Kiel legi la taksonomion | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Troveblo de fosilioj: Malfrua kretaceo | ||||||||||||||||
  | ||||||||||||||||
| Biologia klasado | ||||||||||||||||
| ||||||||||||||||
| Konserva statuso | ||||||||||||||||
|
Konserva statuso: Fosilia | ||||||||||||||||
| ||||||||||||||||
Sinonimoj | ||||||||||||||||
| ||||||||||||||||
Aliaj Vikimediaj projektoj | ||||||||||||||||
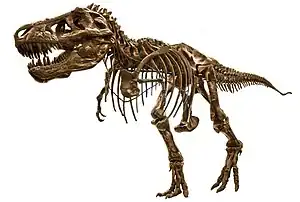
Tiranosaŭro (Tyrannosaurus = 'tirana lacerto') estas dinosaŭra genro. Ankaŭ konata kiel T. rex, la specio Tyrannosaurus rex vivis en tiu landparto, kiu nun estas okcidenta Nordameriko. Iuj sciencistoj konsideras la iom pli malnovan tarbosaŭron (Tarbosaurus bataar) de Azio dua tiranosaŭra specio dum aliaj opinias, ke tarbosaŭro estas aparta genro. La tiranosaŭro estas la dinosauro plej ofte prezentita en la populara kulturo.
Same kiel aliaj tiranosaŭredaj teropodoj, tiranosaŭro estis dupieda karnomanĝulo kun granda kranio kontraŭbalancita per longa, peza vosto. Kompare kun la grandaj, potencaj malantaŭaj kruroj, la antaŭaj kruroj de la tiranosaŭro estis malgrandaj kaj havis nur du fingrojn. Kvankam aliaj terapodoj estis same grandaj aŭ eĉ pli grandaj ol la tiranosaŭro, ĝi estis la plej granda tiranosaŭredo (pri kiu ni scias) kaj unu el la plej grandaj teraj rabobestoj, kun longeco de pli ol 12 metroj kaj pezo de elefanto.
Oni trovis en Nordameriko fosiliojn de T. rex en formacioj de la plej malfrua kretaceo; ĝi estis inter la lastaj dinosaŭroj ekzistantaj antaŭ la kretacea-terciara formorta evento. Oni jam identigis pli ol 30 ekzemplerojn de T. rex, iuj preskaŭ kompletaj, kio permesis profundan esploradon pri multaj aspektoj de ĝia biologio, inkluzive ĝian vivhistorion kaj biomeĥanikon. Ankoraŭ ne estas konsento pri la manĝkutimoj kaj potencia rapideco de T. rex.
Priskribo
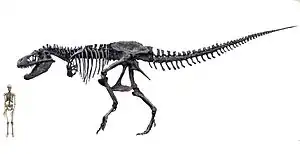
Tyrannosaurus rex estis unu el la plej grandaj teraj karnomanĝuloj iam ajn, ĉ. 12 ĝis 13 metrojn longa kaj 5 metrojn alta kiam plenkreska.[1] Pritaksoj de la pezo de tiranosaŭro multe variis dum la jaroj, de pli ol 7200 kg[2] ĝis malpli ol 4500 kg[3][4] kun la plimulto de la modernaj pritaksoj estante inter 5400 kg kaj 6800 kg.[5][6][7][8]

La plej grandaj kranioj de T. rex konataj estas 1,5m longaj. Kompare kun aliaj teropodoj, la tiranosaŭra kranio estis multe kodifita. Malantaŭe ĝi estis tre larĝa kaj antaŭe mallarĝa, kio permesis iom da binokla vidpovo. Iuj ostoj estis kunfandiĝintaj, kio malhelpis moviĝon inter ili. Grandaj truoj (fenestrae) en la kranio malpligrandigis pezon kaj donis lokojn kie povis ligiĝi muskoloj. La ostoj mem estis masivaj, same kiel la segil-dentaj dentoj kiuj, anstataŭ esti klingecaj, estis ovalaj kversekce. Kiel aliaj tiranosaŭredoj T. rex havis frapan diversdentecon, kun la antaŭaj dentoj ĉe la antaŭa parto de la supra makzelo dense pakitaj kaj D-formaj kversekce. Grandaj mordo-signoj sur la ostoj de aliaj dinosaŭroj indikas ke ĉi tiuj dentoj povis penetri solidajn ostojn. T. rex havis la plej grandan mordo-potencon de ĉiuj dinosaŭroj kaj de plimultaj bestoj entute. Oni ofte trovas frotitajn aŭ rompitajn dentojn, sed malkiel la dentoj de mamuloj, la dentoj de tiranosaŭredoj daŭre anstataŭiĝis dum la vivo de iliaj posedantoj.[1]
La kolo de T. rex havis la formon de natura S-kurbiĝo, same kiel ĉe aliaj teropodoj, sed ĝi estis mallonga kaj fortmuskola por subteni la grandan kapon. La du-fingraj antaŭkruroj estis tre malgrandaj kompare kun la korpo, sed ili estis fortmuskolaj. La malantaŭaj kruroj kontraŭe estis inter la plej longaj, proporcie al korpograndeco, de ĉiuj teropodoj. La vosto estis peza kaj longa, foje kun pli ol kvardek vertebroj, por kontraŭbalanci la grandajn kapon kaj korpon. Por kompensi la grandecon de la besto, multaj ostoj de la skeleto estis kavaj. Ĉi tio malpligrandigis la pezon de la skeleto dum ĝi retenis multon el la forteco de la ostoj.[1]
Klasado
Tiranosaŭro estas genro de la superfamilio de tiranosaŭroideoj, la familio de tiranosaŭredoj, kaj la subfamilio de tiranosaŭrenoj. Inter aliaj membroj de la tiranosaŭrena subfamilio estas la nordamerika daspletosaŭro kaj la azia tarbosaŭro,[9][10] kiuj ambaŭ ofte traktiĝis kiel sinonimoj de tiranosaŭro.[11] Oni iam kredis tiranosaŭedojn esti posteuloj de pli fruaj grandaj teropodoj kiel la megalosaŭroj kaj karnosaŭroj sed intertempe oni reklasigis ilin metante ilin kun la ĝenerale malpli grandaj celurosaŭroj.[12]
En la jaro 1955 la sovetunia paleontologo Evgeny Maleev nomis novan specion, Tyrannosaurus bataar, de Mongolio.[13] Ĝis 1965 oni renomis ĉi tiun specion Tarbosaurus bataar.[14] Spite la renomadon multaj filogenetikaj analizoj traktas Tarbosaurus bataar kiel fratinan taksonon de Tyrannosaurus rex,[10] kaj oni havis konsideris ĝin azia tiranosaŭra specio.[12][15][16] Lastatempa repriskribado de la kranio de Tarbosaurus bataar montris ke ĝi estis multe pli mallarĝa ol Tyrannosaurus rex kaj ke dum mordado la distribuo de premoj en la kranio estus ege alia, pli proksima al tiuj ĉe alioramo (Alioramus), alia azia tiranosaŭredo.[17] Parenca kladistika analizo trovis ke alioramo, ne tiranosaŭro, estis la fratina taksono de tarbosaŭro kio, se vero, sugestus ke tarbosaŭro kaj tiranosaŭro devus resti apartaj genroj.[9]
Aliaj tiranosaŭredaj fosilioj trovitaj en la samaj formacioj kiel T. rex unue klasiĝis kiel apartaj taksonoj, inter ili Aublysodon kaj Albertosaurus megagracilis,[11] la dua poste renomita Dinotyrannus megagracilis in 1995.[18] Oni nun tamen universale konsideras ĉi tiujn fosiliojn aparteni al nematuraj tiranosaŭroj.[19] Malgranda sed preskaŭ kompleto kranio de Montano, 60 cm longa, eble estas escepto tiurilate. Ĉi tiun kranion unue klasis kiel apartenanta al specio de gorgosaŭro (G. lancensis) Charles W. Gilmore en la jaro 1946,[20], sed oni poste proponis ke ĝi apartenas al nova genro nanotirano (Nanotyrannus).[21] Daŭre restas diverseco de opinioj pri tio ĉu la propono pri N. lancensis validas. Multaj paleontologoj konsideras la kranion esti tiu de nematura T. rex.[22] Klare estas malgrandaj diferencoj inter la specioj, inter ili pli granda nombro de dentoj ĉe N. lancensis, kio instigis iujn sciencistoj rekomendi apartan traktadon de la genroj ĝis plia esplorado aŭ pliaj malkovroj klarigos la situacion.[10][23]
La debato pri Manospondylus

La unua fosilia specimeno atribuata al Tyrannosaurus rex konsistas el du partaj vertebroj (de kiuj unu perdiĝis) kiujn trovis Edward Drinker Cope en 1892 kaj kiujn ili priskribis kiel Manospondylus gigas. Osborn rekonis la similecon inter M. gigas kaj T. rex jam en 1917 sed, pro la parta naturo de la vertebroj de Manospondylus, li ne povis ilin definitive sinonimigi.[24]
Debato estiĝis en junio de 2000 kiam la Instituto Black Hills trovis la tipo-lokon de M. gigas en Suda Dakoto kaj tie trovis pliajn tiranosaŭrajn ostojn. Oni decidis ke ĉi tiuj reprezentas pliajn restaĵojn de la sama unuopulo kaj ke ili estas identaj kun tiuj de T. rex. Laŭ la reguloj de la Internacia Kodo de Zoologia Nomenklaturo (IKZN), la sistemo kiu regas la sciencan nomadon de bestoj, Manospondylus gigas sekve devus havi prioritaton super Tyrannosaurus rex, ĉar ĝin oni nomis unue.[25] La Kvara Eldono de la IKZN Arkivigite je 2009-05-24 per la retarkivo Wayback Machine, kiu ekvalidis la 1-an de januaro 2000, tamen asertas ke "la uzado de jam ekzistantaj terminoj daŭru" kiam "la pli aĝa sinonimo aŭ homonimo ne uziĝas kiel valida nomo depost 1899" kaj "la pli juna sinonimo aŭ homonimo uziĝas por certa taksono, kiel ĝia supozata valida nomo, en almenaŭ 25 verkoj, publikigitaj de almenaŭ 10 aŭtoroj en la tuj antaŭaj 50 jaroj..."[26] Laŭ ĉi tiuj kondiĉoj Tyrannosaurus rex facile kvalifikiĝus kiel valida nomo kaj plej verŝajne estus konsiderita protektita nomo (nomen protectum) sub la IKZN se iam oni ĝin kontestus (kaj oni ankoraŭ ne kontestis ĝin). Tiukaze Manospondylus gigas estus juĝita forgesita nomo (nomen oblitum).[27]
Paleobiologio
Kiel ĉe ĉiuj dinosaŭroj konataj de fosilioj, multo pri la tiranosaŭra biologio, inkluzive ĝian agmanieron, koloro, ekologio, kaj fiziologio estas nekonata. Tamen la malkovrado de multaj novaj specimenoj dum la pasintaj dudek jaroj permesis iom da informita spekulativado pri kreskorapideco, seksa dimorfismo, kaj metabolismo.
Vivhistorio

La identigo de kelkaj specimenoj kiel nematuraj tiranosaŭroj permesis al sciencistoj ekscii pri ontogenezaj ŝanĝiĝoj en la specio, pritaksi la vivlongecon, kaj informiĝi pri tio kiel rapide kreskis la tiranosaŭridoj. Oni taksas ke la plej malgranda unuopulo konata (NMLK 28471, la "teropodo Jordan") pezis nur 29.9 kg, dum la plej granda (NHMF PR2081 "Sue") verŝajne pezis pli ol 5400 kg. Histologia analizado de la ostoj de T. rex montris ke NMLK 28471 havis nur 2 jarojn kiam ĝi mortis, dum "Sue" havis 28 jaroj, aĝo kiu eble estis proksima al la maksimumo por tiu specio.[8]
Histologio ankaŭ permesis ekscii la aĝon de aliaj specimenoj. Oni povas fari vivkurbiĝojn indikante la aĝojn de diversaj specimenoj sur grafo kune kun iliaj pezoj. La vivkurbiĝo de T. rex estas S-forma, kun junaj tiranosaŭroj restante sub pezo de 1800 kg ĝis kiam ili havis 14 jarojn, kiam la korpomaso pligrandiĝis rapide. Dum ĉi tiu periodo de rapida kreskado juna T. rex gajnis meze 600 kg jare dum la sekvaj kvar jaroj. Kiam la tiranosaŭro ekhavas 18 jarojn, la kurbiĝo horizontaliĝas denove, kio indikas ke la kreskado forte malrapidiĝis. Ekzemple, nur 600 kg apartigis la 28-jara "Sue" de 22-jara kanada specimeno (RMPT 81.12.1).[8] Alia lastatempa histologia studaĵo farita de aliaj sciencistoj subtenas ĉi tiujn rezultojn, trovante ke rapida kreskado ekmalrapidiĝis kiam la bestoj atingis 16 jarojn.[28] Ĉi tiu subita ŝanĝiĝo de kreskorapideco eble indikis korpan maturecon, hipotezo subtenata de la malkovro de medolaj histoj en la femuro de 16-ĝis-20-jara T. rex de Montano (MRM 1125, ankaŭ konata kiel "B-rex"). Oni trovas medolajn histojn nur en inaj birdoj dum ovproduktado, kio sugestas ke "B-rex" estis sekse matura.[29] Aliaj tiranosaŭredoj montras similajn kreskokurbiĝojn kvankam kun malpli granda kreskorapideco korespondanta al malpli granda plenkreskula grandeco.[30]
Ŝajnas ke pli ol la duono de la konataj specimenoj de T. rex mortis interne de ses jaroj post kiam ili atingis seksan maturecon, kio vidiĝas ankaŭ ĉe aliaj tiranosaŭredoj kaj ĉe iuj grandaj longvivaj birdoj kaj mamuloj hodiaŭ. Ĉi tiujn speciojn karakterizas alta procentaĵo de infanmortado kun relative malalta mortoprocentaĵo inter junuloj. La mortoprocentaĵo denove plialtiĝas post la atingo de seksa matureco, parte pro la premoj de reproduktado. Unu esploraĵo sugestas ke la rareco de junulaj fosilioj de T. rex estas parte pro malalta mortoprocentaĵo inter tiranosaŭroj de ĉi tiu vivoperiodo; la bestoj ne mortis grandnombre en la jaroj tuj antaŭ la seksa maturiĝo kaj sekve ne produktiĝis fosilioj. Ĉi tiu rareco tamen povas ankaŭ ŝuldiĝi al la nekompleteco de la fosiliaro aŭ la prefero de fosili-kolektantoj pri grandaj, pli spektaklaj specimenoj.[30]
Seksa dimorfismo
Kiam la nombro de specimenoj plimultiĝis, sciencistoj ekanalizis la variadon inter unuopuloj kaj malkovris tion kio ŝajnis esti du apartajn korpoformojn, aŭ morfojn, kiel en iuj aliaj teropodoj specioj. Ĉar unu morfo estis de pli solida konstruo oni nomis ĝin fortika kaj la alia gracia. Pluraj morfologiaj diferencoj ligitaj kun la du morfoj uziĝis por analizi seksan dimorfismon ĉe Tyrannosaurus rex, kun la fortika morfo ĝenerale supozata esti ina. La pelvoj de pluraj fortikaj specimenoj, ekzemple, ŝajnis pli larĝaj, eble por permesi la trapason de ovoj.[31] Oni ankaŭ pensis ke estas regula korelacio inter la fortika morfologio kaj reduktiĝo de la ĉevrono sur la unua vosta vertebro, ankaŭ ŝajne por permesi al ovoj pasi el la reprodukta sistemo, kiel estis raportite por krokodiloj.[32]
En lastaj jaroj la argumentoj por seksa dimorfismo malfortiĝis. Studaĵo de 2005 raportis ke antaŭaj asertoj pri seksa dimorfismo en la anatomio de krokodilaj ĉevronoj malĝustis, kio siavice dubigis asertojn pri la ekzisto de seksa dimorfismo ĉe T. rex.[33] Oni trovis plengrandan ĉevronon ĉe la unua vosto-vertebro de "Sue", tre fortika unuopulo, kio indikas ke oni ĉiuokaze ne povas uzi ĉi tiun trajton por konsekvence diferencigi la du morfojn. Ĉar oni trovis specimenojn de T. rex en lokoj tiel disaj kiel Saskaĉevano kaj Nov-Meksiko, povas esti ke diferencoj inter unuopuloj respegulas geografian variadon kaj de seksan dimorfismon. La diferencoj ankaŭ povus rilati al aĝo: eble fortikaj unuopuloj simple estas pli aĝaj bestoj.[1]
Nur koncerne unu specimenon de T. rex oni povis difini sekson kun certeco: esplorado de "B-rex" montris ke konserviĝis molaj histoj ene de iuj ostoj, kaj oni povis identigi iujn el ĉi tiuj histoj kiel medolajn histojn, speciala histo kiun kreskigas modernaj birdoj kiel fonton de calcio por la produktado de ovoŝeloj dum ovolado. Nur inaj birdoj ovolas, kaj sekve oni trovas medolajn histojn nur en inoj, kvankam maskloj povas ĝin produkti se oni injekcias al ili inajn reproduktohormonojn kiel oestrogenon. Ĉi tio forte sugestas ke "B-rex" estis ina kaj ke ŝi mortis dum ovolado.[29] Lastatempaj esploroj montris ke medolaj histoj neniam produktiĝas ĉe krokodiloj, kiuj ŝajne estas la plej proksimaj ankoraŭ vivantaj parencoj de dinosaŭroj, krom la birdoj. La ĉeesto de medolaj histoj ĉe birdoj kaj teropodaj dinosaŭroj plue atestas pri la proksima evolua parenceco inter la birdoj kaj teropodoj.[34]
Teniĝo

_2012-05-11_04.jpg.webp)
Kiel multaj dupiedaj dinosaŭroj, Tyrannosaurus rex estis historie montrita kiel 'vivanta tripiedo', kun la korpo tenata je 45 gradoj (aŭ malpli) de vertikaleco kaj la vosto trenata surtere, iom simile kiel ĉe kanguruo. Ĉi tiu ideo datiĝas de la rekonstruaĵo de hadrosaŭro kiun faris en la jaro 1865 Joseph Leidy, la unua persono montrinta dinosaŭro en dupieda teniĝo.[35] Henry Fairfield Osborn, iama prezidanto de la Amerika Muzeo de Naturhistorio (AMNH) en Novjorko, kiu kredis ke tiranosaŭroj staris rekte, plifortigis la ideon eksponante en la muzeo la unuan kompletan tiranosaŭran skeleton en 1915; ĝi restis en ĉi tiu rektstara teniĝo dum preskaŭ jarcento, ĝis oni malmuntis ĝin en 1992.[36] Ĝis 1970 la sciencistoj konstatis ke ĉi tiu teniĝo estis erara kaj estus neeltenebla ĉe vivanta besto, ĉar ĝi rezultigus la delokiĝon aŭ malfortigon de pluraj artikoloj, inter ili la koksoj kaj la artikado de kapo kun la spino.[37] Spite siajn erarojn, la tiranosaŭra muntaĵo de la AMNH inspiris similajn montraĵojn en multaj filmoj kaj pentraĵoj (kiel la fama murpentraĵo de Rudolph Zallinger The Age Of Reptiles en Naturhistoria Muzeo Peabody de Universitato Yale) ĝis la 1990-aj jaroj, kiam filmoj kiel Jurassic Park prezentis pli ĝustan tiranosaŭran teniĝon al la ĝenerala publiko. Nuntempe reprezentiĝoj de T. rex en muzeoj, arto kaj filmoj montras ĝin kun la korpo pli-malpli horizontala kaj vosto etendita malantaŭ la korpo por kontraŭbilanci la kapon.[11]
Brakoj
.jpg.webp)
Kiam oni unue malkovris Tyrannosaurus rex la humero estis la sola konata elemento de la tiranosaŭra brako.[38] Por la unua muntaĵo de tiranosaŭra skeleto, kiel ĝin ekvidis la publiko en 1915, Osborn anstataŭigis longajn, tri-fingrajn brakojn kiel tiuj de alosaŭro.[24] Jaron pli frue, tamen, Lawrence Lambe jam priskribis la mallongajn, du-fingrajn brakojn de la proksime parenca gorgosaŭro.[39] Ĉi tio forte sugestis ke T. rex havis similajn brakojn, sed ĉi tiu hipotezo ne konfirmiĝis ĝis kiam oni identigis - en 1989 - la unuajn kompletajn tiranosaŭrajn brakojn, kiuj apartenis al MOR 555 (la "Wankel rex").[40] Inter la restaĵoj de "Sue" ankaŭ estas kompletaj brakoj.[1] La brakoj de T. rex estas tre malgrandaj kompare kun la ĝenerala korpograndeco, nur 1 m longaj. Ili tamen ne estas rudimentaj sed havas grandajn regionojn por la alligado de muskoloj, kio indikas grandan forton. Ĉi tion rekonis jam en 1906 Osborn, kiu proponis ke la brakoj uziĝis por teni parulon dum kopulacio.[41] Oni ankaŭ sugesti ke la brakoj uziĝis por helpi la beston kiam ĝi leviĝis de kuŝa pozicio.[37] Plia ebleco estas tio ke la brakoj tenis predobeston dum la enormaj makzeloj de la tiranosaŭro ĝin mortigis. Ĉi tiun hipotezon subtenas biomeĥanika analizo. La brak-ostoj de T. rex havas tre dikan kortikan oston, kio indikas ke ili evoluis por porti grandajn pezojn. La braka bicepso de plenkreska tiranosaŭro povis mem levi 199 kg; ĉi tiu numero nur pligrandiĝus kiam aliaj muskoloj (kiel la brakialo) kunlaborus kun la bicepso. La brako de T. rex havis limigitan movokapablon; la ŝulto- kaj kubuto-artikoloj permesis nur 40 kaj 45 gradojn respektive de moviĝo. La samaj du artikoj en deinoniko permesis 88 kaj 130 gradojn de moviĝo, respektive, dum homa brako povas rotacii 360 gradojn ĉe la ŝultro kaj povas moviĝi tra 165 gradoj ĉe la kubuto. La pezo strukturo de la brak-ostoj, la granda forteco de la muskoloj, kaj limigita moviĝkapablo eble indikas sistemo farita por firme teni spite la premojn de baraktanta predobesto.[42]
Molaj histoj
En la numero de Science de marto 2005, Mary Higby Schweitzer of Ŝtata Universitato de Norda Karolino kaj kolegoj anoncis la regajnon de molaj histoj de medola kavaĵo en fosiligita krur-osto, de 68-milion-jara tiranosaŭro. La oston oni intence, kvankam malvolonte, rompis por transsendado kaj sekve ne konservis en la normala maniero, specife pro tio ke Schweitzer esperis testi ĝin por ekscii ĉu ĉeestas molaj histoj. Kiam oni elfosis la dinosaŭron, kiu ricevis la specimen-numeron MRM 1125, el la Formacio Hell Creek oni rekonis flekseblajn, disforkiĝantajn sangovaskulojn kaj fibran sed elastan matricohistojn de osto. Aldone estis mikrostrukturoj similaj al sangoglobuloj en la osta matrico kaj la vaskuloj. La strukturoj similas al strutaj sango-globuloj kaj vaskuloj. Ĉu la materialon konservis nekonata procedo, aparta de normala fosiliĝo, aŭ ĉu la materialo estas originala, la esploristoj ne scias, kaj ili sin gardas ne fari asertojn pri konservado.[43] Se estos decidite ke la materialo estas originala, oni uzos travivintajn proteinojn por nerekte diveni la DNA-enhavon de la koncernaj dinosaŭroj, ĉar ĉiun proteinon tipe kreas specifa geno. La fakto ke oni ne trovis ĉi tiajn histojn antaŭe povus simple esti sekvo de tio ke homoj supozis la konservadon de molaj histoj neebla kaj sekve ne serĉis ilin. Depost la unua malkovro oni trovis ĉi tiujn histecajn strukturojn ĉe du pliaj tiranosaŭroj kaj hadrosaŭro.[44] Esplorado de iuj el la histoj sugestas ke birdoj estas pli proksime parencaj al tiranosaŭroj ol aliaj ĉiuj aliaj modernaj bestoj.[45]
En sekvaj studaĵoj pri kiuj oni raportis en la gazeto Science en aprilo 2007, Asara kaj kolegoj konkludis ke sep protein-restaĵoj trovitaj en purigita osto de T. rex plej proksime akordiĝas kun tiuj de kokoj, kaj due plej proksime kun tiuj de ranoj kaj trituroj. La malkovrado de proteinoj de besto aĝa dekmilionoj de jaroj, kune kun la malkovrado de similaj kemi-restaĵoj trovitaj en mastodonta osto almenaŭ 160.000 jarojn aĝa, renversas la tradician vidpunkton pri fosilioj kaj eble ŝanĝos la fokuson de paleontologoj de ostoĉasado al biokemio. Ĝis ĉi tiuj malkovroj la plimulto de sciencistoj supozis ke fosiliĝo anstataŭigis ĉiujn vivantajn histojn per nevivantajn mineralojn. Paleontologo Hans Larsson de Universitato McGill en Montrealo, kiu ne partoprenis la esplorojn, nomas la malkovrojn "mejloŝtono" kaj sugestis ke dinosaŭroj povus "enter the field of molecular biology and really slingshot paleontology into the modern world" [eniri la kampon de molekula biologio kaj vere ĵeti paleontologion en la modernan mondon].[46]
Haŭto
En 2004 la scienca gazeto Nature publikigis raporton priskribanta fruan tiranosaŭroideon, Dilong paradoxus, de la fama Formacio Yixian en Ĉinio. Kiel ĉe multaj aliaj teropodoj malkovritaj en Formacio Yixian, la fosilia skeleto konserviĝis kune kun tavolo de fibrecaj strukturoj ĝenerale rekonataj kiel prastrukturoj de plumoj. Oni ankaŭ proponis ke tiranosaŭroj kaj aliaj proksim-parencaj tiranosaŭredoj havis tiajn praplumojn. La raraj haŭtpremaĵoj de plenkreskaj tiranosaŭredoj en Kanado kaj Mongolio tamen montras la gruzecajn skvamojn tipaj de aliaj dinosaŭroj. Certe eblas ke praplumoj kovris partojn de la tiranosaŭra korpo ne konservitaj, sed la manko de izoliga korpo-kovraĵo estus tute konsekvenca se oni konsideras ke tio ankaŭ mankas ĉe modernaj mult-tunaj mamuloj kiel elefantoj, hipopotamoj, kaj la plej multaj rinoceraj specioj. Kiam objekto pligrandiĝas, ĝia kapablo reteni varmecon pligrandiĝas pro la malgrandiĝanta rilato de surfaca areo al volumeno. Sekve, kiam grandaj bestoj evoluas aŭ ekvivas en varmaj klimatoj, kovraĵo de haro aŭ plumoj perdas evoluan avantaĝon kaj povas fariĝi malavantaĝo, ĉar la izolaĵo retenas varmecon en la korpo kaj donas la riskon tro multe varmigi la beston. Povas do esti ke dum grandaj tiranosaŭredoj kiel tiranosaŭro evoluis, ili perdis la praplumojn hereditaj de terapodoj prauloj, precipe en varmaj klimatoj.[47]
Ĉu la tiranosaŭro estis varmsanga?
Tiranosaŭro, kiel la plimulto de dinosaŭroj, estis longe supozata havi malvarm-sangan (ektoterman), reptilian metabolismon. La ideon pri dinosaŭra malvarm-sangecon kontestis sciencistoj kiel Robert Bakker kaj John Ostrom en la fruaj jaroj de la "Dinosaŭra Renesanco" ekde la malfruaj 1960-aj jaroj.[48][49] Oni asertis ke tiranosaŭro mem estis varm-sanga (endoterma), kio implicas tre aktivan vivstilon.[4] Depost tiam, pluraj paleontologoj penas precize scii ĝis kiu grado tiranosaŭro povis reguligi sian korpotemperaturon. Histologiaj atestaĵoj pri alta kreskorapideco ĉe junaj tiranosaŭroj, kompareblaj kun tiuj ĉe mamuloj kaj birdoj, subtenas la hipotezon ke tiranosaŭroj havis altan metabolismon. Kreskokurbiĝoj indikas ke, same kiel ĉe mamuloj kaj birdoj, la kreskado de T. rex limiĝis ĉefe al nematuraj bestoj, malkiel la konstanta kreskado trovata ĉe la plimulto de la aliaj vertebruloj.[28]
Oni foje uzas la rejŝojn de oksigenaj izotopoj en fosiligita osto por ekscii la temperaturo je kiu deponiĝi la osto, ĉar la rejŝo inter certaj izotopoj varias laŭ temperaturo. En unu specimeno, la izotopaj rejŝoj en ostoj de diversaj korpopartoj indikis temperaturan diferencon de ne pli ol 4 ĝis 5 gradusoj inter la torakaj vertebroj kaj la tibio de la kruro. Laŭ la paleontologo Reese Barrick kaj la geokemiisto William Showers ĉi tiel malgranda temperaturdiferenco inter la korpokerno kaj la ekstremaĵoj indikas konstantan internan korpotemperaturon (homeotermeco) indikanta metabolismon inter tiu de la malvarm-sangaj reptilioj kaj la varm-sangaj mamuloj.[50] Aliaj sciencistoj atentigis ke la rejŝo de oksigenaj izotopoj en la fosilioj hodiaŭ ne nepre reprezentas la saman rejŝon en la fora pasinteco kaj eble ŝanĝiĝis dum aŭ post la fosiliĝo.[51] Barrick kaj Showers defendis siajn konkludojn en sekvaj artikoloj, trovante similajn rezultojn en alia teropoda dinosaŭro vivinta sur alia kontinento kaj dekmilionojn da jaroj pli frue (giganotosaŭro).[52] Ornitiskiaj dinosaŭroj montris atestaĵojn de homeotermeco, dum varanedaj lacertoj de la sama formacio ne montris tion.[53] Sed eĉ se tiranosaŭroj fine montras atestaĵojn de homeotermeco, ĉi tio ne nepre implicas ke ili estis endotermaj. Tian reguligon de varmeco povas klarigi ankaŭ gigantotermeco, kion oni vidas ĉe iuj vivantaj kelonioj.[54][55]
Manĝostrategioj

La plej granda parto de debato pri tiranosaŭro fokusiĝas je ĝiaj manĝostrategioj kaj sinmovado. Unu paleontologo, la fama hadrosaŭreda spertulo Jack Horner, asertas ke tiranosaŭro plejparte estis postpredanto kaj tute ne ĉasis aktive.[40] Horner nur prezentis ĉi tion unu fojon en oficiala scienca kuntekston, sed multe diskutas ĝin en siaj libroj kaj en intervjuoj kun ĵurnalistoj. Lia hipotezo havas la sekvan bazon: tiranosaŭroj havis grandajn olfaktajn bulbojn kaj olfaktajn nervojn (kompare kun ilia cerbograndeco). Ĉi tio sugestas alte evoluintan flarsenton, laŭdire uzita por flarserĉi kadavrojn de longe for. Tiranosaŭraj dentoj povis trarompi dentojn, kion ili eble uzis por eltiri kiel eble plej multe da manĝaĵo (medolo) de kadavraj restaĵoj, kutime la malplej nutraj partoj. Ĉar almenaŭ iuj el la predobestoj de tiranosaŭro povis moviĝi rapide, atestaĵoj ke tiranosaŭroj promenis anstataŭ kuri eble atestas pri tio ke ĝis estis postpredanto.[56][57]
La plej multaj sciencistoj publikigintaj artikolojn pri ĉi tiu temo insistas ke tiranosaŭro estis kaj predanto kaj postpredanto, manĝante kiun ajn viandon trovebla laŭ la okazo.[58] Modernaj karnomanĝuloj kiel leonoj kaj hienoj ofte postpredas tion kion mortigis aliaj predantoj, kaj estas verŝajne ke tiranosaŭroj ankaŭ tion faris.[59]
Ankoraŭ aliaj atestaĵoj sugestas ĉasan agadon ĉe tiranosaŭroj. La okulkavaĵoj de tiranosaŭroj havas pozicion permesanta al la okuloj direktiĝi antaŭen, donante al la dinosaŭro binoklan vidpovon.[60] Postpredanto eble ne bezonus la precizan perceptadon de profundeco kiun donas binokla vidpovo; en modernaj bestoj la binoklan vidpovon ĉefe havas predantoj.
Kiam li esploris Sue, la paleontologo Pete Larson trovis rompitajn kaj resaniĝintajn fibulon kaj vostovertebrojn, cikatriĝintajn vizaĝostojn, kaj denton de alia tiranosaŭro enfiksita en kolovertebro. Ĉi tio atestas pri agresiva konduto inter tiranosaŭroj, sed ne estas klare ĉu la agresadon kaŭzis konkurencado por manĝaĵo kaj paruloj ĉu aktiva kanibalismo.[61] Plia esplorado lastatempa de ĉi tiuj "vundiĝoj" tamen montris ke ili plejparte estas infektiĝoj kaj ne vundiĝoj (aŭ simple postmorta damaĝo al la fosilio) kaj la malmultaj veraj vundiĝoj estas tro ĝeneralaj por indiki interspecian konflikton.[62] Ĉe la elfosejo de Sue oni trovis ankaŭ skeleton de edmontosaŭro (Edmontosaurus annectens) kun cikatroj faritaj de tiranosaŭraj dentoj survoste. La fakto ke la vundoj cikatriĝe resaniĝis sugestas aktivan predadon flanke de la tiranosaŭroj.[63][64] Alian atestaĵon provizas triceratopo trovita kun mordoŝpuroj sur ĝia ilio; denove la mordinto estis tiranosaŭro, kaj la vundoj ŝajne resaniĝis.[65]
Sciencistoj malkonsentas inter si pri tio, ĝis kiu grado tiranosaŭroj povis kuri kaj precize kiel rapide ili povis kuri (se entute); iuj sugestis rapidecojn de ĝis 70 km/h aŭ pli. Tamen laŭ James Farlow, palaeontologo ĉe Universitato de Indianao-Purdue en Fort Wayne (Indianao): "If T. rex had been moving fast and tripped, it would have died. [Se T. rex moviĝus rapide kaj stumblus, ĝi mortus.]"[66] Se tiranosaŭro stumblus kaj falus dum kurado, ĝia torako frapus la teron kun malakcelado de 6 g (sesoblo de la akcelado pro gravito, aŭ ĉ. 60 m/s²).[5] (Vidu ankaŭ Sinmovado, sube.)
Iuj diras ke se tiranosaŭroj estis postpredantoj, alia dinosaŭro devis esti la ĉefa predanto en la malfrua kretaceo de Amerazio. La ĉefaj predobestoj estis la pli grandaj marginocefaloj kaj ornitopodoj. La aliaj tiranosaŭredoj dividas tiom da trajtoj kun tiranosaŭroj ke nur la malgrandaj dromeosaŭredoj restas bona alternativo kiel ĉefaj predantoj. De ĉi tiu vidpunkto, subtenantoj de la hipotezo pri tiranosaŭro kiel postpredanto sugestis ke la grandeco kaj potenco de tiranosaŭroj permesis al ili ŝteli predaĵojn de malpli grandaj predantoj.[57]
Sinmovado
Sciencistoj pensantaj ke tiranosaŭroj povis kuri malrapide atentigas pri la kavaj ostoj kaj aliaj trajtoj kiuj malpezigis la korpon tiom ke plenkreskulo eble peze ne pli ol 5 tunojn, kaj ili atentigas ke bestoj kiel strutoj kaj ĉevaloj kun longaj flekseblaj kruroj povas atingi altan rapidecon per malpli rapidaj sed pli longaj paŝoj. Iuj aldone argumentis ke tiranosaŭroj havis relative pli grandajn kruromuskulojn ol iu ajn besto hodiaŭ vivanta, kaj ke ĉi tiuj estus povintaj ebligi rapidan kuradon (40–70 km/h).[67]
Iuj malnovaj studoj de krura anatomio kaj vivantaj bestoj sugestis ke tiranosaŭroj entute ne povis kuri kaj nure promenis. La rilato de femura al tibia longeco (pli ol 1, kiel en la plimulto de teropodoj) povus indiki ke tiranosaŭroj estis specialigitaj promenantoj, kiel modernaj elefantoj. Aldone, ĝi havis etajn 'brakojn', kiuj ne povus haltigi falon, se la tiranosaŭro stumblus dum kurado; kutimaj taksoj de la tiranosaŭra pezo (6 ĝis 8 tunoj) donus mortigan frapoforton al falanta tiranosaŭro.[68] Estas tamen notinde ke ĝirafoj foje galopas je rapideco de 50 km/h.[69] Je ĉi tiuj rapidecoj, ĝirafoj riskas rompi kruron, kio povas esti mortiga eĉ kiam la akcidento okazas en 'sekura' ĉirkaŭaĵo, kiel zoo.[70] Se ĝi povis kuri, tiranosaŭroj eble estis riskuloj, same kiel multaj modernaj bestoj estas riskuloj. Taksoj de la krurosta forteco ĉe tiranosaŭroj tamen indikas ke ĝiaj kruroj apenaŭ estis pli fortaj (se entute tiel fortaj) ol tiuj de elefonatoj, kiuj estas sufiĉe limigitaj koncerne ilian plej altan rapidecon kaj neniam estas 'en la aero', kiel okazas dum kurado.
Subtenantoj de la promena hipotezo taksas ke tiranosaŭroj povis promeni ne pli rapide ol 17 km/h, kio tamen estas rapideco pli alta ol tiu de la plej verŝajnaj predospecioj kunvivantaj kun tiranosaŭroj, la hadrosaŭredoj kaj ceratopoj.[71] Iuj subtenantoj de la preda hipotezo aldone asertas ke tiranosaŭra rapideco ne gravas, ĉar povas esti ke ĝi estis malrapida sed pli bone adaptita por rapida moviĝo ol ĝiaj predaĵoj[72] aŭ eble ĝi embuske kaptis la pli rapidajn predobestojn.[67]
La plej lastatempaj esploroj je tiranosaŭra sinmovado ne specifigas la rapidecon de tiranosaŭra kurado, sed agnoskas ke malfacilas diri iom pli precizan ol tio ke la plej granda rapideco estis inter 17 km/h, promenado aŭ malrapida kurado, kaj 40 km/h, mez-rapida kurado. Ekzemple, artikolo en Nature[71] uzis matematikan modelon (kiun ĝi validigis per aplikado al du vivantaj bestoj, aligatoroj kaj kokoj) por taksi la mason de kruromuskulo bezonata por rapida kurado (pli ol 40 km/h). Ili trovis ke proponitaj rapidecoj pli altaj ol 40 km/h estis neeblaj, ĉar ili necesigus tre grandajn kruromuskolojn (pli ol 40–86% de la tuta korpomaso).[73] Eĉ modere altaj rapidecoj necesigus grandajn kruromuskolojn. Tamen ne eblas findecidi ĉi-teme, ĉar ni ne scias kiom grandaj estis la tiranosaŭraj kruromuskoloj. Se ili estus malpli grandaj, nur promenado/trotado de 18 km/h eblus.[67]
Tamen laŭ Thomas R. Holtz Jr estas notinde ke konsiderante la pezon de la tiranosaŭro (5-7 tunoj), la rejŝoj de tibio al femuro kaj metatarso al femuro kategoriigas la tiranosaŭron inter la plej graciaj bestoj inter ĉiuj bestoj konataj de la mezozoikaj aŭ kenozoikaj fosilioj.
Ĉe la tipa grandeco de plenkreska tiranosaŭro la membroproporcioj de gracia besto ŝajnas malgraciaj, sed se oni komparas ilin kun la malantaŭaj kruroj de aliaj similgrandaj bestoj, ekzemple elefanto, triceratopo aŭ edmontosaŭro, la kruroj de Tyrannosaurus rex estas pli maldikaj kaj havas pli longajn tibiojn kaj metatarsojn.
Kompare kun aliaj familioj de grandaj teropodoj, la tiranosaŭredoj havis membroproporciojn pli graciaj. La malgrandaj tiranosaŭredoj estis ankoraŭ pli graciaj, kaj la plej malgrandaj havis la samajn membroproporciojn kiel la plej grandaj ornitomimedoj: laŭ grandeco, la kruroj de alektrosaŭro kaj galimimo estas identaj.
Tiranosaŭredoj aldone havis ornitomimedecajn piedojn, pli malgrandaj kaj pli maldikaj ol tiuj de la aliaj grandaj teropodoj, kio signifas ke ili havis pli efikan sinmovadon kiam la kruro artikiĝis dum movado.
Laŭ Holtz, kiu vidas la aferon el la vidpunkto de funkcia morfologio, la tiranosaŭredoj estis pli bone adaptitaj por rapideco ol iu ajn alia familio de grandaj teropodoj, tio estas, la alosaŭroideoj, megalosaŭroideoj kaj neoceratosaŭroj. Tio eble ne signifas ke T. rex povis movi je grandaj rapidecoj, sed por besto tiel granda ĝi tamen estis optimume konstruita por rapideco kaj - kio gravas por la debato pri tio ĉu ĝi estis predanto aŭ postpredanto - pli rapida ol ĝiaj predobestoj.[74]
Novaj atestaĵoj kolektitaj per biomeĥanikaj komputil-modeloj sugestas ke tiranosaŭroj havis limigitan eblecon sin turni. Laŭ John Hutchinson, spertulo pri biomeĥaniko ĉe la Reĝa Veterinara Kolegio de la Universitato de Londono en Anglio, tiranosaŭroj verŝajne bezonis unu ĝis du sekundojn por turni sin nur 45° – kion homoj, kun vertikala orientiĝo kaj sen vosto, povas sin turni dum sekundero.[75]
Historio
Henry Fairfield Osborn, prezidanto de la Amerika Muzeo de Naturhistorio, nomis Tyrannosaurus rex en la jaro 1905. La genronomo venas de la malnovgrekaj vortoj τυραννος (= "tirano") kaj σαυρος (= "lacerto"). Osborn uzis la latina vorto rex (= "reĝo") por la specia nomo. Kune ili emfazas la grandecon de la besto kaj supozatan regadon sur aliaj tiutempaj specioj.[38]
Plej fruaj trovaĵoj
Oni povas konsideri la vertebrojn de la dinosaŭro, kiun Cope nomis Manospondylus en 1892 la unua konata specimeno de Tyrannosaurus rex. Barnum Brown, vickuratoro de la Amerika Muzeo de Naturhistorio, trovis la duan tiranosaŭran skeleton en Vajomingo en 1900. Ĉi tiun specimenon unue ricevis la nomon Dynamosaurus imperiosus en la sama artikolo en kiu oni priskribis Tyrannosaurus rex.[76] Nur la paĝordo malhelpis ke Dynamosaurus fariĝu la oficiala nomo. La originala specimeno de "Dynamosaurus" estas en la kolektaĵoj de la Naturhistoria Muzeo (Londono).[77]

Barnum Brown trovis kvin tiranosaŭrajn (partajn) skeletojn entute. Brown kolektis sian duan tiranosaŭron en 1902 kaj 1905 de la Formacio Hell Creek (Montano). Ĉi tiu estas la holotipo uzata por priskribi Tyrannosaurus rex Osborn, 1905. En 1941 oni vendis ĝin al la Muzeo Carnegie de Naturhistorio en Pittsburgh (Pensilvanio). La kvara kaj plej granda trovaĵo de Brown, ankaŭ de Formacio Hell Creek, eksponiĝas ĉe la Amerika Muzeo de Naturhistorio en Novjorko.[40]
Kvankam multaj skeletoj estis trovitaj en diversaj mondopartoj, oni trovis nur unu tiranosaŭran ŝpuron, ĉe Philmont Scout Ranch en nordorienta Nov-Meksiko. Oni trovis ĝin en 1983 kaj identigis kaj formale priskribis ĝin en 1994.[78]
Notindaj specimenoj

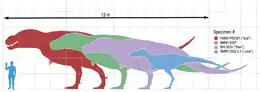
Sue Hendrickson, amatora paleontologo, malkovris la plej (pli ol 90%-e) kompletan kaj, ĝis 2001, plej grandan fosilian skeleton de tiranosaŭro; ŝi trovis ĝin en Formacio Hell Creek proksime de Faith (Suda Dakoto) la 12-an de aŭgusto 1990. Pri ĉi tiu tiranosaŭro, nun nomita "Sue" por honori ŝin, estis leĝa batalo por decidi, kiu ĝin posedas. En 1997 oni decidis ke ĝin posedas Maurice Williams, la posedanto de la tereno kie oni trovis la fosilian skeleton, kaj oni vendis la kolektaĵon aŭkcie por 7,6 milionoj da usonaj dolaroj; tiel ĝi fariĝis la plej multekosta dinosaŭra skeleto ĝis nun. Ĝi nun estas rekunmetita kaj eksponiĝas ĉe la Muzeo Field de Naturhistorio. Surbaze de studado de 'ŝiaj' fosiligitaj ostoj oni kredas ke Sue mortis 20-jaraĝa, atinginte plenan grandecon kiam ŝi havis 19 jarojn. Sciencistoj raportas ke oni trovis skeletojn de nematuran kaj idan tiranosaŭrojn en la sama ŝtonminejo kiel Sue; ĉi tio subtenas la hipotezon ke tiranosaŭroj grupe vivis.[79]
Alian tiranosaŭron, al kiu oni donis la kromnomon "Stan" honore al la amatora paleontologo Stan Sacrison, oni trovis en la Formacio Hell Creek proksime de Buffalo (Suda Dakoto) en printempo de 1987. Post 30 mil horoj da fosado kaj zorgema pretigado aperis 65-procente kompleta skeleto, kiu nun eksponiĝas ĉe la Naturhistoria Muzeo Black Hills en Hill City (Suda Dakoto) post longa rondvojaĝo tra la mondo. Ankaŭ ĉe ĉi tiu tiranosaŭro oni trovis multajn ostopatologiojn, inkluzive rompitajn kaj resaniĝintajn ripojn, rompitan (kaj resaniĝintan) kolon kaj grandan truon en la kapa malantaŭo, kun la proksimuma grandeco de tiranosaŭra dento. Kaj Stan kaj Sue esploris Peter Larson.
En 2001 teamo de Naturhistoria Muzeo Burpee de Rockford (Ilinojo) trovis 50-procente kompletan skeleton de juna tiranosaŭro en la Formacio Hell Creek. Kromnomita "Jane the Rockford T-Rex" la specimeno unue traktiĝis kiel la unua konata skeleto de la nana tiranosaŭredo nanotirano (Nanotyrannus) sed postaj esploroj montris ke pli verŝajne temas pri juna tiranosaŭro.[80] Ĝi estas la kompleta kaj plej bone konservita ekzemplero de juna tiranosaŭro ĝis nun konata. Jane esploris Jack Horner, Pete Larson, Robert Bakker, Greg Erickson kaj pluraj aliaj famaj paleontologoj pro la unikeco de ŝia aĝo. Jane nuntempe eksponiĝas ĉe Naturhistoria Muzeo Burpee en Rockford (Ilinojo).[81][82]
Ankaŭ en la jaro 2001, Jack Horner malkovris tiranosaŭran specimenon ĉ. 10-procente pli granda ol "Sue". Oni ankoraŭ esploras ĉi tiun specimenon, al kiu oni donis la kromnomon C. rex (aŭ "Celeste" honore al la edzino de Jack).
En gazeta komuniko dissendita la 7-an de aprilo 2006, Ŝtata Universitato de Montano anoncis ke ĉi posedas la plej grandan tiranosaŭran kranion ĝis nun malkovrita. Malkovrita dum la 60-aj jaroj kaj nur lastatempe rekonstruita, la kranio estas 150 cm longa, pli granda ol la 141 cm longa kranio de “Sue” je 6,5%.[83][84]
La tiranosaŭro en la populara kulturo
Depost kiam oni ĝin unue priskribis en 1905, la tiranosaŭro fariĝis la plej vaste rekonata dinosaŭro en la populara kulturo. Ĝi ankaŭ estas la sola dinosaŭro kiun membroj de la ĝenerala publiko kutime nomas per ĝia scienca nomo (Tyrannosaurus rex), kaj la scienca mallongiĝo T. rex ankaŭ ĝuas vastan uzadon.[1] Muzeaj eksponaĵoj fokusiĝantaj je la tiranosaŭro estas tre popularaj; proksimume dek mil vizitantoj venis al la Muzeo Field de Ĉikago la tagon kiam malfermiĝis ĝia eksponaĵo pri "Sue" en 2003.[85] T. rex aperis multfoje en televido kaj en filmoj, unuavice en The Lost World, King Kong, The Land Before Time, Jurassic Park, kaj Night at the Museum. En diversaj libroj kaj bildstrioj, inter ili Calvin and Hobbes, ankaŭ aperis la tiranosaŭro, kiun oni kutime montras kiel la plej grandan kaj plej terurigan karnovorulon. Almenaŭ unu muzikgrupo, la bando T. Rex, alprenis sian nomon laŭ la specio. Ludiloj kiuj rilatas al tiranosaŭro, inter ili multaj videoludoj kaj aliaj vendaĵoj, restas popularaj. Multaj komercoj faris kapitalon de la populareco de la tiranosaŭro uzante ĝin en reklamoj.
Referencoj
- 1 2 3 4 5 6 Brochu, C.R. 2003. Osteology of Tyrannosaurus rex: insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull. Memoirs of the Society of Vertebrate Paleontology. 7: 1-138.
- ↑ Henderson, D.M. 1999. Estimating the masses and centers of mass of extinct animals by 3-D mathematical slicing. Paleobiology 25: 88–106.
- ↑ Anderson, J.F., Hall-Martin, A. & Russell, D.A. 1985. Long bone circumference and weight in mammals, birds and dinosaurs. Journal of Zoology 207: 53–61.
- 1 2 Bakker, R.T. 1986. The Dinosaur Heresies. New York: Kensington Publishing. 481pp.
- 1 2 Farlow, J.O., Smith, M.B., & Robinson, J.M. 1995. Body mass, bone "strength indicator", and cursorial potential of Tyrannosaurus rex. Journal of Vertebrate Paleontology 15: 713-725.
- ↑ Seebacher, F. 2001. A new method to calculate allometric length-mass relationships iof dinosaurs. Journal of Vertebrate Paleontology 21(1): 51–60.
- ↑ Christiansen, P. & Fariña, R.A. 2004. Mass prediction in theropod dinosaurs. Historical Biology 16: 85-92.
- 1 2 3 Erickson, G.M., Makovicky, P.J., Currie, P.J., Norell, M.A., Yerby, S.A., & Brochu, C.A. 2004. Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs. Nature 430: 772-775.
- 1 2 Currie, P.J., Hurum, J.H., and Sabath, K. 2003. Skull structure and evolution in tyrannosaurid dinosaurs. Acta Palaeontologica Polonica 48(2): 227–234. (deŝutu ĉi tie Arkivigite je 2007-06-12 per la retarkivo Wayback Machine)
- 1 2 3 Holtz, T.R. 2004. Tyrannosauroidea. En: Weishampel, D.B., Dodson, P., & Osmolska, H. (red.). The Dinosauria (2-a eldono). Berkeley: University of California Press. Pp. 111-136.
- 1 2 3 Paul, G.S. 1988. Predatory Dinosaurs of the World. New York: Simon & Schuster. 464pp.
- 1 2 Holtz, T.R. 1994. The phylogenetic position of the Tyrannosauridae: implications for theropod systematics. Journal of Palaeontology 68(5): 1100-1117.
- ↑ Maleev, E.A. 1955. [Gigantic carnivorous dinosaurs of Mongolia]. Doklady Akademii Nauk S.S.S.R. 104(4): 634-637. [en rusa lingvo]
- ↑ Rozhdestvensky, A.K. 1965. Growth changes in Asian dinosaurs and some problems of their taxonomy. [Paleontological Journal] 3: 95-109.
- ↑ Carpenter, K. 1992. Tyrannosaurids (Dinosauria) of Asia and North America. En: Mateer, N. & Chen P. (red.). Aspects of Nonmarine Cretaceous Geology. Beijing: China Ocean Press Pp. 250-268. (deŝutu ĉi tie Arkivigite je 2007-09-27 per la retarkivo Wayback Machine)
- ↑ Carr, T.D., Williamson, T.E., & Schwimmer, D.R. 2005. A new genus and species of tyrannosauroid from the Late Cretaceous (Middle Campanian) Demopolis Formation of Alabama. Journal of Vertebrate Paleontology 25(1): 119–143.
- ↑ Hurum, J.H. & Sabath, K. 2003. Giant theropod dinosaurs from Asia and North America: Skulls of Tarbosaurus bataar and Tyrannosaurus rex compared. Acta Palaeontologica Polonica 48(2): 161–190. (deŝutu ĉi tie Arkivigite je 2007-06-12 per la retarkivo Wayback Machine)
- ↑ Olshevsky, George (1995). “The origin and evolution of the tyrannosaurids”, Kyoryugaku Saizensen [Dino Frontline] 9-10, p. 92–119 (9) 75–99 (10).
- ↑ Carr, T.D. & Williamson, T.E. 2004. Diversity of late Maastrichtian Tyrannosauridae (Dinosauria: Theropoda) from western North America. Zoological Journal of the Linnean Society 142: 479–523.
- ↑ Gilmore, C.W. 1946. A new carnivorous dinosaur from the Lance Formation of Montana. Smithsonian Miscellaneous Collections 106: 1-19.
- ↑ Bakker, R.T., Williams, M., & Currie, P.J. 1988. Nanotyrannus, a new genus of pygmy tyrannosaur, from the latest Cretaceous of Montana. Hunteria 1(5): 1-30.
- ↑ Carr TD. 1999. Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Theropoda). Journal of Vertebrate Paleontology 19: 497–520.
- ↑ Currie, P.J. 2003. Cranial anatomy of tyrannosaurid dinosaurs from the Late Cretaceous of Alberta, Canada. Acta Palaeontologica Polonica 48(2): 191–226. (deŝutu ĉi tie Arkivigite je 2007-06-12 per la retarkivo Wayback Machine)
- 1 2 Osborn, H.F. 1917. Skeletal adaptations of Ornitholestes, Struthiomimus, Tyrannosaurus. Bulletin of the American Museum of Natural History 35: 733–71.
- ↑ "T. rex may be in for a name change Arkivigite je 2007-09-26 per la retarkivo Wayback Machine" by David McCormick. Discovery Channel Canada. 13-a de junio 2000. Aliro 20-a de julio 2006.
- ↑ International Code of Zoological Nomenclature, Fourth Edition. Article 23.9 - Reversal of Precedence Arkivigite je 2009-06-09 per la retarkivo Wayback Machine. International Commission on Zoological Nomenclature. 1-a de januaro 2000. Aliro 20-a de julio 2006.
- ↑ "So why hasn't Tyrannosaurus been renamed Manospondylus?" by Mike Taylor. 27-a de aŭgusto 2002. Aliro 20-a de julio 2006.
- 1 2 Horner, J.R. & Padian, K. 2004. Age and growth dynamics of Tyrannosaurus rex. Proceedings of the Royal Society of London B 271: 1875-1880.
- 1 2 Schweitzer, M.H., Wittmeyer, J.L., & Horner, J.R. 2005. Gender-specific reproductive tissue in ratites and Tyrannosaurus rex. Science 308: 1456-1460.
- 1 2 Erickson, G.M., Currie, P.J., Inouye, B.D., & Winn, A.A. 2006. Tyrannosaur life tables: an example of nonavian dinosaur population biology. Science 313: 213-217.
- ↑ Carpenter, K. 1990. Variation in Tyrannosaurus rex. En: Carpenter, K. & Currie, P.J. (red.). Dinosaur Systematics: Approaches and Perspectives. New York: Cambridge University Press. Pp. 141-145. (deŝutu ĉi tie Arkivigite je 2007-09-27 per la retarkivo Wayback Machine)
- ↑ Larson, P.L. 1994. Tyrannosaurus sex. En: Rosenberg, G.D. & Wolberg, D.L. Dino Fest. The Paleontological Society Special Publications. 7: 139-155.
- ↑ Erickson, G.M., Lappin, A.K., & Larson, P.L. 2005. Androgynous rex. The utility of chevrons for determining the sex of crocodilians and non-avian dinosaurs. Zoology 108: 277-286.
- ↑ Schweitzer, M.H., Elsey, R.M., Dacked, C.G., Horner. J.R., & Lamm, E.-T. 2007. Do egg-laying crocodilian (Alligator mississippiensis) archosaurs form medullary bone? Bone 40 (4): 1152-1158.
- ↑ Leidy, J. 1865. Memoir on the extinct reptiles of the Cretaceous formations of the United States. Smithsonian Contributions to Knowledge. 14: 1-135.
- ↑ "Tyrannosaurus Arkivigite je 2008-12-08 per la retarkivo Wayback Machine" American Museum of Natural History. (20-a de julio 2006).
- 1 2 Newman, B.H. 1970. Stance and gait in the flesh-eating Tyrannosaurus. Biological Journal of the Linnean Society. 2: 119-123.
- 1 2 Osborn, H.F. 1905. Tyrannosaurus and other Cretaceous carnivorous dinosaurs. Bulletin of the American Museum of Natural History 21: 259-265. (deŝutu ĉi tie Arkivigite je 2008-02-28 per la retarkivo Wayback Machine)
- ↑ Lambe, L.M. 1914. On a new genus and species of carnivorous dinosaur from the Belly River Formation of Alberta, with a description of the skull of Stephanosaurus marginatus from the same horizon. Ottawa Naturalist 27: 129-135.
- 1 2 3 Horner, J.R. & Lessem, D. 1993. The Complete T. rex: How Stunning New Discoveries Are Changing Our Understanding of the World's Most Famous Dinosaur. New York: Simon & Schuster. 235pp.
- ↑ Osborn, H.F. 1906. Tyrannosaurus, Upper Cretaceous carnivorous dinosaur (second communication). Bulletin of the American Museum of Natural History. 22: 281-296. (deŝutu ĉi tie Arkivigite je 2014-12-21 per la retarkivo Wayback Machine)
- ↑ Carpenter, K. & Smith, M.B. 2001. Forelimb osteology and biomechanics of Tyrannosaurus. En: Tanke, D.H. & Carpenter, K. (Eds.). Mesozoic Vertebrate Life. Bloomington: Indiana University Press. Pp. 90-116. (deŝutu ĉi tie Arkivigite je 2007-09-27 per la retarkivo Wayback Machine)
- ↑ Schweitzer M.H., Wittmeyer J.L., Horner J.R., Toporski J.B. 2005. Soft Tissue Vessels and Cellular Preservation in Tyrannosaurus rex. Science 307: 1952-1955.
- ↑ . Smithsonian Magazine Online. Alirita 2006-05-01.
- ↑ . Alirita 12-a de aprilo 2007.
- ↑ Yesterday's T. Rex is today's chicken. Yahoo news, 13-a de aprilo 2007.
- ↑ Xu X., Norell, M.A., Kuang X., Wang X., Zhao Q., & Jia C. 2004. Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids. Nature 431: 680-684.
- ↑ Bakker, R.T. 1968. The superiority of dinosaurs. Discovery 3: 11-22.
- ↑ Bakker, R.T. 1972. Anatomical and ecological evidence of endothermy in dinosaurs. Nature 238: 81-85.
- ↑ Barrick, R.E. & Showers, W.J. 1994. Thermophysiology of Tyrannosaurus rex: Evidence from oxygen isotopes. Science 265: 222-224.
- ↑ Trueman, C., Chenery, C., Eberth, D.A. & Spiro, B. 2003. Diagenetic effects on the oxygen isotope composition of bones of dinosaurs and other vertebrates recovered from terrestrial and marine sediments. Journal of the Geological Society, London 160: 895–901.
- ↑ Barrick, R.E. & Showers, W.J. 1999. Thermophysiology and biology of Giganotosaurus: comparison with Tyrannosaurus. Palaeontologia Electronica 2 (2): 22pp.
- ↑ Barrick, R.E., Stoskopf, M. & Showers, W.J. 1997. Oxygen isotopes in dinosaur bones. In: Farlow, J.O. & Brett-Surman, M. (Eds.). The Complete Dinosaur. Bloomington: Indiana University Press. Pp. 474-490.
- ↑ Paladino, F.V., Spotila, J.R., & Dodson, P. 1997. A blueprint for giants: modeling the physiology of large dinosaurs. En: Farlow, J.O. & Brett-Surman, M. (red.). The Complete Dinosaur. Bloomington: Indiana University Press. Pp. 491-504.
- ↑ Chinsamy, A. & Hillenius, W.J. 2004. Physiology of nonavian dinosaurs. En: Weishampel, D.B., Dodson, P., & Osmolska, H. (red.). The Dinosauria (2nd Edition). Berkeley: University of California Press. Pp. 643-659.
- ↑ Horner, J.R., (1994). Steak knives, beady eyes, and tiny little arms (a portrait of Tyrannosaurus as a scavenger). The Paleontological Society Special Publication 7: 157-164.
- 1 2 Walters, M., Paker, J. (1995). Dictionary of Prehistoric Life. Claremont Books. ISBN 1-85471-648-4.
- ↑ Farlow, J. O. and Holtz, T. R. Jr. 2002. The fossil record of predation in dinosaurs. pp. 251–266, in M. Kowalewski and P. H. Kelley (eds.), The Fossil Record of Predation. The Paleontological Society Papers 8.
- ↑ Dorey, M. (1997). Tyrannosaurus. Dinosaur Cards. Orbis Publishing Ltd. D36045907.
- ↑ Stevens, K.A. (2006) Binocular vision in theropod dinosaurs. Journal of Vertebrate Paleontology 26(2):321-330
- ↑ Tanke, D.H. & Currie, R.J. Head-Biting Behavior in Theropod Dinosaurs: Paleopathological Evidence. May 2000. Gaia 15
- ↑ Goldstone, E. (1997). Injury & Disease, Part 3. Dinosaur Cards. Orbis Publishing Ltd. D36045009.
- ↑ Erickson, G. M., and Olson, K. H. (1996). "Bite marks attributable to Tyrannosaurus rex: preliminary description and implications." Journal of Vertebrate Paleontology, 16(1): 175-178.
- ↑ Carpenter, K. (2000). "Evidence of predatory behavior by carnivorous dinosaurs." Gaia, 15: 135-144.
- ↑ Fowler, D. W., and Sullivan, R. M. (2006). "A ceratopsid pelvis with toothmarks from the Upper Cretaceous Kirtland Formation, New Mexico: evidence of late Campanian tyrannosaurid feeding behavior." New Mexico Museum of Natural History and Science Bulletin, 35: 127-130.
- ↑ "The bigger they come, the harder they fall" New Scientist, October 7 1995, p. 18.
- 1 2 3 Hajdul, R. (1997). Tendons. Dinosaur Cards. Orbis Publishing Ltd. D36044311.
- ↑ Hecht, J. (1998). The deadly dinos that took a dive. New Scientist 2130.
- ↑ . WildlifeSafari.info. Alirita 2006-04-29.
- ↑ . Alirita 2006-04-29.
- 1 2 Hutchinson, J. R. and Garcia, M. (2002). Tyrannosaurus was not a fast runner. Nature 415: 1018-1021
- ↑ . Alirita 11-a de decembro 2005.
- ↑ . Alirita 11-a de decembro 2005.
- ↑ . Dinosauria online.
- ↑ "Tyrannosaurus had poor turning circle" Arkivigite je 2012-05-07 per la retarkivo Wayback Machine Cosmos magazine
- ↑ Osborn, H. F. 1905.Tyrannosaurus and other Cretaceous carnivorous dinosaurs. Bulletin of the American Museum of Natural History 21;259-265
- ↑ White, S. (1997). Tyrannosaurus. Dinosaur Cards. Orbis Publishing Ltd. D36046009.
- ↑ Online guide to the continental Cretaceous-Tertiary boundary in the Raton basin, Colorado and New Mexico. Arkivita el la originalo je 2008-08-20. Alirita 2007-06-21.
- ↑ Guinness World Records Ltd. (2003). 2003 Guinness World Records. pg 90.
- ↑ Currie, P. J., Hurum, J. H., and Sabath, K. 2003. Skull structure and evolution in tyrannosaurid dinosaurs. Acta Palaeontologica Polonica 48: 227-234
- ↑ Croucher, B. (1997). Beast of the Badlands. Dinosaur Cards. Orbis Publishing Ltd. D36045407.
- ↑ Visit Jane.com. Oficala muzea retejo.. Arkivita el la originalo je 2007-09-29. Alirita 2007-06-21.
- ↑ . Alirita 7-a de aprilo 2006.
- ↑ . Alirita 12-a de aprilo 2006.
- ↑ Guinness World Records Ltd. 2003. 2003 Guinness World Records. p. 90.
Literaturo
- Erickson, G. M., Van Kirk, S. D., Su, J., Levenston, M. E., Caler, W. E., kaj Carter, D. R. (1996). “Bite-force estimation for Tyrannosaurus rex from tooth-marked bones.”, Nature, p. 706–708.
- Meers, Mason B. (aŭgusto 2003). “Maximum Bite Force and Prey Size of Tyrannosaurus rex and Their Relationships to the Inference of Feeding Behavior”, Historical Biology: A Journal of Paleobiology 16 (1), p. 1 – 12.
- Schweitzer, Mary H.; Wittmeyer, Jennifer L.; Horner, John R.; Toporski, Jan K. (25-a de marto 2005). “Soft-Tissue Vessels and Cellular Preservation in Tyrannosaurus rex”, Science 307 (5717), p. 1952–1955.
Vidu ankaŭ
- Muzeo Senckenberg - La Muzeo Senckenberg en Frankfurto ĉe Majno (Germanio) havas la plej grandan ekspozicion de grandaj dinosaŭroj en Eŭropo.
Eksteraj ligiloj
- hejma paĝo de Sue Arkivigite je 2007-09-29 per la retarkivo Wayback Machine
- hejma paĝo de Stan Arkivigite je 2007-10-12 per la retarkivo Wayback Machine
- Bildoj de rekonstruaĵo de tiranosaŭro Sue Arkivigite je 2014-06-28 per la retarkivo Wayback Machine
- Historio de la unuaj trovaĵoj de T. rex Arkivigite je 2006-09-28 per la retarkivo Wayback Machine
- Tree of Life diskutas tiranosaŭredojn Arkivigite je 2007-10-12 per la retarkivo Wayback Machine
- Elterigante Tyrannosaurus rex
- juna tiranosaŭro Jane Arkivigite je 2007-09-29 per la retarkivo Wayback Machine
- Cretaceous Hell Creek Faunal Facies Arkivigite je 2008-05-03 per la retarkivo Wayback Machine estas ekzemplo de unu tiranosaŭra medio, en la Formacio Hell Creek de Montano
- Studaĵo de la Universitato de Bristol pri la mordoforto de predantoj Arkivigite je 2006-11-18 per la retarkivo Wayback Machine
- Muzeo de Nenatura Mistero - Mordoforto ktp de T. rex
- Universitato de Tampa pri mordoforto ktp de T. rex
- Universitato Stanford pri la mordoforto de T. rex Arkivigite je 2008-02-27 per la retarkivo Wayback Machine
- Kiel tiranosaŭroj eble seksumis Arkivigite je 2007-11-13 per la retarkivo Wayback Machine