2021-02-15).
- Papagianni, Dimitra. (2015). The Neanderthals rediscovered : how modern science is rewriting their story. (Revised and updated edition. argitaraldia) ISBN 978-0-500-77311-6. PMC 919727485. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Drell, Julia R. R.. (2000). «Neanderthals: A History of Interpretation» Oxford Journal of Archaeology 19 (1): 1–24. doi:. ISSN 1468-0092. (Noiz kontsultatua: 2021-02-15).
- Shaw, Ian. (2007). Dictionary of Archaeology.. John Wiley & Sons ISBN 978-0-470-75196-1. PMC 476170403. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Lycett, Stephen J.; von Cramon-Taubadel, Noreen. (2013-03-01). «A 3D morphometric analysis of surface geometry in Levallois cores: patterns of stability and variability across regions and their implications» Journal of Archaeological Science 40 (3): 1508–1517. doi:. ISSN 0305-4403. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Sorensen, A. C.; Claud, E.; Soressi, M.. (2018-07-19). «Neandertal fire-making technology inferred from microwear analysis» Scientific Reports 8 (1): 10065. doi:. ISSN 2045-2322. PMID 30026576. PMC PMC6053370. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Brittingham, Alex; Hren, Michael T.; Hartman, Gideon; Wilkinson, Keith N.; Mallol, Carolina; Gasparyan, Boris; Adler, Daniel S.. (2019-10-25). «Geochemical Evidence for the Control of Fire by Middle Palaeolithic Hominins» Scientific Reports 9 (1): 15368. doi:. ISSN 2045-2322. PMID 31653870. PMC PMC6814844. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Hayden, Brian. (2012). «Neandertal Social Structure?» Oxford Journal of Archaeology 31 (1): 1–26. doi:. ISSN 1468-0092. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Kedar, Yafit; Barkai, Ran. (2019-06-12). «The Significance of Air Circulation and Hearth Location at Paleolithic Cave Sites» Open Quaternary 5 (1): 4. doi:. ISSN 2055-298X. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Schmidt, Patrick; Blessing, Matthias; Rageot, Maxime; Iovita, Radu; Pfleging, Johannes; Nickel, Klaus G.; Righetti, Ludovic; Tennie, Claudio. (2019-09-03). «Birch tar production does not prove Neanderthal behavioral complexity» Proceedings of the National Academy of Sciences 116 (36): 17707–17711. doi:. ISSN 0027-8424. PMID 31427508. PMC PMC6731756. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Hoffecker, John F.. (2009-09-22). «The spread of modern humans in Europe» Proceedings of the National Academy of Sciences 106 (38): 16040–16045. doi:. ISSN 0027-8424. PMID 19571003. PMC PMC2752585. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Hardy, B. L.; Moncel, M.-H.; Kerfant, C.; Lebon, M.; Bellot-Gurlet, L.; Mélard, N.. (2020-04-09). «Direct evidence of Neanderthal fibre technology and its cognitive and behavioral implications» Scientific Reports 10 (1): 4889. doi:. ISSN 2045-2322. PMID 32273518. PMC PMC7145842. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Ferentinos, George; Gkioni, Maria; Geraga, Maria; Papatheodorou, George. (2012-07-01). «Early seafaring activity in the southern Ionian Islands, Mediterranean Sea» Journal of Archaeological Science 39 (7): 2167–2176. doi:. ISSN 0305-4403. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Hardy, Karen; Buckley, Stephen; Huffman, Michael. (2013/09). «Neanderthal self-medication in context» Antiquity 87 (337): 873–878. doi:. ISSN 0003-598X. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Lev, Efraim; Kislev, Mordechai E.; Bar-Yosef, Ofer. (2005-03-01). «Mousterian vegetal food in Kebara Cave, Mt. Carmel» Journal of Archaeological Science 32 (3): 475–484. doi:. ISSN 0305-4403. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Weyrich, Laura S.; Duchene, Sebastian; Soubrier, Julien; Arriola, Luis; Llamas, Bastien; Breen, James; Morris, Alan G.; Alt, Kurt W. et al.. (2017-04). «Neanderthal behaviour, diet, and disease inferred from ancient DNA in dental calculus» Nature 544 (7650): 357–361. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) «Living to fight another day: The ecological and evolutionary significance of Neanderthal healthcare» Quaternary Science Reviews 217: 98–118. 2019-08-01 doi:. ISSN 0277-3791. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Valensi, Patricia; Michel, Véronique; El Guennouni, Khalid; Liouville, Marie. (2013-12-06). «New data on human behavior from a 160,000 year old Acheulean occupation level at Lazaret cave, south-east France: An archaeozoological approach» Quaternary International 316: 123–139. doi:. ISSN 1040-6182. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Krief, Sabrina; Daujeard, Camille; Moncel, Marie-Hélène; Lamon, Noemie; Reynolds, Vernon. (2015/04). «Flavouring food: the contribution of chimpanzee behaviour to the understanding of Neanderthal calculus composition and plant use in Neanderthal diets» Antiquity 89 (344): 464–471. doi:. ISSN 0003-598X. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Hardy, Karen; Buckley, Stephen; Collins, Matthew J.; Estalrrich, Almudena; Brothwell, Don; Copeland, Les; García-Tabernero, Antonio; García-Vargas, Samuel et al.. (2012-08-01). «Neanderthal medics? Evidence for food, cooking, and medicinal plants entrapped in dental calculus» Naturwissenschaften 99 (8): 617–626. doi:. ISSN 1432-1904. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Dusseldorp, Gerrit L.. (2013). Clark, Jamie L. ed. «Neanderthals and Cave Hyenas: Co-existence, Competition or Conflict?» Zooarchaeology and Modern Human Origins: Human Hunting Behavior during the Later Pleistocene (Springer Netherlands): 191–208. doi:. ISBN 978-94-007-6766-9. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Richards, Michael P.; Pettitt, Paul B.; Trinkaus, Erik; Smith, Fred H.; Paunović, Maja; Karavanić, Ivor. (2000-06-20). «Neanderthal diet at Vindija and Neanderthal predation: The evidence from stable isotopes» Proceedings of the National Academy of Sciences 97 (13): 7663–7666. doi:. ISSN 0027-8424. PMID 10852955. PMC PMC16602. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Henry, Amanda G.; Brooks, Alison S.; Piperno, Dolores R.. (2011-01-11). «Microfossils in calculus demonstrate consumption of plants and cooked foods in Neanderthal diets (Shanidar III, Iraq; Spy I and II, Belgium)» Proceedings of the National Academy of Sciences 108 (2): 486–491. doi:. ISSN 0027-8424. PMID 21187393. PMC PMC3021051. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Shipley, Gerhard P.; Kindscher, Kelly. (2016-10-24). «Evidence for the Paleoethnobotany of the Neanderthal: A Review of the Literature» Scientifica doi:. PMID 27843675. PMC PMC5098096. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Madella, Marco; Jones, Martin K.; Goldberg, Paul; Goren, Yuval; Hovers, Erella. (2002-07-01). «The Exploitation of Plant Resources by Neanderthals in Amud Cave (Israel): The Evidence from Phytolith Studies» Journal of Archaeological Science 29 (7): 703–719. doi:. ISSN 0305-4403. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Gómez-Olivencia, Asier; Sala, Nohemi; Núñez-Lahuerta, Carmen; Sanchis, Alfred; Arlegi, Mikel; Rios-Garaizar, Joseba. (2018-07-12). «First data of Neandertal bird and carnivore exploitation in the Cantabrian Region (Axlor; Barandiaran excavations; Dima, Biscay, Northern Iberian Peninsula)» Scientific Reports 8 (1): 10551. doi:. ISSN 2045-2322. (Noiz kontsultatua: 2021-02-17).
- Trekking the shore : changing coastlines and the antiquity of coastal settlement. Springer 2011 ISBN 978-1-4419-8219-3. PMC 733544112. (Noiz kontsultatua: 2021-02-15).
- Shipman, Pat. (2015). The invaders : how humans and their dogs drove Neanderthals to extinction. ISBN 978-0-674-42538-5. PMC 898113099. (Noiz kontsultatua: 2021-02-15).
- Tattersall, Ian. (2015). The strange case of the rickety cossack : and other cautionary tales from human evolution. ISBN 978-1-4668-7943-0. PMC 906030767. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Majkić, Ana; Evans, Sarah; Stepanchuk, Vadim; Tsvelykh, Alexander; d’Errico, Francesco. (2017-03-29). «A decorated raven bone from the Zaskalnaya VI (Kolosovskaya) Neanderthal site, Crimea» PLOS ONE 12 (3): e0173435. doi:. ISSN 1932-6203. PMID 28355292. PMC PMC5371307. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Hoffmann, Dirk L.; Angelucci, Diego E.; Villaverde, Valentín; Zapata, Josefina; Zilhão, João. (2018-02-01). «Symbolic use of marine shells and mineral pigments by Iberian Neandertals 115,000 years ago» Science Advances 4 (2): eaar5255. doi:. ISSN 2375-2548. PMID 29507889. PMC PMC5833998. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Moncel, M. -H.; Chiotti, L.; Gaillard, C.; Onoratini, G.; Pleurdeau, D.. (2012-03-01). «Non-utilitarian lithic objects from the European Paleolithic» Archaeology, Ethnology and Anthropology of Eurasia 40 (1): 24–40. doi:. ISSN 1563-0110. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Majkić, Ana; d’Errico, Francesco; Stepanchuk, Vadim. (2018-05-02). «Assessing the significance of Palaeolithic engraved cortexes. A case study from the Mousterian site of Kiik-Koba, Crimea» PLOS ONE 13 (5): e0195049. doi:. ISSN 1932-6203. PMID 29718916. PMC PMC5931501. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Turk, Matija; Turk, Ivan; Dimkaroski, Ljuben; Blackwell, Bonnie A. B.; Horusitzky, François Zoltán; Otte, Marcel; Bastiani, Giuliano; Korat, Lidija. (2018-09-01). «The Mousterian Musical Instrument from the Divje babe I cave (Slovenia): Arguments on the Material Evidence for Neanderthal Musical Behaviour» L'Anthropologie 122 (4): 679–706. doi:. ISSN 0003-5521. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Aubert, Maxime; Brumm, Adam; Huntley, Jillian. (2018-12-01). «Early dates for ‘Neanderthal cave art’ may be wrong» Journal of Human Evolution 125: 215–217. doi:. ISSN 0047-2484. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Pike, Alistair W. G.; Hoffmann, Dirk L.; Pettitt, Paul B.; García-Diez, Marcos; Zilhão, João. (2017-03-08). «Dating Palaeolithic cave art: Why U–Th is the way to go» Quaternary International 432: 41–49. doi:. ISSN 1040-6182. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Hoffmann, D. L.; Standish, C. D.; García-Diez, M.; Pettitt, P. B.; Milton, J. A.; Zilhão, J.; Alcolea-González, J. J.; Cantalejo-Duarte, P. et al.. (2018-02-23). «U-Th dating of carbonate crusts reveals Neandertal origin of Iberian cave art» Science 359 (6378): 912–915. doi:. ISSN 0036-8075. PMID 29472483. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Wunn, Ina. (2000). «BEGINNING OF RELIGION» Numen 47 (4): 417–452. doi:. ISSN 0029-5973. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Dediu, Dan; Levinson, Stephen C. (2018-06-01). «Neanderthal language revisited: not only us» Current Opinion in Behavioral Sciences 21: 49–55. doi:. ISSN 2352-1546. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) D’Anastasio, Ruggero; Wroe, Stephen; Tuniz, Claudio; Mancini, Lucia; Cesana, Deneb T.; Dreossi, Diego; Ravichandiran, Mayoorendra; Attard, Marie et al.. (2013-12-18). «Micro-Biomechanics of the Kebara 2 Hyoid and Its Implications for Speech in Neanderthals» PLOS ONE 8 (12): e82261. doi:. ISSN 1932-6203. PMID 24367509. PMC PMC3867335. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Stewart, J. R.; García-Rodríguez, O.; Knul, M. V.; Sewell, L.; Montgomery, H.; Thomas, M. G.; Diekmann, Y.. (2019-08-01). «Palaeoecological and genetic evidence for Neanderthal power locomotion as an adaptation to a woodland environment» Quaternary Science Reviews 217: 310–315. doi:. ISSN 0277-3791. (Noiz kontsultatua: 2021-02-15).
- Kislev, Meidad; Barkai, Ran. (2018). «Neanderthal and Woolly Mammoth Molecular Resemblance: Genetic Similarities May Underlie Cold Adaptation Suite» Human Biology 90 (2): 115–128. doi:. ISSN 0018-7143. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) de Azevedo, S.; González, M. F.; Cintas, C.; Ramallo, V.; Quinto-Sánchez, M.; Márquez, F.; Hünemeier, T.; Paschetta, C. et al.. (2017-10-30). «Nasal airflow simulations suggest convergent adaptation in Neanderthals and modern humans» Proceedings of the National Academy of Sciences 114 (47): 12442–12447. doi:. ISSN 0027-8424. PMID 29087302. PMC PMC5703271. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Rae, Todd C.; Koppe, Thomas; Stringer, Chris B.. (2011-02-01). «The Neanderthal face is not cold adapted» Journal of Human Evolution 60 (2): 234–239. doi:. ISSN 0047-2484. (Noiz kontsultatua: 2021-02-15).
- Helmuth, H.. (1998). «Body height, body mass and surface area of the Neandertals» Zeitschrift für Morphologie und Anthropologie 82 (1): 1–12. ISSN 0044-314X. (Noiz kontsultatua: 2021-02-15).
- Hominid evolution and community ecology : prehistoric human adaptation in biological perspective. Academic Press 1984 ISBN 0-12-261920-X. PMC 11456220. (Noiz kontsultatua: 2021-02-15).
- Ancestors : the hard evidence.. John Wiley 1985 ISBN 0-471-84376-8. PMC 232946395. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Amano, Hideki; Kikuchi, Takeo; Morita, Yusuke; Kondo, Osamu; Suzuki, Hiromasa; León, Marcia S. Ponce de; Zollikofer, Christoph P. E.; Bastir, Markus et al.. (2015). «Virtual reconstruction of the Neanderthal Amud 1 cranium» American Journal of Physical Anthropology 158 (2): 185–197. doi:. ISSN 1096-8644. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Juric, Ivan; Aeschbacher, Simon; Coop, Graham. (2016-11-08). «The Strength of Selection against Neanderthal Introgression» PLOS Genetics 12 (11): e1006340. doi:. ISSN 1553-7404. PMID 27824859. PMC PMC5100956. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Ruebens, Karen. (2013-10-01). «Regional behaviour among late Neanderthal groups in Western Europe: A comparative assessment of late Middle Palaeolithic bifacial tool variability» Journal of Human Evolution 65 (4): 341–362. doi:. ISSN 0047-2484. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Fabre, Virginie; Condemi, Silvana; Degioanni, Anna. (2009-04-15). «Genetic Evidence of Geographical Groups among Neanderthals» PLOS ONE 4 (4): e5151. doi:. ISSN 1932-6203. PMID 19367332. PMC PMC2664900. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Demay, Laëtitia; Péan, Stéphane; Patou-Mathis, Marylène. (2012-10-25). «Mammoths used as food and building resources by Neanderthals: Zooarchaeological study applied to layer 4, Molodova I (Ukraine)» Quaternary International 276-277: 212–226. doi:. ISSN 1040-6182. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Trinkaus, Erik. (1995-01-01). «Neanderthal mortality patterns» Journal of Archaeological Science 22 (1): 121–142. doi:. ISSN 0305-4403. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Green, Richard E.; Krause, Johannes; Briggs, Adrian W.; Maricic, Tomislav; Stenzel, Udo; Kircher, Martin; Patterson, Nick; Li, Heng et al.. (2010-05-07). «A Draft Sequence of the Neandertal Genome» Science 328 (5979): 710–722. doi:. ISSN 0036-8075. PMID 20448178. PMC PMC5100745. (Noiz kontsultatua: 2021-02-15).
- Sankararaman, Sriram; Mallick, Swapan; Patterson, Nick; Reich, David. (2016-05). «The Combined Landscape of Denisovan and Neanderthal Ancestry in Present-Day Humans» Current Biology 26 (9): 1241–1247. doi:. ISSN 0960-9822. PMID 27032491. PMC PMC4864120. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Sankararaman, Sriram; Mallick, Swapan; Dannemann, Michael; Prüfer, Kay; Kelso, Janet; Pääbo, Svante; Patterson, Nick; Reich, David. (2014-03). «The genomic landscape of Neanderthal ancestry in present-day humans» Nature 507 (7492): 354–357. doi:. ISSN 1476-4687. PMID 24476815. PMC PMC4072735. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Peyrégne, Stéphane; Slon, Viviane; Mafessoni, Fabrizio; Filippo, Cesare de; Hajdinjak, Mateja; Nagel, Sarah; Nickel, Birgit; Essel, Elena et al.. (2019-06-01). «Nuclear DNA from two early Neandertals reveals 80,000 years of genetic continuity in Europe» Science Advances 5 (6): eaaw5873. doi:. ISSN 2375-2548. PMID 31249872. PMC PMC6594762. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Kuhlwilm, Martin; Gronau, Ilan; Hubisz, Melissa J.; de Filippo, Cesare; Prado-Martinez, Javier; Kircher, Martin; Fu, Qiaomei; Burbano, Hernán A. et al.. (2016-02). «Ancient gene flow from early modern humans into Eastern Neanderthals» Nature 530 (7591): 429–433. doi:. ISSN 1476-4687. PMID 26886800. PMC PMC4933530. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Prüfer, Kay; Racimo, Fernando; Patterson, Nick; Jay, Flora; Sankararaman, Sriram; Sawyer, Susanna; Heinze, Anja; Renaud, Gabriel et al.. (2014-01). «The complete genome sequence of a Neanderthal from the Altai Mountains» Nature 505 (7481): 43–49. doi:. ISSN 1476-4687. PMID 24352235. PMC PMC4031459. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Pennisi, Elizabeth. (2013-05-17). «More Genomes From Denisova Cave Show Mixing of Early Human Groups» Science 340 (6134): 799–799. doi:. ISSN 0036-8075. PMID 23687020. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Vernot, Benjamin; Akey, Joshua M.. (2014-02-28). «Resurrecting Surviving Neandertal Lineages from Modern Human Genomes» Science 343 (6174): 1017–1021. doi:. ISSN 0036-8075. PMID 24476670. (Noiz kontsultatua: 2021-02-15).
- Nédélec, Yohann; Sanz, Joaquín; Baharian, Golshid; Szpiech, Zachary A.; Pacis, Alain; Dumaine, Anne; Grenier, Jean-Christophe; Freiman, Andrew et al.. (2016-10). «Genetic Ancestry and Natural Selection Drive Population Differences in Immune Responses to Pathogens» Cell 167 (3): 657–669.e21. doi:. ISSN 0092-8674. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Ségurel, Laure; Quintana-Murci, Lluis. (2014-10-01). «Preserving immune diversity through ancient inheritance and admixture» Current Opinion in Immunology 30: 79–84. doi:. ISSN 0952-7915. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Dolgova, Olga; Lao, Oscar. (2018/7). «Evolutionary and Medical Consequences of Archaic Introgression into Modern Human Genomes» Genes 9 (7): 358. doi:. PMID 30022013. PMC PMC6070777. (Noiz kontsultatua: 2021-02-15).
- Reich, David. (2018). Who we are and how we got here : ancient DNA and the new science of the human past. (First edition. argitaraldia) ISBN 0-19-882125-5. PMC 1006478846. (Noiz kontsultatua: 2021-02-15).
- (Ingelesez) Krause, Johannes; Fu, Qiaomei; Good, Jeffrey M.; Viola, Bence; Shunkov, Michael V.; Derevianko, Anatoli P.; Pääbo, Svante. (2010-04). «The complete mitochondrial DNA genome of an unknown hominin from southern Siberia» Nature 464 (7290): 894–897. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2023-01-27).
- Eider, Carton Virto. (2012-08-31). «Denisovako gizakiaren kalitate handiko sekuentzia genetikoa osatu dute» Zientzia.eus (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Ovodov, Nikolai D.; Crockford, Susan J.; Kuzmin, Yaroslav V.; Higham, Thomas F. G.; Hodgins, Gregory W. L.; Plicht, Johannes van der. (2011 uzt. 28). «A 33,000-Year-Old Incipient Dog from the Altai Mountains of Siberia: Evidence of the Earliest Domestication Disrupted by the Last Glacial Maximum» PLOS ONE 6 (7): e22821. doi:. ISSN 1932-6203. PMID 21829526. PMC PMC3145761. (Noiz kontsultatua: 2023-01-27).
- Ana, Galarraga Aiestara. (2019-05-02). «Denisovatik kanpoko lehen denisovartzat jo dute Tibeteko lautadan aurkitutako fosil bat» Zientzia.eus (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Prüfer, Kay; Racimo, Fernando; Patterson, Nick; Jay, Flora; Sankararaman, Sriram; Sawyer, Susanna; Heinze, Anja; Renaud, Gabriel et al.. (2014-01). «The complete genome sequence of a Neanderthal from the Altai Mountains» Nature 505 (7481): 43–49. doi:. ISSN 1476-4687. PMID 24352235. PMC PMC4031459. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Kuhlwilm, Martin; Gronau, Ilan; Hubisz, Melissa J.; de Filippo, Cesare; Prado-Martinez, Javier; Kircher, Martin; Fu, Qiaomei; Burbano, Hernán A. et al.. (2016-02). «Ancient gene flow from early modern humans into Eastern Neanderthals» Nature 530 (7591): 429–433. doi:. ISSN 1476-4687. PMID 26886800. PMC PMC4933530. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Posth, Cosimo; Wißing, Christoph; Kitagawa, Keiko; Pagani, Luca; van Holstein, Laura; Racimo, Fernando; Wehrberger, Kurt; Conard, Nicholas J. et al.. (2017-07-04). «Deeply divergent archaic mitochondrial genome provides lower time boundary for African gene flow into Neanderthals» Nature Communications 8 (1): 16046. doi:. ISSN 2041-1723. PMID 28675384. PMC PMC5500885. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Warren, Matthew. (2018-08-22). «Mum’s a Neanderthal, Dad’s a Denisovan: First discovery of an ancient-human hybrid» Nature 560 (7719): 417–418. doi:. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) «This ancient bone belonged to a child of two extinct human species» www.science.org (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Malyarchuk, B. A.. (2011-10-08). «Adaptive evolution of the Homo mitochondrial genome» Molecular Biology 45 (5): 780. doi:. ISSN 1608-3245. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Brown, P.; Sutikna, T.; Morwood, M. J.; Soejono, R. P.; Jatmiko; Wayhu Saptomo, E.; Awe Due, Rokus. (2004-10). «A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia» Nature 431 (7012): 1055–1061. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Argue, Debbie; Donlon, Denise; Groves, Colin; Wright, Richard. (2006-10-01). «Homo floresiensis: Microcephalic, pygmoid, Australopithecus, or Homo?» Journal of Human Evolution 51 (4): 360–374. doi:. ISSN 0047-2484. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Martin, Robert D.; MacLarnon, Ann M.; Phillips, James L.; Dobyns, William B.. (2006-11). «Flores hominid: New species or microcephalic dwarf?» The Anatomical Record Part A: Discoveries in Molecular, Cellular, and Evolutionary Biology 288A (11): 1123–1145. doi:. (Noiz kontsultatua: 2023-01-27).
- Callaway, Ewen. (2016-06-01). «‘Hobbit’ relatives found after ten-year hunt» Nature 534: 164–165. doi:. ISSN 0028-0836. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Brumm, Adam; van den Bergh, Gerrit D.; Storey, Michael; Kurniawan, Iwan; Alloway, Brent V.; Setiawan, Ruly; Setiyabudi, Erick; Grün, Rainer et al.. (2016-06). «Age and context of the oldest known hominin fossils from Flores» Nature 534 (7606): 249–253. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) van den Bergh, Gerrit D.; Kaifu, Yousuke; Kurniawan, Iwan; Kono, Reiko T.; Brumm, Adam; Setiyabudi, Erick; Aziz, Fachroel; Morwood, Michael J.. (2016-06). «Homo floresiensis-like fossils from the early Middle Pleistocene of Flores» Nature 534 (7606): 245–248. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Schlebusch, Carina M.; Malmström, Helena; Günther, Torsten; Sjödin, Per; Coutinho, Alexandra; Edlund, Hanna; Munters, Arielle R.; Vicente, Mário et al.. (2017-11-03). «Southern African ancient genomes estimate modern human divergence to 350,000 to 260,000 years ago» Science 358 (6363): 652–655. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) «Oldest Homo sapiens bones ever found shake foundations of the human story» the Guardian 2017-06-07 (Noiz kontsultatua: 2023-01-27).
- «Scientists Find the Skull of Humanity’s Ancestor, on a Computer - The New York Times | Ghostarchive» ghostarchive.org (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Mounier, Aurélien; Mirazón Lahr, Marta. (2019-09-10). «Deciphering African late middle Pleistocene hominin diversity and the origin of our species» Nature Communications 10 (1): 3406. doi:. ISSN 2041-1723. PMID 31506422. PMC PMC6736881. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Ambrose, Stanley H.. (1998-06-01). «Late Pleistocene human population bottlenecks, volcanic winter, and differentiation of modern humans» Journal of Human Evolution 34 (6): 623–651. doi:. ISSN 0047-2484. (Noiz kontsultatua: 2023-01-27).
- Huff, Chad D.; Xing, Jinchuan; Rogers, Alan R.; Witherspoon, David; Jorde, Lynn B.. (2010-02-01). «Mobile elements reveal small population size in the ancient ancestors of Homo sapiens» Proceedings of the National Academy of Science 107: 2147–2152. doi:. ISSN 0027-8424. (Noiz kontsultatua: 2023-01-27).
- Fernandez, Luistxo. «Haplotaldeak eta ahaideak [geno-4»] eibar.org (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Clark, Jamie L.. (2011-09-01). «The evolution of human culture during the later Pleistocene: Using fauna to test models on the emergence and nature of “modern” human behavior» Journal of Anthropological Archaeology 30 (3): 273–291. doi:. ISSN 0278-4165. (Noiz kontsultatua: 2023-01-27).
- Boyd & Silk 2003.
- Brues & Snow 1965.
- (Ingelesez) Brunet, Michel; Guy, Franck; Pilbeam, David; Mackaye, Hassane Taisso; Likius, Andossa; Ahounta, Djimdoumalbaye; Beauvilain, Alain; Blondel, Cécile et al.. (2002-07). «A new hominid from the Upper Miocene of Chad, Central Africa» Nature 418 (6894): 145–151. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) White, Tim D.; Asfaw, Berhane; Beyene, Yonas; Haile-Selassie, Yohannes; Lovejoy, C. Owen; Suwa, Gen; WoldeGabriel, Giday. (2009-10-02). «Ardipithecus ramidus and the Paleobiology of Early Hominids» Science 326 (5949): 64–86. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Ko, Kwang Hyun. (2015-Nov-Dec). «Origins of Bipedalism» Brazilian Archives of Biology and Technology 58: 929–934. doi:. ISSN 1516-8913. (Noiz kontsultatua: 2023-01-27).
- DeSalle & Tattersall 2008, 146 orr. .
- (Ingelesez) «Study Identifies Energy Efficiency As Reason For Evolution Of Upright Walking» ScienceDaily (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Sockol, Michael D.; Raichlen, David A.; Pontzer, Herman. (2007-07-24). «Chimpanzee locomotor energetics and the origin of human bipedalism» Proceedings of the National Academy of Sciences 104 (30): 12265–12269. doi:. ISSN 0027-8424. PMID 17636134. PMC PMC1941460. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Dávid-Barrett, Tamás; Dunbar, Robin I. M.. (2016-05-01). «Bipedality and hair loss in human evolution revisited: The impact of altitude and activity scheduling» Journal of Human Evolution 94: 72–82. doi:. ISSN 0047-2484. PMID 27178459. PMC PMC4874949. (Noiz kontsultatua: 2023-01-27).
- Aiello & Dean 1990.
- Kondo 1985.
- Strickberger 2000, 475–476 orr. .
- Trevathan 2011, 20 orr. .
- Curry 2008, 106–109 orr. .
- Hrdy, Sarah Blaffer. (2011). Mothers and others : the evolutionary origins of mutual understanding. (First Harvard University Press pbk. edition. argitaraldia) ISBN 978-0-674-65993-3. PMC 913785175. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Killer whales, grandmas and what men want: Evolutionary biologists consider menopause. 2013-08-19 (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Callan, Hilary, ed. (2018-10-05). The International Encyclopedia of Anthropology. (1. argitaraldia) Wiley doi:. ISBN 978-1-118-92439-6. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Welker, Frido; Ramos-Madrigal, Jazmín; Gutenbrunner, Petra; Mackie, Meaghan; Tiwary, Shivani; Rakownikow Jersie-Christensen, Rosa; Chiva, Cristina; Dickinson, Marc R. et al.. (2020-04-09). «The dental proteome of Homo antecessor» Nature 580 (7802): 235–238. doi:. ISSN 0028-0836. PMID 32269345. PMC PMC7582224. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Dennell, Robin; Roebroeks, Wil. (2005-12). «An Asian perspective on early human dispersal from Africa» Nature 438 (7071): 1099–1104. doi:. ISSN 0028-0836. (Noiz kontsultatua: 2023-01-27).
- (Ingelesez) Schoenemann, P. Thomas. (2006-10-01). «Evolution of the Size and Functional Areas of the Human Brain» Annual Review of Anthropology 35 (1): 379–406. doi:. ISSN 0084-6570. (Noiz kontsultatua: 2023-01-28).
- «Human Evolution» tolweb.org (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Kaplan, H.; Hill, K.; Lancaster, J.; Hurtado, A.. (2000). «A theory of human life history evolution: Diet, intelligence, and longevity» Evolutionary Anthropology: Issues (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Park, Min S.; Nguyen, Andrew D.; Aryan, Henry E.; U, Hoi Sang; Levy, Michael L.; Semendeferi, Katerina. (2007-03). «EVOLUTION OF THE HUMAN BRAIN: CHANGING BRAIN SIZE AND THE FOSSIL RECORD» Neurosurgery 60 (3): 555. doi:. ISSN 0148-396X. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Bruner, Emiliano. (2007-12-01). «Cranial shape and size variation in human evolution: structural and functional perspectives» Child's Nervous System 23 (12): 1357–1365. doi:. ISSN 1433-0350. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Potts, Richard. (2012-10-21). «Evolution and Environmental Change in Early Human Prehistory» Annual Review of Anthropology 41 (1): 151–167. doi:. ISSN 0084-6570. (Noiz kontsultatua: 2023-01-28).
- «For Evolving Brains, a ‘Paleo’ Diet of Carbs - The New York Times | Ghostarchive» ghostarchive.org (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Mann, Neil. (2007-09). «Meat in the human diet: An anthropological perspective» Nutrition & Dietetics 64 (s4 The Role of): S102–S107. doi:. ISSN 1446-6368. (Noiz kontsultatua: 2023-01-28).
- «06.14.99 - Meat-eating was essential for human evolution, says UC Berkeley anthropologist specializing in diet» newsarchive.berkeley.edu (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Barr, W. Andrew; Pobiner, Briana; Rowan, John; Du, Andrew; Faith, J. Tyler. (2022-02). «No sustained increase in zooarchaeological evidence for carnivory after the appearance of Homo erectus» Proceedings of the National Academy of Sciences 119 (5): e2115540119. doi:. ISSN 0027-8424. PMID 35074877. PMC PMC8812535. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Organ, Chris; Nunn, Charles L.; Machanda, Zarin; Wrangham, Richard W.. (2011-08-30). «Phylogenetic rate shifts in feeding time during the evolution of Homo» Proceedings of the National Academy of Sciences 108 (35): 14555–14559. doi:. ISSN 0027-8424. PMID 21873223. PMC PMC3167533. (Noiz kontsultatua: 2023-01-28).
- Dávid-Barrett, T.; Dunbar, R. I. M.. (2013-08-22). «Processing power limits social group size: computational evidence for the cognitive costs of sociality» Proceedings of the Royal Society B: Biological Sciences 280 (1765): 20131151. doi:. PMID 23804623. PMC PMC3712454. (Noiz kontsultatua: 2023-01-28).
- Bown & Rose 1987.
- (Ingelesez) Barton, Robert A.; Venditti, Chris. (2014-10-20). «Rapid Evolution of the Cerebellum in Humans and Other Great Apes» Current Biology 24 (20): 2440–2444. doi:. ISSN 0960-9822. PMID 25283776. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Starowicz-Filip, Anna; Milczarek, Olga; Kwiatkowski, Stanisław; Bętkowska-Korpała, Barbara; Prochwicz, Katarzyna. (2013-09-02). «“Cerebellar cognitive affective syndrome CCAS – a case report”» Archives of Psychiatry and Psychotherapy 15 (3): 57–64. doi:. ISSN 1509-2046. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Weaver, Anne H.. (2005-03-08). «Reciprocal evolution of the cerebellum and neocortex in fossil humans» Proceedings of the National Academy of Sciences 102 (10): 3576–3580. doi:. ISSN 0027-8424. PMID 15731345. PMC PMC553338. (Noiz kontsultatua: 2023-01-28).
- Yu, Feng; Jiang, Qing-jun; Sun, Xi-yan; Zhang, Rong-wei. (2015-06). «A new case of complete primary cerebellar agenesis: clinical and imaging findings in a living patient» Brain: A Journal of Neurology 138 (Pt 6): e353. doi:. ISSN 1460-2156. PMID 25149410. PMC 4614135. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Graham, Kirsty E.; Hobaiter, Catherine. (2023-01-24). de Waal, Frans B. M. ed. «Towards a great ape dictionary: Inexperienced humans understand common nonhuman ape gestures» PLOS Biology 21 (1): e3001939. doi:. ISSN 1545-7885. PMID 36693024. PMC PMC9873169. (Noiz kontsultatua: 2023-02-01).
- Klein, Stefan. (2014). Survival of the nicest : how altruism made us human and why it pays to get along. ISBN 978-1-61519-090-4. PMC 842880031. (Noiz kontsultatua: 2023-01-28).
- Sallet, J.; Mars, R. B.; Noonan, M. P.; Andersson, J. L.; O'Reilly, J. X.; Jbabdi, S.; Croxson, P. L.; Jenkinson, M. et al.. (2011-11-01). «Social Network Size Affects Neural Circuits in Macaques» Science 334: 697. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Dunbar, R. I. M.. (1992-06-01). «Neocortex size as a constraint on group size in primates» Journal of Human Evolution 22 (6): 469–493. doi:. ISSN 0047-2484. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Shultz, Susanne; Dunbar, Robin. (2010-12-14). «Encephalization is not a universal macroevolutionary phenomenon in mammals but is associated with sociality» Proceedings of the National Academy of Sciences 107 (50): 21582–21586. doi:. ISSN 0027-8424. PMID 21098277. PMC PMC3003036. (Noiz kontsultatua: 2023-01-28).
- Potts, R.. (1998). «Environmental hypotheses of hominin evolution» American Journal of Physical Anthropology Suppl 27: 93–136. doi:. ISSN 0002-9483. PMID 9881524. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Mesoudi, Alex; Whiten, Andrew; Laland, Kevin N.. (2006-08). «Towards a unified science of cultural evolution» Behavioral and Brain Sciences 29 (4): 329–347. doi:. ISSN 0140-525X. (Noiz kontsultatua: 2023-01-28).
- Handbook of paleoanthropology. (Second edition. argitaraldia) 2015 ISBN 978-3-642-39979-4. PMC 900727331. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Zeitoun, Valery. (2003-09). «High occurrence of a basicranial feature inHomo erectus: Anatomical description of the preglenoid tubercle» The Anatomical Record 274B (1): 148–156. doi:. ISSN 0003-276X. (Noiz kontsultatua: 2023-01-28).
- Tanner, Nancy Makepeace. (1981). On becoming human. Cambridge University Press ISBN 0-521-23554-5. PMC 6707544. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Reno, Philip L.; Lovejoy, C. Owen. (2015-04-28). «From Lucy to Kadanuumuu: balanced analyses of Australopithecus afarensis assemblages confirm only moderate skeletal dimorphism» PeerJ 3: e925. doi:. ISSN 2167-8359. PMID 25945314. PMC PMC4419524. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Lovejoy, C. Owen. (2009-10-02). «Reexamining Human Origins in Light of Ardipithecus ramidus» Science 326 (5949): 74. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Young, Richard W.. (2003-01). «Evolution of the human hand: the role of throwing and clubbing» Journal of Anatomy 202 (1): 165–174. doi:. ISSN 0021-8782. PMID 12587931. PMC PMC1571064. (Noiz kontsultatua: 2023-01-28).
- Ittyerah, Miriam. (2013). Hand preference and hand ability : evidence from studies in haptic cognition. ISBN 978-90-272-7164-8. PMC 858981530. (Noiz kontsultatua: 2023-01-28).
- «The Hand» archive.nytimes.com (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Kuzawa, Christopher W.; Chugani, Harry T.; Grossman, Lawrence I.; Lipovich, Leonard; Muzik, Otto; Hof, Patrick R.; Wildman, Derek E.; Sherwood, Chet C. et al.. (2014-09-09). «Metabolic costs and evolutionary implications of human brain development» Proceedings of the National Academy of Sciences 111 (36): 13010–13015. doi:. ISSN 0027-8424. PMID 25157149. PMC PMC4246958. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Pontzer, Herman; Brown, Mary H.; Raichlen, David A.; Dunsworth, Holly; Hare, Brian; Walker, Kara; Luke, Amy; Dugas, Lara R. et al.. (2016-05). «Metabolic acceleration and the evolution of human brain size and life history» Nature 533 (7603): 390–392. doi:. ISSN 1476-4687. PMID 27144364. PMC PMC4942851. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Rantala, M. J.. (2007-09). «Evolution of nakedness in Homo sapiens» Journal of Zoology 273 (1): 1–7. doi:. ISSN 0952-8369. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Mcbrearty, Sally; Brooks, Alison S.. (2000-11-01). «The revolution that wasn't: a new interpretation of the origin of modern human behavior» Journal of Human Evolution 39 (5): 453–563. doi:. ISSN 0047-2484. (Noiz kontsultatua: 2023-01-28).
- Diamond 1999, 39 orr. .
- (Ingelesez) Bar-Yosef, Ofer. (2002-10). «The Upper Paleolithic Revolution» Annual Review of Anthropology 31 (1): 363–393. doi:. ISSN 0084-6570. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Henshilwood, Christopher S.; d'Errico, Francesco; Yates, Royden; Jacobs, Zenobia; Tribolo, Chantal; Duller, Geoff A. T.; Mercier, Norbert; Sealy, Judith C. et al.. (2002-02-15). «Emergence of Modern Human Behavior: Middle Stone Age Engravings from South Africa» Science 295 (5558): 1278–1280. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Henshilwood, Christopher S.; Marean, Curtis W.. (2003-12). «The Origin of Modern Human Behavior: Critique of the Models and Their Test Implications» Current Anthropology 44 (5): 627–651. doi:. ISSN 0011-3204. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Backwell, Lucinda; d'Errico, Francesco; Wadley, Lyn. (2008-06-01). «Middle Stone Age bone tools from the Howiesons Poort layers, Sibudu Cave, South Africa» Journal of Archaeological Science 35 (6): 1566–1580. doi:. ISSN 0305-4403. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Nowell, April. (2010-10-21). «Defining Behavioral Modernity in the Context of Neandertal and Anatomically Modern Human Populations» Annual Review of Anthropology 39 (1): 437–452. doi:. ISSN 0084-6570. (Noiz kontsultatua: 2023-01-28).
- d'Errico, Francesco; Stringer, Chris B.. (2011-04-12). «Evolution, revolution or saltation scenario for the emergence of modern cultures?» Philosophical Transactions of the Royal Society B: Biological Sciences 366 (1567): 1060–1069. doi:. PMID 21357228. PMC PMC3049097. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Yong, Ed. (2018-03-15). «A Cultural Leap at the Dawn of Humanity» The Atlantic (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Brooks, Alison S.; Yellen, John E.; Potts, Richard; Behrensmeyer, Anna K.; Deino, Alan L.; Leslie, David E.; Ambrose, Stanley H.; Ferguson, Jeffrey R. et al.. (2018-04-06). «Long-distance stone transport and pigment use in the earliest Middle Stone Age» Science 360 (6384): 90–94. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Rasmussen, Morten; Guo, Xiaosen; Wang, Yong; Lohmueller, Kirk E.; Rasmussen, Simon; Albrechtsen, Anders; Skotte, Line; Lindgreen, Stinus et al.. (2011-10-07). «An Aboriginal Australian Genome Reveals Separate Human Dispersals into Asia» Science 334 (6052): 94–98. doi:. ISSN 0036-8075. PMID 21940856. PMC PMC3991479. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Ambrose, Stanley H.. (2001-03-02). «Paleolithic Technology and Human Evolution» Science 291 (5509): 1748–1753. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Mellars, Paul. (2010-11-23). «Neanderthal symbolism and ornament manufacture: The bursting of a bubble?» Proceedings of the National Academy of Sciences 107 (47): 20147–20148. doi:. ISSN 0027-8424. PMID 21078972. PMC PMC2996706. (Noiz kontsultatua: 2023-01-28).
- Rubin, Paul H.. (2003-07). «Folk Economics» Southern Economic Journal 70 (1): 157. doi:. ISSN 0038-4038. (Noiz kontsultatua: 2022-06-02).
- Bowlby, John. (1969). Attachment. Basic Books ISBN 0-465-09715-4. PMC 7111186. (Noiz kontsultatua: 2022-06-02).
- Dodge, John Vilas, (25 Sept. 1909–23 April 1991), Senior Editorial Consultant, Encyclopædia Britannica, since 1972; Chairman, Board of Editors, Encyclopædia Britannica Publishers, since 1977. Oxford University Press 2007-12-01 (Noiz kontsultatua: 2022-06-02).
- (Ingelesez) Narvaez, Darcia; Gleason, Tracy; Wang, Lijuan; Brooks, Jeff; Lefever, Jennifer Burke; Cheng, Ying. (2013-10-01). «The evolved development niche: Longitudinal effects of caregiving practices on early childhood psychosocial development» Early Childhood Research Quarterly 28 (4): 759–773. doi:. ISSN 0885-2006. (Noiz kontsultatua: 2022-06-06).
- Narvaez, Darcia; Wang, Lijuan; Gleason, Tracy; Cheng, Ying; Lefever, Jennifer; Deng, Lifang. (2013-03-01). «The evolved developmental niche and child sociomoral outcomes in Chinese 3-year-olds» European Journal of Developmental Psychology 10 (2): 106–127. doi:. ISSN 1740-5629. (Noiz kontsultatua: 2022-06-06).
- (Ingelesez) Öhman, Arne; Mineka, Susan. (2001). «Fears, phobias, and preparedness: Toward an evolved module of fear and fear learning.» Psychological Review 108 (3): 483–522. doi:. ISSN 1939-1471. (Noiz kontsultatua: 2022-06-06).
- (Ingelesez) Pinker, Steven. (1999-06). «How the Mind Works» Annals of the New York Academy of Sciences 882 (1 GREAT ISSUES): 119–127. doi:. ISSN 0077-8923. (Noiz kontsultatua: 2022-06-06).
- (Ingelesez) Hagen, Edward H.; Hammerstein, Peter. (2006-05-01). «Game theory and human evolution: A critique of some recent interpretations of experimental games» Theoretical Population Biology 69 (3): 339–348. doi:. ISSN 0040-5809. (Noiz kontsultatua: 2022-06-06).
- Tapper, Katy. (2008-10). «Book Review: Deirdre Barrett Waistland. The (R)evolutionary Science Behind Our Weight and Fitness Crisis. New York: W.W. Norton & Company, 2007, 320 pp. $24.95 (hbk.); ISBN: 978-0-393-06216-8» Journal of Health Psychology 13 (7): 968–969. doi:. ISSN 1359-1053. (Noiz kontsultatua: 2022-06-02).
- Vugt, Mark van. (2011). Naturally selected : the evolutionary science of leadership. (1st U.S. ed. argitaraldia) HarperBusiness ISBN 978-0-06-196383-4. PMC 548626461. (Noiz kontsultatua: 2022-06-06).
- (Ingelesez) Vugt, Mark van; Ronay, Richard. (2014-02). «The evolutionary psychology of leadership: Theory, review, and roadmap» Organizational Psychology Review 4 (1): 74–95. doi:. ISSN 2041-3866. (Noiz kontsultatua: 2022-06-06).
- (Ingelesez) Jabr, Ferris. «Does Thinking Really Hard Burn More Calories?» Scientific American (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Gibbons, Ann. (1998-05-29). «Solving the Brain's Energy Crisis» Science 280 (5368): 1345–1347. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2023-01-28).
- Robinson 2008, 398 orr. .
- (Ingelesez) Semaw, Sileshi; Rogers, Michael J; Quade, Jay; Renne, Paul R; Butler, Robert F; Dominguez-Rodrigo, Manuel; Stout, Dietrich; Hart, William S et al.. (2003-08-01). «2.6-Million-year-old stone tools and associated bones from OGS-6 and OGS-7, Gona, Afar, Ethiopia» Journal of Human Evolution 45 (2): 169–177. doi:. ISSN 0047-2484. (Noiz kontsultatua: 2023-01-28).
- Freeman & Herron 2007, 786–788 orr. .
- (Ingelesez) Ward, Carol V.; Tocheri, Matthew W.; Plavcan, J. Michael; Brown, Francis H.; Manthi, Fredrick Kyalo. (2014-01-07). «Early Pleistocene third metacarpal from Kenya and the evolution of modern human-like hand morphology» Proceedings of the National Academy of Sciences 111 (1): 121–124. doi:. ISSN 0027-8424. PMID 24344276. PMC PMC3890866. (Noiz kontsultatua: 2023-01-28).
- Cochran & Harpending 2009.
- (Ingelesez) Bender, Andrea. (2020-10). «The Role of Culture and Evolution for Human Cognition» Topics in Cognitive Science 12 (4): 1403–1420. doi:. ISSN 1756-8757. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Yi, Xin; Liang, Yu; Huerta-Sanchez, Emilia; Jin, Xin; Cuo, Zha Xi Ping; Pool, John E.; Xu, Xun; Jiang, Hui et al.. (2010-07-02). «Sequencing of 50 Human Exomes Reveals Adaptation to High Altitude» Science 329 (5987): 75–78. doi:. ISSN 0036-8075. PMID 20595611. PMC PMC3711608. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Jeong, Choongwon; Alkorta-Aranburu, Gorka; Basnyat, Buddha; Neupane, Maniraj; Witonsky, David B.; Pritchard, Jonathan K.; Beall, Cynthia M.; Di Rienzo, Anna. (2014-02-10). «Admixture facilitates genetic adaptations to high altitude in Tibet» Nature Communications 5 (1): 3281. doi:. ISSN 2041-1723. PMID 24513612. PMC PMC4643256. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) «Brain disease 'resistance gene' evolves in Papua New Guinea community; could offer insights into CJD» ScienceDaily (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Mead, Simon; Whitfield, Jerome; Poulter, Mark; Shah, Paresh; Uphill, James; Campbell, Tracy; Al-Dujaily, Huda; Hummerich, Holger et al.. (2009-11-19). «A Novel Protective Prion Protein Variant that Colocalizes with Kuru Exposure» New England Journal of Medicine 361 (21): 2056–2065. doi:. ISSN 0028-4793. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Corbett, Stephen; Morin-Papunen, Laure. (2013-07-05). «The Polycystic Ovary Syndrome and recent human evolution» Molecular and Cellular Endocrinology 373 (1): 39–50. doi:. ISSN 0303-7207. (Noiz kontsultatua: 2023-01-28).
- Diamond 1999.
- (Ingelesez) Courtiol, Alexandre; Tropf, Felix C.; Mills, Melinda C.. (2016-07-12). «When genes and environment disagree: Making sense of trends in recent human evolution» Proceedings of the National Academy of Sciences 113 (28): 7693–7695. doi:. ISSN 0027-8424. PMID 27402758. PMC PMC4948334. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Markov, Alexander V.; Markov, Mikhail A.. (2020-06). «Runaway brain‐culture coevolution as a reason for larger brains: Exploring the “cultural drive” hypothesis by computer modeling» Ecology and Evolution 10 (12): 6059–6077. doi:. ISSN 2045-7758. PMID 32607213. PMC PMC7319167. (Noiz kontsultatua: 2023-01-28).
- Stout, Dietrich. (2011-04-12). «Stone toolmaking and the evolution of human culture and cognition» Philosophical Transactions of the Royal Society B: Biological Sciences 366 (1567): 1050–1059. doi:. PMID 21357227. PMC PMC3049103. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Byars, Sean G.; Ewbank, Douglas; Govindaraju, Diddahally R.; Stearns, Stephen C.. (2010-01-26). «Natural selection in a contemporary human population» Proceedings of the National Academy of Sciences 107 (suppl_1): 1787–1792. doi:. ISSN 0027-8424. PMID 19858476. PMC PMC2868295. (Noiz kontsultatua: 2023-01-28).
- Stanford, Allen & Antón 2009.
- (Ingelesez) Wood, Bernard; Richmond, Brian G.. (2000-07). «Human evolution: taxonomy and paleobiology» Journal of Anatomy 197 (1): 19–60. doi:. ISSN 0021-8782. PMID 10999270. PMC PMC1468107. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Varki, Ajit; Nelson, David L.. (2007-09-01). «Genomic Comparisons of Humans and Chimpanzees» Annual Review of Anthropology 36 (1): 191–209. doi:. ISSN 0084-6570. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Sayers, Ken; Raghanti, Mary Ann; Lovejoy, C. Owen. (2012-10-21). «Human Evolution and the Chimpanzee Referential Doctrine» Annual Review of Anthropology 41 (1): 119–138. doi:. ISSN 0084-6570. (Noiz kontsultatua: 2023-01-28).
- Dawkins 2004.
- (Ingelesez) Ruvolo, M.. (1997-10). «Genetic Diversity in Hominoid Primates» Annual Review of Anthropology 26 (1): 515–540. doi:. ISSN 0084-6570. (Noiz kontsultatua: 2023-01-28).
- academic.oup.com doi:. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Patterson, Nick; Richter, Daniel J.; Gnerre, Sante; Lander, Eric S.; Reich, David. (2006-06). «Genetic evidence for complex speciation of humans and chimpanzees» Nature 441 (7097): 1103–1108. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Begun, David R.. (2010-10-21). «Miocene Hominids and the Origins of the African Apes and Humans» Annual Review of Anthropology 39 (1): 67–84. doi:. ISSN 0084-6570. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Wood, Bernard. (1996-12). «Human evolution» BioEssays 18 (12): 945–954. doi:. ISSN 0265-9247. (Noiz kontsultatua: 2023-01-28).
- More word histories and mysteries : from aardvark to zombie. Houghton Mifflin 2006 ISBN 0-618-71681-5. PMC 70199867. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Nested Hierarchies, the Order of Nature: Carolus Linnaeus - Understanding Evolution. 2021-05-12 (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) THE BOOK OF Popular Science. 1963 (Noiz kontsultatua: 2023-01-28).
- Darwin 1859, 367–404 orr. .
- Darwin 1981.
- (Ingelesez) Shipman, Pat; Storm, Paul. (2002-06-13). «Missing links: Eugène Dubois and the origins of paleoanthropology» Evolutionary Anthropology: Issues, News, and Reviews 11 (3): 108–116. doi:. (Noiz kontsultatua: 2023-01-28).
- Dart, Raymond A.. (1925-02-01). «Australopithecus africanus: The Man-Ape of South Africa» Nature 115: 195–199. doi:. ISSN 0028-0836. (Noiz kontsultatua: 2023-01-28).
- Cartmill, Matt. (2009). The human lineage. Wiley-Blackwell ISBN 978-0-471-21491-5. PMC 294066337. (Noiz kontsultatua: 2023-01-28).
- Johanson & Edey 1981, 22 orr. .
- Berger, Lee R; Hawks, John; de Ruiter, Darryl J; Churchill, Steven E; Schmid, Peter; Delezene, Lucas K; Kivell, Tracy L; Garvin, Heather M et al.. (2015-09-10). Krause, Johannes ed. «Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa» eLife 4: e09560. doi:. ISSN 2050-084X. PMID 26354291. PMC PMC4559886. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) «National Geographic» National Geographic (Noiz kontsultatua: 2023-01-28).
- Dirks 2017.
- Dirks, Paul HGM; Roberts, Eric M; Hilbert-Wolf, Hannah; Kramers, Jan D; Hawks, John; Dosseto, Anthony; Duval, Mathieu; Elliott, Marina et al.. (2017-05-09). Perry, George H ed. «The age of Homo naledi and associated sediments in the Rising Star Cave, South Africa» eLife 6: e24231. doi:. ISSN 2050-084X. PMID 28483040. PMC PMC5423772. (Noiz kontsultatua: 2023-01-28).
- (Ingelesez) Sarich, Vincent M.; Wilson, Allan C.. (1967-12). «Immunological Time Scale for Hominid Evolution» Science 158 (3805): 1200–1203. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2023-01-28).
- M'charek 2005, 96 orr. .
- Trent 2005, 6 orr. .
- (Ingelesez) López, Saioa; Van Dorp, Lucy; Hellenthal, Garrett. (2015-01). «Human Dispersal Out of Africa: A Lasting Debate» Evolutionary Bioinformatics 11s2: EBO.S33489. doi:. ISSN 1176-9343. PMID 27127403. PMC PMC4844272. (Noiz kontsultatua: 2022-01-11).
- History alive! : the ancient world.. Teachers' Curriculum Institute 2004 ISBN 1-58371-350-6. PMC 55896144. (Noiz kontsultatua: 2022-01-11).
- (Ingelesez) Harvati, Katerina; Röding, Carolin; Bosman, Abel M.; Karakostis, Fotios A.; Grün, Rainer; Stringer, Chris; Karkanas, Panagiotis; Thompson, Nicholas C. et al.. (2019-07). «Apidima Cave fossils provide earliest evidence of Homo sapiens in Eurasia» Nature 571 (7766): 500–504. doi:. ISSN 0028-0836. (Noiz kontsultatua: 2022-01-11).
- (Ingelesez) Stringer, Chris. (2012-05). «What makes a modern human» Nature 485 (7396): 33–35. doi:. ISSN 0028-0836. (Noiz kontsultatua: 2022-01-11).
- (Ingelesez) Callaway, Ewen. (2018-02). «Israeli fossils are the oldest modern humans ever found outside of Africa» Nature 554 (7690): 15–16. doi:. ISSN 0028-0836. (Noiz kontsultatua: 2022-01-11).
- (Ingelesez) Zimmer, Carl. (2019-07-10). «A Skull Bone Discovered in Greece May Alter the Story of Human Prehistory» The New York Times ISSN 0362-4331. (Noiz kontsultatua: 2022-01-11).
- (Ingelesez) «'Oldest remains' outside Africa reset human migration clock» phys.org (Noiz kontsultatua: 2022-01-11).
- (Ingelesez) Lohse, Konrad; Frantz, Laurent A F. (2014-04-01). «Neandertal Admixture in Eurasia Confirmed by Maximum-Likelihood Analysis of Three Genomes» Genetics 196 (4): 1241–1251. doi:. ISSN 1943-2631. PMID 24532731. PMC PMC3982695. (Noiz kontsultatua: 2022-01-11).
- (Ingelesez) Reich, David; Green, Richard E.; Kircher, Martin; Krause, Johannes; Patterson, Nick; Durand, Eric Y.; Viola, Bence; Briggs, Adrian W. et al.. (2010-12). «Genetic history of an archaic hominin group from Denisova Cave in Siberia» Nature 468 (7327): 1053–1060. doi:. ISSN 0028-0836. PMID 21179161. PMC PMC4306417. (Noiz kontsultatua: 2022-01-11).
- (Ingelesez) Hammer, M. F.; Woerner, A. E.; Mendez, F. L.; Watkins, J. C.; Wall, J. D.. (2011-09-13). «Genetic evidence for archaic admixture in Africa» Proceedings of the National Academy of Sciences 108 (37): 15123–15128. doi:. ISSN 0027-8424. PMID 21896735. PMC PMC3174671. (Noiz kontsultatua: 2022-01-11).
Bibliografia
- Aiello, Leslie; Dean, Christopher. (1990). An Introduction to Human Evolutionary Anatomy. London; San Diego: Elsevier Academic Press ISBN 978-0-12-045591-1. OCLC .33408268.
- Bogin, Barry. (1997). «The Evolution of Human Nutrition» The Anthropology of Medicine: From Culture to Method. (3rd. argitaraldia) Westport, CT: Bergin & Garvey ISBN 978-0-89789-516-3. OCLC .36165190.
- Bown, Thomas M.; Rose, Kenneth D.. (1987). Patterns of Dental Evolution in Early Eocene Anaptomorphine Primates (Omomyidae) From the Bighorn Basin, Wyoming. 23 Tulsa, OK: Paleontological Society OCLC .16997265.
- Boyd, Robert; Silk, Joan B.. (2003). How Humans Evolved. (3rd. argitaraldia) New York: W.W. Norton & Company ISBN 978-0-393-97854-4. OCLC .49959461. (9th edizioa 2021)
- Boyd, Robert; Silk, Joan B.. (2021). How Humans Evolved. (9th. argitaraldia) New York: W.W. Norton & Company ISBN 978-0-393-53315-6. OCLC .1282924892.
- Brues, Alice M.; Snow, Clyde C.. (1965). «Physical Anthropology» Biennial Review of Anthropology 1965. 4 Stanford, CA: Stanford University Press ISBN 978-0-8047-1746-5. ISSN 0067-8503. OCLC .01532912.
- Bryson, Bill. (2004). «The Mysterious Biped» A Short History of Nearly Everything. Toronto: Anchor Canada ISBN 978-0-385-66004-4. OCLC .55016591.
- Cameron, David W.. (2004). Hominid Adaptations and Extinctions. Sydney, NSW: UNSW Press ISBN 978-0-86840-716-6. OCLC .57077633.
- Cochran, Gregory; Harpending, Henry. (2009). The 10,000 urtea Explosion: How Civilization Accelerated Human Evolution. New York: Basic Books ISBN 978-0-465-00221-4. OCLC .191926088.
- Curry, James R.. (2008). Children of God: Children of Earth. Bloomington, IN: AuthorHouse ISBN 978-1-4389-1846-4. OCLC .421466369.
- Darwin, Charles. (1859). On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life. (1st. argitaraldia).
- Darwin, Charles. (1981). The Descent of Man, and Selection in Relation to Sex. (Reprint. argitaraldia) Princeton, NJ: Princeton University Press ISBN 978-0-691-02369-4. OCLC .7197127.
- Dawkins, Richard. (2004). The Ancestor's Tale: A Pilgrimage to the Dawn of Evolution. Boston, MA: Houghton Mifflin ISBN 978-0-618-00583-3. OCLC .56617123.
- DeSalle, Rob; Tattersall, Ian. (2008). Human Origins: What Bones and Genomes Tell Us About Ourselves. 13 (1st. argitaraldia) College Station, TX: Texas A&M University Press ISBN 978-1-58544-567-7. OCLC .144520427.
- Diamond, Jared. (1999). Guns, Germs, and Steel: The Fates of Human Societies. New York: W.W. Norton & Company ISBN 978-0-393-31755-8. OCLC .35792200.
- Freeman, Scott; Herron, Jon C.. (2007). Evolutionary Analysis. (4th. argitaraldia) Upper Saddle River, NJ: Pearson Prentice Hall ISBN 978-0-13-227584-2. OCLC .73502978.
- Johanson, Donald; Edey, Maitland. (1981). Lucy, the Beginnings of Humankind. Granada ISBN 978-0-586-08437-3..
- Montgomery, William M.. (1988). «Germany» The Comparative Reception of Darwinism. Chicago, Illinois: University of Chicago Press ISBN 978-0-226-29977-8. OCLC .17328115. "The Conference on the Comparative Reception of Darwinism was held in Austin, Texas, on April 22 and 23, 1972, under the joint sponsorship of the American Council of Learned Societies and the University of Texas at Austin"
- «Primate Morphophysiology, Locomotor Analyses, and Human Bipedalism» American aldizkaria of Physical Anthropology (Tokyo: University of Tokyo Press) 702: 278–279. doi:. ISBN 978-4-13-066093-8. OCLC .12352830.
- Leakey, Richard E.. (1994). The Origin of Humankind. New York: Basic Books ISBN 978-0-465-03135-1. OCLC .30739453.
- M'charek, Amade. (2005). The Human Genome Diversity Project: An Ethnography of Scientific Practice. Cambridge; New York: Cambridge University Press ISBN 978-0-521-83222-9. OCLC .55600894.
- Martin, Robert D.. (2001). «Primates, Evolution of» (1st. argitaraldia) Amsterdam; New York: Elsevier, 12032–12038 or. doi:. ISBN 978-0-08-043076-8. OCLC .47869490.
- Maxwell, Mary. (1984). Human Evolution: A Philosophical Anthropology. New York: Columbia University Press ISBN 978-0-231-05946-6. OCLC .10163036.
- McHenry, Henry M.. (2009). «Human Evolution» Evolution: The izena Four Billion urteas. Cambridge, Massachusetts: Belknap Press of Harvard University Press ISBN 978-0-674-03175-3. OCLC .225874308.
- Ramachandran, Sohini. (2010). «Genetics and Genomics of Human Population Structure» Vogel and Motulsky's Human Genetics: Problems and Approaches. (4th completely rev.. argitaraldia) Heidelberg; London; New York: Springer doi:. ISBN 978-3-540-37653-8. OCLC .549541244.
- Robinson, J.T.. (2008). «Adaptive Radiation in the Australopithecines and the Origin of Man» African Ecology and Human Evolution. New Brunswick, NJ: Transaction argitaletxeas ISBN 978-0-202-36136-9. OCLC .144770218.
- Srivastava, R.P.. (2009). Morphology of the Primates and Human Evolution. New Delhi: PHI Learning Private Limited ISBN 978-81-203-3656-8. OCLC .423293609.
- Stanford, Craig; Allen, John S.; Antón, Susan C.. (2009). Biological Anthropology: The Natural History of Humankind. (2nd. argitaraldia) Upper Saddle River, NJ: Pearson Prentice Hall ISBN 978-0-13-601160-6. OCLC .187548835.
- Strickberger, Monroe W.. (2000). Evolution. (3rd. argitaraldia) Sudbury, MA: Jones and Bartlett argitaletxeas ISBN 978-0-7637-1066-8. OCLC .41431683.
- Stringer, Chris B.. (1994). «Evolution of Early Humans» The Cambridge Encyclopedia of Human Evolution. (1st paperback. argitaraldia) Cambridge, UK: Cambridge University Press ISBN 978-0-521-32370-3. OCLC .444512451.
- Swisher, Carl C., III; Curtis, Garniss H.; Lewin, Roger. (2001). Java Man: How Two Geologists Changed Our Understanding of Human Evolution. Chicago: University of Chicago Press ISBN 978-0-226-78734-3. OCLC .48066180.
- Trent, Ronald J.. (2005). Molecular Medicine: An Introductory Text. (3rd. argitaraldia) Burlington, MA: Elsevier Academic Press ISBN 978-0-12-699057-7. OCLC .162577235.
- Trevathan, Wenda R.. (2011). Human Birth: An Evolutionary Perspective. New Brunswick, NJ: Transaction argitaletxeas ISBN 978-1-4128-1502-4. OCLC .669122326.
- Human Diet: Its Origin and Evolution. Westport, CT: Bergin & Garvey ISBN 978-0-89789-736-5. OCLC .537239907.
- Walker, Alan. (2007). «Early Hominin Diets: Overview and Historical Perspectives» Evolution of the Human Diet: The Known, the Unknown, and the Unknowable. Oxford; New York: Oxford University Press ISBN 978-0-19-518346-7. OCLC .132816551.
- Wallace, David Rains. (2004). Beasts of Eden: Walking Whales, Dawn Horses, and Other Enigmas of Mammal Evolution. Berkeley: University of California Press ISBN 978-0-520-24684-3. OCLC .53254011.
- Webster, Donovan. (2010). Meeting the Family: One Man's Journey Through His Human Ancestry. Washington, DC: National Geographic Society ISBN 978-1-4262-0573-6. OCLC .429022321.
- Wood, Bernard A.. (2009). «Where Does the Genus Homo Begin, and How Would We Know?» The izena Humans: Origin and Early Evolution of the Genus Homo. Dordrecht, the Netherlands: Springer Netherlands, 17–28 or. doi:. ISBN 978-1-4020-9979-3. ISSN 1877-9077. OCLC .310400980. "Contributions from the Third Stony Brook Human Evolution Symposium and Workshop October 3–7, 2006."
Ikus, gainera
Kanpo estekak
- Giza eboluzioari buruzko zazpi bideo Youtuben.
- Giza eboluzioa berridazten, Amaia Portugalen artikulua Berrian, 2013/09/24.
| Autoritate kontrola |
|
|---|
 Datuak: Q83944
Datuak: Q83944 Multimedia: Human evolution / Q83944
Multimedia: Human evolution / Q83944