Kiel legi la taksonomion | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Troveblo de fosilioj: Karbonio - ĉi-epoko | ||||||||||||||
 | ||||||||||||||
| Biologia klasado | ||||||||||||||
| ||||||||||||||
 | ||||||||||||||
Aliaj Vikimediaj projektoj | ||||||||||||||

Reptilioj (Reptilia)[1] estas klaso de vertebruloj, kiu enhavas la nuntempajn testudojn, krokodilojn, serpentojn, lacertojn kaj tuatarojn, ties formortintajn parencojn, kaj kelkaj el la formortintaj prauloj de mamuloj. Pro sia evolucia historio kaj la diverseco de formortintaj formoj, la valideco de la klaso ne estas jam universale eltenita en sciencaj fakaj etosoj, kvankam en la praktiko, ĝi restas uzata de kelkaj biologoj kaj plej mezedukitaj homoj, ĉefe ĉe amaskomunikilaro. La studado de reptilioj, historie kombinita kun tiu de amfibioj, estas nomata herpetologio.
Temas pri vertebruloj, kun ŝanĝiĝanta korpotemperaturo, spirantaj per pulmoj. Kelkaj estas senpiedaj (sed descendaj de kvarpieduloj), aliaj posedas kvar mallongajn krurojn, tial ili rampas ĉe la tero kaj estas komunuze nomataj rampuloj pro ĉi tiu kvalito. Reptilohaŭto estas seka kun malgranda nombro da glandoj, epidermo tre korniĝinta; ĝi estas ŝatata por produktado de valizoj.
Reptilioj estas tetrapodoj kaj amniuloj, bestoj kies embrion ĉirkaŭas amnia membrano, kaj membroj de la klaso Sauropsida. Modernaj reptiloj loĝas ĉe ĉiuj kontinentoj kun la escepto de Antarkto. Hodiaŭ ilin reprezentas kvar travivintaj ordoj:
- Krokodiluloj Crocodilia (krokodilo, gavialoj, kajmanoj kaj aligatoroj): 25 specioj
- Sfenodontuloj Sphenodontia (sfenodontoj de Nov-Zelando): 1 specio
- Skvamuloj Squamata (lacertoj, serpentoj kaj amfisbenoj amphisbaenia): ĉirkaŭ 9,600 specioj
- Testuduloj Testudines (testudoj kaj kelonioj): ĉirkaŭ 400 specioj
Malkiel amfibioj, reptiloj ne havas akvolarvan etapon. Plej multe reptiloj estas ovo-demetuloj, kvankam kelkaj specioj de skvamuloj estas vivonaskuloj, kiel estis kelkaj formortintaj akvokladoj[2] —la feto disvolviĝas ene de la patrino, enhavante placenton pli ol ovokonko. Kiel amniuloj, reptiliaj ovoj estas ĉirkaŭataj de membranoj por protektado kaj transporto, kiuj adaptigis ilin al reproduktado sur seka tero. Multaj el la vivonaskaj specioj nutras siajn fetojn tra variaj formoj de placentoj similaj al tiuj de mamuloj, kaj kelkaj havigas komencan zorgon al siaj idoj. Vivantaj reptilioj gamas laŭgrande el eta geko, Sphaerodactylus ariasae, kiu povas kreskiĝis nur ĝis 17 mm al la Mara krokodilo, Crocodylus porosus, kiu povas atingi ĝis 6 m de longo kaj pezas ĉirkaŭ 1000 kg.
Evoluo

Kiam oni konsideras la evoluon de reptilioj ("rampuloj"), estas pli logike grupigi ilin en:
Grupo 1 (testud-devenaj):
- Testuduloj (testudoj);
Grupo 2 (lacert-devenaj):
- Lacertojn (igvanoj, etaj lacertoj, kameleonoj/ĥameleonoj, serpentoj, ktp);
- Krokodiluloj (krokodiloj, aligatoroj, gavialoj) kune kun Birdoj.
Klasigo




La reptilioj estas parafiletika aro. Tio signifas, ke ĝi ne enhavas prapatron kaj ĉiujn ties idojn, ĉar en ili estas la birdoj. Male ĝi estas evolua grado, kuniĝante ulojn kiuj intersimilas forme kaj ekologie. Ili do ne estas aro rigardata en arkreskiĝa klasigo. Kun la birdoj, ili estas la klado Sauropsida.
Klasigo ĝis la nivelo de ordoj, laŭ Benton 2004:
- Serio Amniuloj Amniota
- Klaso Sinapsidoj Synapsida
- Ordo Pelycosauria*
- Ordo Terapsidoj Therapsida
- Klaso Mamuloj Mammalia
- Klaso Reptilioj Sauropsida
- Subklaso Anapsidoj Anapsida
- Ordo Testuduloj Testudines
- Subklaso Diapsidoj Diapsida
- Ordo Araeoscelidia
- Ordo Younginiformes
- Infraklaso Fiŝosaŭroj Ichthyosauria
- Infraklaso Lepidosaŭromorfoj Lepidosauromorpha
- Superordo Saŭropterigoj Sauropterygia
- Ordo Plakodontoj Placodontia
- Ordo Notosaŭroj Nothosauroidea
- Ordo Pleziosaŭruloj Plesiosauria
- Superordo Lepidosaŭroj Lepidosauria
- Ordo Sphenodontida (tuataro)
- Ordo Skvamuloj Squamata (lacertuloj kaj serpentoj)
- Superordo Saŭropterigoj Sauropterygia
- Infraklaso Arkosaŭromorfoj Archosauromorpha
- Ordo Prolacertoformaj Prolacertiformes
- Divizio Arkosaŭroj Archosauria
- Subdivizio Crurotarsi
- Superordo Krokodilomorfoj Crocodylomorpha
- Ordo Krokodiluloj Crocodylia
- Superordo Krokodilomorfoj Crocodylomorpha
- Subdivizio Avemetatarsalia
- Infradivizio Ornitodiroj Ornithodira
- Ordo Flugsaŭroj Pterosauria
- Superordo Dinosaŭroj Dinosauria
- Ordo Saŭriskioj Saurischia
- Infradivizio Ornitodiroj Ornithodira
- Subdivizio Crurotarsi
- Subklaso Anapsidoj Anapsida
- Klaso Birdoj Aves
- Ordo Ornitiskioj Ornithischia
- Klaso Sinapsidoj Synapsida
Lamarko
Lamarko konsideris la reptiliojn (li inkludis amfibiojn ene de la sama grupo) kiel dekdua klaso de organizo de animaloj el la plej simplaj (senvertebruloj) al la plej perfekta (la homo). Li difinis la reptiliojn kiel la unua grupo kun spirado per pulmoj, kvankam ankoraŭ kun malvarma sango, kiel la proksima grupo de fiŝoj, pli proksima al senvertebruloj.
Morfologio kaj fiziologio
Cirkulado

Multaj reptilioj havas tri-ĉambran koron konsista el du koraj aŭrikloj, unu variebla partigita ventriklo, kaj du aortoj kiuj kondukas al oksigenhava cirkulado. La grado de miksado de oksigena kaj neoksigena sangoj en la tri-ĉambra koro varias depende el la specio kaj fiziologia stato. Ĉe diferencaj kondiĉoj, neoksigena sango povas esti revenigita al la korpo aŭ oksigena sango povas esti revenigita al la pulmoj. Tiu variado en sangofluo hipoteze permesas pli efikan termoreguladon kaj pli longajn merĝotempojn ĉe akvaj specioj, sed oni ne pruvis, ke temas pri plitaŭgiga avantaĝo.[3]
Estas esceptoj al la ĝenerala fiziologio. Ekzemple, krokodiloj havas anatomie kvar-ĉambran koron, sed ankaŭ havas du oksigenportajn aortojn kaj estas tiele kapablaj preterpasi nur sian pulman cirkuladon.[4] Ankaŭ, kelkaj serpentaj kaj lacertaj specioj (ekz., pitonoj kaj varanoj) havas tri-ĉambrajn korojn kiuj iĝas porfunkcie kvar-ĉambraj koroj dum kuntiriĝo. Tio eblas pro muskola bordo kiu subdividas la ventriklon dum la ventrikla diastolo kaj komplete dividas ĝin dum la sistolo. Pro tiu bordo, kelkaj el tiuj skvamuloj estas kapablaj produkti ventriklajn premodiferencojn kiuj egalas al tiuj viditaj en mamulaj kaj birdaj koroj.[5]
Metabolo
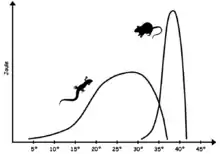
Modernaj reptilioj montras kelkan formon de ektotermiko aŭ sango-malvarmeco, tiele ke ili havas limigitajn fiziologiajn rimedojn elteni la korpotemperaturon konstanta kaj ofte deprende el eksteraj fontoj de varmo. Pro malpli stabila kernotemperaturo ol tiu de birdoj kaj mamuloj, la reptilia biokemio postulas enzimojn kapablajn pluteni efikon super pli granda gamo de temperaturoj ol ĉe la kazo de sango-varmaj animaloj. Optimuma gamo de korpotemperaturo varias laŭ la specioj, sed estas tipe sub tiu de sango-varmaj animaloj; ĉe multaj lacertoj, ĝi falas en la gamo de 24°–35 °C,[6] dum tre varm-adaptitaj specioj, kiaj la uson-meksika dezerta igvano Dipsosaurus dorsalis, povas havi optimumajn fiziologiajn temperaturojn en la mamula gamo, inter 35° kaj 40 °C.[7] Dum la optimuma temperaturo estas ofte trovita kiam la animalo estas aktiva, la malaltaj baza metabolismo faras, ke korpotemperaturo falas rapide kiam la animalo estas neaktiva.
Spirsistemo

Ĉiuj reptilioj spiras uzante pulmojn. Akvaj testudoj disvolvis pli permeablan haŭton, kaj kelkaj specioj modifis sian kloakon por pliigi la areon por gasa interŝanĝo.[8] Eĉ kun tiuj adaptoj, spirado estas neniam tute plenumita sen pulmoj. Pulma aerumado estas plenumita diference en ĉiu ĉefa reptilia grupo. Ĉe skvamuloj, la pulmoj estas aerumitaj preskaŭ ekskluzive pere de la aksa muskolaro. Tiu estas ankaŭ la sama muskolaro kiu estas uzata dum la movado. Pro tiu malfacilaĵo, plej granda parto de skvamuloj devas reteni sian spiradon dum intensa kurado. Kelkaj, tamen, trovis alian rimedon. Varanedoj, kaj ankaŭ kelkaj aliaj specioj de lacertoj, uzas buŝan pumpadon kiel komplementon al sia normala "aksa spirado". Tio ebligas, ke la animaloj komplete plenigu siajn pulmojn dum intensa movado, kaj tiel restas aerobie aktivaj dum longa tempo. Oni scias, ke tupinambaj lacertoj posedas pra-diafragmon, kiu separas la pulmokavaĵon el la interna kavaĵo. Kvankam ĝi fakte ne estas kapabla helpi la movadon, ĝi ebligas pli grandan pulmoŝvelon, forprenante la pezon de la internaĵoj el la pulmoj.[9]
Krokodiloj fakte havas muskolan diafragmon, kiu estas simila al la mamula diafragmo. La diferenco estas ke la muskoloj ĉe la krokodila diafragmo puŝas la pubon (parto de la pelvo, kiu estas movebla ĉe krokodiluloj) reen, kio subenigas la hepaton, tiel liberigante lokon por ke la pulmoj etendiĝu. Tiu tipo de diafragma sistemo estis referencata kiel "hepata piŝto". La aervojoj formas nombrajn duoblajn tuboĉambrojn ene de ĉiu pulmo. Je enspiro kaj elspiro aero moviĝas tra la aervojoj en la sama direkto, tiel kreante unudirektan aerfluon tra la pulmoj. Simila sistemo troviĝas ĉe birdoj,[10] monitorlacertoj[11] kaj igvanoj.[12]
Al plej granda parto de reptilioj mankas duaranga palato, kio rezultas en la fakto, ke ili devas reteni sian spiradon dum englutado. Krokodiluloj evoluis ostecan duarangan palaton kiu ebligas al ili plue spiradi dum restas submerĝitaj (kaj protektas siajn cerbojn kontraŭ damaĝo dum la luktado kontraŭ la predo). Ankaŭ skinkoj (familio Skinkedoj) evoluis ostecan duarangan palaton, je variaj gradoj. Serpentoj adoptas diferencan alproksimiĝon kaj anstataŭe etendas siajn trakeojn. Iliaj trakeaj etendaĵoj elstaras kiel karneca pajleto, kiu ebligas al tiuj animaloj engluti grandajn predojn sen suferin pro asfiksio.[13]
Testudoj kaj emidoj

Kiel testudoj kaj emidoj spiras estis objekto de multa studado. Ĝis nun, nur kelkaj specioj estis sufiĉe studita por havi ideon pri kiel tiuj testudoj spiras. La tre variaj rezultoj indikaj, ke testudoj kaj emidoj trovis varion de solvoj al tiu problemo.
La malfacilo estas ke plej granda parto de testudŝeloj estas rigidaj kaj permesas neniun tipon de etendon aŭ kuntiriĝon kion aliaj amniotoj uzas por aerumi siajn pulmojn. Kelkaj testudoj, kiel la Hindia punktolisemo (Lissemys punctata), havas tavolon de muskolo kiu ĉirkaŭas la pulmojn. Kiam ĝi kuntiriĝas, la testudo povas elspiri. Ripoze, la testudo povas retiri la menbrojn en la korpokavaĵon kaj forpeli aeron el la pulmoj. Kiam la testudo malretiras siajn membrojn, la premo ene de la pulmoj estas reduktita, kaj la testudo povas ensuĉi aeron. La testudaj pulmoj estas ligitaj al la interno de la pinto de la ŝelo (karapaco), kaj la malsupro de la pulmoj ligitaj (pere de konektiva histo) al la cetero de la internaĵoj. Uzante serion de specialaj muskoloj (iom ekvivalenta al diafragmo), testudoj estas kapablaj movi siajn internaĵojn supren kaj malsupren, rezulte en efektiva spirado, ĉar multaj el tiuj muskoloj havas liiglojn kongrue kun ilia antaŭaj membroj (fakte multaj el la muskoloj etendiĝas en membropoŝojn dum la kuntiriĝo).[14]
Spirado dum movado estis studita en tri specioj, kaj ili montris diferencajn modelojn. Plenkreskaj inaj verdaj martestudoj ne spiras dum ili rampas laŭlonge de siaj nestostrandoj. Ili retenas sian spiradon dum surtera movado kaj spiras de tempo en tempo kiel ili ripozas. Nordamerikaj skatoltestudoj spiras kontinue dum movado, kaj la aeruma ciklo ne estas kunordigita kun la membromovado.[15] Tio okazas ĉar ili uzas siajn ventrajn muskolojn por spiri dum la movado. La lasta specio kiu estis studita estas la ruĝorela glittestudo, kiu ankaŭ spiras dum la movado, sed prenas pli malgrandajn spirojn dum la movado ol dum malgrandaj paŭzoj inter movaderoj, kio indikas, ke povas esti mekanika interfero inter la membromovado kaj la spiraparato. Ankaŭ skatoltestudoj estis observitaj kiam ili spiris dum ili estis komplete sigelitaj ene de siaj ŝeloj.[15]
Sonproduktado

Kompare kun ranoj, birdoj kaj mamuloj, reptilioj estas malpli voĉprodukta. Sonproduktado estas kutime limigita al fajfado, kiu estas produktita pere de forpelo de aero tra parte fermita gloto kaj tio ne estas konsiderata kiel vera voĉo. La kapablo voĉi ekzistas en krokodiluloj, kelkaj lacertoj kaj testudoj; kaj tipe konsistas en vibrado de faldecaj strukturoj en la laringo aŭ gloto. Kelkaj gekoj kaj testudoj posedas verajn voĉkordojn, kiuj havas elastin-riĉan konektivan histon.[16][17]
Aŭdado ĉe serpentoj
Aŭdado ĉe homoj dependas el tri partoj de la orelo; la ekstera orelo kiu direktas sonondojn en la orelkanalon, la meza orelo kiu transmisias venintajn sonondojn al la interna orelo, kaj la interna orelo kiu helpas en aŭdado kaj por ekvilibro. Malkiel ĉe homoj kaj ĉe aliaj animaloj, serpentoj posedas nek eksteran orelon, nek mezan orelon, nek timpanon, sed havas internan orelan strukturon kun kokleo rekte konektita al iliaj makzeloj.[18] Ili kapablas senti la vibradon generitan el la sonondoj en siaj makzeloj dum ili moviĝas surgrunde. Tion faras la uzado de mekanikaj riceviloj, sensoricevilaj nervoj kiuj iras laŭlonge de la korpo de serpentoj direktante la vibradon laŭlonge de la spinaj nervoj al la cerbo. Serpentoj havas sensivan aŭdan percepton kaj povas koni el kiu direkto sono farita venas kaj tiel ili povas sensi la proksimecon de predo aŭ de predanto, sed estas ankoraŭ neklare kiel sensivaj serpentoj perceptas la sonondoj kiuj veturas tra la aero.[19]
Haŭto

Reptilia haŭto estas kovrita per korneca epidermo, kio faras ĝin akvoprotekta kaj ebligas, ke reptilioj povas vivi sur seka tero, kontraste kun amfibioj. Kompare kun la mamula haŭto, tiu de reptilioj estas pli fajna kaj al ĝi mankas la dika derma tavolo kiu produktas ledon en mamuloj.[20] Eksponitaj partoj de reptilioj estas protektitaj pere de skvamoj aŭ ŝildaĵoj, foje kun osteca bazo (ostodermoj), formante kvazaŭ armaĵojn. Ĉe lepidosaŭroj, kiel ĉe lacertoj kaj serpentoj, la tuta haŭto estas kovrita per koincidaj epidermaj skvamoj. Tiaj skvamoj estis iam supozitaj tipaj de la klaso Reptilia kiel tuto, sed nun oni scias, ke ili estas nur en lepidosaŭroj. La skvamoj ekzistantaj ĉe testudoj kaj krokodiloj estas de derma, anstataŭ de epiderma, origino kaj estas propre referencataj kiel ŝildetoj. Ĉe testudoj, la korpo estas ŝirmita ene de forta ŝelo komponita de fuzoota ŝildetoj. Ĉar ili ne havas dikan dermon, reptilia ledo ne estas tiom forta kiom la mamula ledo. Ĝi estas fakte uzata en ledaj varoj por dekoracia celo, nome por ŝuoj, zonoj kaj mansakoj, precipe de krokodila haŭto.
Mudo
Reptilioj mudigas sian haŭton pere de procezo nomita "ekdizo" kiu okazas kontinue tra ilia tuta vivo. Pli precize, pli junaj reptilioj tendencas mudi unufoje ĉiun 5an–6an semajnon kaj estante plenkreskuloj 3–4 fojojn jare.[21] Pli junaj reptilioj mudas pli pro siaj pli rapida kreskoproporcio. Post la atingo de plenkreska grando, la frekvenco mudi draste malpliiĝas. La procezo ekdizo konsistas en la formado de nova tavolo de haŭto sub la malnova tavolo. Proteinolizaj enzimoj kaj limfa fluaĵo estas sekreciataj inter la malnova kaj nova tavoloj de haŭto. Poste, tio forigas la malnovan haŭton disde la nova, kio ebligas la okazigon de la mudo.[22] Serpentoj mudas el la kapo ĝis la vosto, dum lacertoj mudas laŭ "makula modelo".[22] Disekdizo, ofta haŭtomalsano ĉe serpentoj kaj lacertoj, okazas kiam ekdizo, aŭ mudo, malsukcesas.[23] Estas nombraj tialoj kial mudo malsukcesas kaj ili povas rilati al maltaŭgaj humideco kaj temperaturo, nutraj mankoj, dehidratiĝo kaj traŭmataj vundoj.[22] Nutraj mankoj malpliigas proteinolizajn enzimojn dum dehidratiĝo malpliigas limfajn fluaĵojn por separi la haŭtotavolojn. Traŭmataj vundoj aliflanke, formas cikatrojn kiuj ne permesas la formadon de novaj skvamoj kaj rompas la procezon de ekdizo.[23]
Ekskrecio
Ekskrecio estas farita ĉefe de du malgrandaj renoj. Ĉe diapsedoj, ureata acido estas la ĉefa nitrogena kromprodukto; testudoj, kiel mamuloj, ekskrecias ĉefe ureon. Malkiel la renoj de mamuloj kaj birdoj, reptiliaj renoj estas malkapablaj produkti likvan ureon pli koncentritan ol ilia korpa fluaĵo. Tio okazas ĉar al ili mankas specializita strukturo nomita Anso de Henle, kiu estas en la nefronoj de birdoj kaj mamuloj. Pro tio, multaj reptilioj uzas la kojlon por helpi en la reabsorbado de akvo. Kelkaj estas ankaŭ kapablaj preni akvon stokitan en la veziko. Ankaŭ troaj saloj estas ekskreciataj pere de nazaj kaj langaj salglandoj en kelkaj reptilioj.
En ĉiuj reptilioj kaj la urinogeneraj duktoj kaj la anuso malpleniĝas en organo nomita kloako. Ĉe kelkaj reptilioj, mezventra mureto en la kloako povas malfermiĝi al urina veziko, sed ne ĉiuj. Ĝi estas en ĉiuj testudoj kaj emidoj same kiel ĉe plej granda parto de lacertoj, sed ĝi mankas ĉe la varanoj, kaj ĉe la senkruraj lacertoj. Ĝi forestas en serpentoj, aligatoroj kaj krokodiloj.[24]
Multaj testudoj, emidoj kaj lacertoj havas proporcie tre grandajn vezikojn. Charles Darwin jam notis, ke la Galapaga testudo havas vezikon kiu povas stoki ĝis 20% de sia korpopezo.[25] Tiaj adaptoj estas la rezulto de medioj kiel foraj insuloj kaj dezertoj kie akvo estas tre malabunda.[26]:143 Aliaj dezert-loĝantaj reptilioj havas grandajn vezikojn kiuj povas stoki longan akvorezervejon por ĝis kelkaj monatoj kaj helpi en osmoregado.[27]
Testudoj havas du aŭ pliajn kromajn urinvezikojn, flanke de la kolo de la urina veziko kaj dorse de la pubo, okupante gravan parton de siaj korpokavaĵo.[28] Iliaj vezikoj estas ankaŭ kutime dulobigitaj kun maldekstra kaj dekstra sekcioj. La dekstra sekcio estas sub la hepato, kiu evitas, je grandaj ŝtonoj restu en tiu loko dum la maldekstra sekcio plej verŝajne povas havi vezikajn ŝtonojn.[29]





Referencoj
- ↑ reptilioj en vortaro.net
- ↑ Sander, P. Martin. (2012). “Reproduction in early amniotes”, Science 337 (6096), p. 806–808. doi:10.1126/science.1224301.
- ↑ (2002) “The Physiological and Evolutionary Significance of Cardiovascular Shunting Patterns in Reptiles”, News in Physiological Sciences 17, p. 241–245.
- ↑ (1997) “From anatomy to angioscopy: 164 years of crocodilian cardiovascular research, recent advances, and speculations”, Comparative Biochemistry and Physiology A 188 (1), p. 51–62. doi:10.1016/S0300-9629(96)00255-1.
- ↑ (2003) “Ventricular haemodynamics in Python molurus: separation of pulmonary and systemic pressures”, The Journal of Experimental Biology 206 (Pt 23), p. 4242–4245. doi:10.1242/jeb.00681.
- ↑ Huey, R.B. & Bennett, A.F. (1987): Phylogenetic studies of coadaptation: Preferred temperatures versus optimal performance temperatures of lizards. Evolution No. 4, vol 5: paĝoj 1098-1115 PDF Arkivigite je 2022-09-11 per la retarkivo Wayback Machine
- ↑ Huey, R.B. (1982): Temperature, physiology, and the ecology of reptiles. Side 25-91. In Gans, C. & Pough, F.H. (red), Biology of the Reptili No. 12, Physiology (C). Academic Press, London.artikkel Arkivigite je 2022-04-19 per la retarkivo Wayback Machine
- ↑ Orenstein, Ronald. (2001) Turtles, Tortoises & Terrapins: Survivors in Armor. Firefly Books. ISBN 978-1-55209-605-5.
- ↑ (2003) “Structure of the posthepatic septum and its influence on visceral topology in the tegu lizard, Tupinambis merianae (Teidae: Reptilia)”, Journal of Morphology 258 (2), p. 151–157. doi:10.1002/jmor.10136.
- ↑ (2010) “Unidirectional airflow in the lungs of alligators”, Science 327 (5963), p. 338–340. doi:10.1126/science.1180219.
- ↑ (2013) “Unidirectional pulmonary airflow patterns in the savannah monitor lizard”, Nature 506 (7488), p. 367–370. doi:10.1038/nature12871.
- ↑ (2014) “New insight into the evolution of the vertebrate respiratory system and the discovery of unidirectional airflow in iguana lungs”, Proceedings of the National Academy of Sciences 111 (48), p. 17218–17223. doi:10.1073/pnas.1405088111.

- ↑ (Januaro 1982) “Gross anatomy of snakes.”, Veterinary Medicine/Small Animal Clinician.
- ↑ (2014) “Origin of the unique ventilatory apparatus of turtles”, Nature Communications 5, p. 5211. doi:10.1038/ncomms6211.
- 1 2 (2003) “Lung ventilation during treadmill locomotion in a terrestrial turtle, Terrapene carolina”, Journal of Experimental Biology 206 (19), p. 3391–3404. doi:10.1242/jeb.00553.
- ↑ Russell, Anthony P.; Bauer, Aaron M. (2020). “Vocalization by extant nonavian reptiles: A synthetic overview of phonation and the vocal apparatus”, The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology 304 (7), p. 1478–1528. doi:10.1002/ar.24553.
- ↑ Capshaw, Grace; Willis, Katie L.; Han, Dawei; Bierman, Hilary S.. (2020) Rosenfeld, Cheryl S.; Hoffmann, Frauke: Neuroendocrine Regulation of Animal Vocalization. Academic Press, p. 101–118. ISBN 978-0128151600.
- ↑ Christensen, C. B., Christensen-Dalsgaard, J., Brandt, C., & Madsen, P. T. (2012). Hearing with an atympanic ear: Good vibration and poor sound-pressure detection in the royal python, Python regius. Journal of Experimental Biology, 215(2), 331–342. https://doi.org/10.1242/jeb.062539
- ↑ YOUNG, BRUCE A. “A REVIEW OF SOUND PRODUCTION AND HEARING IN SNAKES, WITH A DISCUSSION OF INTRASPECIFIC ACOUSTIC COMMUNICATION IN SNAKES.” Journal of the Pennsylvania Academy of Science, vol. 71, no. 1, 1997, pp. 39–46, http://www.jstor.org/stable/44149431. Alirita la 8an de Majo 2022
- ↑ Hildebran, M. & Goslow, G. (2001): Analysis of Vertebrate Structure. 5th edition. John Wiley & sons inc, New York. 635 pp. (ISBN 978-0-471-29505-1)
- ↑ Paterson, Sue. (17a de Decembro 2007) Skin Diseases of Exotic Pets. Blackwell Science, Ltd., p. 74–79. ISBN 9780470752432.
- 1 2 3 (Julio 2012) “Dermatological Diseases in Lizards”, The Veterinary Journal 193 (1), p. 38–45. doi:10.1016/j.tvjl.2012.02.001.
- 1 2 Girling, Simon. (26a de Junio 2013) Veterinary Nursing of Exotic Pets, 2‑a eldono, Blackwell Publishing, Ltd.. ISBN 9781118782941.
- ↑ Herbert W. Rand. (1950) The Chordates. Balkiston.
- ↑ P.J. Bentley. (14-a de marto 2013) Endocrines and Osmoregulation: A Comparative Account in Vertebrates. Springer Science & Business Media. ISBN 978-3-662-05014-9.
- ↑ Paré, Jean (11a de Januaro 2006). “Reptile Basics: Clinical Anatomy 101”, Proceedings of the North American Veterinary Conference 20, p. 1657–1660.
- ↑ (2007-04-15) “The urinary bladder as a physiological reservoir that moderates dehydration in a large desert lizard, the Gila monster Heloderma suspectum”, Journal of Experimental Biology (en) 210 (8), p. 1472–1480. doi:10.1242/jeb.003061.
- ↑ (Februaro 2015) “Urogenital System”, Anatomy of Sea Turtles 1, p. 153–165.
- ↑ Divers, Stephen J.. (2005) Reptile Medicine and Surgery. Amsterdam: Elsevier Health Sciences, p. 481, 597. ISBN 9781416064770.
Literaturo
- Lamarko (Lamarck), Filozofio zoologia, 1809. SAT-Broŝurservo, 1987. Tradukis Valo. (paĝo 127).
- Colbert, Edwin H. (1969). Evolution of the Vertebrates (2nd ed.). New York: John Wiley and Sons Inc. ISBN 0-471-16466-6.
- Klein, Wilfied; Abe, Augusto; Andrade, Denis; Perry, Steven (2003). "Structure of the posthepatic septum and its influence on visceral topology in the tegu lizard, Tupinambis merianae (Teidae: Reptilia)". Journal of Morphology 258 (2): 151–157. doi:10.1002/jmor.10136. PMID 14518009.
- Landberg, Tobias; Mailhot, Jeffrey; Brainerd, Elizabeth (2003). "Lung ventilation during treadmill locomotion in a terrestrial turtle, Terrapene carolina". Journal of Experimental Biology 206 (19): 3391–3404. doi:10.1242/jeb.00553. PMID 12939371.
- Laurin, Michel kaj Gauthier, Jacques A.: Diapsida. Lizards, Sphenodon, crocodylians, birds, and their extinct relatives Arkivigite je 2013-05-05 per la retarkivo Wayback Machine, Versio de 22a de Junio 2000; parto de The Tree of Life Web Project
- Orenstein, Ronald (2001). Turtles, Tortoises & Terrapins: Survivors in Armor. Firefly Books. ISBN 1-55209-605-X.
- Pianka, Eric; Vitt, Laurie (2003). Lizards Windows to the Evolution of Diversity. University of California Press. pp. 116–118. ISBN 0-520-23401-4.
- Pough, Harvey; Janis, Christine; Heiser, John (2005). Vertebrate Life. Pearson Prentice Hall. ISBN 0-13-145310-6.
Eksteraj ligiloj
- http://tolweb.org/tree?group=Amniota&contgroup=Terrestrial_Vertebrates Arkivigite je 2007-09-27 per la retarkivo Wayback Machine
- http://www.nhf.dk/
- http://cheni.0086e.net/qiu/class_reptile/ Arkivigite je 2005-10-27 per la retarkivo Wayback Machine
- LAST: LITOVIO: Reptilioj en la domo el Monato (2002/11, p. 16)
- Reptilia Filogenetiko
- Reptiliaj bildoj
- Sri Lanka Wild Life Information Database
- Biologio de Reptilioj estas reta kopio de la kompleta teksto de 22voluma 13,000-paĝa studo pri la stato de enketado pri reptilioj.