Azido desoxirribonukleiko
Azido desoxirribonukleikoa (oro har DNA siglarekin laburtua; edota, batzuetan, ADN siglarekin)[oh 1] azido nukleiko mota bat da, organismo bizi[1] guztien funtzionamendu eta garapenaren informazio genetikoa duena eta bere transmisioaren erantzulea dena.
| Genetika |
|---|
 |

Ikuspuntu kimikotik, DNA nukleotidoen polimero bat da; hau da, polinukleotido bat.[2] Elkarrekin lotutako unitate sinple askoz osatutako polimero konposatua da, bagoi askoz osatutako tren baten antzekoa. DNAn bagoi bakoitza nukleotido bat da, eta, aldi berean, nukleotido bakoitza azukre (desoxirribosa), base nitrogenatu (adenina (A), timina (T), zitosina (C) alaguanina (G)) eta fosfato talde batez osatuta dago; azken horrek bagoiak elkarren artean lotzen ditu. Bagoi edo nukleotido bat bestetik bereizten duena base nitrogenatua da; horregatik, DNA sekuentzia base nitrogenatuak aipatuz bakarrik izendatzen da. Lau base horiek sekuentzian duten ordena informazio genetikoa kodifikatzen duena da, esaterako, ATCGATCG... Organismo bizietan, DNA nukleotido kate bikoitz moduan agertzen da, non bi kateak hidrogeno zubien bidez lotuta agertzen diren.[3]
Zelulak, DNAk duen informazioa erabili ahal izateko, nukleotidoak RNA (azido erribonukleikoa) molekulak emanez kopiatzen dira. RNA molekulak DNAtik kopiatzen dira transkripzio izeneko prozesu baten bidez. RNA molekula hauek, nukleoan prozesatu ondoren, zitoplasmara irteten dira erabiliak izateko. RNAk duen informazioa kode genetikoa erabiliz itzultzen da proteinen aminoazido sekuentziak osatuz. Nukleotido hirukote (kodon) bakoitzak aminoazido bat emango du, eta, ondoren, aminoazido hauek elkartuko egingo dira lotura peptidikoen bidez proteinak osatuz. RNA erabiliz proteinak sintetizatzeko prozesu honi, itzulpena esaten zaio.
Herentziaz arduratzen diren eta DNAren sekuentzian ezinbestekoak diren unitateak geneak dira. Gene bakoitzak RNAra transkribatuko den zati bat eta noiz eta non espresatuko den definitzeaz arduratuko den beste zati bat ditu. Geneetan dagoen informazioa zelularentzat ezinbestekoak diren RNA eta proteinak sintetizatzeko erabiltzen da.
Zelularen barnean, DNA kromosoma izeneko egitura batzuetan antolatuta dago. Hauek, ziklo zelularrean zelula banandu aurretik, bikoiztu egiten dira. Organismo eukariotoek (animalia, landare eta onddoek) bere DNAren gehiengoa zelularen nukleoan gordetzen dute, eta, gainerakoa, mitokondria eta kloroplastoetan. Prokariotoek (bakterioak eta arkeoak), zelularen zitoplasman gordetzen dute, eta birusek, aldiz, proteinazko kapsidaren barnean. Kromosoma hornidura guztiaren materia genetikoa, genoma izenez ezagutzen da, eta espezie bakoitzaren ezaugarria da.
Historia

DNA 1869ko neguan isolatu zuen lehenengo aldiz Friedrich Miescher suitzar medikuak Tubingengo Unibertsitatean lan egiten zuenean. Erabilitako benda kirurgikoetako zornearen konposaketa kimikoari buruzko esperimentuak egiten ari zela, hauspeatuta zegoen substantzia ezezagun bat ikusi zuen.[4][5] "Nukleina" izena eman zion, zelulen nukleotik erauzi baitzuen.[6] Ia 70 urtetako ikerketa behar izan zen azido nukleikoen egitura eta osagaiak identifikatu ahal izateko.
1919an, Phoebus Levenek argitu zuen nukleotidoak, izan ere, base, azukre eta fosfatoz osatuta zeudela.[7] Levenek proposatu zuen DNAk solenoide (malguki) itxurako egitura eratzen zuela eta nukleotido unitateak fosfato taldeen bidez elkartuta zeudela. 1930ean, Levenek eta bere maisu Albrecht Kosselek frogatu zuten nukleina lau base nitrogenatu (zitosina (C), timina (T), adenina(A) eta guanina(G)), desoxirribosa azukre eta fosfato talde batez osatutako azido desoxirribonukleikoa zela eta bere oinarrizko egitura base eta fosfato bati lotutako azukrea[8] zela. Aitzitik, Levenek katea motza zela eta baseak ordena jakin bati jarraituz errepikatzen zirela uste zuen. 1937an, William Astburyk DNAk egitura erregularra zuela erakusten zuen X izpien lehen difrakzio patroia asmatu zuen.[9]

DNAren funtzio biologikoa 1928an hasi zen argitzen Frederick Griffith Pneumococcus, bakterioaren andui batzuekin lanean ari zela, genetika modernoaren esperimentu batzuk burutu zituenean. Neumokokoaren andui batzuk "leunak" (S) ziren, eta beste batzuk "zimurrak" (R); horren arabera, bakterioaren birulentzia aldakorra zen.
Arratoiei S neumokoko biziak injektatuz, haien heriotza gertatzen zen, baina R neumokoko biziak edo beroaren eraginez hildako S neumokokoekin ez zirela hiltzen ikusi zuen Griffithek. Hala ere, R neumokoko biziak eta S neumokoko hilak, aldi berean, injektatuz gero, arratoiak hil egiten ziren, eta, beren odolean, S neumokoko biziak isolatzen ziren. Hildako bakterioak arratoiaren barnean bikoiztea ezinezkoa denez, Griffithek "printzipio transformatzaile" deitu zuen arrazoinamendu bat egin zuen. Teoria horrek, substantzia aktibo baten transferentziaren bidez, bakterio mota batetik besterako aldaketa gertatzen zela azaltzen zuen. Substantzia honek R neumokokoei kapsula azukretsu bat sortu, eta, horrela, birulentoak bihurtzea ahalbideratuko lieke.
Hurrengo 15 urteetan, bakterioaren andui desberdinen konbinaketak erabiliz, behin eta berriz errepikatu ziren esperimentu hauek, bai arratoietan (in vivo) eta bai saio hodietan (in vitro) ere.[10] Hasiera batean, birulentoak ez ziren anduiak birulento bihurtzeko gaitasuna zuen printzipio transformatzailearen bilaketak 1944ra arte jarraitu zuen. Urte horretan, Oswald Avery, Colin MacLeod eta Maclyn McCartyk gaur egun klasikoa den esperimentu bat eraman zuten aurrera. Ikertzaile hauek frakzio aktiboa (printzipio transformatzailea) erauztea lortu zuten, eta, analisi kimiko, entzimatiko eta serologikoei esker, ez zuela proteina, lipido edo polisakarido aktiborik ikusi zuten. Azido desoxirribonukleikoaren forma likatsu eta oso polimerizatu batez osatuta zegoela ikusi zuten, hau da, DNA[11].
Beroaren bidez hildako S neumokokoetatik erauzitako DNA, hala, R neumokoko biziekin nahastu zuten "in vitro". Horren ondorioz, S kolonia bakterianoak eratu ziren, eta, beraz, zalantzarik gabe printzipio transformatzailea DNA zela frogatu zen.
DNA, printzipio transformatzaile gisa, unibertsalki onartua izateko hainbat urte pasa ziren arren, aurkikuntza hau genetika molekurraren jaiotzan giltzarria izan zen. Azkenik, DNAk herentzian zuen papera 1952an onartu zen Alfred Hershey eta Martha Chasen esperimentuei esker.
Molekularen karakterizazio kimikoari dagokionez, 1940an, Chargaffek DNAren base nitrogenatuen proportzioak ezagutzeko balio izan zuten esperimentu batzuk egin zituen. Purinen proportzioak pirimidinen berdinak zirela ikusi zuen ([A]=[T] eta [G]=[C]), eta, DNA molekula jakin batean, G+C kantitatea ez zela beti A+T kantitatearen berdina, % 36–70 artean aldakorra zela. Informazio honekin eta Rosalind Franklinek lortutako X izpien difrakzio datuekin, James Watson eta Francis Crickek DNAren egitura tridimentsionala azaltzen zuen helize bikoitzaren modeloa proposatu zuten 1953an.
Nature aldizkari zientifikoaren ale berean, Watsonek eta Crickek proposatzen zuten modeloaren alde agertzen ziren bost artikulu argitaratu ziren. Artikulu hauetan lehena, Franklin eta Raymond Goslingena, X izpien difrakzio datuekin modeloaren alde egiten zuena zen.
1962an, Franklin hil eta gero, Watsonek, Crickek eta Wilkinsek Medikuntzaren Nobel Saria jaso zuten. Gaur egun ere, zientziaren arloan, bizirik dirau ea aurkikuntzaren merezimendua norena den eztabaidak.
DNAren egitura

DNA antiparaleloak diren bi desoxirribonukleotido-kateez osatuta dago. Kate hauen osagaiak, lehen aipatu bezala, azukre talde bat (desoxirribosa), fosfato talde bat eta base nitrogenatu bat (adenina (A), guanina (G), zitosina (C) edo timina (T)) dira. Bere egitura molekularrak bi kateek eratutako helize bikoitz baten forma hartzen du, base nitrogenatuak aurrez aurre kokaturik dituela, non A baten aurrean T bat kokatzen den beti eta G baten aurrean C bat. Base puriko bat (tamaina handiagokoa) base pirimidiniko baten (tamaina txikiagokoa) aurrean geratzen denez beti, katea bikoitzaren zabalera konstantea mantentzen da.
Fenomeno horri base-parekatzea deritzo, eta, hari esker, bi kateak osagarriak dira; hau da, kate baten baseen sekuentzia ezagutzen bada, bestearena ondoriozta daiteke. Osagarritasun honi esker, DNAk gordetzen duen informazioa bi kateetan agertuko da; hau da, bikoiztuta dago. Hau funtsezkoa izango da DNAren erreplikazio prozesuan.
DNA molekula egonkortzeko, parekatutako baseen artean hidrogeno zubiak eratzen dira. Lotura hau eratzeko, base batek karga partzial positiboa duen talde baten (−NH2 ala −NH) hidrogeno atomo bat eman behar dio beste basean elektronegatiboki kargatuta dagoen talde bati (C=O ala N). Hidrogeno zubiak lotura kobalenteak baino ahulagoak dira; apurtu, eta berriro sor daitezke zailtasun handirik gabe. Hori dela eta, helize bikoitzaren bi kateak kremailera baten moduan banandu daitezke, indar mekaniko edo tenperaturaren eraginez (desnaturalizazio izeneko prozesua)

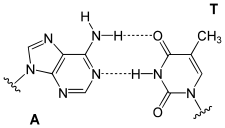
Base nitrogenatu guztiek ez dituzte hidrogeno zubi kopuru berak ematen, A=T bi hidrogeno zubi eratzen dituzte, eta, aldiz, C≡G, hiru. Beraz, C≡G base parea A=T baino sendoagoa da. Horren ondorioz, DNA helizearen sendotasuna bertan aurkitzen diren C≡G base pareen portzentaiaren eta molekularen luzeraren araberakoa izango da. C≡G base pare asko dituzten helize luzeen kateak A=T base pare asko dituzten helize laburren kateak baino indar handiagorekin daude lotuta.
Laborategian, sendotasun hori neurtzeko, hidrogeno zubiak apurtzeko behar den tenperatura neurtzen da; hau da, urtze tenperatura. Molekularen base pareen lotura guztiak puskatzean, bi kateak bereizi egiten dira.
DNA egituraren mailak
DNAren egitura tridimentsionalean, hainbat maila bereizten dira:
Lehen mailako egitura
Kateatutako nukleotido sekuentzia. Informazio genetikoa dagoen kate hauetan, desberdintasuna base nitrogenatuetan dago.
Bigarren mailako egitura
- Helize bikoitz itxurako egitura da. Informazio genetikoaren metaketa eta DNAren erreplikazio mekanismoa azaltzeko balio du. Watson eta Crickek aurkitu zuten Franklinen eta Wilkinsen X izpien difrakzioan oinarrituz.
- Kate bikoitza da destrogiroa (eskuinerantz biratzen dena) edo lebogiroa (ezkerrerantz biratzen dena), DNA motaren arabera. Bi kateak osagarriak dira, kate bateko adenina eta guanina beste kateko timina eta zitosinarekin elkartzen baitira, hurrenez hurren. Gainera, bi kate hauek antiparaleloak dira.
- Hiru DNA mota daude: DNA-A, DNA-B eta DNA-Z. B motakoa da ugariena eta Watson eta Crickek aurkitu zutena.
Hirugarren mailako egitura
- DNA leku murriztu batean antolatzeko moduari dagokiona da, kromosomak eratuz. Organismo eukarioto edo prokariotoek antolamendu desberdina dute.
- Prokariotoetan, DNA super-helize bat balitz gisa zabaltzen da zitoplasman; orokorrean, eraztun itxura izaten du, eta proteina kantitate txiki bati lotuta egoten da. Mitokondrio edo kloroplastoetan ere aurki daiteke modu berean.
- Eukariotoetan, kromosoma bakoitzean sartzen den DNA kantitatea oso handia denez, paketamendua konplexua, eta trinkoa da; horretarako beharrezkoa da proteinen presentzia.
Funtzioak
DNAren funtzio biologikoen artean, informazioa gordetzea (geneak eta genoma), proteinen kodifikazioa (transkripzioa eta itzulpena) eta bere bikoizketa (DNAren erreplikazioa) dira. Azken horren bidez, zatiketa zelularrean informazioa zelula alabetara transmitituko dela ziurtatzen da.
Geneak eta genoma
DNA bera bizi den organismoa eraiki eta mantentzeko beharrezkoa den informazioa gordetzen duen biltegia dela kontsidera genezake. Informazio hau, gero, belaunaldiz belaunaldi transmititua izango da. Organismoan funtzio hau betetzen duen informazioari genoma esaten zaio, eta, bertan dagoen DNAri, DNA genomikoa.
DNA genomikoa kromosometan antolatzen da, eta hori, batez ere, eukariotoen nukleo zelularrean dago; kantitate txiki bat ordea, mitokondrio eta kloroplastoetan ere badago. Prokariotoetan, nukleoide izeneko forma irregularreko gorputz batean dago DNA.[12]
DNA kodetzailea
Genomaren informazio genetikoa genetan dago, eta organismoko informazio guzti horri genotipo deritzo. Genea herentziaren unitatea eta organismo baten ezaugarri konkretu batean eragina duen DNA zatia da, esaterako, begien kolorean.
Herentziaren funtzio nagusia proteinen zehazketa da, DNA baita proteina horiek ekoizteko errezeta. DNAren aldaketak, gehienetan, disfuntzio proteiko bat dakar, eta honek gaixotasun bat sortuko du. Baina, zenbait kasutan, aldaketa horiek onurak ekarriko dituzte, eta, horren ondorioz, izakiak hobeto egokituko dira beren ingurunera.
Giza gorputzak 30.000 proteina inguru ditu, eta hauek 20 aminoazido desberdinez osatuta daude. DNA molekula bat da, aminoazido hauen sekuentzia zehaztuko duena hain zuzen ere. Proteinen sintesian, DNAren gene bat irakurri, eta RNAn transkribatzen da. RNA horrek, DNA eta proteinak sintetizatzen dituen makineriaren artean, mezulari gisa jarduten du. Horregatik, RNA mezulari edo mRNA izena ematen zaio. mRNA hori, proteinen sintesian, eredu gisa erabiltzen da aminoazidoak ordena egokian lotzeko proteinak eraikitzean.
Biologia molekularraren dogma nagusiak dio informazioak DNA → RNA → proteina norabidea jarraitzen duela. Aitzitik, gaur egun, dogma hori zabaldu egin behar dela badakigu beste informazio fluxu batzuk aurkitu direlako, hala nola RNA birusak. Kasu honetan, informazioa RNAtik DNAra doa, eta hori alderantzizko transkripzio izenarekin ezagutzen da. Gainera, RNAra transkribatu eta proteinak eman gabe funtzionalak diren DNA sekuentziak ere badaude. Horiek RNA ez-kodetzaileak dira.
DNA ez-kodetzailea
Organismo baten genomako DNA bi taldetan sailka daiteke: proteinak kodetzen dituena (geneak) eta kodetzen ez dituena. Espezie askotan, genomaren zati txiki batek bakarrik kodetzen ditu proteinak. Giza genomaren kasuan, % 1,5 inguru da proteinak kodetzeko gai; zati hauei exoi izena ematen zaie, eta 20.000–25.000 genek osatzen dute. Gainerako DNA ez-kodetzailea da, eta introi izena ematen zaie zati horiei.
DNA ez-kodetzailea proteinak sintetizatzeko gai ez diren DNA sekuentziak dira. Orain gutxi arte, DNA ez-kodetzaileak ez zuela erabilgarritasunik uste zen, baina azkenaldian egin diren ikerketek hori ez dela horrela erakutsi dute. Beste funtzioen artean, gene-adierazpena erregulatzen dute. Sekuentzia hauek "sekuentzia erregulatzaile" gisa ezagutzen dira, eta ikerlariek uste dute daudenetatik gutxi batzuk besterik ez direla identifikatu. Eukariotoen genoman hainbeste DNA ez-kodetzailearen presentzia eta espezieen arteko genoma tamainaren aldaketa misterio bat izaten jarraitzen du.
Bestalde, DNA sekuentzia batzuek egitura funtzioa dute kromosometan. Telomero eta zentromeroek proteinen gene kodifikatzaile gutxi dituzte, baina, kromosomen egitura egonkortzeko, garrantzitsuak dira.
Transkripzioa eta itzulpena
DNA harizpian agertzen den nukleotido sekuentzia mRNA batera transkribatzen da. Sekuentzia hau, aldi berean, organismoarentzat ezinbestekoa den proteina bat emateko itzultzen da.
Lehenik eta behin, DNAn dagoen informazioa RNAra pasa behar da. Horretarako, DNA helizearen kate bat erabiltzen da. Ondoren, RNAn dagoen informazioa proteinak sintetizatzeko erabiliko da. Nukleotido sekuentzia eta proteina osatzen duten amnioazido sekuentziaren arteko erlazioa kode genetikoak ezartzen du. Hori beharrezkoa da itzulpen edo proteinen sintesian. Kode genetikoaren unitate kodetzailea hiru nukleotidotako talde bat da. Hirukote hauetako nukleotido bakoitza hizki batez adierazten da, nukleotido horrek duen base nitrogenatuaren arabera. DNAren hirukoteak mRNA emanez transkribatzen dira. Hirukote hauei kodoi izena ematen zaie. Ondoren, mRNA-ren kodoi bakoitzak aminoazido bat emango du erribosoman. Aminoazido hauek elkarren artean lotuko dira proteina osatuz.
DNAren erreplikazioa

DNAren erreplikazioaren bidez, DNA molekularen kopia berdin-berdinak lortzen dira. Erreplikazioa beharrezkoa da informazio genetikoa belaunaldiz belaunaldi transmititua izateko. Horretarako, helizearen bi harizpiak banandu, eta kate berri bat sintetizatzen da bakoitzarentzat beraiek eredu gisa hartuz. Amaieran, hasierakoaren berdinak diren bi DNA molekula lortzen dira. Erreplikazio mota hau erdikontserbakorra izenez ezagutzen da kate bat mantendu eta bestea berria sintetizatzen delako.
Aldaketa kimikoak


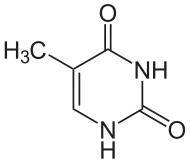
DNA-oinarrien aldaketak
Gene-adierazpena DNA kromosometan biltzeko moduak eragiten du, kromatina izeneko egituran. Base-aldaketek parte har dezakete DNAren bilketan: gene-adierazpen baxua edo hura ez duten eskualdeek, normalean, zitosinaren oinarrien metilazio-maila handia izaten dute. Adibidez, zitosinaren metilizazioak 5-metil-zitosina sortzen du, eta hori garrantzitsua da X kromosomaren inaktibaziorako[13]. Batezbesteko metilazio-maila aldatu egiten da organismoen artean: Caenorhabditis elegans harrak ez du zitosinaren metilaziorik, eta ornodunek, berriz, maila altua dute —% 1 arte—; beren DNAren 5-metil-zitosina dauka[14]. 5-metil-zitosinak garrantzia izan arren, desaminatu daiteke timina-base bat sortzeko. Metilatutako zitosinak, beraz, bereziki sentikorrak dira mutazioen aurrean[15]. Zinetoplastoetan J-basea sortzeko, beste oinarri-aldaketa batzuk bakterioetan adeninaren metilazioa eta urazilaren glikosilazioa dira[16][17].
DNA kaltetua
[[Fitxategi:Benzopyrene DNA adduct 1JDG.png|thumb|Benzopireno molekula, esaterako, [[tabako|tabakoaren] kean dagoen mutagenoa, DNA helize bati lotuta.[18]]] DNAren sekuentzia aldatzen duten mutageno mota askoren ondorioz kaltetu daiteke: agente alkilatzaileak, baita energia handiko erradiazio elektromagnetikoak ere, argi ultramoreak eta X izpiak, adibidez. Sortzen den DNAren kalte mota mutageno motaren araberakoa da. Adibidez, UV argiak DNA kaltetu dezake timina-dimeroak sortuz, Pirimidina-baseen arteko gurutzaketa-loturaz sortzen direnak[19]. Bestalde, erradikal askeak edo hidrogeno peroxido izeneko oxidatzaileek kalte ugari sortzen dituzte, oinarrien aldaketak, batez ere guanina, eta kate bikoitzeko hausturak barne (double-strand breaks)[20]. Edozein giza zeluletan, 500 base inguruk kalte oxidatiboa jasaten dute egunero[21][22]. Kalte oxidatibo hauetatik, kate bikoitzeko hausturak dira arriskutsuenak, konpontzen zailak baitira, eta DNAren sekuentziaren mutazio puntualak, txertaketak eta delezioak sor ditzakete, baita translokazio kromosomikoak ere[23].
Mutageno asko aldameneko bi base-pareren artean kokatzen dira; horregatik, agente interkalatzaileak deitzen zaie. Agente interkalatu gehienak molekula lauak eta aromatikoak dira, hala nola etidio bromuroa, daunomicina, doxorrubizina eta talidomida. Agente interkalatzaile bat bi base pareren artean integratzeko, banandu egin behar dira DNA kateak desitxuratuz eta helize bikoitza irekiz. Honek transkripzioa eta DNAren erreplikazioa galarazten ditu toxikotasuna eta mutazioak eraginez. Hori dela eta, DNAren agente interkalanteak agente kartzinogenoak izan ohi dira: benzopirenoa, akridina, aflatoxina eta etidio bromuroa dira adibide ezagunak[24][25][26]. Hala ere, DNAren erreplikazioak eta DNAren transkripzioak inhibitzeko duten gaitasuna dela eta, toxina hauek ere erabiltzen dira kimioterapia minbizi-zelulen hazkuntza azkarra eragozteko[27].
DNA kaltetuak DNAren lesio espezifikoak ezagutzen dituzten konponketa mekanismo desberdinak aktibatzen dituen erantzuna abiarazten du, eta, jatorrizko DNA sekuentzia berreskuratzeko, berehala konpontzen dira. Era berean, DNA kaltetuak zelula-zikloaren geldialdia eragiten du, eta horrek prozesu fisiologiko ugari aldatzea dakar, eta, aldi berean, sintesia, eta proteinen garraio eta degradazioa (ikus DNA kaltetuak kontrolatzeko puntua). Bestela, kalte genomikoa konpontzeko handiegia bada, kontrol-mekanismoek zelulen heriotzarekin amaituko den bide zelularraren aktibazioa eragingo dute.
Oharrak
- Euskaraz, badirudi zientziarako laburtzapenik erabiliena eta egokiena DNA dela gaur egun:
- Dokumentu honetan, esaten dute euskal herritar zientzialarien artean «forma kanonikoa» dela DNA: Kepa Altonaga (Zoologia eta Animali Zelulen Biologia Saila. Zientzia eta Teknologia Fakultatea. EHU/UPV): «Txontak eta txolarreak: gogoeta bat ereduzko prosa zientifikoaren gainean» (2008. urtea).
- Beste honetan (Alfontso Mujika: «Aholkularitza-lanaren ustiaketa didaktikoa», 2008. urtea), zientzietako euskal terminologian jakintsuenetakoa dugun Alfontso Mujikak adierazi du «arau/gomendio/aholku» hau aplikatzen dutela Elhuyar Fundazioak eginiko testuetan:
EZ BAI (biologian)
ARN, ADN, gene-adierazpena(biologian)
RNA, DNA, gene-espresioa- Elhuyarkoek irizpide hori erabiltzen dutela baieztatzen da erakunde hartako kide diren Antton Gurrutxagak, Saroi Jauregik eta Alfontso Mujikak 2004. urtean idatzitako beste artikulu honetan (Antton Gurrutxaga, Saroi Jauregi, Alfontso Mujika: «Zuzenketen kudeaketa eta hizkuntz zerbitzuak», Senez, 27. zenbakia, 2004) ere: «Elhuyarren irizpidea da ingelesezko siglak erabiltzea: RNA eta DNA».
- EIMAk, Biologiako ikasliburuetarako, DNA forma erabiltzea hobesten du (EIMA: Zientzia eta teknikako euskara arautzeko gomendioak, Eusko Jaurlaritzaren Argitalpen Zerbitzu Nagusia, 2011).
- Azkenik, Berria egunkarian bilaketa eginda (2010-05-17ko 22:00ak diren honetan), 106 aldiz azaltzen da «ADN», eta 317 aldiz «DNA». Argia aldizkariko bilagailuak ez du zenbaketarik egiten, baina Googlen «ADN site:argia.com» bilaketa eginda (egun eta ordu berean), 99 emaitza ateratzen dira; «DNA site:argia.com» bilaketarekin, berriz, 165.
Erreferentziak
- (Gaztelaniaz) Malavé, Dr Antonio Alcalá. (2015-11-04). Genética de la emoción: El origen de la enfermedad. Penguin Random House Grupo Editorial España ISBN 978-84-9069-206-6. (Noiz kontsultatua: 2020-02-18).
- (Gaztelaniaz) Mazzotta, Guillermo Cejas. (2000). Identificación por ADN. Ediciones Jurídicas Cuyo ISBN 978-987-527-014-5. (Noiz kontsultatua: 2020-02-18).
- «Publication Name:» web.archive.org 2013-05-18 (Noiz kontsultatua: 2020-02-18).
- (Ingelesez) Dahm, Ralf. (2005-02-15). «Friedrich Miescher and the discovery of DNA» Developmental Biology 278 (2): 274–288. doi:. ISSN 0012-1606. (Noiz kontsultatua: 2020-02-18).
- «Building Life On Earth» www.terradaily.com (Noiz kontsultatua: 2020-02-18).
- Dahm, Ralf. (2008-01). «Discovering DNA: Friedrich Miescher and the early years of nucleic acid research» Human Genetics 122 (6): 565–581. doi:. ISSN 1432-1203. PMID 17901982. (Noiz kontsultatua: 2020-02-18).
- (Ingelesez) Levene, P. A.. (1919-12-01). «THE STRUCTURE OF YEAST NUCLEIC ACID IV. AMMONIA HYDROLYSIS» Journal of Biological Chemistry 40 (2): 415–424. ISSN 0021-9258. (Noiz kontsultatua: 2020-02-18).
- «Wayback Machine» web.archive.org 2014-03-03 (Noiz kontsultatua: 2020-02-18).
- Astbury, W. (1947). «Nucleic acid». Symp. SOC. Exp. Bbl 1 (66).
- Lorenz, M G; Wackernagel, W. (1994-09). «Bacterial gene transfer by natural genetic transformation in the environment.» Microbiological Reviews 58 (3): 563–602. ISSN 0146-0749. PMID 7968924. (Noiz kontsultatua: 2020-02-18).
- «Journal of Experimental Medicine | Rockefeller University Press» rupress.org (Noiz kontsultatua: 2020-02-18).
- (Ingelesez) Thanbichler, Martin; Wang, Sherry C.; Shapiro, Lucy. (2005). «The bacterial nucleoid: A highly organized and dynamic structure» Journal of Cellular Biochemistry 96 (3): 506–521. doi:. ISSN 1097-4644. (Noiz kontsultatua: 2020-02-18).
- Klose, Robert J.; Bird, Adrian P.. (2006-02). «Genomic DNA methylation: the mark and its mediators» Trends in Biochemical Sciences 31 (2): 89–97. doi:. ISSN 0968-0004. (Noiz kontsultatua: 2023-09-24).
- (Ingelesez) Bird, Adrian. (2002-01-01). «DNA methylation patterns and epigenetic memory» Genes & Development 16 (1): 6–21. doi:. ISSN 0890-9369. PMID 11782440. (Noiz kontsultatua: 2023-09-24).
- (Ingelesez) Walsh, C. P.; Xu, G. L.. (2006). Doerfler, Walter ed. «Cytosine Methylation and DNA Repair» DNA Methylation: Basic Mechanisms (Springer): 283–315. doi:. ISBN 978-3-540-31390-8. (Noiz kontsultatua: 2023-09-24).
- (Ingelesez) Ratel, David; Ravanat, Jean-Luc; Berger, François; Wion, Didier. (2006-03). «N6-methyladenine: the other methylated base of DNA» BioEssays 28 (3): 309–315. doi:. ISSN 0265-9247. PMID 16479578. PMC PMC2754416. (Noiz kontsultatua: 2023-09-24).
- Gommers-Ampt, Janet H.; Van Leeuwen, Fred; de Beer, Antonius L.J.; Vliegenthart, Johannes F.G.; Dizdaroglu, Miral; Kowalak, Jeffrey A.; Crain, Pamela F.; Borst, Piet. (1993-12). «β-d-glucosyl-hydroxymethyluracil: A novel modified base present in the DNA of the parasitic protozoan T. brucei» Cell 75 (6): 1129–1136. doi:. ISSN 0092-8674. (Noiz kontsultatua: 2023-09-24).
- PDB 1JDG-tik sortua
- (Ingelesez) Douki, Thierry; Reynaud-Angelin, Anne; Cadet, Jean; Sage, Evelyne. (2003-08-01). «Bipyrimidine Photoproducts Rather than Oxidative Lesions Are the Main Type of DNA Damage Involved in the Genotoxic Effect of Solar UVA Radiation» Biochemistry 42 (30): 9221–9226. doi:. ISSN 0006-2960. (Noiz kontsultatua: 2023-09-24).
- Cadet, J., Delatour, T., Douki, T., Gasparutto, D., Pouget, J., Ravanat, J., Sauvaigo, S. (1999). «Hydroxyl radicals and DNA base damage». Mutat Res 424 (1-2): 9-21. PMID 10064846
- Shigenaga, M., Gimeno, C., Ames, B. (1989). «Urinary 8-hydroxy-2′-deoxyguanosine as a biological marker of in vivo oxidative DNA damage». Proc Natl Acad Sci U S A 86 (24): 9697-701. PMID 2602371.
- (Ingelesez) Cathcart, R; Schwiers, E; Saul, R L; Ames, B N. (1984-09). «Thymine glycol and thymidine glycol in human and rat urine: a possible assay for oxidative DNA damage.» Proceedings of the National Academy of Sciences 81 (18): 5633–5637. doi:. ISSN 0027-8424. PMID 6592579. PMC PMC391764. (Noiz kontsultatua: 2023-09-24).
- (Ingelesez) Valerie, Kristoffer; Povirk, Lawrence F.. (2003-09). «Regulation and mechanisms of mammalian double-strand break repair» Oncogene 22 (37): 5792–5812. doi:. ISSN 1476-5594. (Noiz kontsultatua: 2023-09-24).
- Ferguson, L., Denny, W. (1991). «The genetic toxicology of acridines». Mutat Res 258 (2): 123-60. PMID 1881402
- Jeffrey, Alan M.. (1985-01-01). «DNA modification by chemical carcinogens» Pharmacology & Therapeutics 28 (2): 237–272. doi:. ISSN 0163-7258. (Noiz kontsultatua: 2023-09-24).
- Stephens, Trent D; Bunde, Carolyn J. W; Fillmore, Bradley J. (2000-06-01). «Mechanism of action in thalidomide teratogenesis» Biochemical Pharmacology 59 (12): 1489–1499. doi:. ISSN 0006-2952. (Noiz kontsultatua: 2023-09-24).
- (Ingelesez) Brana, M. F.; Cacho, M.; Gradillas, A.; Pascual-Teresa, B. de; Ramos, A.. «Intercalators as Anticancer Drugs» Current Pharmaceutical Design 7 (17): 1745–1780. doi:. (Noiz kontsultatua: 2023-09-24).
Bibliografía
- Calladine, Chris R; Drew, Horace R; Ben, F; Travers, Andrew A. (2003). Understanding DNA. Elsevier Academic Press ISBN 978-0-12-155089-9..
- Clayton. (2003). 50 Years of DNA. Palgrave MacMillan Press ISBN 978-1-4039-1479-8..
- Judson, Horace Freeland. (1996). The Eighth Day of Creation: Makers of the Revolution in Biology. Cold Spring Harbor Laboratory Press ISBN 978-0-87969-478-4..
- Olby, Robert. (1974). The Path to The Double Helix: Discovery of DNA. ISBN 978-0-486-68117-7..
- Ridley, Matt. (2006.). Francis Crick: Discoverer of the Genetic Code (Eminent Lives). HarperCollins Publishers, 192 orrialde ISBN 978-0-06-082333-7..
- Rose, Steven. The Chemistry of Life. Penguin, 273-274. or. ISBN 978-0-14-027..
- Watson, James D; Crick, Francis H. C.. (1953-04-25). ««A structure for Deoxyribose Nucleic Acid.» (PDF)» Nature 171: 737-738...
- Watson, James D. DNA: The Secret of Life. ISBN 978-0-375-41546-3..
- Watson. The Double Helix: A Personal Account of the Discovery of the Structure of DNA. Norton Critical Editions. isbn=978-0-393-95075-5.
- Watson, James D. (2007). Avoid boring people and other lessons from a life in science. ISBN 978-0-375-41284-4..
Ikus, gainera
Kanpo estekak
| Autoritate kontrola |
|
|---|