Archaea
Archaea edo Arkeoak[oh 1]. izaki bizidun zelulabakarren domeinu bat da. Organismo hauek ez dute zelula nukleorik, eta beraz prokariotoak dira. Hasiera batean bakterioen barruan sailkatzen ziren, eta horregatik deitzen zitzaien Archaebacteria edo arkeobakterio, baina termino hau jada ez da hainbeste erabiltzen[4].
| Archaea | |
|---|---|
 | |
| Sailkapen zientifikoa | |
| Domeinua | Archaea |
| Azpibanaketa | |
Erreinuak[2]
| |
| Datu orokorrak | |
| Mugimendua | Errodadura |
| Aurkitzailea | Carl Woese (en) |
Archaea zelulek beste bi domeinuetatik, Bacteria eta Eukaryota, bereizten dituzten ezaugarriak dituzte. Archaea hainbat filumetan bereizten da. Hala ere, sailkapena zaila da, gehienak ez direlako laborategian isolatu eta soilik inguruneetako laginetan hartutako sekuentzia genetiko bidez identifikatu direlako. Ez dago argi detektatutako horiek endosporak sortzeko gai ote diren.
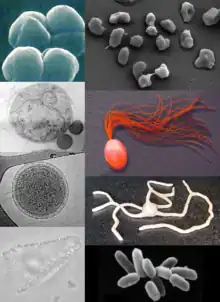
Oro har, arkeoak eta bakterioak antzekoak dira tamainari eta formari dagokienez, nahiz eta arkeo batzuek oso forma desberdinak izan, hala nola Haloquadratum walsbyi zelula lau eta karratuak[5]. Bakterioekin antzekotasun morfologiko hori izan arren, arkeoek eukariotoekin lotura estuagoa duten geneak eta zenbait bide metaboliko dituzte, batez ere transkripzioan eta itzulpenean parte hartzen duten entzimentzat. Arkeoen biokimikaren beste alderdi batzuk bakarrak dira, hala nola eter lipidoekiko mendekotasuna zelula-mintzetan[6], arkeoloak barne. Arkeoek eukariotoek baino energia-iturri gehiago erabiltzen dituzte: konposatu organikoak, hala nola azukreak, amoniakoa, ioi metalikoak edo hidrogeno-gasa. Gatzarekiko tolerantzia duen Haloarchaeak, eguzki-argia erabiltzen dute energia-iturri gisa, eta beste arkeoa espezie batzuek karbonoa finkatzen dute (autotrofoak dira), baina, landareek eta zianobakterioek ez bezala, ez da ezagutzen arkeorik biak batera egiten duenik. Arkeoak asexualki ugaltzen dira fisio bitarraren, fragmentazio edo gemazio bidez; bakterioak ez bezala, ez da endosporak eratzen dituen espezie ezagunik. Behatutako lehen arkeoak extremofiloak ziren eta muturreko inguruneetan bizi ziren, iturri termaletan eta laku gazietan, beste organismorik gabe. Molekulak detektatzeko tresnak hobetuta, ia habitat guztietan arkeoak aurkitu ziren, lurzoruan[7], ozeanoetan eta paduretan barne. Arkeoak oso ugariak dira ozeanoetan, eta planktonean dauden arkeoak planetako organismo-talde ugarienetako bat izan daitezke.
Arkeoak Lurreko biziaren zati garrantzitsua dira. Organismo guztien mikrobiotaren parte dira. Giza mikrobioman garrantzitsuak dira hestean, ahoan eta azalean[8]. Aniztasun morfologiko, metaboliko eta geografikoari esker, hainbat funtzio ekologiko bete ditzakete: karbonoa finkatzea, nitrogenoaren zikloa, konposatu organikoak berritzea eta mikrobio-komunitate sinbiotiko eta sintrofikoak mantentzea, adibidez[9].
Ez da ezagutzen arkeo patogenoen edo parasitoen adibide garbirik. Aldiz, mutualistak edo komentsalistak izan ohi dira, hala nola metanogenoak (metanoa sortzen duten anduiak), gizakien eta hausnarkarien traktu gastrointestinalean bizi direnak, non kopuru handiak digestioa errazten baitu. Metanogenoak biogasa ekoizteko eta hondakin-urak tratatzeko ere erabiltzen dira, eta bioteknologiak tenperatura altuak eta disolbatzaile organikoak jasan ditzaketen arkeo extremofiloen entzimak ustiatzen ditu.
Historia
Kontzeptuaren sorrera
.jpg.webp)
XX. mendearen zati handi batean, prokariotak organismo-talde bakartzat hartu ziren, eta beren biokimikaren, morfologiaren eta metabolismoaren arabera sailkatu ziren. Mikroorganismoak sailkatzeko, haien zelula-hormen egiturak, formak eta kontsumitzen dituzten substantziak hartu zituzten oinarri mikrobiologoek[10]. 1965ean, Emile Zuckerkandlek eta Linus Paulingek[11] prokariotetako geneen sekuentziak erabiltzea proposatu zuten, haien ahaidetasuna jakiteko. Ikuspegi filogenetiko hori da gaur egun erabiltzen den metodo nagusia[12].
1977an, Carl Woesek eta George E. Foxek lehen aldiz sailkatu zituzten arkeak eta bakterioak, RNA erribosomikoaren (rRNA) geneetan oinarrituta[13]. (Garai hartan metanogenoak baino ez ziren ezagutzen). Talde horiei Urkingdom Archaebacteria eta Eubacteria deitu zieten, baina beste ikertzaile batzuek erresuma edo azpierresuma gisa tratatu zituzten. Woesek eta Foxek lehen frogak aurkeztu zituzten, hots, arkeobakterioek "ondorengotza-lerro" independentea osatzen dutela: 1. Zelula-hormetan peptidoglikanorik ez izatea, 2. ezohiko bi koentzima, 3. RNA erribosomikoaren 16S genearen sekuentziazioaren emaitzak. Alde hori azpimarratzeko, Woese, Otto Kandler eta Mark Wheelisek geroago proposatu zuten organismoak hiru domeinuen sistemarekin birsailkatzea: Eukarya, Bacteria eta Archaea[14]. Gaur egun, Woesear iraultza deitzen zaio[15].
Archaea hitza antzinako grezierako ἀρχαῖα hitzetik dator, eta "gauza zaharrak" esan nahi du; izan ere, Archaea domeinuko lehen ordezkariak metanogenoak ziren, eta haien metabolismoak Lurraren hasierako atmosfera eta organismoen antzinatasuna islatzen zituen, baina habitat berriak aztertu ahala, organismo gehiago aurkitu ziren. Mikrobio halofiloak[16] eta muturreko hipertermotofiloak[17] ere Archaean sartu ziren. Denbora luzez, muturreko habitatetan soilik zeuden arkeoak, hala nola ur termaletan eta laku gazietan, baina XX. mendearen amaieran arkeoak ere identifikatu ziren muturrekoak ez ziren inguruneetan. Gaur egun, jakina da naturan oparo banatutako organismo-multzo zabal eta askotarikoa osatzen dutela[18]. Arkeoen garrantzia eta nonahikotasuna berriro hautemateko, polimerasaren kate-erreakzioa (PCR) erabili zen, prokariotak detektatzeko, ingurumeneko laginetan (ura edo lurzorua, esaterako), gene erribosomikoak biderkatuz. Horri esker, laborategian landu ez diren organismoak hauteman eta identifika daitezke[19][20].
Sailkapena

Arkeoen eta, oro har, prokarioten sailkapena bilakaera azkarreko eta polemikoko eremua da. Gaur egungo sailkapen-sistemen helburua da ezaugarri estrukturalak eta arbaso komunak dituzten organismo-multzoetan antolatzea arkeoak[21]. Sailkapen horiek, neurri handi batean, RNA erribosomikoaren gene-sekuentziaren erabileran oinarritzen dira, organismoen arteko erlazioak ezagutarazteko (filogenetika molekularra)[22]. Kultibatu daitezkeen eta ondo ikertutako arkea espezie gehienak bi filum nagusi hauetakoak dira: "Euryarchaeota" eta "Thermoproteota" (lehen Crenarchaeota)[23]. Beste talde batzuk sortu dira behin-behinekoz, hala nola Nanoarchaeum equitans espezie berezia, 2003an aurkitu zena eta bere filum propioa duena, "Nanoarchaeota". Beste filum bat ere proposatu da, "Korarchaeota". Ohikoak ez diren espezie termofiloen talde txiki bat du, bi filum nagusien ezaugarriak partekatzen dituztenak, baina lotura estuagoa du Thermoproteotarekin[24][25]. Antzemandako beste arkea espezie batzuk talde horietako batzuekin bakarrik daude lotuta, hala nola Archaeal Richmond Mine acidophilic nanoorganisms edo ARMAN, Micrarchaeota eta Parvarchachaeota barnean dituen nanoorganismo azidofiloak. 2006an aurkitu ziren[26], eta horietako batzuk ezagutzen diren organismo txikienak dira[27].
2011n proposatu zen TACK superfiloak, Thaumarchaeota (orain Nitrososphaerota), "Aigarchaeota", Crenarchaeota (orain Thermoproteota) eta "Korarchaeota", eukarioten jatorriarekin zerikusia zuela[28]. 2017an proposatu zen Asgard superfiloa, berriki aurkitu eta bataiatua, lotura estuagoa zuela jatorrizko eukariotarekin, eta TACKen talde ahizpa bat zela[29].
2013an, DPANN superfiloa proposatu zen, "Nanoarchaeota", "Nanohaloarchaeota", "Archaeal Richmond Mine acidophilic nanoorganisms" (ARMAN, "Mikrarchaeota" eta "Parvarchaeota") eta antzeko beste arkeo batzuk biltzeko. Arkeo-superfilo horrek gutxienez 10 lerro desberdin hartzen ditu, eta tamaina oso txikiko zelula eta genomak eta ahalmen metaboliko mugatuak dituzten organismoak hartzen ditu. Beraz, DPANNko kide asko beste organismo batzuekiko elkarrekintza sinbiotikoen mende egon daitezke nahitaez, baita parasito berriak ere. Hala ere, beste azterketa filogenetiko batzuetan ikusi zen DPANN ez dela talde monofiletikoa, eta adar luzeen erakarpenaren ondorioz taldekatu zirela horrela. Horrek aditzera ematen du leinu horiek guztiak "Euryarchaeota"koak direla[30][31].
Kladograma
Tom A. Williams et al. (2017)[32] eta Castelle & Banfield (2018)ren arabera[33], eta GTDB 08-RS214ren arabera (2023ko apirila)[34][35]:
| Tom A. Williams et al. 2017[32] eta Castelle & Banfield 2018[33] | 08-RS214 (2023ko apirilaren 28)[34][35] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
Espezie kontzeptua
Arkeoak espezieetan sailkatzea ere eztabaidagarria da. Ernst Mayrrek honela definitu zuen espezie bat: elkarrekin gurutzatzen diren eta ugalkortasunez bakartuta dauden organismoen taldea, baina horrek ez du ezertarako balio, arkeoak asexualki baino ez baitira ugaltzen[37].
Arkeoek geneen transferentzia horizontal handia erakusten dute lerroen artean. Ikertzaile batzuek iradokitzen dute indibiduoak antzeko populazioetan taldeka daitezkeela, oso antzekoak diren genometan, eta geneen transferentzia ezohikoa dela harreman gutxiagoko genomak dituzten zeluletara/zeluletatik, hala nola Ferroplasma generoan[38]. Bestalde, Halorubrumi buruzko azterketetan, lotura txikiagoa duten populazioekiko/horietatik egindako transferentzia genetiko esanguratsua ikusi zen, eta horrek irizpidearen aplikagarritasuna mugatzen du[39]. Ikertzaile batzuek galdetzen dute ea espezie izendapen horiek esanahi praktikoa duten[40].
Arkeoen dibertsitate genetikoari buruzko gaur egungo ezagutzak zatikatuak dira, eta, beraz, espezie-kopurua ezin da zehatz-mehatz zenbatetsi. Filum-kopuruaren zenbatespenak 18 eta 23 bitartekoak dira, eta horietatik 8k bakarrik dituzte zuzenean landu eta aztertu diren ordezkariak. Talde hipotetiko horietako asko rRNA sekuentzia bakar baten bidez ezagutzen dira, eta horrek adierazten du organismo horien arteko aniztasuna iluna dela oraindik[41]. Bakterioek kultibatu gabeko mikrobio asko ere badituzte, eta mikrobio horiek antzeko ondorioak dituzte karakterizatzeko[42].
Filumak
Filum baliagarriak
Kode Bakteriologikoaren arabera argitaratu diren filum baliagarriak hauek dira[43]:
- Nitrososphaerota
- Thermoproteota
Hautagaiak
Honako filum hauek proposatu dira, baina ez dira oraindik Kode Bakteriologikoaren arabera modu egokian argitaratu. Horietako batzuek Candidatus izaera dute:
- "Candidatus Aenigmarchaeota"
- "Candidatus Aigarchaeota"
- "Candidatus Altiarchaeota"
- "Candidatus Asgardaeota"
- "Candidatus Bathyarchaeota"
- "Candidatus Brockarchaeota"
- "Candidatus Diapherotrites"
- "Euryarchaeota"
- "Candidatus Geoarchaeota"
- "Candidatus Hadarchaeota"
- "Candidatus Hadesarchaeota"
- "Candidatus Halobacterota"
- "Candidatus Heimdallarchaeota"
- "Candidatus Helarchaeota"
- "Candidatus Huberarchaeota"
- "Candidatus Hydrothermarchaeota"
- "Candidatus Korarchaeota"
- "Candidatus Lokiarchaeia"
- "Candidatus Lokiarchaeota"
- "Candidatus Mamarchaeota"
- "Candidatus Marsarchaeota"
- "Candidatus Methanobacteriota"
- "Candidatus Micrarchaeota"
- "Candidatus Nanoarchaeota"
- "Candidatus Nanohaloarchaeota"
- "Candidatus Nezhaarchaeota"
- "Candidatus Odinarchaeota"
- "Candidatus Pacearchaeota"
- "Candidatus Parvarchaeota"
- "Candidatus Thermoplasmatota"
- "Candidatus Thorarchaeota"
- "Candidatus Undinarchaeota"
- "Candidatus Verstraetearchaeota"
- "Candidatus Woesearchaeota"
Jatorria eta eboluzioa

Lurraren adina 4.540 milioi urte ingurukoa da[46][47][48]. Froga zientifikoen arabera, bizia duela 3.500 milioi urte hasi zen Lurrean[49][50]. Lurreko lehenengo biziaren frogak hauek dira: Groenlandiako mendebaldean[51] aurkitutako 3.700 milioi urteko arroka metasedimentarioetan biogenikotzat jotako grafitoa eta Australiako mendebaldean aurkitutako 3.480 milioi urteko hareharrietan aurkitutako mikrobio-esteren fosilak[52]. 2015ean, Mendebaldeko Australian 4.100 milioi urteko arroketan aurkitu ziren materia biotikoaren hondarrak[53].
Nahiz eta zelula prokariotoetako fosil probableak duela 3.500 milioi urtekoak izan, prokariota gehienek ez dute morfologia bereizgarririk, eta forma fosilak ezin dira erabili arkeo gisa identifikatzeko[54]. Aldiz, lipido bakarreko fosil kimikoak informazio gehiago ematen dute, konposatu horiek ez baitira beste organismo batzuetan gertatzen[55]. Argitalpen batzuen arabera, duela 2.700 milioi urteko eskistoetan arkea- edo eukariota-lipidoen hondakinak daude[56], nahiz eta datu horiek zalantzan jarri diren ordutik[57]. Groenlandiako mendebaldeko arroka are zaharragoetan ere hauteman dira lipido horiek. Mota horretako hondakin zaharrenak Isuako barrutikoak dira, Lurreko sedimentu zaharrenak barne, duela 3.800 milioi urte eratuak[58]. Arkeen leinua Lurreko zaharrena izan daiteke[59].
Woesek bakterioek, arkeoek eta eukariotoek organismoen antzinako kolonia batetik goiz aldendu ziren ondorengotza-lerro bereiziak adierazten dituztela argudiatu zuen[60][61]. Aukera bat da bereizte hori zelulen eboluzioaren aurretik gertatzea[62], ohiko zelula-mintz baten gabeziak murrizketarik gabeko geneen transferentzia horizontala ahalbidetzen zuenean, eta hiru domeinuetako arbaso komunak gene-azpimultzo espezifikoak finkatzearen ondorioz sortzea. Baliteke bakterioen eta arkeoen azken arbaso komuna termofiloa izatea, eta horrek aukera ematen du tenperatura baxuenak "muturreko inguruneak" izateko arkeoentzat, eta ingurune hotzenetan bizi diren organismoak geroago agertzeko[63]. Arkeoak eta bakterioak eukariotoekin baino ahaidetasun handiagorik ez dutenez, prokariota terminoak haien arteko antzekotasun faltsua adieraz dezake[64]. Hala ere, askotan, antzekotasun estrukturalak eta funtzionalak izaten dira leinuen artean, antzinako ezaugarri partekatuen edo konbergentzia ebolutiboaren ondorioz. Antzekotasun horiei gradu esaten zaie, eta prokariotak bizi-gradutzat hartzen dira, zenbait ezaugarri bereizgarri dituztelako, hala nola, mintzei lotutako organulurik eza.
Beste domeinu batzuekin alderaketa
Taula honek hiru domeinuen arteko ezaugarriak alderatzen ditu, berdintasunak eta aldeak argiago ikus daitezen[65].
| Ezaugarria | Archaea | Bacteria | Eukaryota |
|---|---|---|---|
| Zelula mintza | Eterrekin-lotutako lipidoak | Esterrekin-lotutako lipidoak | Esterrekin-lotutako lipidoak |
| Zelula horma | Glukoproteinaa, edo S-geruza; oso gutxitan pseudopeptiglikanoa | Peptidoglikanoa, S-geruza, edo hormarik ez | Hainbat egitura |
| Gene egitura | Kromosoma zirkularrak, Eukaryotaren itzulpen eta transkripzio antzekoa | Kromosoma zirkularrak, itzulpen eta transkripzio bakuna | Kromosoma lineal ugari, baina itzulpen eta transkripzioa Archaearen antzekoa |
| Zelularen barne egitura | Mintzak mugatu gabeko organulorik[66] edo nukleorik | Mintzak mugatu gabeko organulu eta nukleorik | Mintzek mugatutako organulu eta nukleoa. |
| Metabolismoa[67] | Asko, diazotrofia barne, eta metanogenesia arkeek bakarrik egin dezakete | Asko, fotosintesia, arnasketa aerobikoa eta arnasketa anaerobikoa, hartzidura, diazotrofia eta autotrofia barne | Fotosintesia, arnasketa zelularra eta hartzidura; diazotrofiarik ez |
| Ugalketa | Ugalketa asexuala eta geneen transferentzia horizontala | Ugalketa asexuala eta geneen transferentzia horizontala | Ugalketa asexuala eta sexu ugalketa. |
| Proteinen sintesiaren iniziazioa | Metionina | Formilmetionina | Metionina |
| RNA polimerasa | Bat | Bat | Asko |
| EF-2/EF-G | Difteriaren toxinarekiko sentikorra | Difteriaren toxinari erresistentea | Difteriaren toxinarekiko sentikorra |
Arkeoak hirugarren domeinu batean banatu ziren, RNA erribosomikoaren egituran alde handiak zeudelako. rRNA 16S molekula partikularra funtsezkoa da organismo guztietan proteinak sortzeko. Funtzio hori bizitzarako hain funtsezkoa denez, rRNA 16S-an mutazioak dituzten organismoek bizitzeko aukera gutxi dute, eta horrek egonkortasun handia dakar (baina ez erabatekoa) polinukleotido horren egituran, belaunaldiz belaunaldi. 16S rRNA organismo bakoitzaren berariazko aldaketak erakusteko bezain handia da, baina azkar alderatu ahal izateko bezain txikia. 1977an, organismoen sekuentzia genetikoak aztertzen zituen mikrobiologo batek (Carl Woese) beste konparazio-metodo bat garatu zuen: RNA zatitu, eta beste organismo batzuen zatiekin alderatu. Espezieen arteko patroiak zenbat eta antzekoagoak izan, orduan eta ahaidetasun handiagoa dute[68].
Woesek rRNA konparatzeko metodo berria erabili zuen hainbat organismo sailkatzeko eta kontrastatzeko. Hainbat espezie alderatu zituen, eta edozein prokariota edo eukariota ezagunek ez bezalako RNA bat zuen metanogeno-talde bat aurkitu zuen[69]. Metanogeno horiek beste organismo batzuk baino askoz ere antzekoagoak ziren, eta, horren ondorioz, Woesek arkeoen domeinu berria proposatu zuen. Esperimentuek erakutsi zuten arkeoak eukariotoen antz handiagoa zutela prokariotena baino, nahiz eta haien egitura prokariotena baino handiagoa izan. Ondorioz, Archaeak eta Eukaryak Eukarya eta Bacteria baino arbaso komun berriago bat partekatzen zuten. Bakteria eta arbaso komun horren arteko zatiketaren ondoren garatu zen nukleoa[70].
Arkeoen propietate esklusibo bat da eterrarekin lotutako lipidoen erabilera ugaria beren zelula-mintzean. Eterreko loturak bakterioen eta eukarioten lotura esterrak baino egonkorragoak dira kimikoki, eta horrek lagundu dezake arkeo asko gai izaten zelula-mintzak presio handien eraginpean jartzen dituzten muturreko inguruneetan bizirik irauteko, hala nola beroan eta gazitasunean. Arkeoetako genomen azterketa konparatiboak ere hainbat kontserbatutako sinadura indel eta proteina molekular identifikatu ditu, eta horiek arkeo guztietan edo arkeo talde nagusietan baino ez dira agertzen[71][72][73]. Beste organismoren batean ez dagoen arkeoen beste ezaugarri bat metanogenesia da (metanoaren ekoizpen metabolikoa). Arkeo metanogenikoek funtsezko zeregina dute ekosistemetan, metanoaren oxidaziotik energia lortzen duten organismoak dauden lekuetan. Organismo horietako asko bakterioak dira, metano-iturri garrantzitsua izan ohi baitira ingurune horietan, eta eragile primarioak izan daitezke. Metanogenoek ere funtsezko zeregina dute karbonoaren zikloan, karbono organikoa metanoan deskonposatzen baitute, eta hori ere berotegi-efektuko gas nagusietako bat da[74].
Bakterioen eta arkeoen egitura biokimikoan dagoen desberdintasun hori eboluzio-prozesuen bidez azaldu dute ikertzaileek. Teorizatzen da bi domeinuak ur sakonetako iturri hidrotermal alkalinoetan sortu zirela. Gutxienez bitan, mikrobioek lipidoen biosintesia eta zelula-paretaren biokimika garatu zituzten. Azken arbaso komun unibertsala bizitza librerik gabeko organismoa izan zela iradoki da[75]. Baliteke mintz iragazkorra izatea, kate bakuneko bakterio-anfifiloz (gantz-azidoak) osatua, arkeoen kate bakuneko anfifiloak barne (isoprenoideak). Horiek gantz-azidoen mintzak egonkortu egiten dituzte itsasoko uretan; propietate horrek bakterioen eta arkeoen mintzen dibergentzia bultza dezake, "ondoren fosfolipidoen biosintesiarekin, eta, horren ondorioz, G1P eta G3P buru-taldeak sortu ziren, arkeo eta bakterio bakarrak, hurrenez hurren". Hala bada, mintzezko isoprenoideek emandako propietateek lipidoen banaketa bizitzaren jatorrian kokatzen dute"[76].
Bakterioekin harremana

Hiru domeinuen arteko harremanak funtsezkoak dira biziaren jatorria ulertzeko. Bide metaboliko gehienak, organismo baten gene gehienen objektu direnak, Archaean eta Bacterian komunak dira; genomaren adierazpenean parte hartzen duten gene gehienak, berriz, Archaeen eta Eukaryen komunak dira[77]. Prokariotetan, arkeoen zelula-egitura bakterio gram-positiboen oso antzekoa da, neurri handi batean biek geruza bikoitz lipidiko bakarra[78] dutelako eta osaera kimiko aldakorreko sakulu lodi bat (exoeskeleto bat) izaten dutelako[79]. Zenbait zuhaitz filogenetikotan, homologo prokariotoen geneen/proteinen sekuentzia desberdinetan oinarrituta, arkeoen homologoek lotura estuagoa dute bakterio gram-positiboekin. Arkepek eta bakterio gram-positiboek zenbait proteina garrantzitsutan kontserbatutako indelak ere partekatzen dituzte, hala nola Hsp70 eta glutamina sintetasa I[80]; baina gene horien filogenia domeinuen arteko gene-transferentziaren adierazgarri gisa interpretatu zen[81][82], eta baliteke ez islatzea organismoen arteko erlazioa[83].
Proposatu da arkeoak bakterio gram-positiboetatik abiatuta eboluzionatu zutela, antibiotikoek eragindako hautespen-presioaren ondorioz[78][80][84]. Ideia babesten du arkeoak bakterio gram-positiboek bereziki eragindako antibiotiko mota askorekiko erresistenteak direla ikusteak, eta antibiotiko horiek nagusiki bakterioak eta arkeoak bereizten dituzten geneetan eragiten dutela[78][80]. Proposamena da antibiotiko gram-positiboek sortutako erresistentziarekiko presio selektiboa nahikoa izan zela, azkenean, antibiotikoen diana-gene askotan aldaketa handiak eragiteko, eta andui horiek egungo Archaearen arbasoak direla[84]. Hautespen antibiotikoari edo lehiarako beste edozein presiori erantzunez, arkeoek izan duten eboluzioak ere azal lezake muturreko inguruneetara (tenperatura altuetara edo azidotasunera) nola egokitzen diren, antibiotikoak sortzen dituzten organismoetatik ihes egiteko nitxo hutsak bilatzearen ondorioz; Cavalier-Smithek antzeko iradokizuna egin zuen[85]. Proteinen egiturazko erlazioak ikertzen dituzten beste lan batzuek[86] eta bakterio gram-positiboak prokariotetan adarkatutako lehen leinuak izan daitezkeela iradokitzen duten ikerketek ere babesten dute proposamen hori[87].
Eukariotoekin harremana

Arkeoen eta eukariotoen arteko eboluzio-harremana oraindik ez dago argi. Zelularen egituran eta funtzioan aurrerago aipatzen diren antzekotasunez gain, zuhaitz genetiko askok biak biltzen dituzte[89].
Egoera zailtzen duten faktoreen artean hainbat faktore daude, adibidez eukariotoen eta Thermoproteota filumaren arteko harremana handiagoa dela "Euryarchaeota" eta Thermoproteota filumarena baino[90]. Thermotoga maritima arkeoak hainbat bakterioren geneak ditu, akaso geneen transferentzia horizontalaren ondorioz[91]. Hipotesi estandarraren arabera, eukariotoen arbasoa oso goiz bereizi zen arkeoetatik[92][93], eta eukariotak sinbiogenesiaren, arkeo baten eta mitokondrioak sortu zituen eubakterio baten fusioaren ondorioz sortu ziren; hipotesi horrek azaltzen ditu taldeen arteko antzekotasun genetikoak. Eozitoen hipotesiaren arabera, ordea, nahiko berandu sortu ziren eukariotak arkeoetatik abiatuta[94].
Lokiarchaeum ("Lokiarchaeota" izeneko proposatutako filumekoa), Ozeano Artikoko Loki Gaztelua izeneko iturri hidrotermalean 2015ean aurkitua, ordura arte ezagutzen zen eukariotoekin lotura estuena zuen arkeoa zen. Prokarioten eta eukariotoen arteko trantsizio-organismoa deitu izan zaio[95][96]. Harrezkero, "Lokiarchaeota"ren zazpi filum-ahizpa aurkitu dira (Thorarchaeota, Odinarchaeota, Heimdallarchaeota), eta guztiek supertalde berri bat osatu dute, Asgard izenekoa[97].
Oraindik aztertzen ari dira Asgardeko kideen eta eukariotoen arteko harremanaren xehetasunak[98], baina, 2020ko urtarrilean, zientzialariek jakinarazi zuten Candidatus Prometheoarchaeum syntrophicum, Asgard arkeo mota bat, lotura izan daitekeela duela bi mila milioi urteko mikroorganismo prokarioto sinple eta eukarioto konplexuen artean[99][100][101].
Morfologia

Banakako arkeoek 0,1 mikrometro (μm) eta 15 μm arteko diametroa dute, eta hainbat formatan aurkezten dira, eskuarki esfera, hagatxo, espiral edo plaka gisa[102]. Thermoproteotaren barruan beste morfologia batzuk daude, hala nola zelula lobulatuak dituzte forma irregularrean Sulfolobusen, mikrometro erdia baino zabalera txikiagoa duten orratz moduko filamentuak Thermofilumen, eta hagaxka ia erabat angeluzuzenak Thermoproteus eta Pyrobaculumen. Haloquadratum generoko arkeoak, hala nola Haloquadratum walsbyi, urmael hipergazietan bizi diren espezie lau eta karratuak dira[103]. Forma ezohiko horiek, seguru asko, zelula-hormei eta zitoeskeleto prokariotoari esker mantentzen dira. Arkeoetan beste organismo batzuetako zitoeskeletoaren osagaiekin lotutako proteinak daude[104], eta haien zeluletan harizpiak sortzen dira[105], baina, beste organismo batzuetan ez bezala, zelula-egitura horiek ez dira ondo ezagutzen[106]. Thermoplasman eta Ferroplasman, zelula-paretarik ez dagoenez, zelulek forma irregularrak dituzte eta ameben antzekoak dira[107].
Espezie batzuek 200 μm arteko zelula-agregatuak edo -harizpiak eratzen dituzte[102]. Organismo horiek biofilmetan garrantzitsuak dira[108]. Bereziki, Thermococcus coalescens zelulen agregatuak kultiboetan fusionatzen dira eta banakako zelula erraldoiak sortzen dituzte[109]. Pyrodictium generoko arkeoek zelula anitzeko kolonia oso landua sortzen dute. Kolonia horrek hodi huts luze eta finen multzoak ditu, zelulen azaletik irten eta zuhaixka baten antzeko aglomerazio trinko bat eratuz lotzen dituztenak[110]. Kanula horien funtzioa ez dago argi, baina baliteke inguruko zelulekin elikagaiak trukatu edo komunikatzea[111]. Badira kolonia multiespezifikoak, hala nola 2001ean Alemaniako urtegi batean aurkitutako "perla-lepokoaren" komunitatea. Euryarchaeota espezie berri bateko kolonia biribilak eta zurixkak 15 zentimetro luze arteko luzera izan dezaketen harizpi finetan zehar sakabanatuta daude; harizpi horiek bakterio espezie berezi batez osatuta daude[112].
Egitura eta ekintzak
Arkeoek eta bakterioek, oro har, antzeko zelula-egitura dute, baina zelularen osaerak eta antolaketak arkeoak bereizten dituzte. Bakterioek bezala, arkeoek ez dute barne-mintz eta organulurik. Bakterioak bezala, arkeoetako zelula-mintzak zelula horma batek mugatzen ditu, eta flagelo bat edo gehiago erabiliz igeri egiten dute[113]. Egiturari dagokionez, arkeoek bakterio gram-positiboen antz handiagoa dute. Gehienek mintz plasmatiko bakarra eta zelula-pareta dute, eta ez dute espazio periplasmatikorik; arau orokor horren salbuespena Ignicoccus da, mintzean lotutako besikulak dituen eta kanpoko mintz batez inguratuta dagoen periplasma bereziki handia baitu[114].
Zelula hormak eta arkaelak
Arkeo gehienek (baina ez Thermoplasmak eta Ferroplasmak) zelula-horma dute. Arkeo gehienetan, gainazaleko geruzako proteinek osatzen dute pareta, eta S geruza bat eratzen dute[115]. Zelularen kanpoaldea estaltzen duten molekula proteikoen multzo zurruna da S geruza (sare-kota bat bezala)[116]. Geruza horrek babes kimikoa eta fisikoa ematen du, eta makromolekulek zelula-mintza ukitzea eragotz dezake[117]. Bakterioek ez bezala, arkeoek ez dute peptidoglikanorik beren zelula-hormetan. Metanobakterioek badituzte zelula-paretak, pseudopeptidoglikanoa dutenak (peptidoglikano eubakterianoaren antzekoa da morfologian, funtzioan eta egitura fisikoan), baina pseudopeptidoglikanoa desberdina da egitura kimikoan[118]; ez du D-aminoazidorik eta azido N-azetilmuramikorik; azken horren ordez azido N-azetiltalosaminuronikoa erabiltzen dute[117].
Arkeoetako flageloei arkaela deritze, eta bakterio-flageloek bezala funtzionatzen dute: haien zurtoin luzeak oinarrian motor birakariek bultzatzen dituzte. Mintzean zehar protoi-gradiente batek eragiten ditu motor horiek, baina arkaelak oso desberdinak dira osaeran eta garapenean. Bi flagelo-motak arbaso desberdinetatik abiatuta garatu ziren. Bakterio-flageloak III. motako jariatze-sistemaren arbaso bera du[119][120]; arkeoetako flageloek, berriz, IV. motako bakterio-pilusetatik aurrera eboluzionatu dutela dirudi[121]. Bakterio-flageloa hutsa da eta erdiko porotik flageloaren puntaraino igotzen diren azpiunitateen bidez mihiztatzen da; araelak, berriz, oinarrian azpiunitateak gehituz sintetizatzen dira[122].
Mintzak
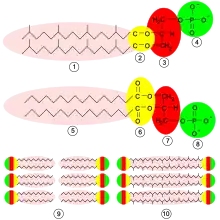
Arkeoen mintzak gainerako talde bizidunenen oso bestelakoak dira, eta horrek frogatzen du arkeoak urrutitik baino ez daudela lotuta bakterioekin eta eukariotoekin[123]. Organismo guztietan, zelula-mintzak fosfolipido izeneko molekulez osatuta daude. Molekula horiek uretan disolbatzen den zati polar bat dute (fosfato-burua), eta disolbatzen ez den zati "koipetsu" ez-polar bat (lipido-buztana). Zati horiek glizerol-frakzio batek lotzen ditu. Uretan, fosfolipidoak elkartu egiten dira, burua uretara begira dutela, eta isatsak kontrako noranzkoan. Zelula-mintzen egitura nagusia fosfolipido horien geruza bikoitza da, geruza bikoitz lipidikoa deritzona[124].
Arkeoen fosfolipidoak ez dira ohikoak lau alderditan:
- Glizerol-eter lipidoz osatutako mintzak dituzte; bakterioek eta eukariotoek, berriz, batez ere glizerol-ester lipidoz osatutako mintzak dituzte[125]. Bien arteko aldea da lipidoak glizerolaren frakzioarekin lotzen dituen lotura-mota; bi motak horiz ageri dira eskuineko irudian. Ester-lipidoetan, ester-lotura da; eter-lipidoetan, berriz, eter-lotura[126].
- Arkeoen glizerol-frakzioaren estereokimika beste organismo batzuen irudi espekularra da. Glizerolaren frakzioa bi eratara ager daiteke: bata bestearen ispilu-irudiak, enantiomero deritzenak. Eskuineko esku bat ezkerreko eskularru batean erraz sartzen ez den bezala, mota bateko enantiomeroak normalean ezin dira bestearentzat egokitutako entzimek erabili edo fabrikatu. Arkeoen fosfolipidoak sn-glizerol-1-fosfatozko eskeleto baten gainean eraikitzen dira. Eskeleto hori sn-glizerol-3-fosfatoaren, bakterio eta eukariotoetan aurkitzen den eskeleto fosfolipidikoaren, enantiomero bat da. Horrek esan nahi du arkeoek erabat desberdinak diren entzimak erabiltzen dituztela fosfolipidoak sintetizatzeko, bakterioekin eta eukariotoekin alderatuta. Entzima horiek oso goiz garatu ziren biziaren historian, eta horrek adierazten du beste bi domeinuetatik goiz bereizi zirela[123].
- Arkeoen isats lipidikoak eta beste organismo batzuenak desberdinak dira; izan ere, alboetan adarkatze ugari dituzten isoprenoide kate luzeetan oinarritzen dira, batzuetan ziklopropano edo ziklohexanozko eraztunekin[127]. Beste organismo batzuetako mintzen gantz-azidoek, aldiz, alboko adarkadurarik eta eraztunik gabeko kate zuzenak dituzte. Organismo askoren biokimikan isoprenoideek zeregin garrantzitsua duten arren, arkeoek bakarrik erabiltzen dituzte fosfolipidoak egiteko. Adarkatutako kate horiek lagungarriak izan daitezke, tenperatura altuetan, arkeoen mintzak ihesik izan ez dezan[128].
- Arkeo batzuetan, geruza bikoitz lipidikoaren ordez geruza bakarra jartzen da. Izan ere, arkeoek bi fosfolipido-molekularen isatsak molekula bakar batean fusionatzen dituzte bi buru polarrekin (bolaanfifilo bat); fusio horren ondorioz, mintzak zurrunagoak izan daitezke eta kontrako inguruneei hobeto eusten diete[129]. Adibidez, Ferroplasmako lipidoak mota horretakoak dira, eta, beraz, organismo horrek bere habitat oso azidoan bizirik irauten du[130].
Metabolismoa
Arkeoek askotariko erreakzio kimikoak dituzte metabolismoan, eta energia-iturri asko erabiltzen dituzte. Erreakzio horiek elikadura-multzotan sailkatzen dira, energia- eta karbono-iturrien arabera. Zenbait arkeok sufrea eta amoniakoa bezalako konposatu ez-organikoetatik energia lortzen dute (kimiotrofoak dira). Horien artean daude nitrifikatzaileak, metanogenoak eta metanoaren oxidatzaile anaerobioak[131]. Erreakzio horietan, konposatu batek elektroiak pasatzen ditu beste batera (erredox erreakzio batean), eta energia askatzen du zelularen jarduerak elikatzeko. Konposatu batek elektroi-emaile gisa jarduten du, eta beste batek elektroi-hartzaile gisa. Askatutako energia adenosina-trifosfatoa (ATP) kimiosmosiaren bidez sortzeko erabiltzen da, zelula eukariotoen mitokondrioan gertatzen den oinarrizko prozesu bera[132].
Beste arkeo-talde batzuek eguzki-argia erabiltzen dute energia-iturri gisa (fototrofoak dira), baina oxigenoa sortzen duen fotosintesia ez da gertatzen organismo horietan. Bizimodu guztiek oinarrizko ibilbide metaboliko asko partekatzen dituzte; adibidez, arkeoek glukolisi-forma aldatu bat (Entner-Doudoroff ibilbidea) eta azido zitrikoaren ziklo oso edo partzial bat erabiltzen dituzte[133]. Beste organismo batzuekiko antzekotasun horiek, seguru asko, biziaren historian izan dituzten jatorri goiztiarrak eta eraginkortasun maila handia islatzen dituzte[134].
| Nutrizio mota | Energia iturria | Karbono iturria | Adibideak |
|---|---|---|---|
| Fototrofoak | Eguzki-argia | Konposatu organikoak | Halobacterium |
| Litotrofoak | Konposatu inorganikoak | Konposatu organikoak edo karbonoaren finkapena | Ferroglobus, Methanobacteria edo Pyrolobus |
| Organotrofoak | Konposatu organikoak | Konposatu organikoak edo karbonoaren finkapena | Pyrococcus, Sulfolobus or Methanosarcinales |
Euryarchaeota batzuk ingurune anaerobikoetan bizi diren metanogenoak dira (metabolismoaren ondorioz metanoa sortzen duten arkeoak), hala nola zingiretan. Metabolismo mota hori oso goiz garatu zen, eta baliteke bizitza libreko lehen organismoa metanogenoa izatea. Erreakzio komun baten ondorioz, karbono dioxidoa erabiltzen da hidrogenoa oxidatzeko elektroi-hartzaile gisa. Metanogenesian, zenbait koentzima esklusibo daude, hala nola M koentzima eta metanofuranoa[135]. Metanogenoek beste konposatu organiko batzuk erabiltzen dituzte, hala nola alkoholak, azido azetikoa edo azido formikoa elektroi-hartzaile alternatibo gisa. Erreakzio horiek ohikoak dira hesteetan bizi diren arkeoetan. Arkea azetotrofoek ere azido azetikoa metanoan eta karbono dioxidoan deskonposatzen dute zuzenean. Azetotrofo horiek Methanosarzinalen ordenako arkeoak dira, eta biogasa sortzen duten mikroorganismoen komunitate garrantzitsuak dira[136].
Beste arkeo batzuek atmosferako erabiltzen dute karbono-iturri gisa, karbonoaren finkapen izeneko prozesu batean (autotrofoak dira). Prozesu horrek Calvinen zikloaren forma oso aldatu bat[137] edo beste bide metaboliko bat eskatzen du, 3-hidroxipropionatoaren/4-hidroxibutiratoaren zikloa izenekoa[138]. Thermoproteotek alderantzizko Krebsen zikloa ere erabiltzen dute; Euryarchaeotek, berriz, Wood–Ljungdahlen bidea ere erabiltzen dute[139]. Karbonoa energia ez-organikoko iturrien bidez finkatzen da. Arkeo ezagun batek ere ez du fotosintesia egiten[140] (Halobacterium da ezagutzen den arkeo fototrofo bakarra, baina fotosintesiaren ordezko prozesu bat erabiltzen du). Arkeoen energia-iturriak oso desberdinak dira: adibidez, Nitrosopumilalesek amoniakoa oxidatzen du[141][142]; Sulfolobus espezieek azido sulfhidrikoa oxidatzen dute, oxigenoa edo ioi metalikoak elektroi-hartzaile gisa erabiliz.

Arkeo fototrofoek argia erabiltzen dute energia kimikoa ATP moduan sortzeko. Halobakterioetan, argiak aktibatutako ioi-ponpek, hala nola bakteriorrodopsinak eta halorrodopsinak, gradiente ionikoak sortzen dituzte, eta ioiak ponpatzen dituzte zelularen barnealderantz eta kanpoalderantz, gradiente elektrokimiko horietan metatutako energia plasmatikoa mintzaren bidez, ATP bihurtzen du ATP sintasak. Prozesu hori fotofosforilazio modu bat da. Argiak bultzatutako ponpa horiek mintzean zehar ioiak mugitzeko duten gaitasuna, argiak proteinaren erdian lurperatutako erretinol-kofaktore baten egituran eragiten dituen aldaketen araberakoa da[143].
Genetika
Arkeoek kromosoma zirkular bakarra izaten dute[144], baina frogatu da Euryarchaea askok kromosoma horren kopia ugari dituztela[145]. 2002an, Methanoscina acetivoransek genoma arkear ezagun handiena zuen: 5.751.492 base-pare[146]. Nanoarchaeum equitans espezieko 490.885 base-pareko genoma ñimiñoa tamaina horren hamarren bat da, eta ezagutzen den arkeoen genomarik txikiena; estimazioen arabera, 537 proteina-kodetzaile baino ez lituzke[147]. DNAren zati txiki independenteak, plasmido deritzenak, aurki daitezke ere arkeotan. Plasmidoak kontaktu fisikoaren bidez transferi daitezke zelulen artean, bakterio-konjugazioaren antzeko prozesu batean[148][149].
Arkeoak genetikoki desberdinak dira bakterioetatik eta eukariotoetatik, eta edozein arkeo-genomek kodetutako proteinen %15 ere domeinuaren esklusiboak dira, nahiz eta gene esklusibo horietako gehienek ez duten funtzio ezagunik[151]. Identifikatutako funtzioa duten gainerako proteina bakarretatik, gehienak Euryarchaeotakoak dira eta metanogenesian parte hartzen dute. Arkeoek, bakterioek eta eukariotoek partekatzen dituzten proteinak zelula-funtzioko nukleo komun bat osatzen dute, eta transkripzioarekin, itzulpenarekin eta nukleotidoen metabolismoarekin du zerikusia, batez ere. Arkeoen beste ezaugarri bereizgarri batzuk funtzio erlazionatuen geneen antolaketa dira —hala nola, bide metaboliko beraren urratsak katalizatzen dituzten entzimak— eragiketa berrietan, eta alde handiak tRNA geneetan eta haien aminoazil tRNA sintetasan[152].
Arkeoetako transkripzioak eukariotoenaren antz handiagoa du bakterioena baino, eta RNA polimerasa eukariotoetan duen baliokidearen oso antzekoa da; arkeoen itzulpenean, berriz, baliokide diren bakterioen eta eukariotoen aztarnak ageri dira[153]. Arkeoek RNA polimerasa mota bakarra duten arren, transkripzioan duten egitura eta funtzioa RNA polimerasa II eukariotikoaren antzekoak dira, RNA polimerasaren eta gene baten sustatzailearen arteko lotura zuzentzen duten antzeko proteina-multzoekin (transkripzio orokorreko faktoreekin)[154], baina arkeoen beste transkripzio faktore batzuk bakterioen antzekoak dira[155]. Transkripzioaren ondorengo aldaketa eukariotoetan baino errazagoa da, arkeoetako gene gehienek ez baitute intronik, nahiz eta transferentziako RNA eta erribosomako RNA geneetan introi asko dauden[156], eta proteinak kodetzen dituzten gene gutxi batzuetan introiak egon daitezkeen[157][158].
Geneen transferentzia eta elkartrukea
Haloferax volcanii, muturreko halofiloa, zelula batetik bestera edozein norabidetan DNA transferitzeko balio duten zelulen artean zubi zitoplasmatikoak eratzen ditu[159]. Sulfolobus solfataricus[160] eta Sulfolobus acidocaldarius[161] arkeo hipertermofilikoak DNA kaltetzen duen UV irradiazioaren menpe edo bleomizinak zein mitomizinak kaltetzen dituenean, espezie bakoitzaren zelula-agregazio espezifikoa eragiten da.
Arkeoen birusak
Arkeoei bakterio edo eukariotoei eragiten ez dieten birus talde ezberdin multzo handiak eragiten die. Orain arte, DNAn oinarritutako 15-18 familia antolatu dira, baina oraindik espezie asko daude isolatu gabe eta sailkatzeko[162][163][164]. Familia horiek bi taldetan bana daitezke informalki: arkeo-espezifikoak eta kosmopolitak. Arkeo-birus espezifikoak espezie arkeoetara bakarrik zuzentzen dira, eta gaur egun 12 familia ezagutzen dira. Talde honetan, orain arte identifikatu ez diren birus-egitura bakar ugari ikusi dira, besteak beste, botila-, ziri-formako, kiribil-formako eta tanta-formako birusak. Arkeo espezie espezifikoen ugaltze-zikloak eta mekanismo genomikoak beste birus batzuetakoen antzekoak izan daitezkeen arren, berariazko ezaugarriak dituzte, eta ezaugarri horiek infektatzen dituzten zelula ostalarien morfologiagatik garatu ziren. Birusak askatzeko mekanismoak eta beste fago batzuenak desberdinak dira. Oro har, bakteriofagoek bide litiko, lisogeniko edo (oso gutxitan) bien nahasketa bati jarraitzen diote[165]. Arkeoen andui biral espezifiko gehienek harreman egonkorra dute, nolabait lisogenikoa, beren ostalariekin, eta infekzio kroniko gisa agertzen dira. Horrek zelula ostalaria hil gabe birioiak pixkanaka eta etengabe sortzea eta askatzea eskatzen du[166].
Birus horiek xehetasun handiagoz aztertu dira termofiloetan, bereziki ordena sulfolobaletan eta termoproteikoetan[167]. Duela gutxi, arkeoak infektatzen dituzten bi DNA kate-bakarreko birus-talde isolatu dira. Horietako bat Halorubrum birus polimorfikoa 1 (Pleolipoviridae) da, eta honek arkeo halofiloak kutsatzen ditu[168]. Bestea Aeropyrumen espiral formako birusa (Spiraviridae) da, ostalari termofiliko bat kutsatzen duena (90-95 °C-ko hazkunde optimoa)[169]. Nabarmentzekoa da azken birus horrek duela orain arte aurkitu den ssDNA-genomarik handiena. Birus horien aurkako defentsek RNAren interferentzia eragin dezakete, birusen geneekin lotutako DNA sekuentzia errepikakorretatik abiatuta[170][171].
Ugalketa
Arkeoak asexualki ugaltzen dira fisio bitar edo anizkoitzaren, zatikatzearen edo gemazioaren bidez; ez da mitosirik ez meiosirik gertatzen; beraz, arkeo espezie berberak itxura bat baino gehiago balu, guztiek dute material genetiko bera. Zelula-zatiketa ziklo zelular batean kontrolatzen da; zelularen kromosoma erreplikatu eta bi kromosoma alaba bereizi ondoren, zelula zatitu egiten da[172]. Sulfolobus generoan, zikloak bakterio eta eukariota sistemen antzeko ezaugarriak ditu. Kromosomak hainbat abiapuntuetatik abiatuta erreplikatzen dira (erreplikazio-jatorria), entzima eukarioto baliokideen antzeko DNA polimerasak erabiliz[173].
Euryarchaeotan, zelula-zatiketako FtsZ proteina zelularen inguruan uzkurdura-eraztun bat eratzen du, eta zelularen erdigunean zehar eraikitzen den trenkadaren osagaiak haren baliokide bakterianoen antzekoak dira. Cren-[174][175] eta Thaumarchaean[176], Cdv zelula-zatiketarako makinak antzeko funtzioa betetzen du. Makina horiek zerikusia dute ESCRT-III makina eukariotoekin; izan ere, zelulen sailkapenean duten zereginagatik ezagunagoa den arren, ikusi da zelula zatituen arteko bereizketan ere zeregin bat betetzen duela, eta, beraz, paper garrantzitsua duela zelulen banaketan[177].
Bai bakterioek bai eukariotoek, baina ez arkeoek, esporak sortzen dituzte[178]. Haloarchaeako espezie batzuek aldaketa fenotipikoa izaten dute, eta hainbat zelula-mota bezala hazten dira, pareta lodietako egiturak barne. Egitura horiek talka osmotikoarekiko erresistenteak dira, eta arkeoek gatz-kontzentrazio txikitan irauten dute uretan, baina ezin dira berriro sortu[179].
Portaera
Komunikazioa
Hasieran arkeoek quorum sensingik ez zutela uste bazen ere, duela gutxi egindako ikerketek frogatu dute espezie batzuk gai direla mekanismo horren bidez komunikazio gurutzatua ezartzeko. Beste ikerketa batzuek frogatu dute arkeoen eta bakterioen arteko elkarreragin sintrofikoa biofilmen hazkundean. Arkeoetan quoruma detektatzeko ikerketa mugatua den arren, zenbait ikerketak LuxR proteinak aurkitu dituzte espezie arkeoetan, LuxR bakterioekin antzekotasunak dituztenak, eta, azken batean, dentsitate handiko komunikazioan erabiltzen diren molekula txikiak detektatzeko aukera ematen dute. Bakterioen antzera, Archaeako luxR-ak AHLekin (laktonak) eta ez-AHL estekatzaileekin bat egiten dutela frogatu dute, eta hori oso garrantzitsua da espezie barruko, espezie arteko eta domeinu arteko komunikazioa egiteko, quoruma detektatuz[180].
Ekologia
Habitata

Arkeoak habitat-sorta zabal batean daude, eta, orain, ekosistema globalen zati garrantzitsu bat direla onartzen da[181], eta ozeanoetako zelula mikrobiarren %20 inguru izan daitezke[182]. Hala ere, aurkitutako lehen arkeoak estremofiloak ziren. Izan ere, arkeo batzuek tenperatura altuetan irauten dute, askotan 100 °C-tik (212 °F) gora, hala nola geyserretan, fumarola beltzetan eta petrolio-putzuetan daudenak. Beste habitat arrunt batzuk oso hotzak eta ur oso gazia, azidoa edo alkalinoa dira, baina arkeoen artean baldintza leunetan hazten diren mesofiloak daude, zingira eta paduretan, hondakin-uretan, ozeanoetan, animalien heste-traktuan eta lurzoruetan. Rhizobacteriak bezala, arkeoak ere landareen hazkundea sustatzeko iturritzat hartzen dira orain[183].
Arkeo estremofiloak lau talde fisiologiko nagusitakoak dira. Halofiloak, termofiloak, alkalofiloak eta azidofiloak dira[184]. Talde horiek ez dira ez zehatzak ez filum batekoak, ez eta elkar baztertzaileak ere, arkeo batzuk talde batekoak baino gehiagokoak baitira. Hala ere, sailkapenerako abiapuntu erabilgarriak dira[185].
Halofiloak, Halobacterium generoa barne, oso ingurune gazietan bizi dira, laku gazietan, esaterako, eta bakterio-homologoak %20-25 baino gazitasun handiagoetan bizi dira. Termofiloak hobeto hazten dira 45 °C-tik gorako tenperaturan , iturri termalen moduan; arkea hipertermofiloak ezin hobeto hazten dira 80 °C-tik gorako tenperaturan[186]. Methanopyrus kandleri 116. anduia arkeoa edozein organismoren erregistratutako tenperaturarik altuenean ere ugal daiteke, 122 °C-tan[187].
Beste arkeo batzuk oso ingurune azido edo alkalinotan bizi dira. Adibidez, arkeo azidofilo muturrekoenetako bat Picrophilus torridus da, pH 0ean hazten dena; horrek esan nahi du 1,2 molar azido sulfurikoan aurrera egiten duela[188].
Muturreko inguruneekiko erresistentzia horrek estralurtarrek izan ditzaketen propietateei buruzko espekulazioen gune bihurtu ditu arkeoak[189]. Muturreko habitat batzuen antzekoak aurki daitezke Marten[190], eta horrek iradoki du mikrobio bideragarriak transferitu daitezkeela planeta batetik bestera meteoritoetan[191].
Orain dela gutxi, zenbait ikerketak erakutsi dutenez, ingurune mesofiloetan eta termofiloetan ez ezik, tenperatura baxuetan ere egoten dira, batzuetan kantitate handitan. Adibidez, arkeoak ohikoak dira itsaso polarrak bezalako ingurune ozeaniko hotzetan[192]. Are esanguratsuagoak dira plankton-komunitatearen (pikoplanktonaren parte gisa[193]) muturreko ez diren habitatetan munduko ozeano guztietan dauden arkeo kopuru handiak. Arkeo horiek oso kantitate handietan egon daitezkeen arren (mikroorganismoen biomasaren %40raino), espezie horietako ia bat ere ez da isolatu eta kultibo hutsean aztertu[194]. Horren ondorioz, ozeanoetako ekologian arkeoek duten zeregina ulertzea funtsezkoa da, eta, beraz, ziklo biogeokimiko globaletan duten eragin osoa aztertu gabe dago neurri handi batean[195]. Itsasoko Thermoproteota batzuk nitrifikatzeko gai dira, eta horrek iradokitzen du organismo horiek eragina izan dezaketela nitrogenoaren ziklo ozeanikoan, nahiz eta Thermoproteota ozeaniko horiek beste energia-iturri batzuk ere erabil ditzaketen[196].
Itsas hondoa estaltzen duten sedimentuetan ere arkeo ugari daude, eta horiek zelula bizidun gehienak metro bat baino gehiagoko sakoneran eratzen dituzte[197][198]. Frogatuta dago azaleko sedimentu ozeaniko guztietan (1.000-10.000 m-ko sakoneran), infekzio birikoaren eragina handiagoa dela arkeoetan bakterioetan baino, eta birusek eragindako arkeoen lisia ezabatutako mikroorganismoen guztizko biomasaren herena dela; horren ondorioz, urtean 0,3 eta 0,5 gigatonelada karbono askatzen dute mundu osoan[199].
Ziklo biokimikoetan duten papera
Hainbat habitatetan karbonoa, nitrogenoa eta sufrea bezalako elementuak birziklatzen dituzte arkeoek[200]. Arkeoek nitrogenoaren zikloaren etapa asko egiten dituzte. Horren barruan sartzen dira bai ekosistemetatik nitrogenoa ezabatzen duten erreakzioak (baita nitratoetan oinarritutako arnasketa eta desnitrifikazioa ere), bai nitrogenoa sartzen duten prozesuak (nitratoak asimilatzea eta nitrogenoa finkatzea)[201][202]. Ikertzaileek berriki jakin dute nola parte hartzen duten arkeoek amoniakoaren oxidazio-erreakzioetan. Erreakzio horiek bereziki garrantzitsuak dira ozeanoetan[203]. Arkeoak ere funtsezkoak dira lurzoruetan amoniakoa oxidatzeko. Nitritoa sortzen dute, ondoren beste mikrobio batzuek nitratoan oxidatzen dutena. Landareek eta beste organismo batzuek azken hori kontsumitzen dute[204].
Sufrearen zikloan, sufre-konposatuak oxidatuz hazten diren arkeoek elementu hori arroketatik askatzen dute, eta beste organismo batzuen esku jartzen dute. Hala ere, Sulfolobus bezalako arkeoek azido sulfurikoa sortzen dute hondakin gisa, eta organismo horiek meatze abandonatuetan hazten badira, minen drainatze azidoa eta beste ingurumen-kalte batzuk eragin ditzakete[205].
Karbonoaren zikloan, arkeo metanogenoek hidrogenoa ezabatzen dute, eta zeregin garrantzitsua dute mikroorganismo-populazioek materia organikoa deskonposatzen dutenean, ekosistema anaerobikoetan deskonposatzaile gisa jarduten baitute, hala nola sedimentuetan, zingiretan eta hondakin-urak tratatzeko instalazioetan[206].
Beste organismo batzuekin elkarrekintzak
Arkepen eta beste organismo batzuen arteko elkarrekintzak mutualistak edo komentsalistak izan ohi dira. Ez dago arkeo patogeno edo parasito ezagunen adibide argirik, baina iradoki da metanogeno-espezie batzuk ahoko infekzioekin lotuta daudela, eta Nanoarchaeum equitans beste arkeo espezie baten parasito izan daiteke, Ignicoccus hospitalisen zeluletan bakarrik bizirauten baitu eta ugaltzen baita, eta ez dirudi batere onurarik ematen dionik haren ostalariari.
Mutualismoa
Mutualismoa espezie desberdinetako indibiduoen arteko elkarreragina da, eta eragin positiboak (onuragarriak) sortzen ditu per capita ugalketan eta/edo elkarrekintza duten populazioen biziraupenean. Zelulosa digeritzen duten animalien digestio-hodiko protozooen eta arkeo metanogenikoen arteko interakzioa da mutualismoaren adibide ezagun bat, hala nola hausnarkariak eta termitak[207]. Ingurune anaerobiko horietan, protozooek landare-zelulosa deskonposatzen dute energia lortzeko. Prozesu horrek hidrogenoa askatzen du hondakin gisa, baina hidrogeno-maila handiak energia gutxiago sortzen du. Metanogenoek hidrogenoa metano bihurtzen dutenean, protozooek energia gehiago jasotzen dute[208].
Protozoo anaerobioetan, hala nola Plagiopyla frontata, Trimyema, Heterometopus eta Metopus contortus, arkeoak protozooen barruan bizi dira eta hidrogenosometan sortutako hidrogenoa kontsumitzen dute[209][210][211]. Arkeoak ere organismo handiagoekin lotzen dira. Adibidez, Cenarchaeum symbiosum itsas arkeoa Axinella mexicana belakiaren endosinbiontea da[212].
Komentsalismoa
Zenbait arkeo komentsalistak dira, eta elkarte baten onura dute, beste organismoari lagundu edo kalterik egin gabe. Esate baterako, Methanobrevibacter smithii metanogenoa da, alde handiarekin, giza floran gehien agertzen den arkeoa da, giza hesteko hamar prokariotatik bat, gutxi gorabehera[213]. Termitetan eta gizakietan, metanogeno horiek mutualistak izan daitezke eta hesteko beste mikrobio batzuekin elkarreraginean jardun dezakete digestioa errazteko[214]. Arkeoen komunitateak beste organismo batzuekin lotzen dira, hala nola koralen gainazalean[215] eta landareen sustraiak inguratzen dituen lurzoruaren eremuan (errizosfera)[216][217].
Parasitismoa
Nahiz eta arkeoek ez duten patogenoen artean erregistro historikorik, sarritan E. coli bezalako patogeno arruntenen antzeko genomekin topo egiten dute[218], eta horrek lotura metabolikoak eta egungo patogenoekin izandako eboluzio-historia erakusten ditu. Archaeak ez dira modu trinkoan aurkitu azterketa klinikoetan, batez ere espezie espezifikoagoetan kategorizatu ez direlako[219].
Garrantzia industrian
Arkeo estremofiloak, batez ere beroarekiko edo azidotasun eta alkalinotasun muturreko baldintzekiko erresistenteak, egoera gogor horietan funtzionatzen duten entzimen iturri dira[220][221]. Entzima horiek erabilera asko aurkitu dituzte. Adibidez, DNA polimerasa termoegonkorrek, hala nola Pyrococcus furiosusen DNA polimerasak, biologia molekularra goitik behera aldatu zuen, ikerketan polimerasaren kate-erreakzioa DNA klonatzeko teknika erraz eta azkar gisa erabiltzea ahalbidetzen baitute. Industrian, 100 °C-tik gora funtzionatzen duten beste Pyrococcus espezie batzuetako amilasa, galaktosidasa eta pullulanasak elikagaiak tenperatura altuetan prozesatzeko aukera ematen dute, hala nola esnearen ekoizpena eta laktosa gutxiko gazura[222]. Arkeo termofilo horien entzimak ere oso egonkorrak izaten dira disolbatzaile organikoetan, eta, horri esker, konposatu organikoak sintetizatzen dituzten kimika berdeko prozesu ekologikoetan erabil daitezke. Egonkortasun horrek biologia estrukturalean erabiltzea errazten du. Horren ondorioz, arkeo muturrekoetako entzima bakterianoen edo eukariotoen homologoak egitura-azterketetan erabiltzen dira maiz[223].
Entzima arkeoen aplikazio-aukerarekin alderatuta, organismoek bioteknologian duten erabilera ez da hain garatua. Arkeo metanogenikoak funtsezkoak dira hondakin-uren tratamenduan, digestio anaerobikoa egiten duten eta biogasa sortzen duten mikroorganismoen komunitatearen parte baitira[224]. Mineralen tratamenduan, arkeo azidofiloak etorkizun oparoa dute metalen erauzketarako, hala nola urrea, kobaltoa eta kobrea[225].
Arkeoak erabilgarriak izan daitezkeen antibiotikoak daude. Arkeozina horietako batzuk karakterizatu dira, baina uste da beste ehunka daudela, batez ere Haloarchaean eta Sulfolobusen. Konposatu horien egitura eta bakterio-antibiotikoena desberdinak direnez, jarduteko modu berriak izan ditzakete. Gainera, arkeoen biologia molekularrean erabiltzeko aukera ematen duten markatzaile berriak sor daitezke[226].
Oharrak
- Euskaraz gutxitan erabili da terminoa, arkeobakterio erabili delako testu gehienetan. Testuetan ikusten denean Arkea ere ager liteke, baina Arkeo terminoa hobesten da[3]
Erreferentziak
- (Ingelesez) Woese, C R; Kandler, O; Wheelis, M L. (1990-06). «Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya.» Proceedings of the National Academy of Sciences 87 (12): 4576–4579. doi:. ISSN 0027-8424. PMID 2112744. PMC PMC54159. (Noiz kontsultatua: 2024-01-08).
- Petitjean, Céline; Deschamps, Philippe; López-García, Purificación; Moreira, David. (2014-12-19). «Rooting the Domain Archaea by Phylogenomic Analysis Supports the Foundation of the New Kingdom Proteoarchaeota» Genome Biology and Evolution 7 (1): 191–204. doi:. ISSN 1759-6653. PMID 25527841. PMC PMC4316627. (Noiz kontsultatua: 2024-01-08).
- «Arkeo» Euskalterm (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Pace, Norman R.. (2006-05). «Time for a change» Nature 441 (7091): 289–289. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2024-01-08).
- (Ingelesez) Stoeckenius, W. (1981-10). «Walsby's square bacterium: fine structure of an orthogonal procaryote» Journal of Bacteriology 148 (1): 352–360. doi:. ISSN 0021-9193. PMID 7287626. PMC PMC216199. (Noiz kontsultatua: 2024-01-09).
- (Ingelesez) «Archaea» Basic Biology (Noiz kontsultatua: 2024-01-09).
- (Ingelesez) Chow, Chanelle; Padda, Kiran Preet; Puri, Akshit; Chanway, Chris P.. (2022-09-20). «An Archaic Approach to a Modern Issue: Endophytic Archaea for Sustainable Agriculture» Current Microbiology 79 (11): 322. doi:. ISSN 1432-0991. (Noiz kontsultatua: 2024-01-09).
- (Ingelesez) Bang, Corinna; Schmitz, Ruth A.. (2015-09). Narberhaus, Franz ed. «Archaea associated with human surfaces: not to be underestimated» FEMS Microbiology Reviews 39 (5): 631–648. doi:. ISSN 1574-6976. (Noiz kontsultatua: 2024-01-09).
- Moissl-Eichinger, Christine; Pausan, Manuela; Taffner, Julian; Berg, Gabriele; Bang, Corinna; Schmitz, Ruth A.. (2018-01). «Archaea Are Interactive Components of Complex Microbiomes» Trends in Microbiology 26 (1): 70–85. doi:. ISSN 0966-842X. (Noiz kontsultatua: 2024-01-09).
- (Ingelesez) Staley, James T. (2006-11-29). «The bacterial species dilemma and the genomic–phylogenetic species concept» Philosophical Transactions of the Royal Society B: Biological Sciences 361 (1475): 1899–1909. doi:. ISSN 0962-8436. PMID 17062409. PMC PMC1857736. (Noiz kontsultatua: 2024-01-09).
- Zuckerkandl, Emile; Pauling, Linus. (1965-03-01). «Molecules as documents of evolutionary history» Journal of Theoretical Biology 8 (2): 357–366. doi:. ISSN 0022-5193. (Noiz kontsultatua: 2024-01-09).
- (Ingelesez) Parks, Donovan H.; Chuvochina, Maria; Waite, David W.; Rinke, Christian; Skarshewski, Adam; Chaumeil, Pierre-Alain; Hugenholtz, Philip. (2018-11). «A standardized bacterial taxonomy based on genome phylogeny substantially revises the tree of life» Nature Biotechnology 36 (10): 996–1004. doi:. ISSN 1546-1696. (Noiz kontsultatua: 2024-01-09).
- (Ingelesez) Woese, Carl R.; Fox, George E.. (1977-11). «Phylogenetic structure of the prokaryotic domain: The primary kingdoms» Proceedings of the National Academy of Sciences 74 (11): 5088–5090. doi:. ISSN 0027-8424. PMID 270744. PMC PMC432104. (Noiz kontsultatua: 2024-01-09).
- (Ingelesez) Woese, C R; Kandler, O; Wheelis, M L. (1990-06). «Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya.» Proceedings of the National Academy of Sciences 87 (12): 4576–4579. doi:. ISSN 0027-8424. PMID 2112744. PMC PMC54159. (Noiz kontsultatua: 2024-01-09).
- (Ingelesez) Sapp, Jan. (2009-07-24). The New Foundations of Evolution: On the Tree of Life. Oxford University Press ISBN 978-0-19-988917-4. (Noiz kontsultatua: 2024-01-09).
- (Ingelesez) Magrum, Linda J.; Luehrsen, Kenneth R.; Woese, Carl R.. (1978-03-01). «Are extreme halophiles actually “bacteria”?» Journal of Molecular Evolution 11 (1): 1–8. doi:. ISSN 1432-1432. (Noiz kontsultatua: 2024-01-09).
- Stetter, K. O.. (1996). «Hyperthermophiles in the history of life» Ciba Foundation Symposium 202: 1–10; discussion 11–18. ISSN 0300-5208. PMID 9243007. (Noiz kontsultatua: 2024-01-09).
- DeLong, Edward F. (1998-12-01). «Everything in moderation: Archaea as ‘non-extremophiles’» Current Opinion in Genetics & Development 8 (6): 649–654. doi:. ISSN 0959-437X. (Noiz kontsultatua: 2024-01-09).
- (Ingelesez) Theron, J.; Cloete, T. E.. (2000-01). «Molecular Techniques for Determining Microbial Diversity and Community Structure in Natural Environments» Critical Reviews in Microbiology 26 (1): 37–57. doi:. ISSN 1040-841X. (Noiz kontsultatua: 2024-01-09).
- Schmidt, Thomas M.. (2006-09). «The maturing of microbial ecology» International Microbiology: The Official Journal of the Spanish Society for Microbiology 9 (3): 217–223. ISSN 1139-6709. PMID 17061212. (Noiz kontsultatua: 2024-01-09).
- (Ingelesez) Gevers, Dirk; Dawyndt, Peter; Vandamme, Peter; Willems, Anne; Vancanneyt, Marc; Swings, Jean; De Vos, Paul. (2006-11-29). «Stepping stones towards a new prokaryotic taxonomy» Philosophical Transactions of the Royal Society B: Biological Sciences 361 (1475): 1911–1916. doi:. ISSN 0962-8436. PMID 17062410. PMC PMC1764938. (Noiz kontsultatua: 2024-01-10).
- Robertson, Charles E; Harris, J Kirk; Spear, John R; Pace, Norman R. (2005-12-01). «Phylogenetic diversity and ecology of environmental Archaea» Current Opinion in Microbiology 8 (6): 638–642. doi:. ISSN 1369-5274. (Noiz kontsultatua: 2024-01-10).
- Huber, Harald; Hohn, Michael J.; Rachel, Reinhard; Fuchs, Tanja; Wimmer, Verena C.; Stetter, Karl O.. (2002-05-01). «A new phylum of Archaea represented by a nanosized hyperthermophilic symbiont» Nature 417: 63–67. doi:. ISSN 0028-0836. (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Barns, S M; Delwiche, C F; Palmer, J D; Pace, N R. (1996-08-20). «Perspectives on archaeal diversity, thermophily and monophyly from environmental rRNA sequences.» Proceedings of the National Academy of Sciences 93 (17): 9188–9193. doi:. ISSN 0027-8424. PMID 8799176. PMC PMC38617. (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Elkins, James G.; Podar, Mircea; Graham, David E.; Makarova, Kira S.; Wolf, Yuri; Randau, Lennart; Hedlund, Brian P.; Brochier-Armanet, Céline et al.. (2008-06-10). «A korarchaeal genome reveals insights into the evolution of the Archaea» Proceedings of the National Academy of Sciences 105 (23): 8102–8107. doi:. ISSN 0027-8424. PMID 18535141. PMC PMC2430366. (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Baker, Brett J.; Tyson, Gene W.; Webb, Richard I.; Flanagan, Judith; Hugenholtz, Philip; Allen, Eric E.; Banfield, Jillian F.. (2006-12-22). «Lineages of Acidophilic Archaea Revealed by Community Genomic Analysis» Science 314 (5807): 1933–1935. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Baker, Brett J.; Comolli, Luis R.; Dick, Gregory J.; Hauser, Loren J.; Hyatt, Doug; Dill, Brian D.; Land, Miriam L.; VerBerkmoes, Nathan C. et al.. (2010-05-11). «Enigmatic, ultrasmall, uncultivated Archaea» Proceedings of the National Academy of Sciences 107 (19): 8806–8811. doi:. ISSN 0027-8424. PMID 20421484. PMC PMC2889320. (Noiz kontsultatua: 2024-01-10).
- Guy, Lionel; Ettema, Thijs J.G.. (2011-12). «The archaeal ‘TACK’ superphylum and the origin of eukaryotes» Trends in Microbiology 19 (12): 580–587. doi:. ISSN 0966-842X. (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Zaremba-Niedzwiedzka, Katarzyna; Caceres, Eva F.; Saw, Jimmy H.; Bäckström, Disa; Juzokaite, Lina; Vancaester, Emmelien; Seitz, Kiley W.; Anantharaman, Karthik et al.. (2017-01). «Asgard archaea illuminate the origin of eukaryotic cellular complexity» Nature 541 (7637): 353–358. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2024-01-10).
- academic.oup.com (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Petitjean, Céline; Deschamps, Philippe; López-García, Purificación; Moreira, David. (2015-01). «Rooting the Domain Archaea by Phylogenomic Analysis Supports the Foundation of the New Kingdom Proteoarchaeota» Genome Biology and Evolution 7 (1): 191–204. doi:. ISSN 1759-6653. PMID 25527841. PMC PMC4316627. (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Williams, Tom A.; Szöllősi, Gergely J.; Spang, Anja; Foster, Peter G.; Heaps, Sarah E.; Boussau, Bastien; Ettema, Thijs J. G.; Embley, T. Martin. (2017-06-06). «Integrative modeling of gene and genome evolution roots the archaeal tree of life» Proceedings of the National Academy of Sciences 114 (23) doi:. ISSN 0027-8424. PMID 28533395. PMC PMC5468678. (Noiz kontsultatua: 2024-01-10).
- Castelle, Cindy J.; Banfield, Jillian F.. (2018-03). «Major New Microbial Groups Expand Diversity and Alter our Understanding of the Tree of Life» Cell 172 (6): 1181–1197. doi:. ISSN 0092-8674. (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) «GTDB - About» gtdb.ecogenomic.org (Noiz kontsultatua: 2024-01-10).
- «GTDB - Taxon History» gtdb.ecogenomic.org (Noiz kontsultatua: 2024-01-10).
- doi:. PMID 31015394. Bibcode: 2019NatCo..10.1822S..
- (Ingelesez) de Queiroz, Kevin. (2005-05-03). «Ernst Mayr and the modern concept of species» Proceedings of the National Academy of Sciences 102 (suppl_1): 6600–6607. doi:. ISSN 0027-8424. PMID 15851674. PMC PMC1131873. (Noiz kontsultatua: 2024-01-10).
- academic.oup.com doi:. PMID 17603112. PMC PMC2013692. (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Papke, R. Thane; Zhaxybayeva, Olga; Feil, Edward J.; Sommerfeld, Katrin; Muise, Denise; Doolittle, W. Ford. (2007-08-28). «Searching for species in haloarchaea» Proceedings of the National Academy of Sciences 104 (35): 14092–14097. doi:. ISSN 0027-8424. PMID 17715057. PMC PMC1955782. (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Kunin, Victor; Goldovsky, Leon; Darzentas, Nikos; Ouzounis, Christos A.. (2005-07-01). «The net of life: Reconstructing the microbial phylogenetic network» Genome Research 15 (7): 954–959. doi:. ISSN 1088-9051. PMID 15965028. PMC PMC1172039. (Noiz kontsultatua: 2024-01-10).
- Hugenholtz, Philip. (2002-01-29). «Exploring prokaryotic diversity in the genomic era» Genome Biology 3 (2): reviews0003.1. doi:. ISSN 1474-760X. PMID 11864374. PMC PMC139013. (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Rappé, Michael S.; Giovannoni, Stephen J.. (2003-10). «The Uncultured Microbial Majority» Annual Review of Microbiology 57 (1): 369–394. doi:. ISSN 0066-4227. (Noiz kontsultatua: 2024-01-10).
- Oren, Aharon; Garrity, George M.. (2021). «Valid publication of the names of forty-two phyla of prokaryotes» International Journal of Systematic and Evolutionary Microbiology 71 (10): 005056. doi:. ISSN 1466-5034. (Noiz kontsultatua: 2024-01-10).
- Battistuzzi, Fabia U.; Feijao, Andreia; Hedges, S. Blair. (2004-11-09). «A genomic timescale of prokaryote evolution: insights into the origin of methanogenesis, phototrophy, and the colonization of land» BMC Evolutionary Biology 4 (1): 44. doi:. ISSN 1471-2148. PMID 15535883. PMC PMC533871. (Noiz kontsultatua: 2024-01-14).
- academic.oup.com (Noiz kontsultatua: 2024-01-14).
- «Geologic Time: Age of the Earth» pubs.usgs.gov (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) crossref. «Chooser» chooser.crossref.org doi:. (Noiz kontsultatua: 2024-01-10).
- Manhes, Gérard; Allègre, Claude J.; Dupré, Bernard; Hamelin, Bruno. (1980-05-01). «Lead isotope study of basic-ultrabasic layered complexes: Speculations about the age of the earth and primitive mantle characteristics» Earth and Planetary Science Letters 47 (3): 370–382. doi:. ISSN 0012-821X. (Noiz kontsultatua: 2024-01-10).
- «The Beginnings of Life on Earth » American Scientist» web.archive.org 2017-06-06 (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Timmer, John. (2012-09-04). «3.5 billion year old organic deposits show signs of life» Ars Technica (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Ohtomo, Yoko; Kakegawa, Takeshi; Ishida, Akizumi; Nagase, Toshiro; Rosing, Minik T.. (2014-01). «Evidence for biogenic graphite in early Archaean Isua metasedimentary rocks» Nature Geoscience 7 (1): 25–28. doi:. ISSN 1752-0908. (Noiz kontsultatua: 2024-01-10).
- Noffke, Nora; Christian, Daniel; Wacey, David; Hazen, Robert M.. (2013-12). «Microbially Induced Sedimentary Structures Recording an Ancient Ecosystem in the ca. 3.48 Billion-Year-Old Dresser Formation, Pilbara, Western Australia» Astrobiology 13 (12): 1103–1124. doi:. ISSN 1531-1074. PMID 24205812. PMC PMC3870916. (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Bell, Elizabeth A.; Boehnke, Patrick; Harrison, T. Mark; Mao, Wendy L.. (2015-11-24). «Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon» Proceedings of the National Academy of Sciences 112 (47): 14518–14521. doi:. ISSN 0027-8424. PMID 26483481. PMC PMC4664351. (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Schopf, J. William. (2006-06-29). «Fossil evidence of Archaean life» Philosophical Transactions of the Royal Society B: Biological Sciences 361 (1470): 869–885. doi:. ISSN 0962-8436. PMID 16754604. PMC PMC1578735. (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Chappe, B.; Albrecht, P.; Michaelis, W.. (1982-07-02). «Polar Lipids of Archaebacteria in Sediments and Petroleums» Science 217 (4554): 65–66. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Brocks, Jochen J.; Logan, Graham A.; Buick, Roger; Summons, Roger E.. (1999-08-13). «Archean Molecular Fossils and the Early Rise of Eukaryotes» Science 285 (5430): 1033–1036. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Rasmussen, Birger; Fletcher, Ian R.; Brocks, Jochen J.; Kilburn, Matt R.. (2008-10). «Reassessing the first appearance of eukaryotes and cyanobacteria» Nature 455 (7216): 1101–1104. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2024-01-10).
- Hahn, Jürgen; Haug, Pat. (1986-05-01). «Traces of archaebacteria in ancient sediments» Systematic and Applied Microbiology 7 (2): 178–183. doi:. ISSN 0723-2020. (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Wang, Minglei; Yafremava, Liudmila S.; Caetano-Anollés, Derek; Mittenthal, Jay E.; Caetano-Anollés, Gustavo. (2007-11-01). «Reductive evolution of architectural repertoires in proteomes and the birth of the tripartite world» Genome Research 17 (11): 1572–1585. doi:. ISSN 1088-9051. PMID 17908824. PMC PMC2045140. (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Woese, Carl R.; Gupta, Ramesh. (1981-01). «Are archaebacteria merely derived ‘prokaryotes’?» Nature 289 (5793): 95–96. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Woese, Carl. (1998-06-09). «The universal ancestor» Proceedings of the National Academy of Sciences 95 (12): 6854–6859. doi:. ISSN 0027-8424. (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Wiegel, Juergen; Michael, Adams W. W.. (1998-08-27). Thermophiles: The Keys to the Molecular Evolution and the Origin of Life?. CRC Press ISBN 978-1-4822-7304-5. (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Gribaldo, Simonetta; Brochier-Armanet, Celine. (2006-06-29). «The origin and evolution of Archaea: a state of the art» Philosophical Transactions of the Royal Society B: Biological Sciences 361 (1470): 1007–1022. doi:. ISSN 0962-8436. (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Woese, C R. (1994). «There must be a prokaryote somewhere: microbiology's search for itself.» Microbiological Reviews 58 (1): 1–9. doi:. ISSN 0146-0749. (Noiz kontsultatua: 2024-01-10).
- Willey, Joanne M.; Sherwood, Linda; Woolverton, Christopher J.; Prescott, Lansing M. Microbiology. (2008). Prescott, Harley, and Klein's microbiology. New York : McGraw-Hill Higher Education ISBN 978-0-07-299291-5. (Noiz kontsultatua: 2024-01-10).
- Heimerl, Thomas; Flechsler, Jennifer; Pickl, Carolin; Heinz, Veronika; Salecker, Benjamin; Zweck, Josef; Wanner, Gerhard; Geimer, Stefan et al.. (2017-06-13). «A Complex Endomembrane System in the Archaeon Ignicoccus hospitalis Tapped by Nanoarchaeum equitans» Frontiers in Microbiology 8 doi:. ISSN 1664-302X. PMID 28659892. PMC PMC5468417. (Noiz kontsultatua: 2024-01-10).
- Jurtshuk, Peter. (1996). Baron, Samuel ed. «Bacterial Metabolism» Medical Microbiology (University of Texas Medical Branch at Galveston) ISBN 978-0-9631172-1-2. PMID 21413278. (Noiz kontsultatua: 2024-01-10).
- (Ingelesez) Howland, John L.. (2000). The Surprising Archaea: Discovering Another Domain of Life. Oxford University ISBN 978-0-19-511183-5. (Noiz kontsultatua: 2024-01-11).
- (Ingelesez) Woese, Carl R.; Fox, George E.. (1977-11). «Phylogenetic structure of the prokaryotic domain: The primary kingdoms» Proceedings of the National Academy of Sciences 74 (11): 5088–5090. doi:. ISSN 0027-8424. PMID 270744. PMC PMC432104. (Noiz kontsultatua: 2024-01-11).
- (Ingelesez) Cavicchioli, Ricardo. (2011-01). «Archaea — timeline of the third domain» Nature Reviews Microbiology 9 (1): 51–61. doi:. ISSN 1740-1534. (Noiz kontsultatua: 2024-01-11).
- (Ingelesez) Gupta, Radhey S.; Shami, Ali. (2011-02-01). «Molecular signatures for the Crenarchaeota and the Thaumarchaeota» Antonie van Leeuwenhoek 99 (2): 133–157. doi:. ISSN 1572-9699. (Noiz kontsultatua: 2024-01-11).
- Gao, Beile; Gupta, Radhey S.. (2007-03-29). «Phylogenomic analysis of proteins that are distinctive of Archaea and its main subgroups and the origin of methanogenesis» BMC Genomics 8 (1): 86. doi:. ISSN 1471-2164. PMID 17394648. PMC PMC1852104. (Noiz kontsultatua: 2024-01-11).
- Gupta, Radhey S.; Naushad, Sohail; Baker, Sheridan. (2015). «Phylogenomic analyses and molecular signatures for the class Halobacteria and its two major clades: a proposal for division of the class Halobacteria into an emended order Halobacteriales and two new orders, Haloferacales ord. nov. and Natrialbales ord. nov., containing the novel families Haloferacaceae fam. nov. and Natrialbaceae fam. nov.» International Journal of Systematic and Evolutionary Microbiology 65 (Pt_3): 1050–1069. doi:. ISSN 1466-5034. (Noiz kontsultatua: 2024-01-11).
- Deppenmeier, Uwe. (2002-01-01). «The unique biochemistry of methanogenesis» Progress in Nucleic Acid Research and Molecular Biology (Academic Press) 71: 223–283. doi:. (Noiz kontsultatua: 2024-01-11).
- (Ingelesez) Martin, William; Russell, Michael J.. (2003-01-29). Allen, J. F. ed. «On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells» Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences 358 (1429): 59–85. doi:. ISSN 0962-8436. PMID 12594918. PMC PMC1693102. (Noiz kontsultatua: 2024-01-12).
- (Ingelesez) Jordan, Sean F.; Nee, Eloise; Lane, Nick. (2019-12-06). «Isoprenoids enhance the stability of fatty acid membranes at the emergence of life potentially leading to an early lipid divide» Interface Focus 9 (6): 20190067. doi:. ISSN 2042-8898. PMID 31641436. PMC PMC6802135. (Noiz kontsultatua: 2024-01-12).
- (Ingelesez) Koonin, Eugene V.; Mushegian, Arcady R.; Galperin, Michael Y.; Walker, D. Roland. (1997-08). «Comparison of archaeal and bacterial genomes: computer analysis of protein sequences predicts novel functions and suggests a chimeric origin for the archaea» Molecular Microbiology 25 (4): 619–637. doi:. ISSN 0950-382X. (Noiz kontsultatua: 2024-01-12).
- (Ingelesez) Gupta, Radhey S.. (1998-12). «Protein Phylogenies and Signature Sequences: A Reappraisal of Evolutionary Relationships among Archaebacteria, Eubacteria, and Eukaryotes» Microbiology and Molecular Biology Reviews 62 (4): 1435–1491. doi:. ISSN 1092-2172. PMID 9841678. PMC PMC98952. (Noiz kontsultatua: 2024-01-12).
- Koch, Arthur L.. (2003-04). «Were Gram-positive rods the first bacteria?» Trends in Microbiology 11 (4): 166–170. doi:. ISSN 0966-842X. (Noiz kontsultatua: 2024-01-12).
- (Ingelesez) Gupta, Radhey S.. (1998-08). «What are archaebacteria: life's third domain or monoderm prokaryotes related to Gram‐positive bacteria? A new proposal for the classification of prokaryotic organisms» Molecular Microbiology 29 (3): 695–707. doi:. ISSN 0950-382X. (Noiz kontsultatua: 2024-01-12).
- (Ingelesez) Gogarten, J. Peter. (1994-11-01). «Which is the most conserved group of proteins? Homology-orthology, paralogy, xenology, and the fusion of independent lineages» Journal of Molecular Evolution 39 (5): 541–543. doi:. ISSN 1432-1432. (Noiz kontsultatua: 2024-01-12).
- (Ingelesez) Brown, J.R.; Masuchi, Y.; Robb, F.T.; Doolittlel, W.F.. (1994-06-01). «Evolutionary relationships of bacterial and archaeal glutamine synthetase genes» Journal of Molecular Evolution 38 (6): 566–576. doi:. ISSN 1432-1432. (Noiz kontsultatua: 2024-01-12).
- (Ingelesez) Katz, Laura A.. (2015-09-26). «Recent events dominate interdomain lateral gene transfers between prokaryotes and eukaryotes and, with the exception of endosymbiotic gene transfers, few ancient transfer events persist» Philosophical Transactions of the Royal Society B: Biological Sciences 370 (1678): 20140324. doi:. ISSN 0962-8436. PMID 26323756. PMC PMC4571564. (Noiz kontsultatua: 2024-01-12).
- (Ingelesez) Gupta, Radhey S.. (2000-01). «The Natural Evolutionary Relationships among Prokaryotes» Critical Reviews in Microbiology 26 (2): 111–131. doi:. ISSN 1040-841X. (Noiz kontsultatua: 2024-01-13).
- Cavalier-Smith, T. (2002). «The neomuran origin of archaebacteria, the negibacterial root of the universal tree and bacterial megaclassification.» International Journal of Systematic and Evolutionary Microbiology 52 (1): 7–76. doi:. ISSN 1466-5034. (Noiz kontsultatua: 2024-01-13).
- Valas, Ruben E.; Bourne, Philip E.. (2011-02-28). «The origin of a derived superkingdom: how a gram-positive bacterium crossed the desert to become an archaeon» Biology Direct 6 (1): 16. doi:. ISSN 1745-6150. PMID 21356104. PMC PMC3056875. (Noiz kontsultatua: 2024-01-13).
- Skophammer, Ryan G.; Herbold, Craig W.; Rivera, Maria C.; Servin, Jacqueline A.; Lake, James A.. (2006-06-26). «Evidence that the Root of the Tree of Life Is Not within the Archaea» Molecular Biology and Evolution 23 (9): 1648–1651. doi:. ISSN 1537-1719. (Noiz kontsultatua: 2024-01-13).
- (Ingelesez) Gargaud, Muriel; López-Garcìa, Purificación; Martin, Hervé. (2011-01-06). Origins and Evolution of Life: An Astrobiological Perspective. Cambridge University Press ISBN 978-1-139-49459-5. (Noiz kontsultatua: 2024-01-13).
- (Ingelesez) Eme, Laura; Spang, Anja; Lombard, Jonathan; Stairs, Courtney W.; Ettema, Thijs J. G.. (2017-12). «Archaea and the origin of eukaryotes» Nature Reviews Microbiology 15 (12): 711–723. doi:. ISSN 1740-1534. (Noiz kontsultatua: 2024-01-13).
- (Ingelesez) Lake, James A.. (1988-01). «Origin of the eukaryotic nucleus determined by rate-invariant analysis of rRNA sequences» Nature 331 (6152): 184–186. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2024-01-13).
- (Ingelesez) Nelson, Karen E.; Clayton, Rebecca A.; Gill, Steven R.; Gwinn, Michelle L.; Dodson, Robert J.; Haft, Daniel H.; Hickey, Erin K.; Peterson, Jeremy D. et al.. (1999-05). «Evidence for lateral gene transfer between Archaea and Bacteria from genome sequence of Thermotoga maritima» Nature 399 (6734): 323–329. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2024-01-13).
- (Ingelesez) Gouy, Manolo; Li, Wen-Hsiung. (1989-05). «Phylogenetic analysis based on rRNA sequences supports the archaebacterial rather than the eocyte tree» Nature 339 (6220): 145–147. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2024-01-13).
- Yutin, N.; Makarova, K. S.; Mekhedov, S. L.; Wolf, Y. I.; Koonin, E. V.. (2008-04-23). «The Deep Archaeal Roots of Eukaryotes» Molecular Biology and Evolution 25 (8): 1619–1630. doi:. ISSN 0737-4038. PMID 18463089. PMC PMC2464739. (Noiz kontsultatua: 2024-01-13).
- (Ingelesez) Williams, Tom A.; Foster, Peter G.; Cox, Cymon J.; Embley, T. Martin. (2013-12). «An archaeal origin of eukaryotes supports only two primary domains of life» Nature 504 (7479): 231–236. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2024-01-13).
- (Ingelesez) Zimmer, Carl. (2015-05-06). «Under the Sea, a Missing Link in the Evolution of Complex Cells» The New York Times ISSN 0362-4331. (Noiz kontsultatua: 2024-01-13).
- (Ingelesez) Spang, Anja; Saw, Jimmy H.; Jørgensen, Steffen L.; Zaremba-Niedzwiedzka, Katarzyna; Martijn, Joran; Lind, Anders E.; van Eijk, Roel; Schleper, Christa et al.. (2015-05). «Complex archaea that bridge the gap between prokaryotes and eukaryotes» Nature 521 (7551): 173–179. doi:. ISSN 1476-4687. PMID 25945739. PMC PMC4444528. (Noiz kontsultatua: 2024-01-13).
- (Ingelesez) Seitz, Kiley W.; Lazar, Cassandre S.; Hinrichs, Kai-Uwe; Teske, Andreas P.; Baker, Brett J.. (2016-07). «Genomic reconstruction of a novel, deeply branched sediment archaeal phylum with pathways for acetogenesis and sulfur reduction» The ISME Journal 10 (7): 1696–1705. doi:. ISSN 1751-7370. PMID 26824177. PMC PMC4918440. (Noiz kontsultatua: 2024-01-13).
- (Ingelesez) MacLeod, Fraser; Kindler, Gareth S.; Wong, Hon Lun; Chen, Ray; Burns, Brendan P.; MacLeod, Fraser; Kindler, Gareth S.; Wong, Hon Lun et al.. (2019). «Asgard archaea: Diversity, function, and evolutionary implications in a range of microbiomes» AIMS Microbiology 5 (1): 48–61. doi:. ISSN 2471-1888. PMID 31384702. PMC PMC6646929. (Noiz kontsultatua: 2024-01-13).
- (Ingelesez) Zimmer, Carl. (2020-01-15). «This Strange Microbe May Mark One of Life’s Great Leaps» The New York Times ISSN 0362-4331. (Noiz kontsultatua: 2024-01-13).
- (Ingelesez) Imachi, Hiroyuki; Nobu, Masaru K.; Nakahara, Nozomi; Morono, Yuki; Ogawara, Miyuki; Takaki, Yoshihiro; Takano, Yoshinori; Uematsu, Katsuyuki et al.. (2020-01). «Isolation of an archaeon at the prokaryote–eukaryote interface» Nature 577 (7791): 519–525. doi:. ISSN 1476-4687. PMID 31942073. PMC PMC7015854. (Noiz kontsultatua: 2024-01-13).
- (Ingelesez) Tirumalai, Madhan R.; Sivaraman, Raghavan V.; Kutty, Layla A.; Song, Eric L.; Fox, George E.. (2023-09-29). «Ribosomal Protein Cluster Organization in Asgard Archaea» Archaea 2023: e5512414. doi:. ISSN 1472-3646. (Noiz kontsultatua: 2024-01-13).
- Garrity, George M.; Brenner, Don J.; Krieg, Noel R.; Staley, James T.. (2005). Bergey's manual of systematic bacteriology. (Second ed. argitaraldia) Springer ISBN 978-0-387-24143-2. (Noiz kontsultatua: 2024-01-13).
- Walsby, A. E.. (1980-01-01). «A square bacterium» Nature 283: 69–71. doi:. ISSN 0028-0836. (Noiz kontsultatua: 2024-01-13).
- (Ingelesez) Hara, Futoshi; Yamashiro, Kan; Nemoto, Naoki; Ohta, Yoshinori; Yokobori, Shin-ichi; Yasunaga, Takuo; Hisanaga, Shin-ichi; Yamagishi, Akihiko. (2007-03). «An Actin Homolog of the Archaeon Thermoplasma acidophilum That Retains the Ancient Characteristics of Eukaryotic Actin» Journal of Bacteriology 189 (5): 2039–2045. doi:. ISSN 0021-9193. PMID 17189356. PMC PMC1855749. (Noiz kontsultatua: 2024-01-13).
- (Ingelesez) Trent, Jonathan D.; Kagawa, Hiromi K.; Yaoi, Takuro; Olle, Eric; Zaluzec, Nestor J.. (1997-05-13). «Chaperonin filaments: The archaeal cytoskeleton?» Proceedings of the National Academy of Sciences 94 (10): 5383–5388. doi:. ISSN 0027-8424. PMID 9144246. PMC PMC24687. (Noiz kontsultatua: 2024-01-13).
- Hixon, William G.; Searcy, Dennis G.. (1993-01-01). «Cytoskeleton in the archaebacterium Thermoplasma acidophilum? Viscosity increase in soluble extracts» Biosystems 29 (2): 151–160. doi:. ISSN 0303-2647. (Noiz kontsultatua: 2024-01-13).
- Golyshina, O V; Pivovarova, T A; Karavaiko, G I; Kondratéva, T F; Moore, E R; Abraham, W R; Lünsdorf, H; Timmis, K N et al.. (2000). «Ferroplasma acidiphilum gen. nov., sp. nov., an acidophilic, autotrophic, ferrous-iron-oxidizing, cell-wall-lacking, mesophilic member of the Ferroplasmaceae fam. nov., comprising a distinct lineage of the Archaea.» International Journal of Systematic and Evolutionary Microbiology 50 (3): 997–1006. doi:. ISSN 1466-5034. (Noiz kontsultatua: 2024-01-13).
- (Ingelesez) Hall-Stoodley, Luanne; Costerton, J. William; Stoodley, Paul. (2004-02). «Bacterial biofilms: from the Natural environment to infectious diseases» Nature Reviews Microbiology 2 (2): 95–108. doi:. ISSN 1740-1534. (Noiz kontsultatua: 2024-01-13).
- Kuwabara, Tomohiko; Minaba, Masaomi; Iwayama, Yukihiro; Inouye, Isao; Nakashima, Miwako; Marumo, Katsumi; Maruyama, Akihiko; Sugai, Akihiko et al.. (2005). «Thermococcus coalescens sp. nov., a cell-fusing hyperthermophilic archaeon from Suiyo Seamount» International Journal of Systematic and Evolutionary Microbiology 55 (6): 2507–2514. doi:. ISSN 1466-5034. (Noiz kontsultatua: 2024-01-13).
- Nickell, Stephan; Hegerl, Reiner; Baumeister, Wolfgang; Rachel, Reinhard. (2003-01-01). «Pyrodictium cannulae enter the periplasmic space but do not enter the cytoplasm, as revealed by cryo-electron tomography» Journal of Structural Biology 141 (1): 34–42. doi:. ISSN 1047-8477. (Noiz kontsultatua: 2024-01-13).
- (Ingelesez) Horn, Christian; Paulmann, Bernd; Kerlen, Gertraude; Junker, Norbert; Huber, Harald. (1999-08-15). «In Vivo Observation of Cell Division of Anaerobic Hyperthermophiles by Using a High-Intensity Dark-Field Microscope» Journal of Bacteriology 181 (16): 5114–5118. doi:. ISSN 1098-5530. PMID 10438790. PMC PMC94007. (Noiz kontsultatua: 2024-01-13).
- (Ingelesez) Rudolph, Christian; Wanner, Gerhard; Huber, Robert. (2001-05). «Natural Communities of Novel Archaea and Bacteria Growing in Cold Sulfurous Springs with a String-of-Pearls-Like Morphology» Applied and Environmental Microbiology 67 (5): 2336–2344. doi:. ISSN 0099-2240. PMID 11319120. PMC PMC92875. (Noiz kontsultatua: 2024-01-13).
- Thomas, Nikhil A.; Bardy, Sonia L.; Jarrell, Ken F.. (2001-04). «The archaeal flagellum: a different kind of prokaryotic motility structure» FEMS Microbiology Reviews 25 (2): 147–174. doi:. ISSN 1574-6976. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Rachel, Reinhard; Wyschkony, Irith; Riehl, Sabine; Huber, Harald. (NaN/NaN/NaN). «The ultrastructure of Ignicoccus: Evidence for a novel outer membrane and for intracellular vesicle budding in an archaeon» Archaea 1: 9–18. doi:. ISSN 1472-3646. PMID 15803654. PMC PMC2685547. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Sára, Margit; Sleytr, Uwe B.. (2000-02-15). «S-Layer Proteins» Journal of Bacteriology 182 (4): 859–868. doi:. ISSN 0021-9193. PMID 10648507. PMC PMC94357. (Noiz kontsultatua: 2024-01-14).
- Engelhardt, Harald; Peters, Jürgen. (1998-12-15). «Structural Research on Surface Layers: A Focus on Stability, Surface Layer Homology Domains, and Surface Layer–Cell Wall Interactions» Journal of Structural Biology 124 (2): 276–302. doi:. ISSN 1047-8477. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Kandler, O.; König, H.. (1998-04-01). «Cell wall polymers in Archaea (Archaebacteria)» Cellular and Molecular Life Sciences CMLS 54 (4): 305–308. doi:. ISSN 1420-9071. (Noiz kontsultatua: 2024-01-14).
- Howland, John L.. (2000). The surprising archaea: discovering another domain of life. Oxford University Press ISBN 978-0-19-511183-5. (Noiz kontsultatua: 2024-01-14).
- Gophna, Uri; Ron, Eliora Z.; Graur, Dan. (2003-07-17). «Bacterial type III secretion systems are ancient and evolved by multiple horizontal-transfer events» Gene 312: 151–163. doi:. ISSN 0378-1119. (Noiz kontsultatua: 2024-01-14).
- Nguyen, L.; Paulsen, I. T.; Tchieu, J.; Hueck, C. J.; Saier, M. H.. (2000-04). «Phylogenetic analyses of the constituents of Type III protein secretion systems» Journal of Molecular Microbiology and Biotechnology 2 (2): 125–144. ISSN 1464-1801. PMID 10939240. (Noiz kontsultatua: 2024-01-14).
- Ng, Sandy Y.M.; Chaban, Bonnie; Jarrell, Ken F.. (2006-09-21). «Archaeal Flagella, Bacterial Flagella and Type IV Pili: A Comparison of Genes and Posttranslational Modifications» Journal of Molecular Microbiology and Biotechnology 11 (3-5): 167–191. doi:. ISSN 1464-1801. (Noiz kontsultatua: 2024-01-14).
- Bardy, Sonia L.; Ng, Sandy Y. M.; Jarrell, Ken F.. (2003). «Prokaryotic motility structures» Microbiology 149 (2): 295–304. doi:. ISSN 1465-2080. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Koga, Yosuke; Morii, Hiroyuki. (2007-03). «Biosynthesis of Ether-Type Polar Lipids in Archaea and Evolutionary Considerations» Microbiology and Molecular Biology Reviews 71 (1): 97–120. doi:. ISSN 1092-2172. PMID 17347520. PMC PMC1847378. (Noiz kontsultatua: 2024-01-14).
- Youssefian, Sina; Rahbar, Nima; Van Dessel, Steven. (2018-05-02). «Thermal conductivity and rectification in asymmetric archaeal lipid membranes» The Journal of Chemical Physics 148 (17) doi:. ISSN 0021-9606. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) De Rosa, M; Gambacorta, A; Gliozzi, A. (1986). «Structure, biosynthesis, and physicochemical properties of archaebacterial lipids.» Microbiological Reviews 50 (1): 70–80. doi:. ISSN 0146-0749. (Noiz kontsultatua: 2024-01-14).
- Balleza, Daniel; Garcia-Arribas, Aritz B.; Sot, Jesús; Ruiz-Mirazo, Kepa; Goñi, Félix M.. (2014-09). «Ether- versus Ester-Linked Phospholipid Bilayers Containing either Linear or Branched Apolar Chains» Biophysical Journal 107 (6): 1364–1374. doi:. ISSN 0006-3495. PMID 25229144. PMC PMC4167531. (Noiz kontsultatua: 2024-01-14).
- Damsté, Jaap S.Sinninghe; Schouten, Stefan; Hopmans, Ellen C.; van Duin, Adri C.T.; Geenevasen, Jan A.J.. (2002-10). «Crenarchaeol» Journal of Lipid Research 43 (10): 1641–1651. doi:. ISSN 0022-2275. (Noiz kontsultatua: 2024-01-14).
- academic.oup.com doi:. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Hanford, Michael J.; Peeples, Tonya L.. (2002-01-01). «Archaeal tetraether lipids» Applied Biochemistry and Biotechnology 97 (1): 45–62. doi:. ISSN 1559-0291. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Macalady, Jennifer L.; Vestling, Martha M.; Baumler, David; Boekelheide, Nick; Kaspar, Charles W.; Banfield, Jillian F.. (2004-10-01). «Tetraether-linked membrane monolayers in Ferroplasma spp: a key to survival in acid» Extremophiles 8 (5): 411–419. doi:. ISSN 1433-4909. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Valentine, David L.. (2007-04). «Adaptations to energy stress dictate the ecology and evolution of the Archaea» Nature Reviews Microbiology 5 (4): 316–323. doi:. ISSN 1740-1534. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Schäfer, Günter; Engelhard, Martin; Müller, Volker. (1999-09). «Bioenergetics of the Archaea» Microbiology and Molecular Biology Reviews 63 (3): 570–620. doi:. ISSN 1092-2172. PMID 10477309. PMC PMC103747. (Noiz kontsultatua: 2024-01-14).
- Zillig, Wolfram. (1991-12-01). «Comparative biochemistry of Archaea and Bacteria» Current Opinion in Genetics & Development 1 (4): 544–551. doi:. ISSN 0959-437X. (Noiz kontsultatua: 2024-01-14).
- Romano, A. H.; Conway, T.. (1996-07-01). «Evolution of carbohydrate metabolic pathways» Research in Microbiology 147 (6): 448–455. doi:. ISSN 0923-2508. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) DiMarco, Anthony A.; Bobik, Thomas A.; Wolfe, Ralph S.. (1990-06). «UNUSUAL COENZYMES OF METHANOGENESIS» Annual Review of Biochemistry 59 (1): 355–394. doi:. ISSN 0066-4154. (Noiz kontsultatua: 2024-01-14).
- Klocke, Michael; Nettmann, Edith; Bergmann, Ingo; Mundt, Kerstin; Souidi, Khadidja; Mumme, Jan; Linke, Bernd. (2008-08-04). «Characterization of the methanogenic Archaea within two-phase biogas reactor systems operated with plant biomass» Systematic and Applied Microbiology 31 (3): 190–205. doi:. ISSN 0723-2020. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Mueller‐Cajar, Oliver; Badger, Murray Ronald. (2007-08). «New roads lead to Rubisco in archaebacteria» BioEssays 29 (8): 722–724. doi:. ISSN 0265-9247. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Berg, Ivan A.; Kockelkorn, Daniel; Buckel, Wolfgang; Fuchs, Georg. (2007-12-14). «A 3-Hydroxypropionate/4-Hydroxybutyrate Autotrophic Carbon Dioxide Assimilation Pathway in Archaea» Science 318 (5857): 1782–1786. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Thauer, Rudolf K.. (2007-12-14). «A Fifth Pathway of Carbon Fixation» Science 318 (5857): 1732–1733. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2024-01-14).
- Bryant, Donald A.; Frigaard, Niels-Ulrik. (2006-11). «Prokaryotic photosynthesis and phototrophy illuminated» Trends in Microbiology 14 (11): 488–496. doi:. ISSN 0966-842X. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Könneke, Martin; Bernhard, Anne E.; de la Torre, José R.; Walker, Christopher B.; Waterbury, John B.; Stahl, David A.. (2005-09). «Isolation of an autotrophic ammonia-oxidizing marine archaeon» Nature 437 (7058): 543–546. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Francis, Christopher A.; Beman, J. Michael; Kuypers, Marcel M. M.. (2007-05). «New processes and players in the nitrogen cycle: the microbial ecology of anaerobic and archaeal ammonia oxidation» The ISME Journal 1 (1): 19–27. doi:. ISSN 1751-7370. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Lanyi, Janos K.. (2004-03-01). «Bacteriorhodopsin» Annual Review of Physiology 66 (1): 665–688. doi:. ISSN 0066-4278. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Allers, Thorsten; Mevarech, Moshe. (2005-01). «Archaeal genetics — the third way» Nature Reviews Genetics 6 (1): 58–73. doi:. ISSN 1471-0064. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Hildenbrand, Catherina; Stock, Tilmann; Lange, Christian; Rother, Michael; Soppa, Jörg. (2011-02). «Genome Copy Numbers and Gene Conversion in Methanogenic Archaea» Journal of Bacteriology 193 (3): 734–743. doi:. ISSN 0021-9193. PMID 21097629. PMC PMC3021236. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Galagan, James E.; Nusbaum, Chad; Roy, Alice; Endrizzi, Matthew G.; Macdonald, Pendexter; FitzHugh, Will; Calvo, Sarah; Engels, Reinhard et al.. (2002-04-01). «The Genome of M. acetivorans Reveals Extensive Metabolic and Physiological Diversity» Genome Research 12 (4): 532–542. doi:. ISSN 1088-9051. PMID 11932238. PMC PMC187521. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Waters, Elizabeth; Hohn, Michael J.; Ahel, Ivan; Graham, David E.; Adams, Mark D.; Barnstead, Mary; Beeson, Karen Y.; Bibbs, Lisa et al.. (2003-10-28). «The genome of Nanoarchaeum equitans: Insights into early archaeal evolution and derived parasitism» Proceedings of the National Academy of Sciences 100 (22): 12984–12988. doi:. ISSN 0027-8424. PMID 14566062. PMC PMC240731. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Schleper, C; Holz, I; Janekovic, D; Murphy, J; Zillig, W. (1995-08). «A multicopy plasmid of the extremely thermophilic archaeon Sulfolobus effects its transfer to recipients by mating» Journal of Bacteriology 177 (15): 4417–4426. doi:. ISSN 0021-9193. PMID 7635827. PMC PMC177192. (Noiz kontsultatua: 2024-01-14).
- Lipps, Georg. (2008). Plasmids: current research and future trends. Caister Academic Press ISBN 978-1-904455-35-6. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Xiang, Xiaoyu; Chen, Lanming; Huang, Xiaoxing; Luo, Yuanmin; She, Qunxin; Huang, Li. (2005-07). «Sulfolobus tengchongensis Spindle-Shaped Virus STSV1: Virus-Host Interactions and Genomic Features» Journal of Virology 79 (14): 8677–8686. doi:. ISSN 0022-538X. PMID 15994761. PMC PMC1168784. (Noiz kontsultatua: 2024-01-14).
- Graham, D. E.. (2000-03-28). «An archaeal genomic signature» Proceedings of the National Academy of Sciences 97 (7): 3304–3308. doi:. PMID 10716711. PMC PMC16234. (Noiz kontsultatua: 2024-01-14).
- Gaasterland, Terry. (1999-10-01). «Archaeal genomics» Current Opinion in Microbiology 2 (5): 542–547. doi:. ISSN 1369-5274. (Noiz kontsultatua: 2024-01-14).
- Dennis, Patrick P. (1997-06). «Ancient Ciphers: Translation in Archaea» Cell 89 (7): 1007–1010. doi:. ISSN 0092-8674. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Werner, Finn. (2007-09). «Structure and function of archaeal RNA polymerases» Molecular Microbiology 65 (6): 1395–1404. doi:. ISSN 0950-382X. (Noiz kontsultatua: 2024-01-14).
- Aravind, L. (1999-12-01). «DNA-binding proteins and evolution of transcription regulation in the archaea» Nucleic Acids Research 27 (23): 4658–4670. doi:. ISSN 1362-4962. PMID 10556324. PMC PMC148756. (Noiz kontsultatua: 2024-01-14).
- Lykke-Andersen, J. (1997-09). «Archaeal introns: splicing, intercellular mobility and evolution» Trends in Biochemical Sciences 22 (9): 326–331. doi:. ISSN 0968-0004. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Watanabe, Yoh-ichi; Yokobori, Shin-ichi; Inaba, Toshiro; Yamagishi, Akihiko; Oshima, Tairo; Kawarabayasi, Yutaka; Kikuchi, Hisasi; Kita, Kiyoshi. (2002-01-02). «Introns in protein‐coding genes in Archaea» FEBS Letters 510 (1-2): 27–30. doi:. ISSN 0014-5793. (Noiz kontsultatua: 2024-01-14).
- Yoshinari, Shigeo; Itoh, Takashi; Hallam, Steven J.; DeLong, Edward F.; Yokobori, Shin-ichi; Yamagishi, Akihiko; Oshima, Tairo; Kita, Kiyoshi et al.. (2006-08-04). «Archaeal pre-mRNA splicing: A connection to hetero-oligomeric splicing endonuclease» Biochemical and Biophysical Research Communications 346 (3): 1024–1032. doi:. ISSN 0006-291X. (Noiz kontsultatua: 2024-01-14).
- (Ingelesez) Rosenshine, Ilan; Tchelet, Ronen; Mevarech, Moshe. (1989-09-22). «The Mechanism of DNA Transfer in the Mating System of an Archaebacterium» Science 245 (4924): 1387–1389. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2024-01-15).
- (Ingelesez) Fröls, Sabrina; Ajon, Malgorzata; Wagner, Michaela; Teichmann, Daniela; Zolghadr, Behnam; Folea, Mihaela; Boekema, Egbert J.; Driessen, Arnold J. M. et al.. (2008-11). «UV‐inducible cellular aggregation of the hyperthermophilic archaeon Sulfolobus solfataricus is mediated by pili formation» Molecular Microbiology 70 (4): 938–952. doi:. ISSN 0950-382X. (Noiz kontsultatua: 2024-01-15).
- (Ingelesez) Ajon, Małgorzata; Fröls, Sabrina; van Wolferen, Marleen; Stoecker, Kilian; Teichmann, Daniela; Driessen, Arnold J. M.; Grogan, Dennis W.; Albers, Sonja-Verena et al.. (2011-11). «UV-inducible DNA exchange in hyperthermophilic archaea mediated by type IV pili: UV-inducible DNA exchange in hyperthermophilic archaea» Molecular Microbiology 82 (4): 807–817. doi:. (Noiz kontsultatua: 2024-01-15).
- Blohs, Marcus; Moissl-Eichinger, Christine; Mahnert, Alexander; Spang, Anja; Dombrowski, Nina; Krupovic, Mart; Klingl, Andreas. (2019-01-01). Schmidt, Thomas M. ed. «Archaea – An Introduction» Encyclopedia of Microbiology (Fourth Edition) (Academic Press): 243–252. doi:. ISBN 978-0-12-811737-8. (Noiz kontsultatua: 2024-01-15).
- Krupovic, Mart; Cvirkaite-Krupovic, Virginija; Iranzo, Jaime; Prangishvili, David; Koonin, Eugene V.. (2018-01-15). «Viruses of archaea: Structural, functional, environmental and evolutionary genomics» Virus Research 244: 181–193. doi:. ISSN 0168-1702. PMID 29175107. PMC PMC5801132. (Noiz kontsultatua: 2024-01-15).
- Pietilä, Maija K.; Demina, Tatiana A.; Atanasova, Nina S.; Oksanen, Hanna M.; Bamford, Dennis H.. (2014-06). «Archaeal viruses and bacteriophages: comparisons and contrasts» Trends in Microbiology 22 (6): 334–344. doi:. ISSN 0966-842X. (Noiz kontsultatua: 2024-01-15).
- Principi, Nicola; Silvestri, Ettore; Esposito, Susanna. (2019). «Advantages and Limitations of Bacteriophages for the Treatment of Bacterial Infections» Frontiers in Pharmacology 10 doi:. ISSN 1663-9812. PMID 31139086. PMC PMC6517696. (Noiz kontsultatua: 2024-01-15).
- Prangishvili, D.. (2013-01-01). Maloy, Stanley ed. «Viruses of the Archaea» Brenner's Encyclopedia of Genetics (Second Edition) (Academic Press): 295–298. doi:. ISBN 978-0-08-096156-9. (Noiz kontsultatua: 2024-01-15).
- Prangishvili, D.; Garrett, R.A.. (2004-04-01). «Exceptionally diverse morphotypes and genomes of crenarchaeal hyperthermophilic viruses» Biochemical Society Transactions 32 (2): 204–208. doi:. ISSN 0300-5127. (Noiz kontsultatua: 2024-01-15).
- (Ingelesez) Pietilä, Maija K.; Roine, Elina; Paulin, Lars; Kalkkinen, Nisse; Bamford, Dennis H.. (2009-04). «An ssDNA virus infecting archaea: a new lineage of viruses with a membrane envelope» Molecular Microbiology 72 (2): 307–319. doi:. ISSN 0950-382X. (Noiz kontsultatua: 2024-01-15).
- (Ingelesez) Mochizuki, Tomohiro; Krupovic, Mart; Pehau-Arnaudet, Gérard; Sako, Yoshihiko; Forterre, Patrick; Prangishvili, David. (2012-08-14). «Archaeal virus with exceptional virion architecture and the largest single-stranded DNA genome» Proceedings of the National Academy of Sciences 109 (33): 13386–13391. doi:. ISSN 0027-8424. PMID 22826255. PMC PMC3421227. (Noiz kontsultatua: 2024-01-15).
- (Ingelesez) Mojica, Francisco J.M.; Díez-Villaseñor, Chc)sar; García-Martínez, Jesús; Soria, Elena. (2005-02-01). «Intervening Sequences of Regularly Spaced Prokaryotic Repeats Derive from Foreign Genetic Elements» Journal of Molecular Evolution 60 (2): 174–182. doi:. ISSN 1432-1432. (Noiz kontsultatua: 2024-01-15).
- Makarova, Kira S.; Grishin, Nick V.; Shabalina, Svetlana A.; Wolf, Yuri I.; Koonin, Eugene V.. (2006-03-16). «A putative RNA-interference-based immune system in prokaryotes: computational analysis of the predicted enzymatic machinery, functional analogies with eukaryotic RNAi, and hypothetical mechanisms of action» Biology Direct 1 (1): 7. doi:. ISSN 1745-6150. PMID 16545108. PMC PMC1462988. (Noiz kontsultatua: 2024-01-15).
- (Ingelesez) Bernander, Rolf. (1998-08). «Archaea and the cell cycle» Molecular Microbiology 29 (4): 955–961. doi:. ISSN 0950-382X. (Noiz kontsultatua: 2024-01-15).
- KELMAN, L; KELMAN, Z. (2004-09). «Multiple origins of replication in archaea» Trends in Microbiology 12 (9): 399–401. doi:. ISSN 0966-842X. (Noiz kontsultatua: 2024-01-15).
- (Ingelesez) Lindås, Ann-Christin; Karlsson, Erik A.; Lindgren, Maria T.; Ettema, Thijs J. G.; Bernander, Rolf. (2008-12-02). «A unique cell division machinery in the Archaea» Proceedings of the National Academy of Sciences 105 (48): 18942–18946. doi:. ISSN 0027-8424. PMID 18987308. PMC PMC2596248. (Noiz kontsultatua: 2024-01-15).
- (Ingelesez) Samson, Rachel Y.; Obita, Takayuki; Freund, Stefan M.; Williams, Roger L.; Bell, Stephen D.. (2008-12-12). «A Role for the ESCRT System in Cell Division in Archaea» Science 322 (5908): 1710–1713. doi:. ISSN 0036-8075. PMID 19008417. PMC PMC4121953. (Noiz kontsultatua: 2024-01-15).
- (Ingelesez) Pelve, Erik A.; Lindås, Ann‐Christin; Martens‐Habbena, Willm; de la Torre, José R.; Stahl, David A.; Bernander, Rolf. (2011-11). «Cdv‐based cell division and cell cycle organization in the thaumarchaeon Nitrosopumilus maritimus» Molecular Microbiology 82 (3): 555–566. doi:. ISSN 0950-382X. (Noiz kontsultatua: 2024-01-15).
- Caspi, Yaron; Dekker, Cees. (2018). «Dividing the Archaeal Way: The Ancient Cdv Cell-Division Machinery» Frontiers in Microbiology 9 doi:. ISSN 1664-302X. PMID 29551994. PMC PMC5840170. (Noiz kontsultatua: 2024-01-15).
- (Ingelesez) Onyenwoke, Rob U.; Brill, Julia A.; Farahi, Kamyar; Wiegel, Juergen. (2004-10-01). «Sporulation genes in members of the low G+C Gram-type-positive phylogenetic branch (Firmicutes)» Archives of Microbiology 182 (2): 182–192. doi:. ISSN 1432-072X. (Noiz kontsultatua: 2024-01-15).
- (Ingelesez) Kostrikina, N. A.; Zvyagintseva, I. S.; Duda, V. I.. (1991-10-01). «Cytological peculiarities of some extremely halophilic soil archaeobacteria» Archives of Microbiology 156 (5): 344–349. doi:. ISSN 1432-072X. (Noiz kontsultatua: 2024-01-15).
- Rajput, Akanksha; Kumar, Manoj. (2017). «Computational Exploration of Putative LuxR Solos in Archaea and Their Functional Implications in Quorum Sensing» Frontiers in Microbiology 8 doi:. ISSN 1664-302X. PMID 28515720. PMC PMC5413776. (Noiz kontsultatua: 2024-01-15).
- DeLong, Edward F. (1998-12-01). «Everything in moderation: Archaea as ‘non-extremophiles’» Current Opinion in Genetics & Development 8 (6): 649–654. doi:. ISSN 0959-437X. (Noiz kontsultatua: 2024-01-16).
- DeLong, Edward F.; Pace, Norman R.. (2001-08-01). «Environmental Diversity of Bacteria and Archaea» Systematic Biology 50 (4): 470–478. doi:. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Chow, Chanelle; Padda, Kiran Preet; Puri, Akshit; Chanway, Chris P.. (2022-09-20). «An Archaic Approach to a Modern Issue: Endophytic Archaea for Sustainable Agriculture» Current Microbiology 79 (11): 322. doi:. ISSN 1432-0991. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Pikuta, Elena V.; Hoover, Richard B.; Tang, Jane. (2007-01). «Microbial Extremophiles at the Limits of Life» Critical Reviews in Microbiology 33 (3): 183–209. doi:. ISSN 1040-841X. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Adam, Panagiotis S.; Borrel, Guillaume; Brochier-Armanet, Céline; Gribaldo, Simonetta. (2017-11). «The growing tree of Archaea: new perspectives on their diversity, evolution and ecology» The ISME Journal 11 (11): 2407–2425. doi:. ISSN 1751-7370. PMID 28777382. PMC PMC5649171. (Noiz kontsultatua: 2024-01-16).
- Madigan, Michael T.; Martinko, John M.; Brock, Thomas D.; Madigan, Michael T.. (2006). Brock biology of microorganisms. (11. ed., international ed. argitaraldia) Pearson, Prentice Hall ISBN 978-0-13-196893-6. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Takai, Ken; Nakamura, Kentaro; Toki, Tomohiro; Tsunogai, Urumu; Miyazaki, Masayuki; Miyazaki, Junichi; Hirayama, Hisako; Nakagawa, Satoshi et al.. (2008-08-05). «Cell proliferation at 122°C and isotopically heavy CH 4 production by a hyperthermophilic methanogen under high-pressure cultivation» Proceedings of the National Academy of Sciences 105 (31): 10949–10954. doi:. ISSN 0027-8424. PMID 18664583. PMC PMC2490668. (Noiz kontsultatua: 2024-01-16).
- Ciaramella, Maria; Napoli, Alessandra; Rossi, Mosè. (2005-02). «Another extreme genome: how to live at pH 0» Trends in Microbiology 13 (2): 49–51. doi:. ISSN 0966-842X. (Noiz kontsultatua: 2024-01-16).
- Javaux, Emmanuelle J.. (2006-01-01). «Extreme life on Earth—past, present and possibly beyond» Research in Microbiology 157 (1): 37–48. doi:. ISSN 0923-2508. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Nealson, Kenneth H.. (1999-01-01). «Post-Viking Microbiology: New Approaches, New Data, New Insights» Origins of life and evolution of the biosphere 29 (1): 73–93. doi:. ISSN 1573-0875. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Davies, P. C. W.. (2007-09-28). Bock, Gregoy R. ed. «The Transfer of Viable Microorganisms Between Planets» Novartis Foundation Symposia (Wiley): 304–317. doi:. ISBN 978-0-471-96509-1. (Noiz kontsultatua: 2024-01-16).
- López-García, P. (2001-07). «Diversity of free-living prokaryotes from a deep-sea site at the Antarctic Polar Front» FEMS Microbiology Ecology 36 (2-3): 193–202. doi:. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Karner, Markus B.; DeLong, Edward F.; Karl, David M.. (2001-01). «Archaeal dominance in the mesopelagic zone of the Pacific Ocean» Nature 409 (6819): 507–510. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Giovannoni, Stephen J.; Stingl, Ulrich. (2005-09). «Molecular diversity and ecology of microbial plankton» Nature 437 (7057): 343–348. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) DeLong, Edward F.; Karl, David M.. (2005-09). «Genomic perspectives in microbial oceanography» Nature 437 (7057): 336–342. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Agogué, Hélène; Brink, Maaike; Dinasquet, Julie; Herndl, Gerhard J.. (2008-12). «Major gradients in putatively nitrifying and non-nitrifying Archaea in the deep North Atlantic» Nature 456 (7223): 788–791. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Teske, Andreas; Sørensen, Ketil B.. (2008-01). «Uncultured archaea in deep marine subsurface sediments: have we caught them all?» The ISME Journal 2 (1): 3–18. doi:. ISSN 1751-7370. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Lipp, Julius S.; Morono, Yuki; Inagaki, Fumio; Hinrichs, Kai-Uwe. (2008-08). «Significant contribution of Archaea to extant biomass in marine subsurface sediments» Nature 454 (7207): 991–994. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Danovaro, Roberto; Dell’Anno, Antonio; Corinaldesi, Cinzia; Rastelli, Eugenio; Cavicchioli, Ricardo; Krupovic, Mart; Noble, Rachel T.; Nunoura, Takuro et al.. (2016-10-07). «Virus-mediated archaeal hecatomb in the deep seafloor» Science Advances 2 (10) doi:. ISSN 2375-2548. PMID 27757416. PMC PMC5061471. (Noiz kontsultatua: 2024-01-16).
- Liu, Xiaobo; Pan, Jie; Liu, Yang; Li, Meng; Gu, Ji-Dong. (2018-10-01). «Diversity and distribution of Archaea in global estuarine ecosystems» Science of The Total Environment 637-638: 349–358. doi:. ISSN 0048-9697. (Noiz kontsultatua: 2024-01-16).
- Cabello, Purificación; Roldán, M. Dolores; Moreno-Vivián, Conrado. (2004). «Nitrate reduction and the nitrogen cycle in archaea» Microbiology 150 (11): 3527–3546. doi:. ISSN 1465-2080. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Mehta, Mausmi P.; Baross, John A.. (2006-12-15). «Nitrogen Fixation at 92°C by a Hydrothermal Vent Archaeon» Science 314 (5806): 1783–1786. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Coolen, Marco J. L.; Abbas, Ben; Van Bleijswijk, Judith; Hopmans, Ellen C.; Kuypers, Marcel M. M.; Wakeham, Stuart G.; Sinninghe Damsté, Jaap S.. (2007-04). «Putative ammonia‐oxidizing Crenarchaeota in suboxic waters of the Black Sea: a basin‐wide ecological study using 16S ribosomal and functional genes and membrane lipids» Environmental Microbiology 9 (4): 1001–1016. doi:. ISSN 1462-2912. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Leininger, S.; Urich, T.; Schloter, M.; Schwark, L.; Qi, J.; Nicol, G. W.; Prosser, J. I.; Schuster, S. C. et al.. (2006-08). «Archaea predominate among ammonia-oxidizing prokaryotes in soils» Nature 442 (7104): 806–809. doi:. ISSN 1476-4687. (Noiz kontsultatua: 2024-01-16).
- Baker, Brett J; Banfield, Jillian F. (2003-05). «Microbial communities in acid mine drainage» FEMS Microbiology Ecology 44 (2): 139–152. doi:. ISSN 0168-6496. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Schimel, Joshua. (2004-08-24). «Playing scales in the methane cycle: From microbial ecology to the globe» Proceedings of the National Academy of Sciences 101 (34): 12400–12401. doi:. ISSN 0027-8424. PMID 15314221. PMC PMC515073. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Chaban, Bonnie; Ng, Sandy Y.M; Jarrell, Ken F. (2006-02-01). «Archaeal habitats — from the extreme to the ordinary» Canadian Journal of Microbiology 52 (2): 73–116. doi:. ISSN 0008-4166. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Schink, B. (1997-06). «Energetics of syntrophic cooperation in methanogenic degradation» Microbiology and Molecular Biology Reviews 61 (2): 262–280. doi:. ISSN 1092-2172. PMID 9184013. PMC PMC232610. (Noiz kontsultatua: 2024-01-16).
- Lewis, William H.; Sendra, Kacper M.; Embley, T. Martin; Esteban, Genoveva F.. (2018). «Morphology and Phylogeny of a New Species of Anaerobic Ciliate, Trimyema finlayi n. sp., with Endosymbiotic Methanogens» Frontiers in Microbiology 9 doi:. ISSN 1664-302X. PMID 29515525. PMC PMC5826066. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Wrede, Christoph; Dreier, Anne; Kokoschka, Sebastian; Hoppert, Michael. (2012-12-27). «Archaea in Symbioses» Archaea 2012: e596846. doi:. ISSN 1472-3646. PMID 23326206. PMC PMC3544247. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Lange, Marianne; Westermann, Peter; Ahring, Birgitte Kiær. (2005-02-01). «Archaea in protozoa and metazoa» Applied Microbiology and Biotechnology 66 (5): 465–474. doi:. ISSN 1432-0614. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Preston, C M; Wu, K Y; Molinski, T F; DeLong, E F. (1996-06-25). «A psychrophilic crenarchaeon inhabits a marine sponge: Cenarchaeum symbiosum gen. nov., sp. nov.» Proceedings of the National Academy of Sciences 93 (13): 6241–6246. doi:. ISSN 0027-8424. PMID 8692799. PMC PMC39006. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Eckburg, Paul B.; Bik, Elisabeth M.; Bernstein, Charles N.; Purdom, Elizabeth; Dethlefsen, Les; Sargent, Michael; Gill, Steven R.; Nelson, Karen E. et al.. (2005-06-10). «Diversity of the Human Intestinal Microbial Flora» Science 308 (5728): 1635–1638. doi:. ISSN 0036-8075. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Samuel, Buck S.; Gordon, Jeffrey I.. (2006-06-27). «A humanized gnotobiotic mouse model of host–archaeal–bacterial mutualism» Proceedings of the National Academy of Sciences 103 (26): 10011–10016. doi:. ISSN 0027-8424. PMID 16782812. PMC PMC1479766. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Wegley, Linda; Yu, Yanan; Breitbart, Mya; Casas, Veronica; Kline, David I.; Rohwer, Forest. (2004-06-08). «Coral-associated Archaea» Marine Ecology Progress Series 273: 89–96. doi:. ISSN 0171-8630. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Chelius, M.K.; Triplett, E.W.. (2001-04-01). «The Diversity of Archaea and Bacteria in Association with the Roots of Zea mays L.» Microbial Ecology 41 (3): 252–263. doi:. ISSN 1432-184X. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Simon, Holly M.; Dodsworth, Jeremy A.; Goodman, Robert M.. (2000-10). «Crenarchaeota colonize terrestrial plant roots» Environmental Microbiology 2 (5): 495–505. doi:. ISSN 1462-2912. (Noiz kontsultatua: 2024-01-16).
- Gophna, Uri; Charlebois, Robert L.; Doolittle, W.Ford. (2004-05). «Have archaeal genes contributed to bacterial virulence?» Trends in Microbiology 12 (5): 213–219. doi:. ISSN 0966-842X. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Shiffman, Miriam E.; Charalambous, Bambos M.. (2012-07). «The search for archaeal pathogens» Reviews and Research in Medical Microbiology 23 (3): 45. doi:. ISSN 2770-3150. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Breithaupt, Holger. (2001-11). «The hunt for living gold: The search for organisms in extreme environments yields useful enzymes for industry» EMBO reports 2 (11): 968–971. doi:. ISSN 1469-221X. PMID 11713183. PMC PMC1084137. (Noiz kontsultatua: 2024-01-16).
- Egorova, Ksenia; Antranikian, Garabed. (2005-12-01). «Industrial relevance of thermophilic Archaea» Current Opinion in Microbiology 8 (6): 649–655. doi:. ISSN 1369-5274. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Synowiecki, Józef; Grzybowska, Beata; Zdziebło, Anna. (2006-04). «Sources, Properties and Suitability of New Thermostable Enzymes in Food Processing» Critical Reviews in Food Science and Nutrition 46 (3): 197–205. doi:. ISSN 1040-8398. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Jenney Jr, Francis E.; Adams, Michael W. W.. (2008-01-01). «The impact of extremophiles on structural genomics (and vice versa)» Extremophiles 12 (1): 39–50. doi:. ISSN 1433-4909. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Schiraldi, Chiara; Giuliano, Mariateresa; De Rosa, Mario. (NaN/NaN/NaN). «Perspectives on biotechnological applications of archaea» Archaea 1: 75–86. doi:. ISSN 1472-3646. PMID 15803645. PMC PMC2685559. (Noiz kontsultatua: 2024-01-16).
- (Ingelesez) Norris, P. R.; Burton, N. P.; Foulis, N. A. M.. (2000-04-01). «Acidophiles in bioreactor mineral processing» Extremophiles 4 (2): 71–76. doi:. ISSN 1431-0651. (Noiz kontsultatua: 2024-01-16).
- Blum, Paul. (2008). Archaea: new models for prokaryotic biology. Caister academic press ISBN 978-1-904455-27-1. (Noiz kontsultatua: 2024-01-16).
Ikus, gainera
Kanpo estekak
| Autoritate kontrola |
|---|