Birus
Birus bat organismo baten zelula bizien barruan soilik erreplikatzen den agente infekzioso txiki bat da. Birusek era guztietako izaki bizidunak infektatu ditzakete, animalia eta landareetatik mikroorganismoetaraino, bakterioak eta arkeoak barne.[1]
- Oharra: Wikipediak ez du mediku aholkurik ematen. Tratamendua behar duzula uste baduzu, jo ezazu sendagilearengana.
 Ohar medikoa
Ohar medikoa
| Birus | |
|---|---|
| Sailkapen zientifikoa | |
| Domeinua | Virus [[|]] |
| Taldeak | |
I: kbDNA birusak II: kaDNA birusak | |
| Datu orokorrak | |
| Kolorea | baliorik ez |
| Gaixotasuna | gaixotasun biriko kutsakorra eta pneumonia birikoa |
1892an Dmitri Ivanovskik artikulu batean tabako landareak kutsatzen dituen bakterio ez den patogeno bat deskribatu zuenetik, eta 1898an Martinus Beijerinckek tabakoaren mosaikoaren birusa aurkitu zuenetik[2], 5.000 birus espezie inguru deskribatu dira xehetasunez[3], milioika mota dauden arren[4]. Birusak Lurreko ia ekosistema guztietan daude, eta entitate biologiko mota ugarienak dira[5][6]. Birusaren azterketa birologia izeneko zientziak egiten du, mikrobiologiaren azpiespezialitate bat.
Infektatutako zelula baten barruan edo zelula bat infektatzeko prozesuan ez badaude, birusak partikula independente edo birioi moduan existitzen dira. Birioi batek honako atalak ditu: i) material genetikoa, DNA ala RNA molekula luzeak (bata ala bestea, inoiz ez biak), birusak eragiten dituen proteinen egitura kodetzen dutenak; ii) geruza proteiniko bat, kapsidea, materiala inguratzen eta babesten duen; eta, kasu batzuetan, iii) lipidozko kanpoaldeko bilgarri bat. Birus partikula horien formak helikoidal eta ikosaedrikoak dira espezie batzuentzat, eta egitura konplexuagoak beste batzuentzat. Birus espezie gehienek birioi txikiegiak dituzte mikroskopio optiko batekin ikusteko, bakterio gehienen tamainaren ehunena gutxi gorabehera.
Birusen jatorriak biziaren historia ebolutiboan ez daude argi: batzuk plasmidoetatik abiatuta eboluzionatu zutela uste da -zelulen artean mugi daitezkeen DNA piezak-; beste batzuk, berriz, bakterioetatik abiatuta eboluziona zezaketen. Eboluzioan, birusak geneen transferentzia horizontalerako bitarteko garrantzitsu bat dira, eta aniztasun genetikoa ugalketa sexualaren antzera handitzen dute[7]; baliteke ere eboluzioaren muinean paper garrantzitsua edukitzea[8]. Birusak izaki biziduntzat hartzen dituzte zientzialari batzuek, material genetikoa garraiatzen dutelako, ugaldu eta eboluzionatu egiten direlako hautespen naturalaren bidez. Hala ere, ez dute ezaugarri giltzarririk (zelulen egitura, adibidez), normalean bizitza gisa hartzeko beharrezkotzat jotzen direnak. Eta ez dute metabolismo propiorik ere. Ezaugarri horietako batzuk dituztelako, baina ez guztiak, birus horiek "bizitzaren ertzeko organismo"[9] eta erreplikatzaile gisa deskribatu dira[10]. Biziaren mugan dauden egitura sinpleak dira beste zientzialari batzuen ustez [11]
Birusak modu askotan hedatzen dira. Transmisio bide bat bektore bezala ezagutzen diren gaixotasunak daramatzaten organismoen bidez da: adibidez, birusa landaretik landarera transmititzen dute landareen izerdiaz elikatzen diren intsektuek, adibidez Aphidoidea taldekoak; eta animaliei erasotzen dieten birusak, berriz, odola xurgatzen duten intsektuek garraiatu ditzakete. Gripearen birusak eztul eta doministiku egitean hedatzen dira. Norobirusak eta errotabirusak, gastroenteritis biralaren kausa arruntak, bide fekal-oraletik hartzen dira, ukipenaren bidez eta gorputzera sartzen dira janarian edo uretan. GIBa sexu kontaktuagatik eta infektatutako odolaren eraginpean egoteagatik transmititzen den hainbat birusetako bat da. Birus batek kutsa ditzakeen zelula gonbidatuen barietatea "ostalari" deitzen da. Estua izan daiteke, birusa bat gai bada espezie gutxi kutsatzeko, edo zabala, gai bada asko infektatzeko[12].
Animaliengan birus infekzioek immunitate erantzun bat eragiten dute, infekzioa sortzen duen birusa ezabatu ohi duena. Erantzun immunitarioak txertoen bidez ere gerta daitezke, infekzio biral espezifikoari artifizialki hartutako immunitatea ematen diotenak. Birus batzuek, hiesa eta hepatitis birala eragiten dutenak barne, erantzun immunitario horiek saihestu eta infekzio kronikoak eragiten dituzte. Birusen aurkako hainbat sendagai garatu dira, antibiral izenekoak.
Historia

Louis Pasteurrek ezin izan zuen amorrua eragin zuen agenterik aurkitu, eta mikroskopioek antzemateko txikiegia zen patogeno baten inguruan espekulatu zuen[13]. 1884an Charles Chamberland mikrobiologo frantziarrak Chamberland iragazkia (edo Pasteur-Chamberland iragazkia) asmatu zuen, bakterio guztiak bertatik igarotzen zen soluzio batetik ezabatzeko bezain txikiak ziren poroekin[14]. 1892an, Dimitri Ivanovski biologo errusiarrak iragazki hau erabili zuen orain tabakoaren mosaikoaren birusa bezala ezagutzen dena aztertzeko: kutsatutako tabako landareen hosto birrinduen estraktuak infekziosoak izaten jarraitzen zuten, baita bakterioak desagerrarazteko filtrazioa egin ondoren ere. Ivanovskik iradoki zuen bakterioek eragindako toxina batek eragin zezakeela infekzioa, baina ez zuen ideiarekin jarraitu[15]. Une horretan pentsatzen zen agente infekzioso guztiak iragazkiek atxiki eta inguru nutritibo batean landu zitezkeela, eta hori gaitzen teoria mikrobiarraren parte zen[2]. 1898an Martinus Beijerinck herbeheretar mikrobiologoak esperimentuak errepikatu zituen eta iragazitako soluzioak agente infekzioso forma berri bat zuela konbentzitu zen[16]. Agentea zatitzen ari ziren zeluletan soilik biderkatzen zela ikusi zuen, baina bere esperimentuek partikulez egina zegoela frogatu ez zutenez, contagium vivum fluidum (ernamuin bizi disolbagarria) deitu zion, eta birus hitza berriz sartu zuen terminologia zientifikoan. Beijerinckek birusak likidoak zirela esan zuen, gerora Wendell Stanleyk ezeztatutako teoria bat, partikulak zirela frogatu zuenean. Urte horretan bertan, Friedrich Loeffler eta Paul Froschek antzeko iragazki baten bidez pasatu zuten lehen animalia birusa: aphthobirusa, sukar aftosoaren agentea[17].
XX. mendearen hasieran, Frederick Twort bakteriologo ingelesak bakterioak kutsatzen dituzten birus talde bat aurkitu zuen, orain bakteriofagoak (edo normalean fagoak) deituak[18], eta Felix d'Herelle mikrobiologo franko-kanadarrak, berriz, agar plaka batean gehitzerakoan bakterioak hiltzen zituzten birusak deskribatu zituen. Birus horiek zehaztasunez diluitu zuen, eta diluzio altuenek (birus kontzentrazio baxuenak), bakterio guztiak hil beharrean, organismo hilen eremu diskretuak osatzen zituztela aurkitu zuen. Eremu horiek kontatzeak eta diluzio-faktorearekin biderkatzeak jatorrizko esekiduran zegoen birus kopurua kalkulatzea ahalbidetu zion[19]. Fagoak sukar tifoidea eta kolera bezalako gaixotasunetarako tratamendu potentzial gisa iragarri ziren, baina haien promesa penizilinaren garapenarekin ahaztu egin zen. Bakterioen antibiotikoekiko erresistentziaren garapenak bakteriofagoak terapeutikoki erabiltzeko interesa berritu du[20].
XIX. mendearen amaieran, birusak honela definitzen ziren: infektagarritasuna, iragazkiak pasatzeko gaitasuna eta ostalari bizien beharra. Birusak landare eta animalietan baino ez ziren isolatu. 1906an, Ross Granville Harrisonek linfan ehunak hazteko metodo bat asmatu zuen, eta, 1913an, E. Steinhardt, C. Israeli eta R. A. Lambertek Vaccinia birusa hazteko akurien ehun kornealaren zatietan erabili zuten metodo hori[21]. 1928an, H. B. Maitlandek eta M. C. Maitlandek Vaccinia birusa garatu zuten, oiloen giltzurrun xehatuen esekidura bidez. Bere metodoa ez zen oso zabaldua izan 1950eko hamarkadara arte, poliobirusa eskala handian hazi zenean txertoak ekoizteko.
Ernst Ruska eta Max Knoll ingeniari alemaniarrek mikroskopio elektronikoa asmatu bezain laster, 1931ean, lehen birusen irudiak lortu ziren [22]. 1935ean, Wendell Stanley biokimikari eta birologo estatubatuarrak tabakoaren mosaikoaren birusa aztertu zuen eta nagusiki proteinez osatua zegoela aurkitu zuen. Tabakoaren mosaikoaren birusa izan zen kristalizatu zen lehena, eta, beraz, haren egitura xehetasunez ezagutu ahal izan zen. Birus kristalizatuaren X izpien difrakzioagatiko lehen irudiak Bernalek eta Fankuchenek lortu zituzten 1941ean. Bere irudietan oinarrituz, Rosalind Franklinek birusaren egitura osoa aurkitu zuen 1955 [23]. Urte berean, Heinz Fraenkel-Conratek eta Robley C. Williamsek frogatu zuten tabakoaren mosaikoaren birusaren ARN purifikatua eta bere proteinak beren kabuz mihiztatu daitezkeela, eta birus funtzionalak sortu, eta zelula ostalarietan birusak sortzeko modua hori bera izan zitekeela iradoki zuten.
XX. mendearen bigarren erdia birusen aurkikuntzaren urrezko aroa izan zen; animalien, landareen eta bakterioen 2.000 birus espezie inguru urte hauetan aurkitu ziren [24]. 1963an Baruch Blumbergek B hepatitisaren birusa aurkitu zuen, eta 1965ean Howard Teminek lehen erretrobirusa deskribatu zuen. Alderantzizko transkriptasa, erretrobirusek euren RNA DNA bihurtzeko erabiltzen duten entzima garrantzitsua, 1970ean deskribatu zuten, modu independentean, Howard Teminek eta David Baltimorek [25]. 1983an, Frantziako Pasteur Institutuko Luc Montagnierren taldeak lehen aldiz isolatu zuen VIH.41 izenarekin ezagutzen den erretrobirusa, HIESaren eragilea [26]. Michael Houghtonek eta Chiron Corporationeko bere taldeak C hepatitisaren birusa aurkitu zuten, 1989an.
Bestalde, biologia molekularraren garapenak eta PCR teknikaren asmakizunak, XX. mendearen 80ko hamarkadan, birusen genomaren ikerketa ahalbidetu dute, laguntza handikoa izan dena txerto berrien bilaketan eta botika antibiralen garapenean. XX. mendearen bukaerak eta XXI mendearen hasierak birulentzia handiko birus berrien agerpena ikusi dute (GIB, Ebolako birusa, SARS-ekoa edo COVID-19koa -azken biak SARS CoV eta SARS-CoV-2 birusek, hurrenez hurren sortuak-). Patogeno berri hauek izurrite latzak eragin dituzte eta birologoen arreta erakarri dute azken urteotan, bere osasun ondorio latzak direla-eta.
Ezaugarriak
Birusek eztute organulurik eta ezta autonomia metabolikorik ere, beraz, eztira zelulatzat hartzen. Beren bizi zikloak bi fase ditu: Bata extrazelularra (zelulatik kanpokoa) eta metabolikoki inertea, eta bestea intrazelularra, ugalketa ematen denekoa.
Historikoki, agente infekzioso bat birus gisa hartzen zen beste aukerak baztertzerakoan, hau da, agente ezezaguna bakterio ez bazen, birus bat izan behar zuen. Horren emaitza izan zen birusen taldean oker sartu zirela zelula bizietan hazten ziren eta oso txikiak ziren agente jakin batzuk (adibidez, mikoplasmak).
Birusen tamaina txikiak bakterioetatik bereizten ditu hiru alderditan:
- Iragazki bakterianoek ez dituzte birusak atxikitzen
- Birusak ezin dira mikroskopio optikoan ikusi
- Ez dute jalkitzen zentrifugatzaile arruntetan; jalkitzeko, zentrifugazio-abiadura handiko zentrifugagailuak erabili behar dira.
Horrez gain, birusek beste ezaugarri bereizgarri batzuk dituzte:
- Azido nukleiko mota bakarra dute (DNA ala RNA, inoiz ez biak)
- Hazteko edo erdibiditzeko ezintasuna
- Entzima-konplexurik ez izatea
- Aurrekoaren ondorioz, zelula barneko parasito hertsiak izatea.
Aipatutako lau ezaugarrietatik hiru lehenak bakarrak dira unibertso biologikoan. Ezaugarri hauek birusak bizia eta bizirik ezaren arteko mugan ezartzen dituzte.
Tamaina
Birusen tamaina 24 nanometrotik hasi (ahoeriaren birusa) eta 300 nanometroetarainokoa izan daiteke (poxbirusena).
Hain izaki txikiak izateagatik, beren aurkikuntza oso berandu gertatu zen. Birusen existentziaz dagoen lehen erreferentzia Dimitri Ivanovski errusiar botanikoak eginikoa da. Ikertzaile hau, tabakoaren mosaiko izeneko gaixotasun eragilearen bila zebilen eta zera ondorioztatu zuen, bakterioak baino toxina edo organismo txikiago bat izango zela eragile hori, bakterioak atxiki zitzaketen iragazkiak iragan baitzitzazkeen. Eragile patogeno hauei, birus iragazkor deitu zien.
1897an Martinus Beijerinck holandar mikrobiologoak Ivanovskik eginiko saiakeren antzekoak egin zituen, eta toxinen ideia alboratu zuen, gaixotasunaren eragileak ugaltzeko ahalmena zuela ikusi baitzuen, landare batetik bestera bere ahalmen infekziosoa mantentzen zuelako, bere ahalmen patogenoa murriztu gabe. Handik gutxira Frederick Loeffer eta Paul Frosch mikrobiologo alemaniarrek, ganaduaren ahoeria birus iragazkor batek sortzen zuela ikusi zuten.
30eko hamarkadan, poro txikiagoko iragazkiak erabiliz, birus kantitate handiak lortzea ahalbideratzen duen in vitro hazkuntza zelular teknikekin, ultrazentrifugazioarekin eta azkenik mikroskopio elektronikoa eta X izpien difrakzioaz, birusak ikustea lortu zen.
Kristalizagarritasuna
1935an W. Stanleyk frogatu bezala, birusak kristalizagarriak dira. Partikula birikoek forma geometriko doiak dituzte eta beren artean berdin-berdinak dira, beste organulu, organismo eta zelulak ez bezala. Bolumen eta forma berdinekoak izanik, partikula birikoek pauta tridimentsional erregularretan ordenatzeko joera dute, kristalizatzekoa alegia.
Nahitaezko parasito intrazelularrak
Zelulen derrigorrezko parasitoak dira eta ostalaritik kanpo inerteak. Halere, birus batzuek 40 bat egunetan iraun dezakete ostalariren batean ugaldu gabe. Birus barneko azido nukleikoa erreplikatu eta birus berriak sor daitezen, ezinbestekoa da zelula bizi bat inbaditu (ostalaria), eta bere egitura eta entzimak erabiltzea, horrela, birusak ondorengoak izan ditzan. Ondorioz, birus asko eta asko patogenoak dira, bakterio, landare eta animalietan gaixotasunak eragiten dituztelako.
Egitura
Birus partikula oso batek, birioi izena hartzen du. Azido nukleiko bat dauka (DNA ala RNA, inoiz ez biak), kapside izeneko proteinazko babes geruza batez inguratuta. Batzuek, lipidoz osatutako bilgarria izan dezakete kapsidearen gainetik, ostalariaren zelula mintzetik eratorritakoa. Hau oso ohikoa da animaliak infektatzen dituzten birusetan [27]. Bilgarri lipidikoa duten birusak erraz inaktibatzen dira xaboiarekin edo alkoholarekin, baina bilgarririk gabeko birusak (birus biluziak) erresistenteagoak dira, orokorrean [28].
Kapsidea birusaren genomak kodetutako proteinez egiten da eta bere formak, birusak morfologikoki eta antigenikoki bereizteko balio du. Kapsomero izeneko azpiunitatez osatuta dago kapsidea. Hauek proteina globularrak izaten dira eta batzuetan zati gluzidiko bat izaten dute lotuta. Kapsomeroak beren artean elkartzen dira, kapsideari forma geometriko bat emanez.
Birus konplexuek proteina berezi batzuek sor ditzakete, kapsidea osatzen lagun dezaketenak.
Orokorrean birusen artean hiru mota morfologiko daude kapsidearen itxuraren arabera:
| Birus helikoidalak | |
 Kapside helikoidalaren diagrama |
Kapside helikoidala subunitate mota bakar batez osatua dago. Subunitateak ardatz zentral batera lotzen dira barnean hutsunea duen egitura helikoidala osatuaz. Honela, harizpi itxurako birioiak osatzen dira zeinak motz eta zurrun edo luze eta oso malgu izan daitezkeen. Material genetikoa, gehienetan kate bakarreko RNA, baina batzuetan ssDNA, proteinazko helizeari lotuta dago, karga negatibodun azido nukleikoa eta karga positibodun proteinen arteko interakzioen ondorioz. Orohar, kapside helikoidalaren luzera, barnean daraman azido nukleikoaren luzeraren araberakoa da eta diametroa berriz protomeroen tamaina eta antolamenduaren araberakoa. Asko ikertutako tabakoaren mosaikoaren birusa da birus helikoidal baten adibidea. |
| Birus ikosaedrikoak | |
 Adenobirusen diagrama |
Kapside ikosaedrikoek esfera itxura dute behar baino handipen txikiagoaz begiratuz gero, baina berez ikosaedro baten itxura hartzen dute. Mota ezberdinetako kapsomeroek lotura ez kobalente bidez ikosaedro erregular bat osatzen dute, 20 aurpegi triangular eta 12 ertzeko poliedroa alegia. Erdialdean hutsune bat geratzen da non azido nukleikoa kokatzen den, bildu bildu eginda. Batzuek aurpegi gehiagoko poliedroak osatzen dituzte eta batzuek kapsidetik kanpokaldera irteten diren zuntz proteikoak ager ditzakete. Birus ikosaedrikoen adibide moduan ditugu pikornabirusak (hotzeri eta faringitien sortarazleak) eta baita adenobirusak ere . |
| Birus konplexuak | |
 Bakteriofago baten diagrama |
Aldaera txiki batzuk ager daitezke, baina orokorrean beren egiturak zati hauek agertzen ditu:
Azido nukleikoa daraman buru bat, ikosaedro itxurakoa. Egitura helikoidala duen isatsa, zilindro huts formakoa. Kapsomeroz osatutako lepoko bat burua eta isatsaren artean. Oinaldeko plaka bat isatsaren azken muturrean, birusa ostalariaren mintz zelularrari lotzen laguntzeko ainguratze puntu batzuekin. Plaka honetatik, proteinazko zuntz batzuk ere irteten dira, finkapen hori gauzatzen laguntzen dutenak. Birus bakteriofago gehienek itxura konplexu hau agertzen dute. |
| Poxbirusak morfologia ezohikoa duten birus handi eta konplexuak dira. Beren genoma proteinekin elkartuta dago nukleoide izeneko egituraren barnean. Nukleoideak disko itxura du eta mintz batez inguratuta dago, eta alboetan funtzio ezezaguneko bi gorputz ditu. Birusak kanpoaldean proteinazko geruza lodi bat dauka. | |
Birusen genoma

Azido nukleiko mota bakarra (DNA ala RNA) izateak birusak bereizten ditu gainontzeko bizidunetatik; izan ere, eukarioto eta prokarioto guztiek DNA eta RNA, biak, dituzte.
Birusen azido nukleikoa askotarikoa izan daiteke: [29]
- kate bikoitzeko DNA (DNA duten birus gehienetan),
- kate bakarreko DNA (DNA duten birus gutxi batzuetan),
- kate bakarreko RNA (RNA duten birus gehienetan),
- kate bikoitzeko RNA (RNA duten birus gutxi batzuetan).
Salbuespenak badaude ere, orokorrean landareak infektatzen dituzten birusek RNA izan ohi dute, animaliak infektatzen dituztenek DNA ala RNA, eta bakterioak infektatzen dituztenek (bakteriofagoek), DNA.
Horretaz gain, azido nukleiko horietako batzuk zirkularrak dira, eta beste batzuk linealak.
Kate bakarreko RNA duten birus batzuek (erretrobirusek) funtsezko entzima bat dute, RNAtik abiatuta DNA sortzen duena: alderantzizko transkriptasa. Gero, birus hauek RNA-m sortzen dute transkripzioaren bidez, eta hortik abiatuta behar dituzten proteinak
- RNA → DNA
RNA duten birusetan RNA hori polaritate positibokoa (+) ala negatibokoa (-) izan daiteke, RNA-m-arekiko duen osagarritasunaren arabera. RNA (+) baldin bada, zelula ostalarian zuzenean proteinak sortuko ditu. RNA (-) baldin bada, RNA-m bihurtu behar da lehen, RNA polimerasa baten bitartez. Alderantzizko transkriptasa duten RNA birusek, aldiz, goian aipatu den prozesua jarraitzen dute.
Birusen genomaren tamaina aldakorra da, baina oso txikia bakterioen edo zelula eukariotoen genomaren aldean. Birus txikienek, esaterako, 5 gene edo gutxiago izaten dituzte [30]. Giza genomak, aldiz, 30.000 gene inguru ditu. ΦX174 bakteriofagoak 5.386 base ditu (kate bakarreko DNA dauka). Epstein-Barr birusak (kate bikoitzeko DNA duenak) 172.000 pare-base ditu, eta 85 gene. GIBak, HIESaren eragileak, 9 gene besterik ez dauzka [31].
Bestalde, RNA duten birusek mutazioak jasaten dituzte sarri, RNAren kopia-prozesuan akatsak ohikoak direlako [32]. RNA duten beste birus batzuek (esaterako, gripearenak) genoma oso zatituta dute kapsidearen barruan, genoma horrek ez baitu zati bakarra osatzen. Genomaren egitura berezi horrek aldakortasun genetikoa areagotzen du. Hori dela eta, egitura genetiko desberdinetako birusak agertzen dira etengabe, RNA-ren zatien arteko birkonbinazio genetikoa gertatzen delako. Mutazio edo birkonbinazio genetikoaren ondorioz birusek ezaugarri bereziak berenganatu ditzakete, batzuetan oso onuragarriak haientzat (birulentzia gehiago garatu, adibidez).
Birkonbinazio genetikoa gerta daiteke, halaber, genetikoki gertu dauden birusen bi anduik zelula ostalari bera infektatzen dutenean. DNA zatiak apurtu egin daitezke eta jatorrizko bi DNAren artean truke bat gertatu, konbinazio genetiko berri bat sortuz. Batzuetan, sortutako DNA horrek birus oso birulento berriak sor ditzake [33] [34].
Erreplikazioa
Birusek ostalaria infektatu eta gaixotasuna hedatu ahal izateko ugalketa ezinbestekoa dute. Horretarako ostalariaren zelula erabiltzen dute, beren kasa erreplikatzeko gaitasunik ez baitute. Birusen errreplikazio zikloa asko aldatzen da birus mota batetik bestera, baina funtsean sei oinarrizko urratsez osatzen da.
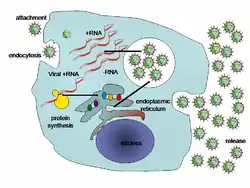
- Finkapena. Birusak eta ostalariak "talka" egiten dute eta ezagutza ematen da. Finkapena ondo burutzeko, birusak errezeptore espezifikoak behar ditu zelula-hartzailearen mintzean. Hain zuzen, birusek gorputzeko zelula espezifikoak infektatzen dituzte, hartzaile espezifikoak dituzten gorputzeko zelulak, alegia [35] (HIESaren birusak T linfozitoak, adibidez, A hepatitisaren birusak gibeleko zelulak, eta gripearen birusak arnas-aparatuko zelulak. SARS-CoV-2 birusak, COVID-19aren eragileak, ACE2 hartzaile zelularrari atxikitzen zaio, gorputzeko hainbat ehunetan dagoena; horregatik birus honek gorputzeko atal ezberdinak kaltetu ditzake). Zelula edo ehun espezifiko batekiko birus batek duen afinitateari tropismo biriko deritzo. Birusaren kapsideko proteina eta ostalariaren zelulako hartzaile espezifikoen arteko lotura itzulezina da.
- Barneraketa. Birus espeziearen arabera barneraketa desberdina izan daitekeen arren, orokorrean endozitosiaren bidez sartzen dira animalia-birusak infektatutako zeluletan. Bakteriofagoen kasuan genomaren injekzioa ematen da, eta zelula barrura ez da birus osoa sartzen, bere material genetikoa soilik baizik.
- Biluztea. Birusaren azido nukleikoa ostalariaren zelulan askatzen den momentua da, horretarako zelularen entzimek kapsidea degradatzen dute.
- Erreplikazioa. Birus berrien eraketarako beharrezkoak diren osagaien sintesia da, hots, RNAm eta proteinena. Fase horretan birusaren material genetikoak (DNAk edo RNAk) zelularen egiturak zuzentzen ditu birusaren material genetiko berria eta birusaren proteinak sintetizatzeko.
- Heltzea. Kapsidearen proteinak eta birusaren material genetiko berria (DNA ala RNA) sortu direnean, elementu horiek akoplatzen dira birus berriak sortzeko.
- Askapena. Birusak zelulatik lisi ala gemazioren bidez askatzen dira. Estalkidun birusek gemazioa erabiltzen duten bitartean, estalki gabekoak zelulan pilatzen joaten dira denbora batez eta ondoren, zelula lisiz puskatzen dute. Bigarren birus hauek zelula puskatzen duten bitartean, lehenengoek ez dute zelula apurtzen askatzen direnean (zelula geroago hiltzen da, birusek sortutako aldaketen ondorioz)
Birusen ziklo litikoa eta lisogenikoa

Goian azaldu den prozesua da birus gehienek jarraitzen dutena, bakterio, landare edo animalia baten zelulak infektatzen dituztenean. Prozesu horretan zelula barruan sortutako birus berri horietako batzuek zelula puskatzen dute, hots, zelularen lisia eragiten dute, azaldu dugun bezala. Ziklo litikoa, horrenbestez, burutzen dute birus horiek [36].
Beste birus batzuek, aldiz, ez dute portaera hori erakusten zelula infektatzen dutenean. Birus hauek denbora luzean iraun dezakeen harreman berezi bat eratzen dute zelula ostalariarekin, zelula ez baitute hasiera batean puskatzen: fenomeno horri lisogenia deritzo, eta bakteriofagoek zein animalia-birusek burutu dezakete.
Lisogenian birusaren material genetikoa zelula ostalariaren genoman txertatzen da (giza zeluletan eukariotoen kasuan, eta bakterioetan prokariotoen kasuan). Hots, goiko ataleko erreplikazio fasea ez du burutzen birusak: material genetiko berria eta proteina birikoak sortu beharrean, material genetiko hori zelularen DNAn integratzen da eta hor egon daiteke hainbat belaunalditan, sor edo latentzia egoeran. Luzaroan egon daiteke hor, harik eta une jakin batean "esnatu" arte.
Zelula ostalariaren DNAn integratuta dauden birusei profago deitzen zaie (bakteriofagoen kasuan) edo probirus (animalia-birusen kasuan) [37]. Infektatutako zelularen genoma zatitzen den bakoitzean profagoa edo probirusa ere zatitzen da harekin batera.
Zelula eukariotoen DNAn integratuta dauden probirus horietako batzuek beste ezaugarri bat dute: tumoreak sortzen dituzte. Probirusak zelula normalaren eraldaketa edo transformazioa eragiten du, zelula kantzerigeno bihurtuz. Probirusaren material genetikoak onkogeneak ditu, eraldaketa hori sortzen dutenak. Fenomeno horri transformazio birikoa deritzo, eta hainbat tumoreen atzean dago (T zelulen leuzemia edo umetoki-lepoko minbiziaren atzean, esaterako).
Profagoak eta probirusak denbora luzean egon daitezke zelula ostalariaren barnean, belaunaldiz belaunaldi, sor egoeran. Hori gertatzen da, adibidez, HIESaren birusarekin. Probirusak edo profagoak egoera horretan daude harik eta kanpoko eragile batek (argi ultramoreak edo zenbait konposatu kimikok, profagoaren kasuan) berpizten dituzten arte. Hori gertatutakoan profagoak edo probirusak birus berriak sortzeari ekiten dio eta ziklo litikoa hasten da, bakterioaren edo zelula eukariotoaren suntsiketarekin bukatuko dena [38].
Birusek, azkenik, beste harreman mota bat eratu dezakete infektatutako zelulekin: infekzio latentea. Birusak sor egoeran egoten dira kutsatutako zelulen barruan, partikula biriko berririk sortu gabe; ondorioz, infektatutako zelulak ez dira suntsitzen. Hala ere, une jakin batean zenbait estimuluk birusaren aktibazioa eragiten dute, eta orduan birusa erreplikatzen da eta kutsatutako zelula suntsitu egiten du. Lisogenian ez bezala, infekzio latentean genoma birikoa ez da txertatzen kutsatutako zelularen DNAn.
Portaera mota hori barizela-zoster birusak jarraitzen du. Barizela pasatu ondoren, birusa urtetan zehar infektatutako neuronetan mantentzen da, zenbait estimuluk aktibatu egiten duten arte; hori gertatzerakoan, herpes zoster mingarria eragiten du. [39].
Gaixotasun biriko batzuk
Beheko taulan birusek gizakian eragindako gaixotasun garrantzitsuenak daude:
| Gaitza | Birus eragilea | Sintoma nagusiak | Tratamendua edo prebentzioa |
|---|---|---|---|
| A hepatitisa | Pikornabirus | Ikterizia, sukarra eta astenia | Atsedena; txertoa badago |
| Amorrua | Errabdobirus | Sintoma neurologikoak | Immunoglobulinak; txertoa badago |
| B hepatitisa | Hepadnaviridiae | Ikterizia, sukarra eta ondoeza | Interferoia; txertoa badago |
| Barizela | Barizela-zoster birusa | Sukarra, nekea, besikula gorriak larruazalean | Txertoa badago |
| Baztanga | Poxbirus | Lurretik desagertutako gaixotasun infekziosoa | Txertoa badago |
| C hepatitisa | Flaviviridae | Sintomarik gabeko gaitza ala nekea eta ondoeza | Sofosbubir |
| Covid-19 | SARS-CoV-2 birusa | Sukarra, eztula, nekea eta ondoeza; arnasteko arazoak | Txertoa badago |
| Ebola sukar hemorragikoa | Ebola birusa | Sukar handia, nekea, odoljarioak, beherakoak | Tratamendurik eta txertorik gabe |
| Elgorria | Paramixobirusa | Sukarra, eztula, orban gorri txikiak larruazalean | Atsedena; txertoa badago |
| Errubeola | Togabirus | Sintomarik gabeko gaitza ala sukarra, eztula eta orban arrosa txikiak larruazalean | Txertoa badago |
| Gripea | Influenzabirus | Sukarra, nekea, giharretako mina | Atsedena; txertoa badago |
| Hazizurria | Paramixobirus | Buruko mina, sukar arina, listu-guruinen hantura | Ez dago tratamendu espezifikorik; txertoa badago |
| Herpes | Herpes sinplex birusa | Erremina, besikulak eta orban gorriak | Antibiralak |
| Herpes zosterra | Barizela-zoster birusa | Erremina eta larruazaleko besikula txikiak | Kasu larrietan, antibiralak |
| HIESa | GIBa | Infekzio errepikakorrak | Antibiralak |
| Mononukleosi infekziosoa | Epstein-Barr birusa | Sukarra, faringitis, nekea, gongoil linfatikoen hantura | Tratamendurik eta txertorik gabe |
| Poliomielitisa | Poliobirus | Sintomarik gabeko gaitza edo sukarra, ondoeza, goragaleak eta batzuetan kalte neurologikoak | Tratamendurik gabe; txertoa badago |
| Sukar horia | Flabibirusa | Gripearen antzeko sintomak; kasu larrietan gibeleko eta giltzurrunetako kalteak | Txertoa badago |
Birusen aurkako borroka
Birusak zelula barneko parasito hertsiak dira. Horrek esan nahi du zelulak infektatu behar dituztela ezinbestez "bizi" ahal izateko, jarduera biologikoa izateko. Zelulatik kanpo daudenean, birusak partikula inerteak dira, ez dute jarduera biologikorik erakusten.
Beste mikrobioen aldean (bakterioak, protozooak, legamiak...), modu autonomoan ere bizi daitezkeenak -infektatu gabe-, birusek ezin dute infektatu gabe jardun, parasito hertsiak direlako. Horrenbestez, birusak bakterio edo legamiak baino askoz kaltegarriagoak dira, zelulak beti infektatu behar dituztelako, sarritan gaixotasunak sortuz gizakiari, animaliei edo landarei [40].
Birus patogenoen aurkako borrokan hainbat elementu daude tartean: alde batetik, gure organismoaren defentsa naturalak; beste aldetik, gizakiak sortutako eragile kimikoak (antibiralak).
Immunitate-sistemaren defentsak birusen aurka

Birus patogeno bat gure organismoan sartzen denean gure immunitate-sistemak elementu asko abian jartzen ditu mikrobio arrotza suntsitzeko. Lehenengo defentsa-lerroa mekanismo ez-espezifikoak dira (berdin jarduten dutenak edozein mikrobioren aurka): esaterako, sudurretik sartzen diren birusen aurka (arnas-aparatuko infekzioak sortzen dituztenak: gripea edo pneumonia, adibidez) sudurreko eta errinofaringeko mukiak eta bertako zelula ziliodunek jarduten dute, birusak harrapatu eta erauzten dituztenak [41].
Bigarren defentsa mekanismo ez-espezifikoak ehunetako eta odoleko fagozitoak dira. Fagozitoak immunitate-sistemaren zelula bereziak dira, mikrobioak irentsi eta suntsitzen dituztenak. Fagozito garrantzitsuenak makrofagoak, neutrofiloak eta zelula dendritikoak dira. Fagozitoen jarduera ez da selektiboa, partikula arrotzak ez baitituzte bereizten eraso egiteko orduan [42]. Makrofagoak eta zelula dendritikoak, bereziki, eraginkorrak dira birusen aurka. Immunitate-sistemaren beste zelula batzuk, NK zelulak, birusen suntsiketa prozesuan parte hartzen dute (bereziki, birusak dituzten eta infekzio kronikoak pairatzen dituzten zelulak suntsitzen dituzte NK zelulek) [43].
Interferoia da gure organismoak birusen aurka erabiltzen duen beste estrategia bat. Funtsean, interferoia birusek infektatutako zelulek eta immunitate-sistemaren zelulek ekoizten duten proteina da, ondoko zelulak babesten dituena infekzio birikoen aurrean [44]. Molekula oso garrantzitsua da interferoia birusen aurkako infekzioetan. Birus batzuek interferoiaren sintesia beste batzuek baino gehiago areagotzen dute, oso ondo ezagutzen ez den mekanismo baten bidez.
Organismoaren bigarren defentsa-lerroa mekanismo espezifikoak dira: erantzun immunitarioa. Birusek lehenengo defentsa-lerroa gainditzen badute, defentsa-sistema konplexu eta eraginkor honekin topo egingo dute. Immunitate-sistemaren defentsa mekanismo espezifikoek osatzen dute defentsa-lerro hori, eta hauen bidez organismoak mikrobio patogenoa hobeto identifikatu eta erantzun egokiagoa ematen du haren aurka. Erantzun immunitarioak oroimen immunologikoa du, eta horrek bere eraginkortasuna nabarmen areagotzen du. Eta antigorputzak eta linfozitoak dira erantzun immunitarioaren funtsezko elementuak. Bi motatakoa izan daiteke erantzun immunitarioa: immunitate humorala, alde batetik, antigorputzen bidez gauzatzen dena; eta immunitate zelularra, T linfozitoen bidez bideratzen dena.
Birus patogeno baten sarrerak gure organismoan gure immunitate-sistema aktibatzen du: B linfozitoek birus hori aztertzen dute beren gainazaleko errezeptore proteikoen bidez, eta zelula plasmatiko bihurtzen dira, antigorputzak sortuko dituztenak. Antigorputz horiek birusaren aurka jardungo dute, birusaren antigenoekin lotuko baitira. Antigeno-antigorputz konplexuak fagozitoak erakartzen ditu, opsonizazio prozesuaren bidez, eta fagozito horiek partikula arrotzak irensten dituzte. Fagozitoak ez ezik, antigeno-antigorputz konplexuak konplementu sistema ere aktibatzen du, birus arrotzaren lisia eragiten duena.
Birusaren aurkako antigorputzak sortu dituen B linfozitoen klonaren zelula batzuk oroimen-zelulak bihurtuko dira; zelula hauek oso bizitza luzea dute (hainbat hamarkadatan daude gorputzean) [45] eta egunen batean topo egiten badute antigeno berarekin erantzun immunologiko oso azkarra eta handia emango dute (zelula plasmatiko azkar bihurtuko dira eta antigorputz asko sortuko dituzte) [46].
Bi antigorputz mota dira garrantzitsuak. Lehenengoak IgM izena du, eta oso eraginkorra da birusak neutralizatzeko, baina immunitate sistemako zelulek bakarrik ekoizten dute aste gutxi batzuetan. Bigarrena, IgG izenekoa, denbora luzean sortzen da. Ostalariaren odolean IgM egotea infekzio akutu bat zehazteko erabiltzen da; IgGk, berriz, iraganean izandako infekzio bat adierazten du [47].
Erantzun immunearen bigarren defentsa lerroa immunitate zelularra da, T linfozitoen bidez gauzatzen dena. Immunitate zelularraren kasuan antigeno arrotzak neutralizatzen dituztenak ez dira antigorputzak, immunitate-sistemaren zelula berezi batzuk (T linfozitoak) baizik. Bi T linfozito mota daude: Tc linfozitoak (CD8+ ere deituak) eta Th linfozitoak (CD4+). Lehenengoek partikula arrotz eta inbaditzaileak zuzenean suntsitzen dituzte. Bigarrenak gai dira antigeno arrotz bat ezagutzeko eta berarekin konbinatzeko. CD8+ linfozitoek, bereziki, funtsezko zeregina burutzen dute birus patogenoen suntsiketa prozesuan.
Antibiralak
Zelula barneko parasito hertsiak direnez, birusek lotura oso estua mantentzen dute zelula ostalariarekin. Ondorioz, zaila da zelula ostalaria kaltetzen ez duten substantzia antibiralak erabiltzea. Bakterioen aurkako agente selektibo eraginkorrekin (antibiotikoekin) lortutako arrakasta terapeutikoak ezin dira lortu antibiralekin [48]. Kontuan hartu behar da bakterioen egitura zelularra eta gure gorputzeko zelulena ezberdinak direla, eta horrek bakterioen aurkako antibiotikoen jarduera errazten du, erraza baita bakterioetan soilik dauden egitura zelularrei eraso egitea, gure gorputzeko zelulak kaltetu gabe. Birusekin ez da gauza bera gertatzen.
Lehenengo botika antibirikoak 1960ko hamarkadan agertu ziren, baina botika horien sintesian aurrerakada handia eman zen 1985tik aurrera, hein handi batean HIESaren agerpenarekin batera. Gaur egun, antibiotikoen aldean, botika antibiriko gutxi dago, nahiz eta haien zerrenda urtez urte handitzen den [49].
Antibirikoei esker hainbat gaitzen aurkako tratamendua dago egun: HIESa, herpesbirusek sortzen dituzten gaitzak (barizela, herpes arrunta, herpes zosterra...), B eta C hepatitisa.... A influenzaren birus (gripea) eta papilomabirusen aurka ere antibiriko espezifikoak daude.
Botika antibiralek modu ezberdinetan jarduten dute:
- batzuek genoma birikoaren (DNA ala RNA) sintesia oztopatzen dute (nukleosidoen analogoak dira); hauek arrakasta handiz erabili dira gaixotasun birikoen tratamenduan (zidobudina da horietako bat, timidinaren analogo bat dena; aziklobirra da beste bat, etab.)
- beste batzuek birusaren sarrera zelula ostalarian blokeatzen dute: amantadina (birusaren deskapsidazioa blokeatzen du), errimantidina…
- RNA eta DNAren polimerasak inhibitzen dituztenen artean, azido fosfonoformikoa aipatu behar da.
- alderantzizko transkriptasa entzima (funtsezkoa hainbat RNA birusentzat) inhibitzen dutenak: nebirapina, delabirdina…
- proteasa inhibitzaileak: GIBaren proteasa entzima blokeatzen dute, birusak behar duen entzima polipeptidoen garapena egiteko eta birusaren heltze-prozesua burutzeko. Sakinabirra, erritonabirra eta indinabirra talde honetako antibiralak dira.
- interferoia: goian aipatu den bezala, interferoia gure gorputzak birusen aurka ekoizten duen substantzia naturala da, gaur egun ingeniaritza genetikoaren bidez kopuru handitan lortzen dena. Interferoiak birusaren ugalketa inhibitzen duten proteinen sintesia induzitzen du.
Ondoko taulan antibiral garrantzitsuenak agertzen dira, bere jarduera mekanismoaren arabera sailkatuak eta bakoitzaren aplikazioekin:
| Jarduera mekanismoa | Antibirala | Birus diana |
|---|---|---|
| Birusaren sarrera oztopatzen dutenak | Amantadina, errimantadina, tromantadina | gripearen birusa (amantadina), herpesbirusa (beste biak) |
| Nukleosidoen analogoak | Aziklobirra, erribabirina, remdesivir, zidobudina, zidofobirra, didesoxizitidina | herpesbirus, arnas birus sintzitiala, GIB, herpesbirus eta GIB, hurrenez hurren. |
| Polimerasak inhibitzen dituztenak | Azido fosfonoformikoa | Zitomegalobirusa |
| Alderantzizko transkriptasaren inhibitzaileak | Nebirapina, delabiridina | GIB |
| Proteasaren inhibitzaileak | Sakinabirra, erritonabirra, indinabirra | GIB |
| Proteina antibiralen sintesia eragiten dutenak | Interferoia | B eta C hepatitisen birusak, herpes zosterra, papilomabirus |
Txertoak
Botika antibiriko gutxi daudenez (bakterioen aurkako antibiotikoekin alderatuta), infekzio birikoei aurre egiteko modurik onena profilaxia eta txertoen erabilpena izaten da. Txertoek gaixotasun biriko batzuen hilgarritasun-tasa nabarmen jaitsi dute azken hamarkadetan: poliomielitisarena, elgorriarena, errubeolarena edota hazizurriarena, esaterako. Eta txertaketa masiboak baztangaren desagerpena lortu du [50]. Gaur egun (2021ean) infekzio biriko askoren aurkako txertoak daude, nahiz eta beste batzuen aurka (HIESa, Ebola...) oraindik emaitza positiborik ez egon, ahalegin handiak egin arren.
Txerto antibiriko gehienak birus motelduekin prestatzen dira. Mikrobio motelduak dituzten txertoak, orokorrean, askoz eraginkorragoak dira eta erantzun immunitario iraunkorragoa eragiten dute mikrobio inaktibatuak dituztenak baino. Halere, txerto motelduak arriskutsuak izan daitezke paziente immunogutxituei aplikatzen zaizkienean, pertsona hauengan birusak gaixotasuna sor dezakeelako.[51]
Bioteknologiaren eta ingeniaritza genetikoaren bitartez, birus patogeno baten zati batzuk (antigenoak) besterik ez erabiltzea txertoen prestaketan lortu da, kapsidearen proteinak, batez ere. Txerto horiek askoz ziurragoak dira gaixo immunogutxituentzat, gaitza sortu ezin dutelako.[52] B hepatitisaren txertoa da horietako bat.
COVID-19aren pandemiak txertoen belaunaldi berri baten sorrera ekarri zuen: RNA-m-aren txertoak, hain zuzen. SARS-CoV-2 birusaren material genetikoaren zati bat erabiltzean datza teknologia berri hori.
Gaixotasun biriko baten erabateko desagerpena txertoen bidez baztangarekin bakarrik lortu da. Edozein gaixotasun biriko desagerrarazteko, beharrezkoa da animalia-gordailu bakar bat ere ez egotea, txerto eraginkorra egotea, infektibitate errepikakorrik ez egotea eta serotipo egonkor bat edo gutxi egotea. Baztangaren ondoren, gure planetatik laster desager daitezkeen gaixotasun birikoak poliomielitisa eta elgorria dira [53]
Birusen jatorria
Hiru dira hipotesi nagusiak birusen jatorriari dagokionez:
- Erregresio zelularraren hipotesia: hasiera batean birusak bakterioen antzeko zelula txikiak izango ziren, zelula handiagoak parasitatuko zituztenak. Parasitismo horren ondorioz zelula txiki horiek zelula-endekapen bat jasango zuten, eta beharrezkoak ez zituzten egitura zelular asko galdu, gaur egungo egitura sinpleraino iritsi arte (material genetikoa eta proteinazko kapsidea) [54]. Zelula barneko parasitismoak tresneria metabolikoaren (entzimak, erribosomak, organuluak....) galera erraztuko luke, zelula ostalarian daudelako beharrezko metabolitoak. Horrenbestez, birusak material genetikoa eta proteina gutxi batzuk besterik ez lituzke beharko infektatu ahal izateko. Bakterioen artean badaude birusen antzeko Zelula barneko parasito hertsiak: Chlamydia eta Rickettsia generoak. Bakterio horiek ere metabolismo mugatua dute, eta molekula asko zelula ostalariatik hartu behar dituzte. Hala ere, bakterio horiek ez dira iritsi birusen egitura sinpleetaraino.
- Jatorri zelularraren hipotesia: aurreko hipotesiaren aldean, hipotesi honek suposatzen du birusen jatorria zeluletan dagoela. Zelula batzuek DNA edo RNAren zati batzuk galduko zituzten; zati horiek independente bihurtuko ziren eta infektatzeko gaitasuna garatu. Posible da irudikatzea aldaketa genetiko batzuen bidez zelularen DNA puska bat DNA biriko bihurtuko zela, bere proteina biriko propioak sintetizatzeko gai zena, eta DNA zelularretik desintegratzeko gai zena, horiekin bat egin eta birus berri bat osatzeko. DNA birusen jatorria hori izan liteke, nahiz eta RNA birusen jatorria azaltzea zailagoa den. Argi dagoena da aspektu askotan antzekotasun handia dagoela plasmidoen eta birusen artean, eta baita transposoien eta birusen artean ere.
- Koeboluzioaren hipotesia: birusak azido nukleikoetatik eta proteinetatik abiatuta eratu ziren bizitzaren sorreraren lehen etapetan, lehen zelulak agertzearekin batera, horiekin batera eboluzionatuz eta horietako batzuk infektatuz. Hipotesi honen arabera, beraz, birusak oso forma sinpleak direla jada sorreratik bertatik. Biroideak, birusak baino are sinpleagoak diren egitura infekziosoak (RNAz soilik osatuak, bilgarri proteikorik gabe), erdiko fase ebolutibo bat izan litezke, birus primitiboen eta gaur egungoen artean [55]
Bizidunak al dira birusak?

Eztabaida luzeak egon dira aspalditik biologoen artean birusen jarduera biologikoa aztertzerakoan. Bizidunak al dira birusak? Elementu genetiko hutsak dira batzuentzat. Hainbat zientzialarik, aldiz, molekula hutsak baino zerbait gehiago direla proposatzen dute.
Gaur egun biologo gehienek uste dute birusak biziaren mugan dauden elementuak direla, ez benetako bizidunak [56] [57].
Alde batetik, gainontzeko bizidunen aldean birusek ez dute egitura zelularrik, ez dira zelulak. Biziaren unitate txikienak zelulak direla onartzen baldin bada, birusak -horrenbestez- ez lirateke bizidunak izango.
Bestalde, birusek ez dute jarduera metabolikorik, bizidunen funtsezko beste ezaugarri bat. Izan ere, zelula ostalari bat infektatzen ez dutenean birioiak guztiz inerteak dira, ez dute jarduera biologiko txikienik ere erakusten.
Halere, infektatzen dutenean birusek erreplikatzeko ("ugaltzeko") gaitasuna dute, biziaren ezaugarri hori garatzeko gai dira. Zelula ostalariaren barnean proteinak eta azido nukleikoak sortu eta mihiztatzen dituzte, birus berriak osatzeko. Partikula berriak eratzeko duten modua ez da kristal huts batena.
Gainontzeko bizidunen antzera, birusen material genetikoak ere aldaketak ditu mutazio edo birkonbinazio genetikoaren bidez, eta hautespen naturalaren menpe dago, honek aldaketa onuragarriak hautatuko baititu eta besteak baztertu.
Baina egitura zelularra, ugalketa autonomoa eta metabolismoa hartzen baldin badira biziaren ezaugarritzat, birusak, argi eta garbi, bizidunen kategoriatik kanpo kokatu behar dira.
Sailkapena
Birusak sailkatzeko hiru modu daude: Baltimore sailkapena, ICTV eta Holmes.
Baltimore sailkapena
David Baltimore Nobel saridunak Baltimoreko sailkapen-sistema asmatu zuen. Sailkapen honek mRNA produkzioaren mekanismoan oinarritzen da. ICTV (International Committee of Taxonomy of Viruses) sailkapena gaur egungo birusen sailkapen-sistemarekin batera erabiltzen da.
Birusek mRNAk genomatik sortu behar dituzte, proteinak sintetizatzeko eta erreplikatzeko. Hori lortzeko, mekanismo ezberdinak erabiltzen dira birus familia bakoitzean. Genoma biralak honelakoak izan daitezke:
- Kate bakunekoak (ss) edo kate bikoitzekoak (s).
- RNA edo DNAdunak.
- Alderantzizko transkriptasa entzima erabiltzen dutenak edo hura erabiltzen ez dutenak.
Gainera, ssRNA birusak sense (+) edo antisense (-) izan daitezke. Sailkapen horrek zazpi taldetan kokatzen ditu birusak [58] :
- I: Kate bikoitzeko DNA birusa.
- II: Kate bakuneko DNA birusa.
- III: Kate bikoitzeko RNA birusa.
- IV: Kate bakun (+) RNA birusa.
- V: Kate bakun (-) RNA birusa.
- VI: Kate bakuneko alderantzizko transkripziodun RNA birusa.
- VII: Kate bikoitzeko alderantzizko transkripziodun DNA birusa.
Material genetikoa eta birusak erreplikatzeko metodoa desberdinak dira:
DNA birusa
DNA birus gehienen genomaren erreplikazioa zelularen nukleoan gertatzen da. Zelula ostalariak gainazalerako hartzailea egokia badu, birusak zelula-mintzarekin fusionatuz edo endozitosiz sartzen dira. DNA birus gehienak zelula ostalariaren DNA eta RNAren sintesi-makineriaren eta RNA prozesatzeko makineriaren mende daude. Bi talde bereizten dira: [59] [60] [61][62]
Kate bikoitzeko DNA birusa
Birus mota horrek kate bikoitzeko DNAz osatutako material genetikoa du, eta zelula ostalariaren polimerasak behar dira genoma birala erreplikatzeko [60]. Birusaren infekzioa eta ekoizpena egin ahal izateko, zelula erreplikazio-fasean egon behar da, orduan baitaude zelularen polimerasak aktibo. Adibideak:
- Adenoviridae
- Papovaviridae
- Herpesviridae
- Poxviridae
- Pleolipoviridae
Kate bakuneko DNA birusa
Birus mota horrek kate bakuneko DNA material genetikoa du, eta DNAren mendeko DNA polimerasa bat erabiliz erreplikatzen da, kate bikoitzeko DNA birusa bezala [63]. Kate bakuneko DNA kate bikoitzeko DNA bihurtu behar da erreplikazioa egon dadin [61]. Adibideak:
- Parvoviridae
- Anelloviridae
- Pleolipoviridae
RNA birusa
RNA birusak apartekoak dira beren informazio genetikoa RNAn kodetuta dagoelako; horrek esan nahi du azido erribonukleikoa (RNA) material genetiko gisa erabiltzen dutela, edo erreplikazio-prozesuan RNA behar duela. Erreplikazioa zitoplasman gertatu ohi da. Oro har, RNA birusak lau taldetan sailka daitezke, erreplikazio-moduaren arabera. RNAren polaritateak zehazten du, neurri handi batean, erreplikazio-mekanismoa zein den, eta ea material genetikoa kate bakunekoa edo bikoitzekoa den. Gainera, RNA birusek beren RNA polimerasa erabiltzen dute beren genomaren kopiak sortzeko.
Kate bikoitzeko RNA birusa
Kate bikoitzeko RNA birusak bitarteko DNA erabiliz erreplikatzen ez diren birusak dira. RNA birus gehienak bezala, zitoplasman erreplikatzen dira, eta ez daude zelula ostalarien polimerasen mende, DNA birusek egiten duten bezala, entzima horiek birioan sartzen baitira.[63]
Itzulpena monozistronikoa izaten da, eta horrek esan nahi du segmentu bakoitzak proteina bakarra kodetzen duela, itzulpen konplexuagoa erakusten duten beste birus batzuek ez bezala. Ezaugarri berezi bat RNAko segmentu bikatenarioen transkripzioa baldintza egokietan egiteko gaitasuna da, kapsideren barruan. Adibideak:
Bakterioak infektatzen dituzten birusak:
- Chrysoviridae
- Quadriviridae
- Hypoviridae
- Megabirnaviridae
- Curvulaviridae
Animaliak infektatzen dituzten birusak:
- Cystoviridae
- Picobirnaviridae
Kate bakun (+) RNA birusa
Kate bakun positiboko RNA birusek zentzu positiboko azido erribonukleikoko (RNA) kate sinplea dute material genetiko gisa, eta ez dira erreplikatzen tarteko DNA erabiliz. RNA birus positiboak RNA mezulariaren berdinak dira eta, beraz, zelula ostalariak berehala itzul ditzake. Birus positibo baten RNA araztuak zuzenean infekzioa eragin dezakeen arren, ez da birus osoa bezain infekziosoa. Erreplikazioa zitoplasman gertatzen da batez ere, eta ez dago zelula-zikloaren mende, DNA birusetan bezala. Zentzu positiboko RNA birusek RNA mezulariaren polaritate bera duten genomak dituzte, eta zuzenean erabil daitezke proteinen sintesirako, zelula ostalariaren itzulpen-makineria erabiliz. Kodetutako proteina horietako bat RNA erreplikasa da. Hura RNA polimerasa bat da eta RNA birala kopiatzen du bitarteko DNA kate batetik igaro beharrik gabe. Talde honetako birusek edozein organismo infekta dezakete (animaliak, landareak, onddoak, protistak, bakterioak eta arkeoak). Hala ere, animalia eta landareetan nagusitzen dira. Adibideak:
- Picornaviridae
- Caliciviridae
- Togaviridae
- Flaviviridae
- Coronaviridae
- Astroviridae
- Arteriviridae
- Hepeviridae
Kate bakun (-) RNA birusa
Birus honek azido erribonukleikoa du (RNA), kate bakun (-) da, eta ez da erreplikatzen bitarteko DNA erabiliz. RNA biral negatiboa RNA mezulariaren osagarria da. Hori dela eta, RNA positibo bihurtu behar da itzulpenaren aurretik. Prozesu hori RNA polimerasa batek egingo du. Bestalde, birus negatibo baten RNA purifikatua ez da berez infekziosoa, RNA positibo bihurtu behar baita. RNA (-) birusek RNA polimerasa edo RNA transkriptasa erabiltzen dute RNA (+) bihurtzeko. Horretarako, birusak RNA polimerasa entzima erabili behar du, RNAren mendekoa baita entzima hura. Ondorioz, RNA (+) molekulak mezulari biral gisa jokatzen du. Ondoren, ostalariak RNA hura proteina bihurtzen ditu (erribosometan gertatzen da prozesu hura). Azkenik, sorturiko proteinei esker, birioi berrien elementuak ekoizten dira, hala nola, kapsideko eta RNA erreplikasako proteinak. Azken hori RNA (-) molekula berrien ekoizpenaz arduratzen da. Adibideak:
- Arenaviridae
- Orthomyxoviridae
- Paramyxoviridae
- Bunyaviridae
- Rhabdoviridae
- Filoviridae
- Bornaviridae
Alderantzizko transkripziodun birusak
Alderantzizko transkripziodun birusek bi genoma mota izan ditzakete:
Kate bakuneko alderantzizko transkripziodun RNA birusa:
Kate bakuneko alderantzizko transkripziodun RNA birusa (edo ssRNA-RT birusa) kate bakuneko RNA duen birus bat da, eta zelula ostalarian erreplikatzen da alderantzizko transkripzioaren bidez, hau da, RNA-tik abiatuz DNA sortzen du[63]. Birus horiek alderantzizko transkriptasa erabiltzen dute, hau da, RNAren mendeko DNA polimerasa bat. Era horretan, RNA genoma biraletik abiatuta DNA sortzen da. DNA hori askotan ostalariaren genoman txertatzen da. Kasu horietan, ostalariak DNA erreplikatu eta transkribatzen du, erretrobirusen eta pseudobirusak kasu. Adibideak:
- Pseudoviridae
- Metaviridae
- Belpaoviridae
Kate bikoitzeko alderantzizko transkripziodun DNA birusa:
Birusak alderantzizko transkripzioaren bidez erreplikatzen dira, aurreko kasuan aipatu den bezala[63]. Batetik, RNA genomadun birusek DNA bitarteko bat erabiltzen dute genoma erreplikatzeko, eta, bestetik, DNA genomadun birusek, RNA bitarteko bat erabiltzen dute genoma erreplikatzeko. Adibideak:
- Hepadnaviridae
- Caulimoviridae
Eskema eta jarraibideak
Birusen Taxonomiaren Nazioarteko Batzordea (ICTV, ingelesezko sigletan) 1970eko hamarkadaren hasieran hasi zen birusak izendatzeko eta sailkatzeko arauak diseinatzen eta ezartzen. ICTV da Elkarte Mikrobiologikoen Nazioarteko Batasunak birus unibertsalaren taxonomia garatu, findu eta mantentzeko ardura duen erakunde bakarra. Sistemak ezaugarri asko partekatzen ditu organismo zelularrak sailkatzeko erabiltzen den sistemarekin, hala nola, taxoiaren egitura. Hala ere, badira desberdintasun batzuk, adibidez, izen taxonomiko guztietarako letra etzanaren erabilera unibertsala; aldiz, alga, onddo eta landareentzako, Nomenklaturaren Nazioarteko Kode, Bakterioen Nomenklaturaren Nazioarteko Kode eta Nomenklatura Zoologikoaren Nazioarteko Kodeetan ez da erabiltzen.
Sailkapen birala domeinu-mailan hasten da, eta honela idazten da, atzizki taxonomikoekin parentesi artean jarrita.
Taxonomiaren egitura orokorra hurrengoa da:
- Domeinua (-viria)
- Erreinua (-virae)
- Filuma (-viricota)
- Subfiluma (-viricotina)
- Klasea (-viricetes)
- Ordena (-virales)
- Subordena (-virineae)
- Familia (-viridae)
- Subfamilia (-virinae)
- Generoa (-virus)
- Espeziea (-virus)
Izaki bizidunen espezieetan hartutako nomenklatura binomialaren sisteman ez bezala, gaur egun ez dago forma estandarizaturik birus-espezieen izenetarako. Gaur egun, ICTVk espezie baten izenak ahalik eta hitz gutxien izan behar dituela eskatzen du, desberdina izateari utzi gabe, eta ez du soilik birus hitza eta ostalariaren izena eduki behar. Espezieen izenek gaixotasun-birus itxura hartzen dute askotan, batez ere goiko landare eta animalien birusen kasuan. 2019an ICTVk proposamen bat argitaratu zuen birus-espezieen izenetarako nomenklatura binomialeko sistema formalizatuagoa ezartzeko, eta 2020an bozkatu zen.
Espezie biralen definizioa
Espezieak sailkapen biologikoko edozein sistemaren oinarri dira. 1982 baino lehen, uste zen birusak ezin zirela egokitu Ernst Mayr espeziearen ugalketa-kontzeptura, eta, beraz, ezin zirela horrela sailkatu. 1982an ICTV espezie bat "andui talde" gisa definitzen hasi zen, eta handik aurrera andui horiek identifikatzeko ezaugarri jakin batzuk kontuan hartu behar ziren. 1991n oinarri espezifiko bat onartu zen: birus-espezie bat birus klase politetiko bat da, erreplikazio-leinu bat osatzen duena eta nitxo ekologiko jakin bat betetzen duena.
2013ko uztailean, ICTV espeziearen definizioa aldatu egin zen honako hau adierazteko: "Espezie bat birusen talde monofiletiko bat da, eta birusen propietateak beste espezie batzuengandik bereiz daitezke, hainbat irizpideren arabera". Birusak entitate fisiko errealak dira, eta eboluzio biologikoak zein genetikak sortu dituzte. Birus-espezieak eta goi-mailako taxoiak, berriz, pentsamendu arrazionalak eta logikak sortutako kontzeptu abstraktuak dira.
Erabilitako irizpide errealak aldatu egiten dira taxoiaren arabera, eta koherentziarik gabekoak izan daitezke edo leinuarekin (geografia) loturarik ez dutenak batzuetan. Oraindik konpondu gabe dagoen arazoa da.
Bestelako sailkapen historikoak: Holmes sailkapena
Holmesek (1948) nomenklatura binomiala zeukan taxonomia lineal bat erabili zuen birusak ordena bakar baten barnean (Virales ordenean) eta 3 taldetan sailkatzeko. Honela zeuden sailkatuta:
- I. taldea: Phaginae (bakterio-erasotzaileak)
- II. taldea: Phytophaginae (landare-erasotzaileak)
- III. Taldea: Zoophaginae (animalia-erasotzaileak)
Beste ikertzaileek ez zuten sistema onartu, antzekotasun morfologikoak alde batera uzten zituelako.[64]
Erreferentziak
- Koonin, Eugene V; Senkevich, Tatiana G; Dolja, Valerian V. (2006). «[No title found»] Biology Direct 1 (1): 29. doi:. PMID 16984643. PMC PMC1594570. (Noiz kontsultatua: 2019-10-03).
- Dimmock 2007, 4 orr. .
- Dimmock 2007, 49 orr. .
- (Ingelesez) Breitbart, Mya; Rohwer, Forest. (2005-6). «Here a virus, there a virus, everywhere the same virus?» Trends in Microbiology 13 (6): 278–284. doi:. (Noiz kontsultatua: 2019-10-03).
- (Ingelesez) Lawrence, C. Martin; Menon, Smita; Eilers, Brian J.; Bothner, Brian; Khayat, Reza; Douglas, Trevor; Young, Mark J.. (2009-05-08). «Structural and Functional Studies of Archaeal Viruses» Journal of Biological Chemistry 284 (19): 12599–12603. doi:. ISSN 0021-9258. PMID 19158076. PMC PMC2675988. (Noiz kontsultatua: 2019-10-03).
- (Ingelesez) Edwards, Robert A.; Rohwer, Forest. (2005-06). «Viral metagenomics» Nature Reviews Microbiology 3 (6): 504–510. doi:. ISSN 1740-1534. (Noiz kontsultatua: 2019-10-03).
- Canchaya, Carlos; Fournous, Ghislain; Chibani-Chennoufi, Sandra; Dillmann, Marie-Lise; Brüssow, Harald. (2003-08-01). «Phage as agents of lateral gene transfer» Current Opinion in Microbiology 6 (4): 417–424. doi:. ISSN 1369-5274. (Noiz kontsultatua: 2019-10-03).
- Ryan, Frank, 1944-. Virolution : the most important evolutionary book since Dawkins' Selfish Gene. ISBN 9780007315123. PMC 317747594. (Noiz kontsultatua: 2019-10-03).
- Rybicki, EP. The classification of organisms at the edge of life, or problems with virus systematics. South African Journal of Science. 1990;86:182–86
- Koonin, Eugene V.; Starokadomskyy, Petro. (2016-10-01). «Are viruses alive? The replicator paradigm sheds decisive light on an old but misguided question» Studies in History and Philosophy of Science Part C: Studies in History and Philosophy of Biological and Biomedical Sciences 59: 125–134. doi:. ISSN 1369-8486. PMID 26965225. PMC PMC5406846. (Noiz kontsultatua: 2019-10-03).
- Albero, Josu: Mikrobioen Mundu Liluragarria, EHUak argitaratuta (2019)29-30 orr. ISBN:978-84-1319-082-2
- Shors 2008, 49-50 orr. .
- .
- Shors 2008, 76-77 orr. .
- Topley, W. W. C. (William Whiteman Carlton), 1886-1944.. (1998). Topley & Wilson's microbiology and microbial infections.. (9th ed.. argitaraldia) Arnold ISBN 0340614706. PMC 38290809. (Noiz kontsultatua: 2019-10-07).
- Dimmock 2007, 4-5 orr. .
- Desk encyclopedia of general virology. Academic Press 2010 ISBN 0123751624. PMC 480587729. (Noiz kontsultatua: 2019-10-07).
- Shors 2008, 589 orr. .
- Publications service. (2007-09-01). «On an invisible microbe antagonistic toward dysenteric bacilli: brief note by Mr. F. D'Herelle, presented by Mr. Roux» Research in Microbiology 158 (7): 553–554. doi:. ISSN 0923-2508. (Noiz kontsultatua: 2019-10-07).
- (Ingelesez) Domingo‐Calap, P.; Georgel, P.; Bahram, S.. (2016). «Back to the future: bacteriophages as promising therapeutic tools» HLA 87 (3): 133–140. doi:. ISSN 2059-2310. (Noiz kontsultatua: 2019-10-07).
- (Ingelesez) Steinhardt, Edna; Israeli, C.; Lambert, R. A.. (1913-09-01). «Studies on the Cultivation of the Virus of Vaccinia» The Journal of Infectious Diseases 13 (2): 294–300. doi:. ISSN 0022-1899. (Noiz kontsultatua: 2019-10-07).
- From Nobel Lectures, Physics 1981-1990, (1993) Editor-in-Charge Tore Frängsmyr, Editor Gösta Ekspång, World Scientific Publishing Co
- Creager, A. N., Morgan, G. J. (2008ko ekaina). «After the double helix: Rosalind Franklin's research on Tobacco mosaic virus.» Isis 99 (2): 239-72
- Norrby, E. (2008). «Nobel Prizes and the emerging virus concept.» Arch. Virol. 153 (6): 1109-23
- Temin, H. M., Baltimore, D. (1972). «RNA-directed DNA synthesis and RNA tumor viruses.» Adv. Virus Res. 17: 129-86. doi:10.1016/S0065-3527(08)60749-6
- Barré-Sinoussi, F., Chermann, J. C., Rey, F., Nugeyre, M. T., Chamaret, S., Gruest, J., Dauguet, C., Axler-Blin, C., Vezinet-Brun, F., Rouzioux, C., Rozenbaum, W., and Montagnier, L. (1983). «Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS).» Science 220 (4599): 868-871
- Madigan M.T., Martinko J.M., Parker J. Brock Mikroorganismoen biologia (2007) E.H.U-ak euskaratua: 240 orr. ISBN: 978-84-9860-026-1.
- Basaras, M., Umaran, A.: Mikrobiologia medikoa, EHUak argitaratuta , 390 orr. ISBN: 84-8373-658-6
- Aranda Anzaldo, A.La estructura de los virus 6-7 orr.
- Madigan M.T., Martinko J.M., Parker J. Brock Mikroorganismoen biologia (2007) E.H.U-ak euskaratua: 238-239 orr. ISBN: 978-84-9860-026-1.
- Infosida Glosario del VIH; Genoma
- The history of vaccines Los virus y su evolución 2018ko urtarrila
- Worobey M, Holmes EC (1999). "Evolutionary aspects of recombination in RNA viruses". J. Gen. Virol. 80 ( Pt 10): 2535–43
- Lukashev AN (2005). "Role of recombination in evolution of enteroviruses". Rev. Med. Virol. 15 (3): 157–67
- Basaras, M., Umaran, A.: Mikrobiologia medikoa, EHUak argitaratuta , 410-411 orr. ISBN: 84-8373-658-6
- Aldaba, J., Lopez, P., Pascual M.M., Urzelai, A.: Biologia, 2. Batxillergoa, Elkar, 2006, 322-323 orr. ISBN: 84-9783-222-1
- Madigan M.T., Martinko J.M., Parker J. Brock Mikroorganismoen biologia (2007) E.H.U-ak euskaratua: 259-260 orr. ISBN: 978-84-9860-026-1.
- Campbell and Reece (2005). Biology. San Francisco: Pearson. 338–339 orr.
- Ingraham, J.L., Ingraham, C. Introducción a la Microbiologia Vol. 2, Ed. Reverté, (1998), 372-373 orr. ISBN 84-291-1871-3
- Albero, Josu: Mikrobioen Mundu Liluragarria, EHUak argitaratuta (2019)19-20 orr. ISBN:978-84-1319-082-2
- Boyton R, Openshaw P «Pulmonary defences to acute respiratory infection». Br Med Bull, 61, 2002, 1-12 orr.
- Janeway CA, Jr. et al (2005). Immunobiology. (6th ed.). Garland Science
- Madigan M.T., Martinko J.M., Parker J. Brock Mikroorganismoen biologia (2007) E.H.U-ak euskaratua:829 orr. ISBN: 978-84-9860-026-1.
- Basaras, M., Umaran, A.: Mikrobiologia medikoa, EHUak argitaratuta , 47 orr. ISBN: 84-8373-658-6
- Crotty, Shane, et al. "Cutting edge: long-term B cell memory in humans after smallpox vaccination." The Journal of Immunology 171.10 (2003): 4969-4973
- Sistema immune indartsua Miren Basaras, BERRIA, 2020-04-1ean
- Greer, S., Alexander, G. J. «Viral serology and detection.» Baillieres Clin Gastroenterol. 1995 Dec;9(4):689-721
- Madigan M.T., Martinko J.M., Parker J. Brock Mikroorganismoen biologia (2007) E.H.U-ak euskaratua:762 orr. ISBN: 978-84-9860-026-1
- Ingraham, J.L., Ingraham, C. Introducción a la Microbiologia Vol. 2, Ed. Reverté, (1998), 509 orr. ISBN 84-291-1871-3
- Lane, J. M. (2006). «Mass vaccination and surveillance/containment in the eradication of smallpox». Curr. Top. Microbiol. Immunol. 304: 17-29
- Thomssen, R. (1975). «Live attenuated versus killed virus vaccines». Monographs in allergy 9: 155-76
- Casswall, T. H., Fischler, B. (2005). «Vaccination of the immunocompromised child». Expert review of vaccines 4 (5): 725-38
- Forbes, B., Sahm, D., Weissfeld, A.: Diagnóstico microbiológico Ed. Panamericana (2002), 888-889 orr. ISBN: 84-7903-810-1
- Mahy WJ & Van Regenmortel MHV (eds) (2009). "Desk Encyclopedia of General Virology". Oxford: Academic Press: 24. ISBN 0-12-375146-2
- Tsagris E. M., de Alba A. E., Gozmanova M., Kalantidis K. «Viroids». Cell. Microb. 2008ko iraila
- Albero, Josu: Mikrobioen mundu liluragarria, EHUak argitaratuta (2019) 28-30 orr. ISBN: 978-84-1319-082-2
- Rybicki, EP (1990). "The classification of organisms at the edge of life, or problems with virus systematics". S Afr J Sci 86: 182–186.
- «LAS VEGAS SANDS CORP., a Nevada corporation, Plaintiff, v. UKNOWN REGISTRANTS OF www.wn0000.com, www.wn1111.com, www.wn2222.com, www.wn3333.com, www.wn4444.com, www.wn5555.com, www.wn6666.com, www.wn7777.com, www.wn8888.com, www.wn9999.com, www.112211.com, www.4456888.com, www.4489888.com, www.001148.com, and www.2289888.com, Defendants.» Gaming Law Review and Economics 20 (10): 859–868. 2016-12 doi:. ISSN 1097-5349. (Noiz kontsultatua: 2023-11-03).
- Shors, Tracey J.. (2009-03). «Saving New Brain Cells» Scientific American 300 (3): 46–54. doi:. ISSN 0036-8733. (Noiz kontsultatua: 2023-11-03).
- Dimmock, Nigel J.; Easton, Andrew J.; Leppard, Keith N.. (2008). Introduction to modern virology. (6. ed., [Nachdr.]. argitaraldia) Blackwell Publ ISBN 978-1-4051-3645-7. (Noiz kontsultatua: 2023-11-03).
- Patton, John T.. (2008). Segmented double-stranded RNA viruses: structure and molecular biology. Caister academic press ISBN 978-1-904455-21-9. (Noiz kontsultatua: 2023-11-03).
- (Ingelesez) Robertson, Michael P; Igel, Haller; Baertsch, Robert; Haussler, David; Ares, Manuel; Scott, William G. (2004-12-28). Wickens, Marv ed. «The Structure of a Rigorously Conserved RNA Element within the SARS Virus Genome» PLoS Biology 3 (1): e5. doi:. ISSN 1545-7885. (Noiz kontsultatua: 2023-11-03).
- Kornberg, Arthur; Baker, Tania A.. (2005). DNA replication. (2. ed., paperback ed. argitaraldia) University Science Books ISBN 978-1-891389-44-3. (Noiz kontsultatua: 2023-11-03).
- (Ingelesez) Kuhn, Jens H.. (2021). «Virus Taxonomy» Encyclopedia of Virology (Elsevier): 28–37. doi:. ISBN 978-0-12-814516-6. PMC PMC7157452. (Noiz kontsultatua: 2023-11-03).
Bibliografia
- Dimmock, N. J.. (2007). Introduction to modern virology. (6th ed. argitaraldia) Blackwell Pub ISBN 1405136456. PMC 65207057. (Noiz kontsultatua: 2019-10-03).
- Shors, Teri. (2008). Understanding viruses. Jones and Bartlett Publishers ISBN 9780763729325. PMC 124505697. (Noiz kontsultatua: 2019-10-07).
- Mahy, W.J.. (2009). Desk Encyclopedia of General Virology. Van Regenmortel MHV ISBN 0123751462...
Ikus, gainera
Kanpo estekak
- (Ingelesez) Viralzone Birus mota guztien informazioa, maila molekularrean zein epidemiologikoan.
- (Ingelesez) Viruses-Open Acces Virology Journal Aldizkari espezializatua
- (Ingelesez) Birusak hiru dimentsiotan The European Bioinformatics Institute
- Birusek sendatzeko ere balio dute EITB, Teknopolis (2020ko martxoa)
| Autoritate kontrola |
|---|