| Ĉi tiu artikolo temas pri ordo de bestoj. Por eklezia titolo rigardu la paĝon Primaso. |
Kiel legi la taksonomion | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Troveblo de fosilioj: Plej malfrua Kretaceo - Nun | ||||||||||||||
 | ||||||||||||||
| Biologia klasado | ||||||||||||||
| ||||||||||||||
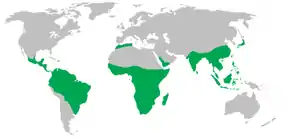 Arealo de ne-homaj primatoj Arealo de ne-homaj primatoj | ||||||||||||||
Familioj | ||||||||||||||
|
vidu tekston | ||||||||||||||
Aliaj Vikimediaj projektoj | ||||||||||||||

Primatoj estas ordo de mamuloj. La termino devenas el latina vorto por "unua rango".[1][2] En taksonomio, primatoj inkludas prosimiulojn kaj simiulojn.[3] Primatoj aperis el prauloj kiuj vivis en arboj de tropikaj arbaroj; multaj primataj karakteroj reprezentas adaptaĵojn al la vivo en tiu defia tri-dimensia medio. La plimulto de primataj specioj restas almenaŭ parte arboloĝantoj.
Kun la escepto de homoj, kiuj loĝas en ĉiu kontinento, la plimulto de primatoj loĝas en tropikaj aŭ subtropikaj regionoj de Ameriko, Afriko kaj Azio.[4] Ili gamas laŭgrandece el Berta muslemuro, kiu pezas nur 30 g, al la Orienta gorilo de malaltaj teroj, kiu pezas ĉirkaŭ 200 kg. Baze sur fosilia pruvaro, la plej fruaj konataj veraj primatoj, reprezentataj de la genro Teilhardina, datas el antaŭ 55.8 milionoj da jaroj.[5] Frua proksima primata parenco konata el abundaj restoj estas de fina Paleoceno Plesiadapis, de antaŭ ĉirkaŭ 55–58 milionoj da jaroj.[6] Studoj per molekula horloĝo sugestas, ke primata branĉo povas esti eĉ pli antikva, origine en la mezo de la periodo de Kretaceo de antaŭ ĉirkaŭ 85 ma.[6]
La ordo Primatoj estis tradicie dividita en du ĉefaj grupoj: prosimiuloj kaj antropoidoj (simiuloj). Prosimiuloj havas karakterojn pli kiel tiuj de la plej fruaj primatoj, kaj inkludas la lemurojn de Madagaskaro, la lorisojn, kaj la tarsiedojn. Simiuloj inkludas simiojn, grandsimiojn kaj homojn. Pli ĵuse, taksonomiistoj preferis disigi primatojn en la subordo Strepsirinoj, aŭ malseknazaj primatoj, konsistaj el ne-tarsiedaj prosimiuloj, kaj la subordo Haplorinoj, aŭ seknazaj primatoj, konsistaj el tarsiedoj kaj simiuloj. Simiuloj estas dividataj en du grupoj: katarinoj (mallarĝ-nazaj) simioj kaj grandsimioj de Afriko kaj sudorienta Azio kaj platirinoj ("eben-nazaj") aŭ Amerikaj simioj de Suda kaj Centra Ameriko. Katarinoj konsistas el Cerkopitekedoj aŭ Malnovmondaj simioj (kiaj pavianoj kaj makakoj), gibonoj kaj Homedoj aŭ grandsimioj aŭ senvostaj simioj; Amerikaj simioj inkludas la kapuĉulojn, hurlulojn kaj sciur-simiojn. Homoj estas la ununuraj vivantaj katarinoj kiuj disvastiĝis sukcese for de Afriko, Suda Azio, kaj Orienta Azio, kvankam fosilia pruvaro montras ke multaj aliaj specioj estis iam ĉeestaj en Eŭropo. Novaj primataj specioj estas ankoraŭ malkovrotaj. Pli ol 25 specioj estis taksonomie priskribitaj en la jardeko de la 2000-aj jaroj kaj dekunu estis priskribitaj ekde la jardeko de la 2010-aj jaroj.
Konsiderita ĝeneralistaj mamuloj, primatoj elmontras larĝan gamon de karakterizaĵoj. Kelkaj primatoj (inkluzive de kelkaj homsimioj kaj pavianoj) estas ĉefe surteremaj prefere ol surarbemaj, sed ĉiuj specioj posedas adaptiĝojn por surgrimpado de arboj. Movadoteknikoj uzitaj inkludas salti de arbo ĝis arbo, piediradon sur du aŭ kvar membroj, maniradoj, kaj saltadon inter branĉoj de arboj (pendiro aŭ braksvingado). Primatoj estas karakterizitaj per grandaj cerboj relative al aliaj mamuloj, same kiel pliigita dependeco en stereoskopa vidkapablo prefere al flarsento, la domina sensa sistemo en la plej multaj mamuloj. Tiuj ecoj estas plievoluintaj en simioj kaj homsimioj kaj videble malpli en lorisoj kaj lemuroj. Trikolora vidkapablo formiĝis en kelkaj primatoj. La plej granda parto ankaŭ havas kontraŭstareblajn dikfingrojn kaj kelkaj havas prenipovajn vostojn. Multaj specioj estas sekse dimorfaj; diferencoj inkludas korpomason, kaningrandecon, kaj kolorigon. Primatoj havas pli malrapidajn rapidecojn de evoluo ol aliaj simile grandaj mamuloj kaj atingas maturecon poste, sed havas pli longajn vivotempojn. Depende de la specio, plenkreskuloj povas vivi en soleco, en parigitaj paroj, aŭ en grupoj de ĝis centoj da membroj.

Anatomio kaj fiziologio
Kapo

La primata kranio havas grandan, kupolhavan kranion, kiu estas aparte elstara ĉe antropoidoj. Tiu kranio protektas la grandan cerbon, distingitan karakteron de tiu grupo.[7] La endokrania volumo (nome la volumo ene de la kranio) estas trifoje pli granda ĉe homoj ol en la pli grandaj nehomaj primatoj, kio montras pli grandan cerbograndon.[8] La averaĝa endokrania volumo estas de 1 201 kubaj centimetroj ĉe homoj, 469 cm3 en goriloj, 400 cm3 en ĉimpanzoj kaj 397 cm3 en orangutanoj.[8] La ĉefa evolucia tendenco de primatoj estis la kreado de la cerbo, kaj pli precize de la novkortekso (parto de la cerba kortekso), kiu rilatas al sensa perceptado, generado de movordonoj, spaca rezonado, konscia pensaro kaj, ĉe homoj, parolkapablo.[4] Dum aliaj mamuloj ege dependas el sia flarkapablo, la arborema vivmaniero de primatoj kondukis ilin al ĉefe tuŝa, vida dominanta sensosistemo,[4] redukto en la flar-regiono de la cerbo kaj pliiĝanta kompleksa socia kutimaro.[9]
Primatoj havas antaŭ-direktitajn okulojn sur la fronto de la kranio; duokula vidkapablo ebligas akuratan distancopercepton, tre utilan por la braksvingemaj prauloj de ĉiuj grandaj simioj.[7] Osta bordo super la okuloj plinfortigas la pli malfortajn ostojn de la vizaĝo kiu suferas fortojn dum maĉado. Strepsirinoj havas superorbitan ostostrion, nome osto ĉirkaŭ la okulkavo, por protekti siajn okulojn; kontraste, la pli altaj primatoj, nome haplorinoj, evoluigis tute fermitajn okulkavojn.[10]

Primatoj montras evolucian tendencon al reduktita muzelo.[11] Teknike, malnovmondaj simioj estas distingeblaj disde la novmondaj simioj per la strukturo de la nazo, kaj el grandaj simioj per la aranĝo de siaj dentoj.[9] En novmondaj simioj, la naztruoj frontas flanken; en malnovmondaj simioj, ili frontas suben.[9] La dentara modelo ĉe primatoj varias konsiderinde; kvankam kelkaj perdis plej el siaj incizivoj, sed ĉiuj retenas almenaŭ unu malsupran incizivon.[9] Ĉe plej strepsirinoj, la malsupraj incizivoj formas dentarkombilon, kiu estas uzata por aranĝado kaj foje por manĝado.[9][12] Malnovmondaj simioj havas ok premolarojn, kompare kun 12 ĉe la novmondaj simioj. La specioj de malnovmondaj simioj estas dividataj en simioj kaj grandaj simioj depende de la nombro de bordopintoj en siaj molaroj: simioj havas kvar, grandsimioj havas kvin[9] - sed homoj povas havi kvar aŭ kvin.[13] La ĉefe homeda molara bordopinto (hipokonuso) evoluis ĉe la frua primata historio, dum la bordopinto de la koresponda primitiva malsupra molaro (parakonusedo) perdiĝis. Prosimiuloj estas distingeblaj pro siaj senmovaj supraj lipoj, la malseka pinto de siaj nazoj kaj la antaŭ-frontaj malsupraj antaŭaj dentoj.
Korpo
Primatoj ĝenerale havas kvin fingrojn en ĉiu membro (kvindaktileco), kun karaktera tipo de keratina ungo sur la fino de ĉiu fingro kaj piedfingro. La malsupraj flankoj de manoj kaj piedoj havas tuŝsensajn kusenetojn ĉe la falangopintoj. Plej havas oponeblajn dikfingrojn, karaktera primata trajto plej disvolvigita ĉe homoj, kvankam ne limigitaj al tiu ordo (ankaŭ didelfoj kaj koaloj, por ekzemplo, havas ilin).[7] Dikfingroj permesas al kelkaj specioj uzi ilojn. Ĉe primatoj, la kombino de opoziciantaj dikfingroj, mallongaj fingrungoj (anstataŭ krifoj) kaj longan, interne-fermeblaj fingroj estas restaĵo de la prapraktiko kapti branĉojn, kaj tio parte ebligis, ke kelkaj specioj disvolvis braksvingadon (svingado per brakoj el arbobranĉo al arbobranĉo) kiel grava rimedo ekhavi movokapablon. Prosimiuloj havas krifecajn ungojn sur la dua fingro de ĉiu piedo, nomitaj purigungoj, kiujn ili uzas por memaranĝado.[7]

La primata kolumosto estas elstara elemento de la brusta krono; tio permesas al la ŝultraj artikoj tre ampleksan moveblon.[11] Kompare kun la malnovmondaj simioj, la grandaj simioj havas pli moveblajn ŝultrajn artikojn kaj brakojn pro la dorsa sinteno de la skapoloj, grandajn ripujojn kiuj estas pli ebenaj antaŭ-malantaŭen, pli mallongan, malpli moveblan spinon, kun la pli malsupraj vertebroj ege reduktitaj - rezulte en vostoperdo en kelkaj specioj. Pren-povaj vostoj estas en la nomvmondaj ateledoj, kiel ĉe la hurluloj, aranesimioj, lanaranesimioj, lansimioj; kaj ĉe cebenoj.[14][15] Masklaj primatoj havas pendantajn penisojn kaj testikojn descendintajn en skroto.[12][16]
Seksa dimorfismo

Seksa dimorfismo estas ofte vidita en simiuloj, kvankam je pli granda grado en malnovmondaj specioj (grandaj simioj kaj kelkaj simioj) ol en novmondaj specioj. Ĵusaj studoj per komparo de DNA por ekzameni kaj la variadon en la esprimo de la seksa dimorfismo inter primatoj kaj la fundamentajn kaŭzojn de seksa dimorfismo. Primatoj kutime havas dimorfismon en la korpopezo[17][18][19] kaj en la grando de kaninoj[20][21][22] kun la felo kaj haŭtkoloro.[23] La dimorfismo povas esti atribuita al kaj tuŝita de diferencaj faktoroj, kiel pariĝado,[17][24] grando,[24] habitato kaj dieto.[25]
Perkomparaj analizoj generis pli kompletan komprenon de la rilatoj inter seksa selekto, natura selekto, kaj pariĝadaj sistemoj en primatoj. Studoj montris, ke dimorfismo estas la produkto de ŝanĝoj kaj en masklaj kaj en inaj trajtoj.[17][26] Ontogenetika skalado, laŭ kiu okazas relativa etendo de komuna kreskotrajektorio, povas havigi ioman vidon en la rilataro inter seksa dimorfismo kaj kreskomodeloj.[27] Kelka informaro el la fosilia registro sugestas, ke estis konverĝa evoluo de dimorfismo, kaj kelkaj formortintaj homedoj probable havis pli grandajn dimorfismojn ol ĉe ajna nuntempe vivanta primato.[26]
Movmaniero

Primataj specioj moviĝas per braksvingado, dupiedado, saltado, kaj arba kaj surtera kvarpiedado, grimpado, fingrartika-piedirado aŭ tre ofte per kombinado de tiuj metodoj. Kelkaj prosimiuloj estas ĉefe vertikale grimpistoj kaj saltuloj. Tiuj estas multaj galagoj, ĉiuj indriedoj (t.e., propitekoj, avahioj kaj indrioj), lepilemuroj, kaj ĉiuj tarsiedoj.[28] Aliaj prosimiuloj estas arbaj kvarpieduloj kaj grimpistoj. Kelkaj estas ankaŭ surteraj kvarpieduloj, dum aliaj estas saltuloj. Plej simioj estas kaj arbaj kaj surteraj kvarpieduloj kaj grimpistoj. Gibonoj, braĥiteloj kaj aranesimioj estas ĉiuj braksvinguloj etende,[29] kaj gibonoj foje faras tion laŭ rimarkinde akrobata maniero. Ankaŭ Lagotriksoj braksvingas foje.[30] Orangutanoj uzas similan formon de movmaniero nomita kvarmana grimpado, en kiu ili uzas siajn brakojn kaj krurojn por porti pezajn korpojn tra la arboj.[29] Ĉimpanzoj kaj goriloj fingrartike piediras,[29] kaj povas moviĝi dupiede dum mallongajn distancojn. Kvankam nombraj specioj, kiel membroj de Australopithecina kaj fruaj homedoj, montris plenan dupiedan movmanieron, homoj estas la nura nuntempe vivanta specio havanta tiun trajton.[31]
Vidkapablo

La evolucio de la kolora vidkapablo en primatoj estas unika inter plej el la eŭteriaj mamuloj. Kvankam la malproksimaj vertebrulaj prauloj de la primatoj posedis trikoloran vidkapablon (triĥromatio), la noktulaj, varmsangaj, mamulaj prauloj perdis unu el la tri konusojn de la retino dum la epoko de la Mezozoiko. Fiŝoj, reptilioj kaj birdoj estas tial triĥromataj aŭ tetraĥromataj, dum aliaj mamuloj, kun la escepte pri kelkaj primatoj kaj marsupiuloj,[32] estas duĥromatoj aŭ unuĥromatoj (totale kolorblindaj).[12] Noktulaj primatoj, kiel la noktosimioj kaj la galagoj, estas ofte monoĥromaaj. Katarinoj estas rutine triĥromataj ĝis genduobliĝo de la ruĝ-verda opsina geno ĉe la bazo de ties stirpo, antaŭ 30 ĝis 40 milionoj da jaroj.[12][33] Platirinoj, aliflanke, estas triĥromataj nur en kelkaj malmultaj kazoj.[34] Specife, unuopaj inoj devas esti heterozigota por du aleloj de la opsina geno (ruĝa kaj verda) en la sama genetika loko de la Kromosomo X.[12] Maskloj, siavice, povas nur esti duĥromataj, dum inoj povas esti ĉu duĥromataj aŭ triĥromataj. Kolora vidkapablo ĉe strepsirinoj ne estas bone komprenita; tamen, esplorado indikas gamon de kolora vidkapablo simila al tiu trovita ĉe platirinoj.[12]
Kiel katarinoj, ankaŭ hurluloj (familio de platirinoj) montras rutine triĥromatecon kiu estis sekvita ĝis evolucia ĵusa genduobliĝo.[35] Hurluloj estas grupo el plej specializitaj folimanĝuloj inter la novmondaj simioj; fruktoj ne estas ĉefa parto de iliaj dietoj,[30] kaj la tipo de folioj kiujn ili preferas konsumi (novaj, nutraj, kaj digesteblaj) estas detekteblaj nur per ruĝ-verda signalo. Kampolaboro esploranta la dietaj preferencoj de hurluloj sugestas, ke rutina triĥromateco estas produkto de selekto pro la medio.[34]
Taksonomio
- ORDO PRIMATOJ
- Subordo Strepsirinoj: lemuroj, lorisoj, ktp.
- Subordo Haplorinoj: tarsioj, simioj, grandsimioj
- Infraordo Tarsioformaj
- Familio Tarsiedoj: tarsioj
- Infraordo Simioformaj: simioj, grandsimioj
- Mikroordo Platirinoj: Amerikaj simioj
- Familio Kalitriksedoj: kalitriksoj, tamarinoj
- Familio Cebedoj: kapuĉuloj, sciursimioj
- Familio Aotedoj: noktosimioj
- Familio Piteciedoj: sakioj, barbosakioj, uakarioj, kaliceboj
- Familio Ateledoj: hurluloj, aransimioj, kaj lansimioj
- Flavvosta lansimio, Oreonax flavicauda
- Mikroordo Katarinoj Malnovmondaj antropoidoj
- Superfamilio Cercopithecoidea: Malnovmondaj simioj
- Familio Cerkopitekedoj
- Superfamilio Homoideoj: grandsimioj
- Familio Hilobatedoj: malgrandaj grandsimioj (gibonoj)
- Familio Homedoj: grandsimioj kaj homoj
- Superfamilio Cercopithecoidea: Malnovmondaj simioj
- Mikroordo Platirinoj: Amerikaj simioj
- Infraordo Tarsioformaj
Arbeto



| Primatoj (Primates) |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
- N.N.
- ne nomata (nomen nudum)
Ordo primatoj (Primates)
La simbolo † indikas grupon nun elmortintan.
subordo prasimioj (Prosimiae)
superfamilio † plesiadapoideoj (Plesiadapoidea)
- familio † plesiadapedoj (Plesiadapidae)
- familio † karpolestedoj (Carpolestidae)
- familio † paromomiedoj (Paromomyidae)
superfamilio lemuroideoj (Lemuroidea)
- familio † adapedoj (Adapidae)
- familio lemuredoj (Lemuridae)
- familio indriedoj (Indriidae)
- familio daŭbentoniedoj (Daubentoniidae)
- familio † arĥeolemuredoj (Archaeolemuridae)
superfamilio lorisoideoj (Lorisoidea)
- familio lorisedoj (Lorisidae)
superfamilio tarsioideoj (Tarsioidea)
- familio † anaptomorfedoj (Anaptomorphidae)
- familio † omomiedoj (Omomyidae)
- familio tarsiedoj (Tarsiidae)
subordo simiuloj (antropoidoj) (Simiae = Anthropoidea)
superfamilio ceboideoj (okcidentaj simiuloj) (Ceboidea)
- familio kalitriksedoj (Callithricidae)
- familio cebedoj (Cebidae)
superfamilio cerkopitekoideoj (orientaj simiuloj) (Cercopithecoidea)
- familio cerkopitekedoj (Cercopithecidae)
superfamilio † parapitekoideoj (Parapithecoidea)
- familio † parapitekedoj (Parapithecidae)
superfamilio homoideoj (Hominoidea)
Esplorado
Primatologio estas fako de zoologio, kiu temas pri primatoj. Ĝi estas diversa fako ĉe la limo inter mamalogio kaj antropologio. Primatologoj studas kaj vivantajn kaj formortintajn speciojn de primatoj en la naturaj vivejoj kaj en laboratorioj, kaj ili faras eksperimentojn por pli bone kompreni ilin.
Primatoj en homa kulturo
Jen kelkaj romanoj kie primatoj ludas ŝlosilan rolon:
- Les Animaux dénaturés de Vercors.
- La Grandaj Simioj de Will Self.
- La Planète des singes de Pierre Boulle.
- Evoluo de Stephen Baxter.
Vidu ankaŭ
Referencoj
- ↑ Primate, Merriam-Webster Online Dictionary, Merriam-Webster http://www.merriam-webster.com/dictionary/primate 2008-07-21
El Malnovfranca lingvo aŭ moderna francaprimat, el substantiva uzado de la latina primat-, el primus ("unua rango") - ↑ La angla singularo primate estis derivita tra reena formado el la modernlatina formo kiun Carl Linnaeus enkondukis en sia influa verko de 1758 nome 10a eldono de Systema Naturae ĉar li pensis, ke tiu estas la "plej alta" ordo de mamuloj.
- ↑ Goodman, M., Tagle, D. A., Fitch, D. H., Bailey, W., Czelusniak, J., Koop, B. F., Benson, P. & Slightom, J. L. (1990). “Primate evolution at the DNA level and a classification of hominoids”, Journal of Molecular Evolution 30 (3), p. 260–266. doi:10.1007/BF02099995.
- 1 2 3 en Encyclopædia Britannica, alirita la 23an de Aŭgusto 2022.
- ↑ O'Leary, M. A. (2013). “The placental mammal ancestor and the post–K-Pg radiation of placentals”, Science 339 (6120), p. 662–667. doi:10.1126/science.1229237.
- 1 2 Helen J Chatterjee, Simon Y.W. Ho, Ian Barnes & Colin Groves (2009). “Estimating the phylogeny and divergence times of primates using a supermatrix approach”, BMC Evolutionary Biology 9, p. 259. doi:10.1186/1471-2148-9-259.
- 1 2 3 4 [1979] (2005) “Characteristics of Primates”, Vertebrate Life, 7‑a eldono, Pearson, p. 630. ISBN 0-13-127836-3.
- 1 2 (1990) An Introduction to Human Evolutionary Anatomy. Academic Press, p. 193. ISBN 0-12-045590-0.
- 1 2 3 4 5 6 Myers, P. (1999). "Primates" (On-line). Animal Diversity Web. Alirita 2008-06-03.
- ↑ (2000) Humankind Emerging, 8‑a eldono, Allyn & Bacon, p. 85. ISBN 0-673-52364-0.
- 1 2 Archonta: Primates. Palaeos (2006-01-08). Arkivita el la originalo je 2008-05-12. Alirita 2008-06-03.
- 1 2 3 4 5 6 “Primates”, The Encyclopedia of Mammals.
- ↑ Ash, M. M.. (2003) Wheeler's Dental Anatomy, Physiology, and Occlusion. W.B. Saunders. ISBN 978-0-7216-9382-8.
- ↑ (November 1999) “The ecological role of the prehensile tail in white-faced capuchins (Cebus capucinus)”, American Journal of Physical Anthropology 110 (3), p. 325–39. doi:[[doi:10.1002%2F%28SICI%291096-8644%28199911%29110%3A3%3C325%3A%3AAID-AJPA5%3E3.0.CO%3B2-D|10.1002/(SICI)1096-8644(199911)110:3<325::AID-AJPA5>3.0.CO;2-D]].
- ↑ (Novembro 2011) “Tail growth tracks the ontogeny of prehensile tail use in capuchin monkeys (Cebus albifrons and C. apella)”, American Journal of Physical Anthropology 146 (3), p. 465–73. doi:10.1002/ajpa.21617.
- ↑ Friderun Ankel-Simons. (27a de Julio 2010) Primate Anatomy: An Introduction. Academic Press, p. 442, 521. ISBN 978-0-08-046911-9.
- 1 2 3 (1998) “Phylogenetic analyses of primate size evolution: the consequences of sexual selection”, Biological Journal of the Linnean Society (en) 64 (4), p. 413–447. doi:10.1111/j.1095-8312.1998.tb00342.x.
- ↑ Ralls, K. (1976). “Mammals in Which Females are Larger Than Males”, The Quarterly Review of Biology 51 (2), p. 245–76. doi:10.1086/409310.
- ↑ Lindstedtand & Boyce (Julio 1985). “Seasonality, Fasting Endurance, and Body Size in Mammals”, The American Naturalist 125 (6), p. 873. doi:10.1086/284385.
- ↑ (2006) “Phylogenetic analyses of dimorphism in primates: Evidence for stronger selection on canine size than on body size”, American Journal of Physical Anthropology (en) 130 (1), p. 50–59. doi:10.1002/ajpa.20321.
- ↑ Frisch, J. E. (1963). “Sex-differences in the canines of the gibbon (Hylobates lar)”, Primates 4 (2), p. 1–10. doi:10.1007/BF01659148.
- ↑ Kay, R. F. (1975). “The functional adaptations of primate molar teeth”, American Journal of Physical Anthropology 43 (2), p. 195–215. doi:10.1002/ajpa.1330430207.
- ↑ Crook, J. H.. (1972) “Sexual selection, dimorphism, and social organization in the primates”, Campbell, B. G.: Sexual selection and the descent of man. Aldine Transaction, p. [htt://archive.org/details/sexualselection00came/246 246]. ISBN 978-0-202-02005-1.
- 1 2 (Novembro 1985) “The quantitative assessment of phylogenetic constraints in comparative analyses: Sexual dimorphism in body weight among primates”, Evolution 39 (6), p. 1335–1351. doi:10.2307/2408790.
- ↑ (1982) “Correlates of sexual dimorphism in primates: Ecological and size variables”, International Journal of Primatology 3 (4), p. 387–402. doi:10.1007/BF02693740.
- 1 2 Plavcan, J. M. (2001). “Sexual dimorphism in primate evolution”, American Journal of Physical Anthropology 33, p. 25–53. doi:10.1002/ajpa.10011.
- ↑ (2002) “Sexual dimorphism and facial growth in papionine monkeys”, Journal of Zoology 257 (2), p. 255–72. doi:10.1017/S0952836902000857.
- ↑ Sussman, R. W.. (1999) Primate Ecology and Social Structure Volume 1: Lorises, Lemurs and Tarsiers. Needham Heights, MA: Pearson Custom Publishing & Prentice Hall, p. [htt://archive.org/details/imateecologyso0001suss/e/78 78, 89–90, 108, 121–123, 233]. ISBN 0-536-02256-9.
- 1 2 3 Strier, K. (2007). Primate Behavioral Ecology (3a eld.). Allyn & Bacon. pp. 7, 64, 71, 77, 182–185, 273–280, 284, 287–298. ISBN 978-0-205-44432-8.
- 1 2 Sussman, R. W.. (2003) Primate Ecology and Social Structure, Volume 2: New World Monkeys, ‑a eldono, Needham Heights, MA: Pearson Custom Publishing & Prentice Hall, p. 77–80, 132–133, 141–143. ISBN 0-536-74364-9.
- ↑ Glazier, S. D.. (2003) Selected Readings in the Anthropology of Religion: Theoretical and Methodological Essays. Greenwood Publishing Group, p. 53. ISBN 9780313300905.
- ↑ (2005) “Cone topography and spectral sensitivity in two potentially trichromatic marsupials, the quokka (Setonix brachyurus) and quenda (Isoodon obesulus)”, Proceedings of the Royal Society B 272 (1565), p. 791–6. doi:10.1098/rspb.2004.3009.
- ↑ (1991) “Photosensitive and photostable pigments in the retinae of Old World monkeys”, The Journal of Experimental Biology 156 (1), p. 1–19. doi:10.1242/jeb.156.1.1. Alirita 2008-06-16..
- 1 2 (2003) “Evolution and selection of trichromatic vision in primates”, Trends in Ecology and Evolution 18 (4), p. 198–205. doi:10.1016/S0169-5347(03)00012-0.
- ↑ (2003) “Evolution and function of routine trichromatic vision in primates”, Evolution 57 (11), p. 2636–43. doi:10.1554/03-168.
Literaturo
- David J. Chivers, Bernard A. Wood & Alan Bilsborough, ed. (1984). Food Acquisition and Processing in Primates. New York & London: Plenum Press. ISBN 0-306-41701-4.
- Louis de Bonis: Vom Affen zum Menschen 1 & 2. Spektrum Compact 2004,1. Verlag Spektrum der Wissenschaft, Heidelberg 2004, ISBN 3-936278-70-9.
- Thomas Geissmann: Vergleichende Primatologie. Springer, Berlin 2003, ISBN 3-540-43645-6.
- Colin Groves: Primate Taxonomy. Smithsonian Institution Press, Washington 2001, ISBN 1-56098-872-X.
- Andreas Paul: Von Affen und Menschen. Wissenschaftliche Buchgesellschaft, Darmstadt 1998, ISBN 3-534-13869-4.
- Daris Swindler: Introduction to the Primates. University of Washington Press, Washington 1998, ISBN 0-295-97704-3.
- Thomas S. Kemp: The Origin and Evolution of Mammals. Oxford University Press, Oxford 2005, ISBN 0-19-850761-5.
- Sharon Lisa Gursky, K. A. Kimberly Anne-Isola Nekaris (2007), Primate Anti-Predator Strategies ; Springer, 31a de majo 2007 - 420 paĝoj.
Eksteraj ligiloj
- Primate Info Net
- Primates ĉe Animal Diversity Web
- Primate Research Institute, Universitato de Kioto
- High-Resolution Cytoarchitectural Primate Brain Atlases
- EUPRIM-Net: European Primate Network
- PrimateImages: Natural History Collection
- Tree of Life web project Arkivigite je 2011-04-26 per la retarkivo Wayback Machine