
Reproduktado estas la biologia procezo dum kiu produktiĝas novaj individuaj organismoj, plej ofte el la "gepatroj". Reproduktado estas fundamenta trajto de ĉiuj konataj formoj de la vivo; ĉiu individua organismo ekzistas kiel rezulto de reproduktado. La konataj manieroj de reproduktado dividiĝas vaste inter du ĉefaj tipoj: seksa kaj neseksa.
En neseksa reproduktado, organismo povas reproduktiĝi sen implico de alia organismo. Neseksa reproduktado ne estas limigita al unuĉelaj organismoj. La klonado de organismo estas formo de neseksa reproduktado. Per neseksa reproduktado organismo kreas genetike simila aŭ eĉ identa kopio de si mem. La evoluo de seksa reproduktado estas grava kaprompilo por biologoj. La duflanka kosto de sekso estas ke nur 50% el organismoj reproduktiĝas[1] kaj el organismoj nur 50% pasas siajn genojn.[2]
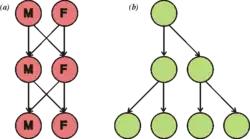
Seksa reproduktado tipe postulas la seksan interagadon de du specializitaj organismoj, nomitaj gametoj, kiuj enhavas duonon el la nombro de kromosomoj el tiuj de normalaj ĉeloj kaj estas kreataj per mejozo, per kiu tipe masklo fekundas inon de la sama specio por krei fekunditan zigoton. Tio produktas idajn organismojn kies genetikaj trajtoj estas derivaj el tiuj de la du gepatraj organismoj.
La tuto de organoj kaj histoj servataj por reproduktado en la homo estas la homa reprodukta sistemo.
Neseksa reproduktado

Neseksa reproduktado estas metodo de reproduktado kiu produktas idojn kun tute sama DNA. Ĝi malsimilas seksan reproduktadon kiu bezonas du gepatrojn, kaj produktas idojn kun DNA kiu malsamas tiun de la patro aŭ la patrino.
Neseksa reproduktado estas reĝimo de reproduktado laŭ kiu idoj ekestiĝas de unuopa organismo, kaj heredas nur la genojn de tiu patro; ĝi estas reproduktado kiu preskaŭ neniam implikas ploidon aŭ reproduktadon sen fuzio de gametoj. La idoj estos precizaj genetikaj kopioj de la patro, krom en la specifa kazo de aŭtomikso. Pli rigora difino estas agamogenezo kiu estas reproduktado sen la fuzio de gametoj. Neseksa reproduktado estas la primara formo de reproduktado por unu-ĉelaj organismoj kiel ekzemple la arĥeoj, eŭbakterioj, kaj protistoj. Ankaŭ multaj plantoj kaj fungoj reproduktiĝas sensekse.
Dum ĉiuj prokariotoj reproduktiĝas sensekse (sen la formado kaj fuzio de gametoj), mekanismoj por laterala gentransigo kiel ekzemple kunjugiĝo, transformo kaj transdukcion foje estas komparitaj kun seksa reproduktado (aŭ almenaŭ kun sekso, en la signifo de genetika rekombinigo).[3] Kompleta manko de seksa reproduktado estas relative rara inter multĉelaj organismoj, precipe bestoj. Estas ne totale komprenita kial la kapablo reproduktiĝi sekse estas tiel ofta inter ili. Nunaj hipotezoj[4] indikas ke neseksa reproduktado povas havi mallongperspektivajn avantaĝojn kiam rapida loĝantarkresko estas grava aŭ en stabilaj medioj, dum seksa reproduktado ofertas retan avantaĝon permesante pli rapidan generaciadon de genetika diverseco, permesante adaptadon al ŝanĝado de medioj. Evoluaj limigoj[5] povas substreki kial malmultaj bestoj cedis seksan reproduktadon tute en siaj vivocikloj. Alia limigo sur enrangigo de seksa ĝis neseksa reproduktado estus la akompananta perdo de mejozo kaj la protekta rekombineca riparo de DNA-difekto havigita kiel unu funkcio de mejozo.[6][7]
Seksa reproduktado

Seksa reproduktado estas biologia procedo, dum kiu estiĝas nova organismo per unuiĝo de seksa ĉelo kutime de du organismoj (escepte de unu). La seksa reproduktado havas do minimume du fazojn: produkton de seksaj ĉeloj kaj ties unuiĝon (la t.n. fekundigon). Alie inter la produkton de la seksaj ĉeloj kaj ties unuiĝon povas esti enmetita ankoraŭ la t.n. gametofito. Post la fekundigo povas sekvi ankoraŭ ia formo de zorgo pri idaro (mamula gravedeco, gardado de idoj, nutrado de idaro, nutrigado de fekundiga ovo per patrina kreskaĵo...).
Avantaĝo de la seksa reproduktado estas diverseco de la estiĝinta idaro, kiu estas grava el du motivoj. Ĝi parte ebligas al unuopuloj de la sama specio ekdiferencigi siajn vivpretendojn en diversa medio, do ili teorie ne bezonas tiom konkurenci unu al la alia kaj ili povas okupi pli ampleksan (ekologian) niĉon, parte (kaj tio estas eble ankoraŭ pli grava) ĝi kaŭzas, ke unuopaj unuopuloj de diversa populacio reagas al la samaj faktoroj de la medio diverse. Estas ekzemple nur minimuma ŝanco, ke troviĝus patogeno, rilate al kiu estos specife sentema la tuta populacio (diference de populacio de klonoj, kie tio povas okazi tute facile). Malavantaĝo de la seksa reprodukatdo estas ties relative malalta efektiveco kaj rapideco (kompare kun neseksa reproduktado).
La homa reproduktado estas ajna maniero de seksa reproduktado kiu havas kiel konsekvenco la homan fekundadon. Ĝi tipe koncernas la vaginan koiton inter viro kaj virino. Dum la koito, la interagado inter la viraj seksorganoj kaj la virinaj seksorganoj povas rezulti en la fekundigo de la ovolo de la virino fare de la spermatozoo de la viro. Tiu reproduktaj ĉeloj specializitaj nomitaj gametoj estas kreitaj per procezo nomita mejozo. Dum la normalaj ĉeloj enhavas 46 kromosomojn, nome 23 parojn, la gametoj enhavas nur 23 kromosomojn, kaj kiam tiuj du ĉeloj fuziiĝas en unu zigoto okazas la genetika rekombinado kaj la nova zigoto enhavas 23 kromosomojn de ĉiu gepatro, parigitaj en 23 paroj.[8] Post tipa periodo de gravedeco naŭmonata, okazas la nasko. La fekundigon de la ovulo oni povas atingi ankaŭ pere de metodoj de artefarita fekundigo kiu ne necesigas la vaginan koiton. Ekzistas ankaŭ la teknologio de helporeprodukta teknologio.
Fremdpolenado aŭ alogamio

La alogamio estas tipo de seksa reproduktado en plantoj kiuj konsistas en la kruc-polenado kaj fekundigo inter individuoj genetike diferencaj.[9] Tiu tipo de reproduktado favoras la produktadon de individuoj genetike novaj kaj, tial la konstantan generadon de genetika variableco en la populacioj. Ekzistas mekanismoj kiuj helpas la krucpolendon en plantoj tiaj kiaj la memneakordigeblo,[10] la heterostilio, la hercogamio kaj la dioikeco.[11]
En la evolucio de plantoj aperis mekanismoj de reproduktado kiuj favoris la alogamion kaj kiuj ekskludis ĉu totale ĉu parte la aŭtogamion. La dioikeco estas la plej grava el ili ĉar la inaj kaj malinaj gametoj originiĝas apartigite en diferencaj individuoj[12] (la asparago estas ekzemplo de dioika planto).[13] La inaj kaj malinaj gametoj disvolviĝas en diferencaj lokoj de unu planto (por ekzemplo, ĉe maizo) aŭ disvolviĝas nesinkronie, tio estas maturiĝas antaŭ ĉu la gineceo ĉu la androceo de la floro (Ekzemplo: kelkaj filikoj kaj kelkaj arboj kun anemofila polenado).
Granda nombro de angiospermoj posedis hermafroditajn florojn kun kaj stamenoj kaj karpeloj en la sama floro kaj ili estis plejparte memneakordigeblaj. Ankoraŭ nuntempe tiu situacio pluas. La plej modernaj angiospermoj estas karakterizataj ĉu kiel monoikaj ĉu kiel dioikaj. Krome oni priskribis kelkajn speciojn kiel tiuj de la genro Bryonia, por ekzemplo kiu prezentas individuojn kaj monoikajn kaj dioikajn. En la plej granda parto de la familioj de la angiospermoj la flora strukturo estas tia ke ĝi evitas la hazardan transdonon de la poleno al la stigmo. Alivorte, la flora strukturo estas tiom prilaborita ke nur la helpo de polenigistoj (insektoj, birdoj ktp) garantias la transdonon de la poleno al la stigmo de alia floro.
La androceo kaj la gineceo de la floroj de kelkaj specioj maturiĝas unu post alia, fenomeno nomita "dikogami" —en la homogamio, male ambaŭ floraj verticiloj maturiĝas samtempe—. Ĉe la protandriaj floroj, la androceo maturiĝas antaŭ la gineceo faras tion, dum ĉe la protoginaj floroj, la gineceo estas tiu kiu plej frue maturiĝas.
Memproduktado aŭ aŭtogamio

En botaniko, estas nomita "aŭtogamio" la maniero de seksa reproduktado konsistanta en la fuzio de inaj kaj malinaj gametoj produktitaj de la sama individuo. La polenado per poleno produktita de la sama floro aŭ de la sama planto estas nepra kondiĉo sed ne sufiĉa por ke okazu aŭtogamio, ĉar la poleno devas esti kapabla ĝermi ene de la stigmo, kreskiĝi en la stiluso kaj fari la duoblan fekundigon. La aŭtogamio estas tre disvastigita inter trudherboj, pioniraj plantoj kaj insulaj specioj, kiuj bezonas, ke la izolaj individuoj fruktiĝu kaj produktu semojn.[14] Ĉe la aŭtogamiaj specioj, la floroj ofte estas malgrandaj, kun nerimarkindaj floreroj, produktas malabundan kvanton de poleno kaj havas nek odoron nek nektaron.[15]
La aŭtogamio povas esti nepra aŭ elekta. La semoj produktitaj de nepra aŭtogamia planto devenas nur de mempolenado kaj memfekundigo. La klejstogameco, procezo laŭ kiu la polenado kaj fekundigo okazas antaŭ ol la floro malfermiĝas, estas mekanismo kiu kondukas al la nepra aŭtogamio. La semoj de elekta aŭtogamia planto, male, povas ĝermi ankaŭ pere de alogamio.
La floroj de la klejstogamaj specioj ĝenerale estas malgrandaj, nerimarkindaj, kaj havas mallongajn pedicelojn kaj tre ofte estas asociaj al subtera florado (Arachis, Paspalum amphicarpum). Krome, ofte ili havas ankaŭ pli malgrandajn anterojn aŭ pli malgrandan nombron de anteroj ol la rilataj ĉasmogamiaj specioj. En la genro Briza, por ekzemplo, la especies alogamiaj kaj memneakordigeblaj havas tri anterojn dum la du klejstogamaj specioj havas nur unu anteron kaj tiu havas nur unu dekonon de la grando.[16][17]
Kiam la polenado okazas inter diversaj floroj de la sama individuo, temas pri gejtonogamio, situacio ofta ĉe specioj kun amasa florado, kiel la Tabebuia heptaphylla aŭ la Ceiba speciosa. El vidpunkto de la genetika variableco, la gejtonogamio estas ekvivalenta al la aŭtogamio.
Mitozo kaj mejozo
Mitozo kaj mejozo estas metodoj de ĉeldivido.
Mitozo

Mitozo[18] estas ĉefa tipo de ĉeldividaj procesoj, dum kiu el unu patrina ĉelo estiĝas du, genetike samaj filinaj ĉeloj (restas la kromosoma nombro). En pli mallarĝa senco, mitozo estas duobliĝo kaj disiĝo de la kromosomoj kaj la korpetoj (organeloj), portantaj la genetikan informon.
Antaŭ la komenco de la mitozo, la kromosomoj duobliĝas kaj okazas sintezo de proteionoj de la pli posta kernobobeno. En la unua fazo de la mitozo (antaŭfazo), la kromosomoj dikiĝas kaj volviĝas. La nukleolo en la ĉelkerno ŝrumpiĝas kaj malaperas. Fine de la antaŭfazo, komenciĝas malkonstruo de kenomembrano, kaj organiziĝo de la kernobobeno el la kernofadenoj.

En la mezfazo, la kromosomoj konsistas el du-du kromatidoj kaj ordiĝas en la centro de la ĉelo (ekvatora ebeno). En la posta (malfrua) metafazo, ĉiu kromatida paro disiĝas al du samaj kromosomoj kaj fadenoj de la kernobobeno tiras ilin en la malajn polusojn.
En la finfazo, la strukturo de la kromosomo loziĝas, malkonstruiĝas la kerna bobeno, rekonstruiĝas la kernomembranoj kaj la nukleoloj, dividiĝas la citoplasmo de la patrina ĉelo kaj estiĝas la du posteulaj ĉeloj. Tiuj estiĝintaj ĉeloj havas la saman nombron de kromosomoj kaj saman strukturon kiel la patrina ĉelo. La staton post la finfazo, oni nomas interfazo.
La mitozo estas baza proceso de la vivo, ĉar ĝi ebligas estiĝon de novaj ĉeloj por la kresko kaj regeneriĝo. Tempodaŭro de la mitozo dependas de la ĉeltipo kaj specio, de minutoj ĝis horoj. Efikas je tio la tagperiodo, temperaturo, certaj kemiaĵoj.
Mejozo

Mejozo aŭ reduktoĉeldividiĝo estas binara dividiĝoprocezo kiu produktas generajn ĉelojn: nome ovolojn kaj spermatozoojn. La homologaj kromosomparoj pro tio ne plu kune troviĝas en la patrina ĉelo. Kio estas genetika de la patro kaj de la parino estos hazarde dividita al la filinaj ĉeloj. La patrina ĉelo estas duploida kaj enhavas homologajn kromosomojn. La filinaj ĉeloj enhavas nur unu kromosomon de ĉiu homologa paro kaj nomiĝas unuploida.
Interfazo
La evoluo estas preskaŭ sama al tiu de la mitozo. La unuan kreskofazon, la sintezan periodon kaj la duan kreskofazon ni povas resumi jene.
- Okazas DNA-replikado: ĉiu kromatido-drato sin mem kopias al du kromatidodratoj.
- Produktado de grandaj kvantoj da H1-histonoj (proteinoj).
- La kromatidodratoj sin pli spiraligas kaj kondensas al kromosomoj.
- Duobliĝo de la centrosomo (centriolparo) kaj membranpliiĝo.
- La homologaj kromosomoj serĉas unu la alian kaj povas juntiĝi: genetika interŝanĝo.

Strategioj de reproduktado
Estas ampleksa gamo de reproduktaj strategioj uzitaj de tre diversaj specioj. Kelkaj animaloj, kiel homo inter mamuloj aŭ norda sulo inter birdoj, ne atingas seksan maturiĝon dum multaj jaroj post nasko kaj eĉ tiam ili produktas malmultajn idojn. Aliaj reproduktiĝas rapide; sed, laŭ normalaj cirkonstancoj, plej parto de idoj ne survivas ĝis plenkreskeco. Por ekzemplo, kuniklo (matura post 8 monatoj) povas produkti 10–30 idojn jare, kaj drosofilo (matura post 10–14 tagoj) povas produkti ĝis 900 idojn jare. Tiuj du ĉefaj strategioj estas konataj kiel K-strategio (malmultaj idoj) kaj R-strategio (multaj idoj). Kiu strategio estis preferita en la evolucio dependas el granda vario de cirkonstancoj. Animaloj kun malmultaj idoj povas dediĉi pli da rimedoj al nutrado kaj protektado de ĉiu individua ido, tiel malpliigante la neceson havi multajn idojn. Aliflanke, animaloj kun multaj idoj povas dediĉi malpli da rimedoj al nutrado kaj protektado de ĉiu individua ido; ĉe tiuj tipoj de animaloj oftas ke multaj idoj mortas tuj post la nasko, sed sufiĉe da individuoj tipe survivas por elteni la populacion. Kelkaj organismoj kiel mielabeloj kaj drosofiloj retenas spermo en procezo nomita "insperma stokado", kaj tio pliigas la daŭradon de iliaj fekundebloj.

Helporeprodukta teknologio aŭ helpita reproduktado estas la teknologio uzata por atingi gravedecon en proceduroj kiaj fekundiga medicino, in vitro fekundigo kaj lu-patrineco. Ĝi estas reproduktiga teknologio uzata ĉefe por kontraŭnefekundaj traktaĵoj, kaj estas konata ankaŭ kiel fekundiga traktado. Ĝi ĉefe apartenas al la kampo de reprodukta endokrinologio kaj nefekundeco, kaj povas inkludi ankaŭ intracitoplasman sperminjekton kaj kriokonservadon. Kelkaj formoj de helporeprodukta teknologio estas uzataj ankaŭ rilate al fekundaj paroj por genetikaj tialoj (antaŭplanta genetika diagnozo). Helporeprodukta teknologio estas uzata ankaŭ por paroj kiuj estas malkongruaj pro kelkaj komunikeblaj malsanoj; por ekzemplo, HIV por limigi la riskon de infekto kiam oni deziras gravedecon.

Seksa kaj neseksa reproduktadoj
Organismoj kiuj reproduktiĝas pere de neseksa reproduktado tendencas kreskiĝi en eksponencialaj nombroj. Tamen, ĉar ili dependas el mutacio por variaĵoj en sia DNA, ĉiuj membroj de la specio havas similajn vundeblojn. Organismoj kiuj reproduktiĝas pere de seksa reproduktado atingas pli malgrandan nombron de idoj, sed la granda kvanto de variado en iliaj genoj faras ili malpli suferantaj pro malsanoj.
Multaj organismoj povas reproduktiĝis kaj sekse kaj nesekse. Afidoj, muko-ŝimoj, maranemonoj, kelkaj specioj de marsteloj (pre de fragmentado), kaj multaj plantoj estas tiaj ekzemploj. Kiam mediaj faktoroj estas favoraj, neseksa reproduktado estas uzata por profiti tiujn taŭgajn kondiĉojn por survivado kiel ekzemple abunda manĝodisponeblo, taŭga ŝirmado, favoraj klimatoj, malsanoj, optimuma pH aŭ taŭga miksaĵo de aliaj vivmanieraj postuloj. Populacioj de tiuj organismoj pliiĝas eksponenciale pere de neseksaj reproduktaj strategioj por atingi plenan avantaĝon de riĉaj manĝorimedoj.[19]
Kiam manĝofontoj elĉerpiĝas, la klimato malboniĝas, aŭ la individua survivafo estas atakita pere de aliaj kontraŭaj ŝanĝoj en la vivkondiĉoj, tiuj organismoj ŝanĝiĝas al seksaj manieroj reproduktiĝi. Seksa reproduktado sekurigas miksaĵon de la genaro de la specio. La variaĵoj troveblaj en la idaro de seksa reproduktado ebligas, ke kelkaj individuoj estas pli taŭgaj por survivado kaj havigas mekanismon por ke okazu selekta adaptado. La mejoza etapo de la seksa ciklo ankaŭ ebligas speciale efektivan riparadon de DNA damaĝoj (vidu artikolon pri mejozo).[19] Aldone, seksa reproduktado kutime rezultas en la formado de vivetapo kiu kapablas elteni la kondiĉojn kiuj minacus la idarojn de neseksa patro. Tiel, semoj, sporoj, ovoj, pupoj, krizalidoj aŭ aliaj "super-vintraj" stadioj de seksa reproduktado sekurigas la survivadon dum malfavoraj tempoj kaj la organismo povas "atendi" super kontraŭaj situacioj ĝis okazu reŝanĝo al taŭgaj vivkondiĉoj.
Senreprodukta vivo
La ekzistado de vivo sen reproduktado estas celo de ioma spekulativado. La biologia studado kiel la origino de la vivo produktis reproduktantajn organismojn el ne-reproduktantaj elementoj estas nomita abiogenezo. Ĉu okazis aŭ ne okazis kelkaj sendependaj abiogenezaj okazaĵoj, biologoj kredas, ke la "lasta universala praulo" de ĉiu nuntempa vivantaĵo sur la Tero vivis antaŭ ĉirkaŭ 3.5 mil milionoj da jaroj.

Sciencistoj spekulativadis pri la eblo kreadi vivon ne-reproduktade en laboratorio. Kelkaj sciencistoj ja jam sukcesis produkti simplajn virusojn el tute ne-vivantaj materialoj.[20] Tamen, virusoj estas ofte konsiderataj kiel ne vivantuloj. Estante nenio pli ol iom da RNA aŭ de DNA en proteina kapsulo, ili ne havas metabolon kaj povas nur mem-replikadiĝi per la helpo de elprenita metabola mekanismo de ĉelo.
La produktado de vera vivanta organismo (ekz. simpla bakterio) sen prauloj estus multe pli kompleksa tasko, sed povus esti ebla je ioma grado laŭ la nuna biologia sciaro. Sinteza genaro estis jam transigita al ekzistanta bakterio kie ĝi anstataŭis la indiĝenan DNA, rezulte en la artefarita produktado de nova organismo de Mycoplasma mycoides.[21]
Estas ioma debato ene de la sciencz komunumo ĉu tiu ĉelo povas esti konsiderata komplete sinteza[22] sur la bazo ke la kemie sinteza genaro estis preskaŭ 1:1 kopio de nature ekzistanta genaro kaj ke la ricevinta ĉelo estis nature ekzistanta bakterio. La Craig Venter Institute eltenas la esprimon "sinteza bakteria ĉelo" sed ili ankaŭ klarigas "...ni ne konsideras, ke tio estas "krei vivon el elŝiraĵo" sed anstataŭe ni estas kreante novan vivon el jam ekzistinta vivo uzante sintezan DNA".[23] Venter planas patenti siajn eksperimentajn ĉelojn, asertante, ke "ili estas tute klare homaj inventaĵoj".[22] Iliaj kreintoj sugestas, ke konstrui 'sintezan vivon' ebligus, ke esploristoj lernu pri la vivo konstruante ĝin, anstataŭ havi ĝin aparte. Ili proponas ankaŭ ŝanĝi la limojn inter vivo kaj maŝinoj ĝis ambaŭ koincidu por produkti "verajn programeblajn organismojn".[24] Koncernaj esploristoj asertis, ke la kreado de "vera sinteza biokemia vivo" estas relative proksime atingebla pere de la nuntempa teknologio kaj malmultekosta kompare kun la klopodoj necesaj por porti homon sur la Lunon.[25]
Vidu ankaŭ
Referencoj
- ↑ Ridley M (2004) Evolution, 3rd edition. Blackwell Publishing, p. 314.
- ↑ John Maynard Smith The Evolution of Sex 1978.
- ↑ Narra HP, Ochman H (2006). “Of what use is sex to bacteria?”, Current Biology 16 (17), p. R705–710. doi:10.1016/j.cub.2006.08.024.
- ↑ Dawson KJ (October 1995). “The Advantage of Asexual Reproduction: When is it Two-fold?”, Journal of Theoretical Biology 176 (3), p. 341–347. doi:10.1006/jtbi.1995.0203.
- ↑ Engelstädter J (November 2008). “Constraints on the evolution of asexual reproduction”, BioEssays 30 (11–12), p. 1138–1150. doi:10.1002/bies.20833.
- ↑ Bernstein H, Hopf FA, Michod RE (1987). “The molecular basis of the evolution of sex”, Adv. Genet. 24, p. 323–70. doi:10.1016/s0065-2660(08)60012-7.
- ↑ Avise, J. (2008) Clonality: The Genetics, Ecology and Evolution of Sexual Abstinence in Vertebrate Animals. See pp. 22-25. Oxford University Press. ISBN 019536967X ISBN 978-0195369670
- ↑ «gamete | Definition, Formation, Examples, & Facts». Encyclopedia Britannica (en angla). Konsultita la 11an de julio 2021.
- ↑ Curso Regional Sobre de la Caficultura Moderna. Bib. Orton IICA / CATIE. Konsultita la 2an de Januaro 2018.
- ↑ El cultivo del olivo. Mundi-Prensa Libros. 2007-12. ISBN 9788484764830. Konsultita la 2an de Januaro 2018.
- ↑ Peña, José Ramón Alonso (2011). MANUAL DE HISTOLOGÍA VEGETAL. Editorial Paraninfo. ISBN 9788484764298. Konsultita la 2an de Januaro 2018.
- ↑ Linné, Carl von (1787). Parte práctica de botánica del caballero Cárlos Linneo, que comprehende las clases, órdenes, géneros, especies y variedades de las plantas, con sus caracteres genéricos y específicos, sinónimos mas selectos, nombres triviales, lugares donde nacen y propiedades. en la Imprenta Real. Konsultita la 2an de Januaro 2018.
- ↑ JOSÉ IGNACIO, CUBERO SALMERÓN (2013). Introducción a la mejora genética vegetal. Mundi-Prensa Libros. ISBN 9788484766551. Konsultita la 2an de Januaro 2018.
- ↑ González, A.M. «Polinización». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Arkivita el la originalo la 9an de Aŭgusto 2014. Konsultita la 10an de Januaro 2013.
- ↑ Frankel, R. & Galun, E. «Pollination mechanisms, reproduction, and plant breeding.» Monographs on Theoretical and Applied Genetics, no. 2. Springer-Verlag. Berlin. 1977. 281 p.
- ↑ Connor, H.E. 1979. "Breeding systems in the grasses: a survey". New Zealand Journal of Botany 17:547-74.
- ↑ Campbell, C., J. Quinn, G. Cheplick, & T J Bell. 1983. «Cleistogamy in Grasses.» Annual Review of Ecology and Systematics Vol. 14: 411-441.
- ↑ mitozo en vortaro.net
- 1 2 (Junio 2020) “Reproductive polyphenism and its advantages in aphids: Switching between sexual and asexual reproduction”, Journal of Integrative Agriculture 19 (6), p. 1447–1457. doi:10.1016/S2095-3119(19)62767-X.
- ↑ Chemical synthesis of poliovirus cDNA: generation of infectious virus in the absence of natural template Arkivigite je 2022-08-31 per la retarkivo Wayback Machine
Scientists Create Artificial Virus Arkivigite je 2005-10-31 per la retarkivo Wayback Machine - ↑ (2010) “Creation of a Bacterial Cell Controlled by a Chemically Synthesized Genome”, Science 329 (5987), p. 52–56. doi:10.1126/science.1190719.
- 1 2 Robert Lee Hotz. "Scientists Create First Synthetic Cell", May 21, 2010.
- ↑ Craig Venter Institute FAQ. Arkivita el la originalo je 2010-12-28. Alirita 2011-04-24. Arkivigite je 2010-12-28 per la retarkivo Wayback Machine Arkivita kopio. Arkivita el la originalo je 2010-12-28. Alirita 2022-12-09.
- ↑ W. Wayte Gibbs (Majo 2004). “Synthetic Life”, Scientific American. Alirita 2012-12-22.. Arkivigite je 2012-10-13 per la retarkivo Wayback Machine Arkivita kopio. Arkivita el la originalo je 2012-10-13. Alirita 2022-12-09.
- ↑ NOVA: Artificial life. Arkivita el la originalo je 2007-01-21. Alirita 2007-01-19.
Literaturo
- Tobler, M. & Schlupp,I. (2005) Parasites in sexual and asexual mollies (Poecilia, Poeciliidae, Teleostei): a case for the Red Queen? Biol. Lett. 1 (2): 166-168.
- Zimmer, Carl. Parasite Rex: Inside the Bizarre World of Nature's Most Dangerous Creatures, New York: Touchstone, 2001.
- GardenWeb Glossary of Botanical Terms, eldono 2.1, 2002, Allogamy, cross-fertilization, cross-pollination, hybridization
- Stedman's Online Medical Dictionary, eldono 27a, 2004, Allogamy
Plia legado
- Frankel, R.; kaj E. Galun (1977). «Pollination mechanisms, reproduction, and plant breeding». Monographs on Theoretical and Applied Genetics (2). Berlín: Springer-Verlag, 281 p.
- Judson, Olivia (2003) Dr.Tatiana's Sex Advice to All Creation: Definitive Guide to the Evolutionary Biology of Sex. ISBN 978-0-09-928375-1
- The Evolution of Sex: An Examination of Current Ideas Richard E. Michod kaj Bruce E. Levin, eldonistoj (1987) Sinauer Associates Inc., Publishers, Sunderland, Massachusetts ISBN 0878934596 ISBN 978-0878934591
- Michod, R.E. Eros and Evolution: A natural philosophy of sex (1994). Addison-Wesley Publishing Company, Reading, Massachusetts ISBN 0201442329 ISBN 978-0201442328
Eksteraj ligiloj
- Asexual Reproduction Arkivigite je 2018-01-22 per la retarkivo Wayback Machine
- Journal of Biology of Reproduction
- Journal of Andrology Arkivigite je 2005-11-07 per la retarkivo Wayback Machine
- Roa Meggo, Ysis J. & Vara Horna, Arístides A. (2005). Aspectos Psicológicos y legales de la mujer frente a las nuevas tecnologías reproductivas en el Perú, Asociación por la Defensa de las Minorías: Lima. En