Ozpin-euli
Drosophila melanogaster euli-espezie bat da, Diptera ordenakoa eta Drosophilidae familiakoa. Oro har, ozpin-euli edota fruta-euli[1] esaten zaio; izan ere, hartzidura fasean dauden fruituez elikatzen da. Charles W. Woodworth entomologo amerikarrak euli-espezie hau organismo eredu moduan erabiltzeko proposatu zuenetik, D. melanogaster oso erabilia izan da zenbait arlotan genetikari buruzko ikerketak egiteko, besteak beste, biologian, fisiologian, mikroorganismoen patogenesian eta biziaren historiaren garapenean. Halaber, hainbat izan dira D. melanogaster euliarekin egindako ikerketei esker Nobel saria [2] irabazi duten ikerlariak.
| Ozpin-euli | |
|---|---|
.jpg.webp) | |
| Sailkapen zientifikoa | |
| Erreinua | Animalia |
| Filuma | Arthropoda |
| Klasea | Insecta |
| Ordena | Diptera |
| Familia | Drosophilidae |
| Generoa | Drosophila |
| Espeziea | Drosophila melanogaster (Meigen, 1830)
|
| Eguneko zikloa | eguneko |
| Genomaren kokapena | ensembl.org… eta metazoa.ensembl.org… |
Drosophila oso erabilia da ikerkuntzan, espezieak dituen ezaugarriak direla eta. Hasteko, euliak laborategian hazteko errazak dira eta haien ugalketa-zikloa oso azkarra izaten da. Bigarrenik, espezie honetako euliek lau kromosoma pare dituzte bakarrik [3]. Hirugarrenik, bizi-ziklo laburra dute, 15-21 egunekoa hain zuzen ere. Laugarrenik, zenbait gizakiren gaixotasunak eragiten dituzten geneen % 61 bat datoz ozpin-eulien genomarekin, eta eulien proteina-sekuentzien % 50ek analogoak dituzte ugaztunetan [4]. Azkenik, haien habitata oso zabala da; izan ere, kontinente guztietan aurki daitezke, baita uharteetan ere [5].
Tephritidae familiako euliei ere esaten zaie fruta-euli. Hori dela eta, denominazioa nahasgarria izan daiteke, batez ere, Mediterraneo aldean, Australian eta Hego Afrikan, non Mediterraneoko fruta-eulia, Ceratitis capitata, plaga ekonomikoa den.
Bizi-zikloa eta ugalketa
D.melanogasterren bizi-zikloak 21 egun irauten du, arrautzatik irteten denetik hiltzen den arte.[6]
D.melanogasterren garapen prozesua tenperaturaren menpekoa da. Garapen prozesuak arrautzatik hasi eta heldu bihurtu arte irauten du. Garapen-denbora laburrena 7 egunekoa izaten da, eta 28 °C-tan lortzen da. Garapen prozesua luzatu egiten da tenperatura igo ahala: 19 egunekoa da 18 °C-tan, eta 50 egun baino gehiagokoa tenperatura 12 °C-koa denean. Tenperatura optimoa 25 °C-koa da, euliak 8,5 egunetan garatzea eragiten duena. Emeek gutxi gorabehera 0,5 mm-ko 400 arrautza inguru erruten dituzte usteldutako frutan edo perretxikoetan. Baldintza optimoetan (25 °C) 12-15 ordu iragandakoan eklosionatzen dute arrautzek, eta horietatik irteten diren larbek, 4 egun pasatu ondoren, pupa bihurtu eta metamorfosia jasaten dute. Metamorfosiaren ostean, heldu bilakatzen dira. Larba direlarik, bitan aldatzen dute azala (exubiazioa), arrautzatik irten eta 24 eta 48 ordura.[7] Garapen-fase horretan, fruta usteltzen duten mikroorganismoez eta frutak berak dauzkan azukreez elikatzen dira.[8]
Ugalketa
Ozpin-euli arrek emeak gorteatu behar dituzte, ugaldu baino lehen. Gorteatze prozesu horrek bost fase ditu, zeinetan arrak emea erakartzen saiatzen den hegoen bibrazioen eta bestelako taktiken bitartez, emeak onartzen duen arte. Emeek arrak errefusa ditzakete horiei ostikoak emanez edo alde eginez.[9] Euli emeak arra onartzen badu, kopulazioak 15-20 minutu irauten du.[10] Emeek bikote batekin baino gehiagorekin kopulatu ohi dute. Beraz, eme bakarrak errunaldi bakarrean ar baten baino gehiagoren arrautzak ipintzen ditu. Hori gertatzen da D.melanogaster emeek kopulatzen dituzten arren esperma gordetzen dutelako euren gorputzaren barruan. Ar desberdinen espermek lehiatu egiten dute emearen obuluak ernaltzeko.[11] Zientzialariek uste dute kopulatzen azkena izan den arrak arrautzen % 80ren aita izango dela lehia horren ondorioz. Ernalketa gertatu ostean, emearen umetokiko neurona sentsorialek arren esperman dagoen proteina bati, sexu-peptidoari, erantzuten diote. Proteina horrek emeak ernalketa gertatu osteko 10 egunetan kopulatzea ekiditen du. Neurona sentsorialak sexu-peptidoaren aurrean kitzikatzen dira eta eulietan hipotalamoaren homologoa den garunaren gunera bidaltzen dute seinalea. Ondoren, hipotalamoak desira eta jokaera sexuala kontrolatzen ditu. Prozesu horren bide metabolikoa ondo deskribatuta dago.[12]
D. melanogaster bizitzaren luzera aztertzen duten ikerketetan erabiltzen da, mutatutakoan eulien bizitza luzatzen duten geneak identifikatzeko adibidez.[13]
Emeak
Emeak azaleratu eta 8-12 ordu igaro ostean arrentzako harkor bihurtzen dira eta, gainera, arren hautapenean eta horiekiko portaeran eragiten duten neurona talde espezifikoak aurkitu dira.[14] Emeek badituzte nerbio abdominaleko neurona jakin batzuk zeinek gorputzeko mugimenduak pausaraz ditzaketen kopulatzeko.[15] Neurona horien aktibazioak emeak bere mugimenduak gelditu eta arraren norabidean orientatzea eragiten du, estalketa baimentzeko helburuarekin. Neurona talde espezifiko hori aktibo ez badago, aldiz, emeak bere mugimenduarekin jarraitzen du eta ez da kopulaziorik gertatzen. Arren feromonak bezalako seinale kimikoek neuronen talde horien aktibazioa susta dezakete.[15] Horrez gain, emeek arren hautapena kopiatzen dute; hau da, birjinak direnean, beste emeak ar mota jakinekin kopulatzen ikusten badituzte, mota horretako arrekin kopulatzeko joera izaten dute. Portaera hori ingurumen-aldaketekiko sentikorra da: emeek gutxiago kopulatzen dute eguraldia txarra denean.[16]
Arrak
D. melanogaster espezieko arrek haien esperientzia sexuala erabiltzen dute etorkizuneko portaera sexuala aldatzeko. Aldaketa horiek, besteak beste, harkortasun-denbora motzagoak eta espezifikoagoak izatea eragiten dutela dirudi. D. simulans espezieko euliekin gertatzen den moduan, D. melanogaster espezieko ar "inozoek" nahiko denbora behar dute harkorrak izateko, eta hasieran, oraindik sexualki aktiboak ez diren emeak gorteatzen saiatzen dira, baita beste ar batzuk ere. D. melanogaster espezieko arrek ez dute lehentasunik erakusten D. melanogaster espezieko emeekiko, beste espezie batzuetako emeekiko edo beste euli arrekiko. Hala ere, D. simulans edo beste euli-espezie batzuekiko gorteatzea eraginkorra ez dela ikusita, D. melanogaster espezieko arrek euren portaera moldatzen ikasi dute, ugalketa-helbururik ez duten prozesuetan gehiegizko energia ez galtzeko.[17]
Horrez gain, esperientzia sexual zabala duten euliek azkarrago ugaltzeko gaitasuna dute; izan ere, denbora gutxiago ematen dute emea gorteatzen eta, beraz, estaltze-sortasun baxuagoak izango dituzte. Horren ondorioz, esperientzia duten arrek etekin handiagoa lortzen dute estalketan ar sineskorrek baino.[18] Horrek abantaila ebolutiboak eskaintzen dizkie, oso garrantzitsuak direnak hautespen naturalaren aldetik ikusita.
Itxura fisikoa eta sexuaren determinazioa
Itxura fisikoa

Fruta-euli basatiak hori-marroixkak dira, begi gorriak dituzte eta zeharkako eraztun beltza ere sabelean. Dimorfismo sexuala erakusten dute: emeek 2,5 mm-ko luzera dute eta, arrak, aldiz, apur bat txikiagoak dira, eta atzeko alde beltzagoa dute. Arrak erraz bereizten dira emeetatik begi-bistaz, koloreen ezberdintasuna dela eta. Alde batetik, sabelean gune beltz bereizgarria dute, jaioberrietan gutxiago nabaritzen dena. Bestetik, arrek sexu-orrazia dute, hau da, zurda beltzen ilara lehenengo hankaren tartsoan. Gainera, emeari lotzeko duten gunea ile zorrotzez inguratuta dago. Argazki ugari aurki daitezke FlyBase orrialdean.[19]
Sexuaren determinazioa
Drosophila euliek X eta Y kromosomak eta autosomak dituzte. Gizakietan ez bezala, Y kromosomak ez du ar-ezaugarririk eragiten, baina hazia sortzeko ahalmena duten geneak kodetzen ditu. Sexua, ordea, X kromosomen eta autosomen arteko proportzioak determinatzen du. Bestetik, zelula bakoitzak arra edo emea izatea "erabaki" dezake, organismoaren beste zelulak "kontuan hartu" gabe. Jokaera horrek ohikoa den ginandromorfismoa eragiten du, alegia, bi sexuen ezaugarri fisikoak adieraztea.
Hiru gene nagusik parte hartzen dute Drosophilaren sexu-determinazioan. Horiek dira: sex-lethal, sisterless, eta deadpan. Gene horien kantitateak eragin handia du euli baten sexuan, hiru geneek elkar inhibitzen baitute. Hiru geneetatik zeintzuk adierazten diren eta zein kantitatetan adierazten diren, horren arabera erabakiko da euliaren amaierako sexua.
| X Kromosomak | Autosomak | X:A proportzioa | Sexua |
|---|---|---|---|
| XXXX | AAAA | 1 | Eme arrunta |
| XXX | AAA | 1 | Eme arrunta |
| XXY | AA | 1 | Eme arrunta |
| XXYY | AA | 1 | Eme arrunta |
| XX | AA | 1 | Eme arrunta |
| XY | AA | 0.50 | Ar arrunta |
| X | AA | 0.50 | Ar arrunta (antzua) |
| XXX | AA | 1.50 | Metaemea |
| XXXX | AAA | 1.33 | Metaemea |
| XX | AAA | 0.66 | Intersexua |
| X | AAA | 0.33 | Metaemea |
Erabilpen historikoa analisi genetikorako eredu gisa

D. melanogaster espezieko euliak analisi genetikorako hautatu ziren lehen organismoak dira eta, beraz, organismo eukariotoen artean, genetikoki hobekien ezagutzen den eta gehien erabiltzen den eredua. Organismo guztiek antzeko sistema genetikoak erabiltzen dituzte eta, ondorioz, transkripzioa eta erreplikazioa bezalako prozesuak ozpin-eulietan ulertzeak ahalbidetzen du gainerako eukariotoetan ere antzeko prozesuak ulertzea, gizakiarenak ere bai.[20] Fruitu-eulia lehenengo aldiz erabili zen herentzia aztertzeko ikerkuntzetan 1910.urtean Columbia Unibertsitatean, Thomas Hunt Morganen esku.[20]
Esan bezala, D.melanogaster espezieko euliak oso erabiliak izan dira ikerkuntza biologikoan, zenbait arrazoi direla eta:
- Eulien hazkuntzarako behar den ekipamendua ez da garestia, ezta horretarako beharrezkoa den espazioa ere.
- Era errazean anestesiatu daitezke (karbono dioxidoa, eterra, hoztuz edota FlyNap bezalako produktuen bidez).
- Behin anestesiatuta, horien morfologia bereizteko erraza da.
- Ugalketa-denbora motza dute (giro-tenperaturan 10 egun ingurukoa) eta horrek aste gutxi batzuetan hainbat belaunaldi ikertzeko aukera ematen du.
- Emankortasun altua dute (emeek 100 arrautza baino gehiago jar ditzakete egunean, eta horien bizitza osoan zehar 2000 arrautzetara irits daitezke). [21]
- Emeen eta arren arteko bereizketa azkar egin daiteke eta eme birjinak ere azkar isola. Prozesu guztiak gurutzaketa genetikoa errazten dute.
- Larba helduek kromosoma erraldoiak erakusten dituzte listu-guruinetan (kromosoma politenikoak), zeinak aktibitate genetikoaren erakusle diren.
- Lau kromosoma pare ditu soilik, hiru autosoma pare eta bi sexu-kromosoma pare.
- Arrek ez dute birkonbinazio meiotikorik, eta horrek ikerketa-genetikoak errazten ditu.
- Organismoaren garapena (arrautza ernaltzen denetik euli heldura arte) ondo ezagutzen da.
- Transformazio genetikorako teknikak 1987.urtetik aurrera eskuragai daude.
- Haren genoma osoa 2000.urtean sekuentziatu eta argitaratu zen. [22]
- Dibertsitate sexuala erraz azter daiteke, eulien garapenari eta jokabideari buruzko informazio erabilgarria lortzeko. [23]
Markatzaile genetikoak
Markatzaile genetikoak oso erabiliak dira Drosophila melanogaster espezieko eulietan eta, horrela, fenotipo gehienak erraz identifikatu daitezke mikroskopio baten laguntzarekin. Jarraian, markatzaile genetiko batzuk zerrendatu dira: aleloaren ikurra eta erasandako genearen izena zehaztu dira, baita fenotipo desberdinen azalpenak ere. Alelo errezesiboak letra xehez ageri dira eta alelo dominanteak, aldiz, letra larriz.
- Cy1: curly; eulien hegoak gorputzarekiko okertuta daude, eta, ondorioz, batzuetan hegaldia oztopa dezakete.
- e1: ebony; gorputza eta hegoak beltzak dira (heterozigotoak basatiak baino ilunago ikusten dira).
- Sb1: stubble; ileak basatiena baino motzagoak eta lodiagoak dira.
- w1: white; begiek ez dute pigmentaziorik eta, beraz, zuri ikusten dira.
- y1: yellow; gorputzaren eta hegoen pigmentazioak horia dirudi, albinismoaren euli analogoa izango litzateke.
Genoma

D. melanogaster euli-espezieak lau kromosoma pare ditu: X/Y parea eta hiru autosoma pare, hain zuzen ere. Laugarren kromosoma oso txikia da, eta askotan ez da kontuan hartzen. Hala ere, begien gabezia eragiten duen geneak kromosoma horretan kokaturik dago. D. melanogaster espeziearen genoma 139.5 milioi [24] base parek osatzen dute, eta 15682 gene ditu. Genomaren % 60 baino gehiago, geneen adierazpena kontrolatzen duen proteina kodetzen ez duen DNA [25] funtzionala dela dirudi. Sexuaren determinazioa D. melanogaser eulian X kromosomen eta autosomen arteko proportzioaren araberakoa da, gizakietan ez bezala. Nahiz eta Y kromosoma guztiz heterokromatikoa izan, gutxienez 16 gene ditu eta horietako askok arrekin erlazionaturiko funtzioak dituzte [26].
Gizakiekiko antzekotasuna
2000. urtean fruta-euliaren genomaren eta giza genomaren alderaketa gauzatu zen National Human Genome Research Istitutek egindako ikerketa batean. Egindako estimazioaren arabera, geneen % 60 bat datoz giza genomarekin [27]. Zenbait gizakiren gaixotasunak eragiten dituzten geneen % 75 ozpin-eulien genoman ageri dira, eta eulien proteina-sekuentzien % 50ek analogoak dituzte ugaztunetan . D. melanogaster eredu genetiko modura erabiltzen da zenbait gaixotasun aztertzeko, besteak beste, Parkinson gaixotasuna eta Alzheimer gaixotasuna [28]. Euli-espezie hau estres oxidatiboan, zahartze prozesuan, diabetes gaixotasunean eta minbizian eragiten dituzten mekanismoak aztertzeko erabiltzen da [29][30][31].
Garapena
D.melanogasterren garapen prozesuak lau fase ditu: arrautza, larba, pupa eta euli heldua. [32]
D.melanogaster ikerketa genetikoetan oso erabilia izan den organismo modeloa da. Haren garapen prozesua sakon ikertu da, abantaila asko eskaintzen baititu beste animalia eredu batzuekin alderatuta: tamaina txikia, garapen-denbora laburra eta arrautza kopuru handia errunaldi bakoitzean.
Oogenesia
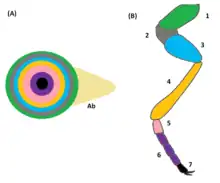
Oozitoa ernaldu ostean, sortu berri den enbrioiak hainbat DNA-bikoizketa jasaten ditu, eta 13 nukleo-zatiketa. Prozesu horren ondoren, 5000 eta 6000 nukleo artean egoten dira zelula bakarrean, zitoplasma partekatzen. Nukleo horiek zitoplasmaren gune desberdinetara migratzen dute, gehienek enbrioiaren azalera, eta beste batzuek enbrioiaren erdialdera. Azken horiek bitelo-zakua osatzen dute. Nukleo-zatiketak oso azkar gertatzen dira, kontrol handirik gabe, eta, beraz, akatsak egon daitezke DNAn. Akasdun nukleoek enbrioiaren erdialdera migratzen dute, bitelo-zakuko zelula bihurtzeko, gune hori ez baita euliaren gorputzaren parte izango. Hasierako garapen-faseetan aldaketa asko gertatzen dira enbrioian, eta ezagunak diren gene askok parte hartzen dute horietan. Gene-sare hau hobekien deskribatuta dagoen gene-sareetako bat da.
Gastrulazioan eta hasierako garapen-faseetan zehar enbrioiak mugimendu morfogenetiko ugari jasaten ditu: mesodermoaren eta endodermoaren hainbat inbaginazio, gorputz-atalen segmentazioa, etab. Arrautzatik irten ostean, larba fasean sartzen da. Fase horretan disko imaginalak sortzen hasten dira larban. Disko horiek zelula epitelialen multzoak dira, pupa fasean euliaren kanpoko egitura bilakatuko direnak. Zelula multzoak disko moduan antolatzen dira, hortik datorkie izena.
Larba fasea amaitzean, metamorfosia gertatzen da. Prozesu horretan zehar, larba pupa bihurtzen da. Larba osatzen zuten ehunak birxurgatu egiten dira, eta disko imaginalek mugimendu morfogenetiko asko jasaten dituzte, euli helduen egiturak sortu arte, adibidez: burua, hegoak, toraxa eta sexu-organoak.[33]
Erreferentziak
- (Ingelesez) «Vinegar Flies (Department of Entomology)» Department of Entomology (Penn State University) (Noiz kontsultatua: 2018-10-19).
- (Ingelesez) «Nobel Prizes | ari.info | ari.info» ari.info (Noiz kontsultatua: 2018-10-19).
- (Ingelesez) Reeve, Eric C. R.; Black, Isobel. (2001). Encyclopedia of Genetics. Taylor & Francis ISBN 9781884964343. (Noiz kontsultatua: 2018-10-19).
- Reiter, L. T.. (2001-06-01). «A Systematic Analysis of Human Disease-Associated Gene Sequences In Drosophila melanogaster» Genome Research 11 (6): 1114–1125. doi:. ISSN 1088-9051. (Noiz kontsultatua: 2018-10-19).
- Markow, Therese Ann. «The secret lives of Drosophila flies» eLife 4 doi:. ISSN 2050-084X. PMID 26041333. PMC PMC4454838. (Noiz kontsultatua: 2018-10-19).
- McGurk, Leeanne; Berson, Amit; Bonini, Nancy M.. (2015-10). «Drosophila as an In Vivo Model for Human Neurodegenerative Disease» Genetics 201 (2): 377–402. doi:. ISSN 1943-2631. PMID 26447127. PMC PMC4596656. (Noiz kontsultatua: 2018-11-16).
- M.,, Ashburner,. (2005). Drosophila : a laboratory handbook. (2nd ed. argitaraldia) Cold Spring Harbor Laboratory Press ISBN 9780879697068. PMC 56324270. (Noiz kontsultatua: 2018-11-16).
- Blum, Jessamina E.; Fischer, Caleb N.; Miles, Jessica; Handelsman, Jo. (2013-11-05). «Frequent replenishment sustains the beneficial microbiome of Drosophila melanogaster» mBio 4 (6): e00860–00813. doi:. ISSN 2150-7511. PMID 24194543. PMC PMC3892787. (Noiz kontsultatua: 2018-11-16).
- (Ingelesez) Connolly, Kevin; Cook, Robert. (1973-01-01). «Rejection Responses By Female Drosophila Melanogaster : Their Ontogeny, Causality and Effects Upon the Behaviour of the Courting Male» Behaviour 44 (1): 142–165. doi:. ISSN 1568-539X. (Noiz kontsultatua: 2018-11-16).
- Houot, Benjamin; Svetec, Nicolas; Godoy-Herrera, Raùl; Ferveur, Jean-François. (2010-07-01). «Effect of laboratory acclimation on the variation of reproduction-related characters in Drosophila melanogaster» The Journal of Experimental Biology 213 (Pt 13): 2322–2331. doi:. ISSN 1477-9145. PMID 20543131. (Noiz kontsultatua: 2018-11-16).
- Price, C. S.; Dyer, K. A.; Coyne, J. A.. (1999-07-29). «Sperm competition between Drosophila males involves both displacement and incapacitation» Nature 400 (6743): 449–452. doi:. ISSN 0028-0836. PMID 10440373. (Noiz kontsultatua: 2018-11-16).
- Ekanayake, Darshika. [leave for Adam - compressed fileThe mating system and courtship behaviour of the Queensland fruit fly, Bactrocera tryoni (Froggatt) (Diptera: Tephritidae). ] Queensland University of Technology (Noiz kontsultatua: 2018-11-16).
- Carnes, Megan Ulmer; Campbell, Terry; Huang, Wen; Butler, Daniel G.; Carbone, Mary Anna; Duncan, Laura H.; Harbajan, Sasha V.; King, Edward M. et al.. (2015). «The Genomic Basis of Postponed Senescence in Drosophila melanogaster» PloS One 10 (9): e0138569. doi:. ISSN 1932-6203. PMID 26378456. PMC PMC4574564. (Noiz kontsultatua: 2018-11-16).
- (Ingelesez) Pitnick, Scott. (1996-07). «Investment in Testes and the Cost of Making Long Sperm in Drosophila» The American Naturalist 148 (1): 57–80. doi:. ISSN 0003-0147. (Noiz kontsultatua: 2018-10-19).
- (Ingelesez) «Fruit fly research may reveal what happens in female brains during courtship, mating» ScienceDaily (Noiz kontsultatua: 2018-10-19).
- (Ingelesez) Anne-Cecile, Dagaeff,; Arnaud, Pocheville,; Sabine, Nöbel,; Adeline, Loyau,; Guillaume, Isabel,; Etienne, Danchin,. «Drosophila mate copying correlates with atmospheric pressure in a speed learning situation» Animal Behaviour 121 ISSN 0003-3472. (Noiz kontsultatua: 2018-10-19).
- (Ingelesez) Dukas, R.. (2004-07-01). «Male fruit flies learn to avoid interspecific courtship» Behavioral Ecology 15 (4): 695–698. doi:. ISSN 1465-7279. (Noiz kontsultatua: 2018-10-19).
- Saleem, Sehresh; Ruggles, Patrick H.; Abbott, Wiley K.; Carney, Ginger E.. (2014-05-07). «Sexual Experience Enhances Drosophila melanogaster Male Mating Behavior and Success» PLoS ONE 9 (5) doi:. ISSN 1932-6203. PMID 24805129. PMC PMC4013029. (Noiz kontsultatua: 2018-10-19).
- FlyBase Homepage. 2009-08-15 (Noiz kontsultatua: 2018-10-19).
- A., Pierce, Benjamin. (2005). Genetics : a conceptual approach. (2nd ed. argitaraldia) W.H. Freeman ISBN 0716788810. PMC 57061864. (Noiz kontsultatua: 2018-10-26).
- Encyclopedia of genetics. Fitzroy Dearborn 2001 ISBN 1884964346. PMC 46984073. (Noiz kontsultatua: 2018-10-26).
- Adams, M. D.; Celniker, S. E.; Holt, R. A.; Evans, C. A.; Gocayne, J. D.; Amanatides, P. G.; Scherer, S. E.; Li, P. W. et al.. (2000-03-24). «The genome sequence of Drosophila melanogaster» Science (New York, N.Y.) 287 (5461): 2185–2195. ISSN 0036-8075. PMID 10731132. (Noiz kontsultatua: 2018-10-26).
- Hotta, Y.; Benzer, S.. (1972-12-29). «Mapping of behaviour in Drosophila mosaics» Nature 240 (5383): 527–535. ISSN 0028-0836. PMID 4568399. (Noiz kontsultatua: 2018-10-26).
- (Ingelesez) «Drosophila melanogaster (ID 47) - Genome - NCBI» www.ncbi.nlm.nih.gov (Noiz kontsultatua: 2018-10-26).
- Halligan, Daniel L.; Keightley, Peter D.. (2006-7). «Ubiquitous selective constraints in the Drosophila genome revealed by a genome-wide interspecies comparison» Genome Research 16 (7): 875–884. doi:. ISSN 1088-9051. PMID 16751341. PMC PMC1484454. (Noiz kontsultatua: 2018-10-26).
- (Ingelesez) «Origin and evolution of the Drosophila Y chromosome» Current Opinion in Genetics & Development 12 (6): 664–668. 2002-12-01 doi:. ISSN 0959-437X. (Noiz kontsultatua: 2018-10-26).
- (Ingelesez) «Background on Comparative Genomic Analysis» National Human Genome Research Institute (NHGRI) (Noiz kontsultatua: 2018-10-26).
- Jaiswal, M.; Sandoval, H.; Zhang, K.; Bayat, V.; Bellen, H.J.. (2012). «Probing Mechanisms That Underlie Human Neurodegenerative Diseases in Drosophila» Annual review of genetics 46: 371–396. doi:. ISSN 0066-4197. PMID 22974305. PMC PMC3663445. (Noiz kontsultatua: 2018-10-26).
- (Ingelesez) Fly Models of Human Diseases. Academic Press 2017-01-02 ISBN 9780128029053. (Noiz kontsultatua: 2018-10-26).
- (Ingelesez) Buchon, Nicolas; Silverman, Neal; Cherry, Sara. (2014-12). «Immunity in Drosophila melanogaster — from microbial recognition to whole-organism physiology» Nature Reviews Immunology 14 (12): 796–810. doi:. ISSN 1474-1733. (Noiz kontsultatua: 2018-10-26).
- Kaun, Karla R.; Devineni, Anita V.; Heberlein, Ulrike. (2012-6). «Drosophila melanogaster as a model to study drug addiction» Human Genetics 131 (6): 959–975. doi:. ISSN 0340-6717. PMID 22350798. PMC PMC3351628. (Noiz kontsultatua: 2018-10-26).
- Markow, Therese Ann. (2015-06-04). «The secret lives of Drosophila flies» eLife 4 doi:. ISSN 2050-084X. PMID 26041333. PMC PMC4454838. (Noiz kontsultatua: 2018-11-16).
- Weigmann, Katrin; Klapper, Robert; Strasser, Thomas; Rickert, Christof; Technau, Gerd; Jäckle, Herbert; Janning, Wilfried; Klämbt, Christian. (2003-6). «FlyMove--a new way to look at development of Drosophila» Trends in genetics: TIG 19 (6): 310–311. doi:. ISSN 0168-9525. PMID 12801722. (Noiz kontsultatua: 2018-11-16).
Kanpo estekak
| Gai honi buruzko informazio gehiago lor dezakezu Scholian |
| Autoritate kontrola |
|
|---|
 Datuak: Q130888
Datuak: Q130888 Multimedia: Drosophila melanogaster / Q130888
Multimedia: Drosophila melanogaster / Q130888 Espezieak: Drosophila melanogaster
Espezieak: Drosophila melanogaster