Burmuin
Burmuina[1] entzefaloaren zatirik handiena eta garrantzitsuena da, goi mailako ornodunengan eta, bereziki, gizakiarengan garapen eta konplexutasun handia duena. Nerbio-sistemaren egitura zuzendaria da.
| Burmuin | |
|---|---|
 | |
| Xehetasunak | |
| Honen parte | burua Nerbio-sistema zentrala sistema neurobaskularra |
| Kokapena | cranial cavity (en) |
| Konponenteak | Prosenzefaloa mesentzefaloa enbor entzefalikoa erronbentzefaloa |
| Artikulazioa | Bizkarrezur-muina Nerbio optikoa cranium (en) |
| Identifikadoreak | |
| Latinez | cerebrum |
| MeSH | A08.186.211 |
| TA | A14.1.03.001 |
| Terminologia anatomikoa | |
Ugaztunen entzefalo barruan, burmuina prosentzefaloan dago, hots, eboluzioaren eskalan berriagoa den egituran. Entzefaloaren beste egitura zaharragoek, metentzefaloak eta mielentzefaloak (erronbentzefaloa osatzen dutenak) ez dute adimen-funtziorik garatzen, gorputzaren funtzio mekanikoez arduratzen baitira (mugimenduen koordinazioaz, arnas eta bihotz-jardueraren kontrolaz, etab.).
Burmuineko neuronen kopurua egundokoa da: ikertzaileak ez dira ados jartzen, baina 50.000 milioi eta bilioi bat artean dagoela uste dute. Era berean, burmuineko neurona bakoitzak milaka harreman sinaptiko eratzen ditu aldameneko neuronekin. Zahartu ahala, ordea, neuronen kopurua jaisten da.
Izena
Euskara batuan, organo honen izen hobetsia "burmuin" da.[2] Horretaz gain, batez ere Hegoaldean, burmuinari garun[3] —Euskaltzaindiaren Hiztegiaren arabera, "entzefalo" adigaitik gertuago dagoena—[4] edo zerebro[5] —etekin txikiko irtenbidetzat jo izan dena—[6] esatea ohikoa da. Iparraldean, "burmuin" deitzeaz gain, nafar-lapurteraz burumuin[7] eta zubereraz bürün esaten diote.[3] Nafar-lapurterazko testuetan, buru muin edo buru fuin aldaerak, bi hitzetan idatzirik, agertzen dira oroz gainetik, eta pluralean.[3]
Ezaugarri anatomikoak
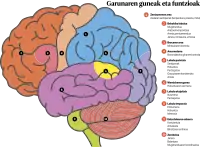
Garuna bi zelula-motaz osatua dago: neuronak eta neuroglia. Neuronak bata besteari lotzen zaizkio, neurona-zirkuituak sortzeko. Entzefaloa burezurrak babesten du eta horrez gain, meningeek estaltzen dute. Garunean bi atal bereiz daitezke: aurreko garuna eta atzeko garuna[8].
Aurreko garuna edo prosentzefaloa
Entzefaloaren aurrealdeko bi herenak hartzen ditu. Garuneko luzetarako fisura izeneko ildo sakon batek bereizirik, bi burmuin-hemisferiotan banatzen da. Garun-hemisferioak beheko aldetik lotuta daude, bata bestearekin konektatzen dituen gorputz kailukara deituriko nerbio-zuntzen bidez[8].
Hemisferio bakoitzaren barruan, alboko bentrikulu izenez ezagutzen diren likidoz beteriko tarte batzuk daude. Alboko bentrikulu hauek, hemisferio bien artean dagoen hirugarren bentrikulu batekin lotzen dira, eta hirugarren bentrikulu hau laugarren batera doa, Sylviusen akueduktu deitzen den kanal fin baten bidez. Bentrikulu hauen barnean dagoen eta nerbio-sistema inguratzen duen likido zefalorrakideoak babes mekaniko gisa eta substantzia kimikoak garraiatzeko balio du. Likido zefalorrakideo hau alboko bentrikuluetan sortzen da, plexu koroideoak osatzen dituzten egitura baskularretan[8].
Garunaren zati azalekoena burmuin-azalak osatzen du. Hemisferioen azalean zabaltzen den substantzia grisezko geruza bat da. Garunaren barrualdea substantzia zuriz inguraturiko oinaldeko nukleo grisez, bi hemisferioak lotzen dituzten ertzetako formazioez, eta barrunbe (edo hutsune) bentrikularrez osatuta dago[8].
Atzeko garuna edo erronbentzefaloa
Bizkarrezur-muinaren gaineko zatian dago eta bi egiturak osatzen dute: entzefalo-enborrak eta garuntxoak. Bien artean laugarren bentrikulua dago[8].
Hainbat zirkunboluziodun egitura da garuntxoa, eta entzefalo-enborrera zuntz aferenteen bidez lotua dago. Zuntz hauek hezur-muineko, erraboileko, zubiko eta erdialdeko eta aurreko garuneko bulkadak eramaten dizkiote. Era berean, garuntxoko nukleoetatik zuntz eferenteak sortzen dira. Borondatezko mugimenduen zehaztasun eta koordinazioan esku hartzen du garuntxoak, eta jarrera eta oreka mantentzeko ezinbestekoa da. Entzefalo-enborra entzefaloaren beheko aldean dago eta arnasa, eztula, bihotz-taupadak eta goragale-erreflexua gobernatzen ditu. Haren lesioa larria da eta gizabanakoaren heriotza ekar dezake, aipatutako bizi-funtzioen asalduraren ondorioz[8].
Substantzia grisa eta substantzia zuria
Substantzia grisa
Soma edo gorputz neuronalak aurkitzen diren tokia da. Substantzia grisak burmuin-hemisferioen geruza azalekoena osatzen du, baina bada barrurago dagoen substantzia grisik ere. Barruragoko substantzia honek entzefaloaren oinaldeko nukleoak eta bikote kranialen nukleoak osatzen ditu[8].
Garun-azalak edo kortex zerebralak 2-3 mm-ko lodiera du, eta hamar zelula-geruzaz osaturik dago. Toles ugari (burmuin-zirkunboluzioak) izatearen ondorioz, burmuin-azala burezur-azala baino 30 aldiz handiagoa da. Zimurrek eta pitzadurek hemisferioak lobuluetan banatzen dituzte. Lobulu hauetariko lau frontala, parietala, tenporala eta okzipitala deitzen dira. Bosgarren lobulua, intsula, ezin da burmuinaren kanpoaldetik ikusi, oso barrenean kokatuta dagoelako[8].
Entzefaloaren oinaldeko nukleoak entzefaloaren oinalde inguruan dauden substantzia grisaren formazioak dira. Nukleoak honakoetan banatzen dira: talamoa, nukleo kaudatua, nukleo lentikularra, putamena, palliduma (azken bi hauek batera nukleo ildaskatua osatzen dute) eta claustruma. Talamoa forma oboideko substantzia grisezko nukleo handi bat da, burmuinaren erdialdean, bi burmuin-hemisferioen artean kokatua. Seinale sentsorialak jasotzen dituen integrazio-gune garrantzitsua da. Hainbat nukleoz osatua dago: aurrekoa, atzekoa, bentrala eta dortsala. Garunera doazen bide aferente guztiak, usaimenezkoak izan ezik, talamora heltzen dira. Bulkadak azalera eta beste gune batzuetara eramaten dituzten hainbat zuntz sortzen dira talamotik. Entzefaloaren oinaldeko nukleoen funtzioa nahi gabeko mugimenduak integratu eta koordinatzea da. Nukleo hauen lesioak mugimendu-desorekak, korea (San Vitoren dantza) eta balismoa adibidez, edo Parkinsonen gaixotasunean gertatzen den mugimendu-murriztea (hipozinesia) sor ditzake[8].
Substantzia zuria
Garunean substantzia grisa baino barrurago dagoen substantzia da, eta nerbio-zuntzez osatua dago batik bat. Zuntz hauek burmuin-azaleko hainbat puntu bata bestearekin lotzen dituzte, edota burmuin-azala bera entzefalo-enborreko eta ornomuineko hainbat nukleorekin. Uniformeki zabaltzen da burmuin-azalaren azpitik eta laminatan banatzen da; barneko, kanpoko eta muturreko kapsula osatzen ditu[8].
Ertzetako formazioak bi hemisferioak lotzen dituzten zuntz-sistemak dira. Gorputz kailukara, fornixa edo trigonoa, aurreko ertz zuria eta septum peluciduma (septu edo trenkada peluzidoa) dira[8].
Garapena
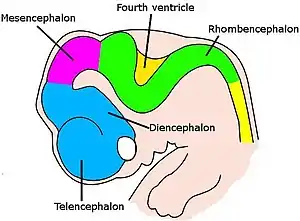
Burmuina fase-sekuentzia korapilatsu orkestratu batean garatzen da[9]. Enbrioi-etaparik goiztiarrenetan nerbio-sokaren aurrealdeko hantura sinple batetik eremu eta konexio multzo konplexu batera aldatzen da forman. Neuronak zelula amak dituzten gune berezietan sortzen dira, eta, gero, ehunetik migratzen dute beren azken kokapenetara iristeko. Neuronak kokatu ondoren, haien axoiak sortu, eta burmuinean zehar nabigatzen dute adarkatuz eta zabalduz aurrera egiten duten heinean, harik eta puntak beren helburuetara iritsi eta konexio sinaptikoak eratzen dituzten arte. Nerbio-sistemaren hainbat lekutan, neuronak eta sinapsiak erruz sortzen dira lehen faseetan, eta, gero, behar ez direnak eliminatu egiten dira[9].
Ornodunei dagokienez, neurona-garapenaren hasierako faseak antzekoak dira espezie guztietan[9]. Enbrioia zelula-bloke biribil batetik zizare itxurako egitura bihurtzen den heinean, bizkarreko erdiko lerrotik doan ektodermo-zerrenda estu bat eragiten da nerbio-sistemaren aitzindari den plaka neuronal bihurtzeko. Plaka neuronala barrurantz tolesten da neurona-lerroa osatzeko, eta, gero, zimurra marrazten duten ezpainek bat egiten dute neurona-hodia ixteko, zelula-lokarri huts bat erdian fluidoz betetako bentrikulu bat duelarik. Aurreko muturrean, bentrikuluak eta kordoia puztu egiten dira, prosentzefaloa (aurre-garuna), mesentzefalo (erdi-garuna) eta erronbentzefaloa (gibel-garuna) aitzindariak diren hiru besikula osatzeko. Hurrengo fasean, burmuina bi besikulatan banatzen da: telentzefaloa (garun-kortexa, ganglio basala eta antzeko egiturak izango dituena) eta dientzefaloa (talamoa eta hipotalamoa izango dituena). Aldi berean, burmuina mesentzefaloan (zerebeloa eta ponsak izango dituena) eta mielentzefaloan (bizkarrezur-erraboila izango duena) banatzen da. Eremu horietako bakoitzak neuronak eta glia-zelulak sortzen diren eremu ugalkorrak ditu; ondoren, ondoriozko zelulak migratzen dute, batzuetan distantzia luzeetara, beren azken kokapenetara[9].
Behin neurona bat bere lekuan dagoenean, dendritak eta axoi bat zabaltzen ditu inguruan. Axoiak, normalean, modu bereziki konplexuan hazten dira gorputz zelularrarekiko distantzia handia hedatzen dutelako eta helburu zehatzak lortu behar dituztelako. Hazten ari den axoi baten punta, izan ere, hazkuntza-kono izeneko protoplasma bloke batek osatzen du, hartzaile kimikoz josia. Hartzaile horiek tokiko ingurunea nabaritzen dute hazkunde-konoa elementu zelular ezberdinek erakartzea edo uxatzea eraginez, eta, horrela, norabide jakin batean erakartzen dute bere bidearen puntu bakoitzean. Patologia horren ondorioz, hazkunde-konoa burmuinean zehar nabigatzen du helmugara iritsi arte, non beste seinale kimiko batzuek sinapsiak sortzen dituzten. Garun osoa kontuan hartuta, milaka genek patologia axonaletan eragina duten produktuak sortzen dituzte[9].
Azkenean sortzen den sare sinaptikoa geneek bakarrik determinatzen dute aldez. Burmuineko alde askotan, axoiak gehiegi hazten dira hasieran, eta, gero, inausi egiten dira neuronen jardueraren menpe dauden mekanismoen bidez[9]. Begitik mesentzefalorako proiekzioan, adibidez, helduaren egiturak mapeaketa oso zehatza du, erretinaren azaleko puntu bakoitza mesentzefaloko geruza batean dagokion puntu batekin lotzen duena. Garapenaren lehen faseetan, erretinatik datorren axoi bakoitza mesentzefaloren eskuineko inguru orokorrera eramaten da, baina, gero, adarkatze handiak sortzen ditu, eta mesentzefaloko neuronen multzo zabal batekin kontaktuan jartzen da. Erretinak, jaio aurretik, mekanismo bereziak ditu, eta, horien ondorioz, berez sortzen diren aktibitate-uhinak sortzen dira ausaz, eta, gero, mantso-mantso hedatzen dira erretina-geruzan zehar. Uhin horiek erabilgarriak dira ondoko neuronak, aldi berean, aktibo egotea eragiten dutelako; hau da, neuronen antolaketa espazialari buruzko informazioa duen aktibitate eredu neuronal bat sortzen dute. Informazio horretaz, mesentzefaloa baliatzen da sinapsiak ahultzen dituen mekanismo baten bidez, eta, azkenean desagertu egiten da, baldin eta axoi batean jarduera ez badu xede-zelularen jarduera jarraitzen. Prozesu sofistikatu horren emaitza da mapeaketa pixkanaka doitzea eta estutzea, azkenean bere heldu-forma zehatzean utziz[10].
Antzeko gauzak gertatzen dira garuneko beste eremu batzuetan: matrize sinaptiko iniziatiko bat sortzen da genetikoki zehaztutako gida kimikoaren ondorioz, baina, gero, pixkanaka, jardueraren menpeko mekanismoek fintzen dute, neurri batean barne-dinamikak edo kanpoko ekarpen sentsorialek bultzatuta. Kasu batzuetan, erretina-mesentzefalo sistemarekin gertatzen den antzera, aktibitate ereduak garunean bakarrik funtzionatzen duten mekanismoen menpe daude, eta, dirudienez, garapena gidatzeko bakarrik existitzen dira[10].
Gizakietan eta beste ugaztun askotan, neurona berriak batez ere jaio aurretik sortzen dira, eta haurren garunak garun helduak baino neurona gehiago ditu[9]. Hala ere, badira eremu batzuk, non neurona berriak bizitzan zehar sortzen diren. Helduentzako, neurogenesia ongi finkatuta dagoen bi eremuak usaimen-fokua eta hippocampuse deniseko hortz-giroa dira, non ebidentziak dauden ezen neurona berriek paper bat betetzen dutela eskuratu berri diren oroitzapenak gordetzean. Baina salbuespen horiekin, lehen haurtzaroan dagoen neurona-multzoa da bizitza osorako dagoena. Zelula glialak desberdinak dira: hau da, gorputzeko zelula mota gehienetan, bizitza osoan zehar sortzen dira[11].
Luzaroan eztabaidatu izan da ea gogoaren, nortasunaren eta adimenaren nolakotasunak herentziari edo heziketari egotzi ahal zaizkion; hau da naturari eta elikadurari buruzko eztabaida[12]. Oraindik xehetasun asko zehazteke dauden arren, neurozientziako ikerketek argi erakutsi dute bi faktore horiek garrantzitsuak direla. Garunaren forma orokorra eta burmuinak, esperientziaren aurrean, nola erreakzionatzen duen geneek zehazten dute. Hala ere, esperientzia behar da lotura sinaptikoen matrizea fintzeko, zeinak bere forma garatuan genomak baino askoz informazio gehiago baitu. Zenbait alderditan, garapen aldi kritikoetan esperientziarik ez izatea da axola duen gauza bakarra[13]. Beste alde batetik, esperientziaren kantitatea eta kalitatea garrantzitsuak dira; adibidez, funtsezko ebidentzia dago ingurune aberastuetan hazitako animaliek garun-kortex lodiagoak dituztela (lotura sinaptikoen dentsitate handiagoa adierazten dutenak) estimulazio-maila murriztua duten animaliek baino[14].
Sistema linbikoa
Garunaren erdian kokatzen da eta honako atal hauetaz osatua dago: talamoa, hipotalamoa, hipokanpoa, amigdala, gorputz kailukara, septuma eta mesentzefaloa. Garuneko unitate funtzional bat da. Sistema linbikoak interakzio biokimiko eta nerbio-interakzio estuak ditu burmuin-azalarekin eta memoria, emozioak, atentzioa eta ikasketaz arduratzen den entzefaloko elementua da[8].
Hipotalamoa talamoaren azpian dago, burmuin-oinarriaren erdiko lerroan. Hainbat gunez eta nukleoz osatua dago. Hauen zeregina oinarrizko bulkadak eta gorputzaren barne-egoeraren faktoreak (tenperatura, gosea, egarria, logura, etab.) erregulatzea da. Hipotalamoak nerbio-sistema zentrala eta sistema endokrinoaren arteko lotura ere egiten du, hainbat hormona sortzen dituen hipofisiaren funtzionamendua erregulatzen baitu[8].
Funtzioak

Garunak informazio sentsoriala prozesatzen du, mugimendua kontrolatu eta koordinatzen du, portaera eta gorputzeko oinarrizko funtzioak, hala nola bihotz-taupadak, odolaren presioa, fluidoen balantzea eta gorputzeko tenperatura kontrolatzen ditu. Garuna ezagutzaren, emozioen, memoriaren eta ikasketaren arduraduna da[8].
Garun-kortexa funtzioaren araberako eremuetan banatzen da. Brodmannek deskribatu zituen eremu hauek eta horregatik Brodmannen area gisa ere ezagunak dira[8].
Mugikortasuna
Area motorretan antolatua dago. Nagusia area motorra edo Brodmannen 4. area da. Erdiko artekaren edo Rolandoren arteka deiturikoaren aurrean dago. Zelula piramidal gisa ezagutzen diren neurona erraldoiak ditu. Zelula horietatik ornomuinera eta entzefalo-enborrera doazen bide motorrak sortzen dira eta entzefalo-enborretik informazioa giharretaraino igortzen da. Borondatezko mugimenduen arduradunak dira. Area motorretik sortzen diren zuntzak bide piramidal gisa ezagutzen dira eta kontrako aldera gurutzatzen dira erraboilean, entzefalo-enborraren alde baxuenean. Horregatik, eskuin hemisferioko lesioek ezkerreko gorputz-erdiko indar-galera dakarte, eta alderantziz[8].
Bada beste area bat, Rolandoren artekaren aurrean dagoena, area premotorra, mugimendu ez-borondatezkoak antolatzeko ardura duena. Area edo eremu honetan lesioak gertatzeak pertsonak ikasitako trebetasunen galera dakar[8].
Sentikortasuna
Area edo eremu sentsorialetan antolatzen da. Area honetan amaitzen dira gorputzaren hainbat bulkada bidaltzen dituzten zuntz sentikorrak. Besteak beste, ikusmen-bulkadak, entzumenezkoak eta usaimenezkoak eta, horiez gain, larruazalaren eta organoen azaleko sentsibilitatea eta sentsibilitate sakona bidaltzen dituzte. Aipatu diren areak honela daude banatuta[8]:
- Area somestesikoa
- gorputzaren beste aldeko azaletik, ehunetatik, muskuluetatik eta artikulazioetatik datorren sentikortasun orokorra jasotzen du. Atzeko zirkunboluzio zentralean kokatzen da, Rolandoren artekaren atzean. Area honetako neuronak objektuen arteko pisu-ezberdintasunak hautemateko gai dira (barognosia). Horrez gain, azaleko estimuluen jatorria zehazteko eta estimulu horiek bakarrak edota askotarikoak diren bereizteko gaitasuna dute aipatutako neuronek; objektuak bere tamainaren eta formaren bidez ezagutzeko (esterognosia) gaitasuna ere badute, eta objektuen tenperatura-aldeak bereizteko ere gai dira, besteak beste.
- Ikusmen-area
- lobulu okzipitalean dago, burmuinaren atzealdean. Betsaretik jasotako irudiak ispilu-eran proiektatzen dira burmuin-azalean: ikus-eremuaren goialdean geratzen dena burmuin-azalaren beheko aldera doa eta behean ikusten duguna goiko gune kortikalera doa. Beste horrenbeste gertatzen da eskuinaldean eta ezkerraldean ikusten dugunarekin. Bada ikusten duguna prozesatzen duen lehen mailako ikus-eremu bat, eta baita zeregin landuagoak egiten dituen bigarren mailako ikus-eremu bat ere. Bigarren mailako ikus-eremu honek ikusitako objektuak ezagutzea, aurpegiak ezagutzea, lengoaiaren eremuekin asoziazioa egitea, mugimenduaren pertzepzioa eta abar ahalbidetzen du.
- Entzumen-area
- lobulu tenporalean kokatua, alboko edo Sylviusen artekaren azpian. Belarri bakoitzak aurkezpen bikoitza du burmuin-azalean, nekez gerta daiteke, beraz, pertsona batek gorreri kortikala izatea. Bada entzumen-area nagusi bat, entzumenezko informazioa prozesatzen duena, eta baita entzuten ari garena (abesti bat, adibidez) ezagutu eta burmuin-azaleko beste area batzuekin lotzen duen bigarren mailako entzumen-area bat ere.
- Usaimen-area
- lobulu tenporalen behealdean dago, entzumen-areatik gertu.
- Dastamen-area
- lobulu tenporalen beheko muturrean kokatzen da.
Norberaren kontzientzia
Norberaren kontzientzia, arima, nortasuna, jokaera, esperientzia afektiboa, iniziatiba, aurrealdeko lobuluetan integratuta daude[8].
Hizkuntza
Beste funtzio batzuek ez bezala, hizkuntzak alde bakarrekoak diren area kortikalak betetzen ditu. Area kortikal hauek kokatzen diren hemisferioari hemisferio gainartzaile deitzen zaio, eta normalean ezkerrekoa izaten da, nahiz eta pertsona ezkertiek eskuineko dominantzia edota bi aldeetakoa eduki ahal izan. Eremu hauek honela banatzen dira[8]:
Area motorra edo Brocaren area: aurreko lobuluaren beheko aldean dago eta hizkuntzaren adierazpenaz arduratzen da (bai ahozkoaz eta bai idatzizkoaz). Area honetan lesio bat izanez gero, hitz egiteko eta idazteko ezintasuna pairatzen da.
Sentitze-area edo Wernickeren area: lobulu tenporalaren goiko aldean dago eta hizkuntzaren ulermenaz arduratzen da (bai ahozkoaz eta bai idatzizkoaz). Area honetan lesio bat izanez gero, esaten digutena eta irakurtzen ari garena ulertzeko ezintasuna pairatzen da.
Fisiologia
Garunaren fisiologia ongi ulertzeko azpimarratu behar da nerbio-zuntzak (zentzumenezkoak zein motorrak ) gurutzatuta daudela. Hots, gorputzaren eskuinaldeko zentzumen-guneak eta gune motorrak burmuineko ezkerreko hemisferioan daudela, eta alderantziz. Horrek esan nahi du burmuinaren eskuinaldeak gorputzaren ezkerraldea kontrolatzen duela, eta burmuinaren ezkerraldeak gorputzaren eskuinaldea.
Ulertzekoa da, beraz, pertsona batek ezkerreko hemisferioa kaltetzen duen burmuineko odoljarioa jasatean gorputzeko ezkerraldean paralisia sufritzea.
Garuneko neuronen arteko komunikazioa neurotransmisore izeneko substantzia batzuen bitartez gauzatzen da. Neurotransmisore garrantzitsuenak azetilkolina, dopamina, noradrenalina eta serotonina dira. Substantzia hauen zeregina nerbio-bulkadaren transmisioan datza.
Neurotransmisoreak neurona baten axoian askatu eta hurrengo neuronaren dendritetan jasotzen dira.
Neurotransmisoreak eta hartzaileak
Neurotransmisoreak sinapsietan askatzen diren produktu kimikoak dira, bertako mintza despolarizatzen denean eta Ca2+ zelulan sartzen denean, normalean ekintza-potentzial bat sinapsiara iristen denean: neurotransmisoreak sintaxiaren xede-zelularen (edo zelulen) mintzaren gaineko molekula hartzaileei atxikitzen zaizkie, eta, horrela, hartzailearen molekulen propietate elektrikoak edo kimikoak aldatzen dituzte. Salbuespen gutxi batzuk izan ezik, garuneko neurona bakoitzak neurotransmisore kimiko bera edo neurotransmisoreen konbinazio bera askatzen du beste neuronekin egiten dituen lotura sinaptiko guztietan; Daleren printzipioa[15] izenarekin ezagutzen da arau hori. Horrela, neurona bat ezaugarri daiteke askatzen dituen neurotransmisoreengatik. Farmako psikoaktibo gehienek sistema neurotransmisore espezifikoak aldatzen dituzte. Hori cannabinoide, nikotina, heroina, kokaina, Alkohol, fluoxetina, klorpromazina eta beste droga askori aplikatzen zaie[16].
Garun ornodunean gehien aurkitzen diren bi neurotransmisoreak glutamatoa eta azido gamma-aminobutrikoa (GABA) dira, ia beti inhibitzaileak. Transmisore horiek erabiltzen dituzten neuronak garunaren ia alde guztietan aurki daitezke[17]. Nonahikotasuna dela eta, glutamatoaren edo GABAren eragina duten botikek efektu zabalak eta indartsuak izaten dituzte. Anestesia orokor batzuek glutamatoaren efektuak murriztuz jarduten dute; lasaigarri gehienek GABAren efektuak indartuz egiten dituzte lasaigarriak[18].
Beste dozenaka neurotransmisore kimiko daude garunaren eremu mugatuagoetan erabiltzen direnak, askotan funtzio jakin baterako erabiltzen direnak. Serotonina, adibidez —depresioaren aurkako botika askoren eta laguntza dietetiko askoren helburu nagusia—, rafe-nukleoak izeneko burmuineko eremu txiki batetik dator soilik[19]. Norepinefrina, kitzikapenean parte hartzen duena, locus coeruleus[20] izeneko hurbileko eremu txiki batetik dator soil-soilik. Beste neurotransmisore batzuek, hala nola azetilkolinak eta dopaminak, hainbat iturri dituzte garunean, baina ez daude glutamatoa eta GABA bezain nonahiko banatuak[21].
Jarduera elektrikoa
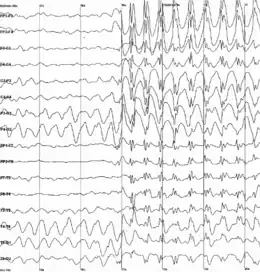
Seinaleztapenerako, neuronek erabiltzen dituzten prozesu elektrokimikoen albo-efektu gisa, burmuineko ehunak eremu elektrikoak sortzen ditu aktibo dagoenean. elektroentzefalografia (EEG)[22] edo magnetoentzefalografia (MEG) erabiliz, neuronen kopuru handiak jarduera sinkronizatua erakusten duenean sortzen dituzten eremu elektrikoak garezurretik kanpo detektatzeko bezain handiak izan daitezke. EEG grabazioek, arratoiak eta antzeko animalien garunetan ezarritako elektrodoekin egindako grabazioekin batera, animalia bizi baten garuna etengabe aktibo dagoela erakusten dute, baita lo dagoen bitartean ere[23]. Garunaren zati bakoitzak jarduera erritmikoaren eta ez-erritmikoaren arteko nahasketa bat erakusten du, jokabide-egoeraren arabera alda daitekeena. Ugaztunetan, garun-kortexak delta-uhin geldo handiak erakutsi ohi ditu loaldian, alfa uhin azkarragoak animalia esna baina arretarik jarri gabe dagoenean, eta kaotiko itxurako aktibitate irregularra animalia lanean ari denean, beta eta gamma uhinak deritzona. Konbultsio epileptiko batean, garunaren kontrol mekanismo inhibitzaileek ez dute funtzionatzen, eta aktibitate elektrikoa maila patologikoetara igotzen da EEG arrastoak sortuz eta garun osasuntsu batean ikusten ez diren uhin eta pika ereduak erakutsiz. Neurofisiologiaren egungo ikerketaren ardatz nagusia da populazio-mailako eredu horiek neurona indibidualen funtzio konputazionalekin erlazionatzea[23].
Metabolismoa
Ornodun guztiek odol- eta burmuin-hesi bat dute garunaren barruko metabolismoak gorputzeko beste atal batzuetan ez bezala funtziona dezan. Unitate neurobaskularrak garuneko odol-fluxua erregulatzen du aktibatutako neuronak energiaz hornitu ahal izateko. Zelula glialek funtzio garrantzitsua betetzen dute garunaren metabolismoan neuronak inguratzen dituen fluidoaren konposizio kimikoa kontrolatuz, ioien eta mantenugaien mailak barne[24].
Garuneko ehunak energia asko kontsumitzen du bere bolumenaren arabera, eta, beraz, garun handiek eskakizun metaboliko larriak ezartzen dizkiete animaliei. Adibidez, hegan egiteko gorputzaren pisua mugatu beharrak, dirudienez, hautespena ekarri du espezie batzuetan (saguzarretan adibidez) garunaren tamaina murrizteko[25]. Garunaren energia kontsumo gehiena neuronen karga elektrikoari eustera doa (mintz potentziala)[24]. Ornodun gehienek metabolismo basalaren % 2 eta % 8 artean eskaintzen diote garunari. Primateetan, ordea, ehunekoa askoz handiagoa da —gizakietan % 20-25era igotzen da[26]. Burmuinaren energia-kontsumoa ez da asko aldatzen denboran zehar, baina garun-kortexaren eskualde aktiboek eskualde inaktiboek baino energia gehiago kontsumitzen dute; hori da PET, fMRI[27] eta NIRSen[28] burmuineko irudi funtzionalen metodoen oinarria. Burmuinak, normalean, glukosaren oxigenoaren menpeko metabolismotik (hau da, odoleko azukretik) lortzen du energia gehiena[24], baina zetonek iturri alternatibo nagusi bat ematen dute, kate ertaineko gantz-azidoen (kaprilikoak eta heptanoikoak)[29][30], laktatoaren[31], azetatoaren[32] eta, beharbada, aminoazidoen ekarpenekin batera[33].
Baskularizazioa
Oxigenoa eta glukosa barneko arteria karotidaren eta orno-arterien bidez heltzen dira burmuinera. Bi arteria karotidak kanpoko eta barneko adarretan banatzen dira. Kanpoko adarra kanpoko karotida da, eta odola garezurraren kanpoaldera eramaten du, eta barneko adarrak (barneko karotidak), berriz, burmuinaren aurreko eta erdiko eremura eramaten du odola. Bi orno-arteriek burmuinaren atzeko aldea irrigatzen duen arteria basilarra osatzen dute. Garunaren oinaldean bada Willisen poligonoa deitzen den sistema bat. Bi sistemak lotzen ditu eta arteriaren bat ixten denean iskemia eragozteko balio du[8].
Ornodunen burmuina
Ugaztunen artean gizakiarena da burmuinik garatuena. Tolestura edo inguraketa ugari ditu, eta azpimarratzekoa da lobulu frontalaren garapena (bertan kokatzen dira adimenaren guneak).
Ugaztun eboluzionatuenengan burmuineko inguraketak oso ugariak dira. Eboluzioa aurrera joan ahala inguraketen bidez burmuinaren azalera zabaltzen da, gai grisaren kopurua handituz; burmuinaren bolumenak ez du aldatzen, baina neuronen kopurua nabarmen areagotzen da.
Ornodunen artean arrainen eta anfibioen burmuinak ez du kortexik (burmuin azalik), narrastiengan eta hegaztiengan oso txikia da, eta ugaztunengan batik bat garatzen da. Telentzefaloaren garapenak azaltzen du ugaztunen burmuinaren berezitasuna.
Erreferentziak
- «burmuin» Euskaltzaindiaren Hiztegia (Noiz kontsultatua: 2021-05-15).
- «burmuin - EH - Euskaltzaindiaren Hiztegia» www.euskaltzaindia.eus (Noiz kontsultatua: 2021-05-22).
- «cerveau | frantses-euskara hiztegi elektronikoa | Nola Erran» www.nolaerran.org (Noiz kontsultatua: 2021-05-22).
- «garun - EH - Euskaltzaindiaren Hiztegia» www.euskaltzaindia.eus (Noiz kontsultatua: 2021-05-22).
- «zerebro - EH - Euskaltzaindiaren Hiztegia» www.euskaltzaindia.eus (Noiz kontsultatua: 2021-05-22).
- Ibon Sarasola. (1998). «zerebro» Euskara batuaren ajeak. Alberdania, 176-177 or. ISBN 848866950X..
- «burumuin - EH - Euskaltzaindiaren Hiztegia» www.euskaltzaindia.eus (Noiz kontsultatua: 2021-05-22).
- «ZT Hiztegi Berria» zthiztegia.elhuyar.eus (Noiz kontsultatua: 2018-12-18).
- Purves, Dale.; Lichtman, Jeff W. (1985). Principles of neural development. Sunderland, Mass.: Sinauer Associates. ISBN 978-0-87893-744-8. OCLC 10798963
- (Ingelesez) Wong, Rachel O. L.. (1999-03). «RETINAL WAVES AND VISUAL SYSTEM DEVELOPMENT» Annual Review of Neuroscience 22 (1): 29–47. doi:. ISSN 0147-006X. (Noiz kontsultatua: 2023-08-17).
- (Ingelesez) Rakic, Pasko. (2002-02-01). «Adult Neurogenesis in Mammals: An Identity Crisis» Journal of Neuroscience 22 (3): 614–618. doi:. ISSN 0270-6474. PMID 11826088. PMC PMC6758501. (Noiz kontsultatua: 2023-08-17).
- (Ingelesez) Ridley, Matt. (2004-07-06). The Agile Gene: How Nature Turns on Nurture. Harper Collins ISBN 978-0-06-000679-2. (Noiz kontsultatua: 2023-08-17).
- (Ingelesez) Wiesel, Torsten N.. (1982-10). «Postnatal development of the visual cortex and the influence of environment» Nature 299 (5884): 583–591. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2023-08-17).
- (Ingelesez) van Praag, Henriette; Kempermann, Gerd; Gage, Fred H.. (2000-12). «Neural consequences of enviromental enrichment» Nature Reviews Neuroscience 1 (3): 191–198. doi:. ISSN 1471-0048. (Noiz kontsultatua: 2023-08-17).
- Kandel, Eric R.; Schwartz, James H. (James Harris); Jessell, Thomas M.. (2000). Principles of neural science. New York : McGraw-Hill, Health Professions Division ISBN 978-0-8385-7701-1. (Noiz kontsultatua: 2023-08-17).
- (Ingelesez) Pardridge, William M.. (2005-01-01). «The blood-brain barrier: Bottleneck in brain drug development» NeuroRX 2 (1): 3–14. doi:. ISSN 1545-5343. PMID 15717053. PMC PMC539316. (Noiz kontsultatua: 2023-08-17).
- McGeer, PL; McGeer, EG (1989). "Chapter 15, Amino acid neurotransmitters". In G. Siegel; et al. (eds.). Basic Neurochemistry. University of Michigan: Raven Press. pp. 311–332. ISBN 978-0-88167-343-2
- Foster, Alan C; Kemp, John A. (2006-02-01). «Glutamate- and GABA-based CNS therapeutics» Current Opinion in Pharmacology 6 (1): 7–17. doi:. ISSN 1471-4892. (Noiz kontsultatua: 2023-08-17).
- Frazer, A; Hensler, JG (1999). "Understanding the neuroanatomical organization of serotonergic cells in the brain provides insight into the functions of this neurotransmitter". In Siegel, GJ (ed.). Basic Neurochemistry (Sixth ed.). Lippincott Williams & Wilkins. ISBN 978-0-397-51820-3
- Mehler, Mark F.; Purpura, Dominick P.. (2009-03-01). «Autism, fever, epigenetics and the locus coeruleus» Brain Research Reviews 59 (2): 388–392. doi:. ISSN 0165-0173. PMID 19059284. PMC PMC2668953. (Noiz kontsultatua: 2023-08-17).
- Rang, H. P., ed. (2003). Pharmacology. (5. ed. argitaraldia) Churchill Livingstone ISBN 978-0-443-07145-4. (Noiz kontsultatua: 2023-08-17).
- Speckmann E, Elger CE (2004). "Introduction to the neurophysiological basis of the EEG and DC potentials". In Niedermeyer E, Lopes da Silva FH (eds.). Electroencephalography: Basic Principles, Clinical Applications, and Related Fields. Lippincott Williams & Wilkins. pp. 17–31. ISBN 978-0-7817-5126-1
- Buzsáki, Gyorgy (2006). Rhythms of the Brain. Oxford University Press. ISBN 9780199828234
- Nieuwenhuys, R; Donkelaar, HJ; Nicholson, C (1998). The Central Nervous System of Vertebrates, Volume 1. Springer. pp. 11–14. ISBN 978-3-540-56013-5
- (Ingelesez) Safi, Kamran; Seid, Marc A; Dechmann, Dina K.N. (2005-09-22). «Bigger is not always better: when brains get smaller» Biology Letters 1 (3): 283–286. doi:. ISSN 1744-9561. (Noiz kontsultatua: 2023-08-18).
- (Ingelesez) Mink, J. W.; Blumenschine, R. J.; Adams, D. B.. (1981-09-01). «Ratio of central nervous system to body metabolism in vertebrates: its constancy and functional basis» American Journal of Physiology-Regulatory, Integrative and Comparative Physiology 241 (3): R203–R212. doi:. ISSN 0363-6119. (Noiz kontsultatua: 2023-08-18).
- (Ingelesez) Raichle, Marcus E.; Gusnard, Debra A.. (2002-08-06). «Appraising the brain's energy budget» Proceedings of the National Academy of Sciences 99 (16): 10237–10239. doi:. ISSN 0027-8424. (Noiz kontsultatua: 2023-08-18).
- (Ingelesez) Mehagnoul-Schipper, D. Jannet; van der Kallen, Bas F.W.; Colier, Willy N.J.M.; van der Sluijs, Marco C.; van Erning, Leon J.Th.O.; Thijssen, Henk O.M.; Oeseburg, Berend; Hoefnagels, Willibrord H.L. et al.. (2002-05). «Simultaneous measurements of cerebral oxygenation changes during brain activation by near-infrared spectroscopy and functional magnetic resonance imaging in healthy young and elderly subjects» Human Brain Mapping 16 (1): 14–23. doi:. ISSN 1065-9471. (Noiz kontsultatua: 2023-08-18).
- (Ingelesez) Ebert, Douglas; Haller, Ronald G.; Walton, Marlei E.. (2003-07-02). «Energy Contribution of Octanoate to Intact Rat Brain Metabolism Measured by 13C Nuclear Magnetic Resonance Spectroscopy» Journal of Neuroscience 23 (13): 5928–5935. doi:. ISSN 0270-6474. PMID 12843297. (Noiz kontsultatua: 2023-08-18).
- (Ingelesez) Marin-Valencia, Isaac; Good, Levi B; Ma, Qian; Malloy, Craig R; Pascual, Juan M. (2013-02). «Heptanoate as a Neural Fuel: Energetic and Neurotransmitter Precursors in Normal and Glucose Transporter I-Deficient (G1D) Brain» Journal of Cerebral Blood Flow & Metabolism 33 (2): 175–182. doi:. ISSN 0271-678X. (Noiz kontsultatua: 2023-08-18).
- (Ingelesez) Boumezbeur, Fawzi; Petersen, Kitt F.; Cline, Gary W.; Mason, Graeme F.; Behar, Kevin L.; Shulman, Gerald I.; Rothman, Douglas L.. (2010-10-20). «The Contribution of Blood Lactate to Brain Energy Metabolism in Humans Measured by Dynamic 13C Nuclear Magnetic Resonance Spectroscopy» Journal of Neuroscience 30 (42): 13983–13991. doi:. ISSN 0270-6474. PMID 20962220. PMC PMC2996729. (Noiz kontsultatua: 2023-08-18).
- (Ingelesez) Deelchand, Dinesh K.; Shestov, Alexander A.; Koski, Dee M.; Uğurbil, Kâmil; Henry, Pierre-Gilles. (2009-05). «Acetate transport and utilization in the rat brain» Journal of Neurochemistry 109: 46–54. doi:. (Noiz kontsultatua: 2023-08-18).
- Soengas, José L.; Aldegunde, Manuel. (2002-03-01). «Energy metabolism of fish brain» Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology 131 (3): 271–296. doi:. ISSN 1096-4959. (Noiz kontsultatua: 2023-08-18).