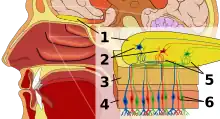
Flar(ad)o, flarsenso aŭ olfakto, estas la senso pri odoro. Ĉi tiun senson efektivigas specialaj sensaj ĉeloj de la naza kavo de vertebruloj, kiuj povas esti konsiderataj analogaj al sensaj ĉeloj de la palpiloj de senvertebruloj. En homoj, olfakto okazas kiam odoraj molekuloj ligiĝas al specifaj kemiajoj sur la olfaktaj ricevantoj.[1] Ĉi tiuj ricevantoj estas uzataj sensi la ĉeeston de odoro. Ili kuniĝas ĉe la glomerulo, strukturo kiu sendas signalojn al la olfakta ampolo (cerba strukturo rekte super la naza kavo kaj sub la fronta lobo).[2] Multaj vertebruloj, inkluzive plejmultajn mamulojn kaj rampulojn, havas du apartajn olfaktajn sistemojn—la ĉefa olfakta sistemo kaj la akcesora olfakta sistemo (uzata plejparte sensi feromonojn). Por aerospirantaj bestoj, la ĉefa olfakta sistemo sensas volatilajn kemiaĵojn kaj la akcesora flara sistemo sensas fluidajn kemiaĵojn.[3] Olfakto, kune kun gusto, estas formo de kemisensilo. La kemiaĵoj kiuj aktivigas la olfaktan sistemon, ĝenerale je tre malaltaj koncentriĝoj, nomiĝas odorantoj. Kvankam gusto kaj odoro estas apartaj sensaj sistemoj en landaj bestoj, akvoloĝantaj organismoj ofte havas unu kemian senson.[4]
Volatilaj etmolekulaj odorantoj, nevolatilaj proteinoj kaj nevolatilaj hidrokarbonoj povas produkti flarajn sensojn. Iuj bestaj specioj kapablas flari karbonan dioksidon en etaj koncentriĝoj.[5]
Studo de olfakto
_L'Odorat_(La_Dame_%C3%A0_la_licorne)_-_Mus%C3%A9e_de_Cluny_Paris.jpg.webp)
Frua scienca studado de olfakto inkluzivas la vastan doktorecan disertacion de Eleanor Gamble, eldonita en 1898, kiu komparis la flaran al aliaj sensoj kaj implicis ke odoro havas malpli intensan distingopovon.[6] Kiel la epikura kaj atomisma roma filozofo Lucretius (1a Jarcento KE) konjektis, malsamaj odoroj estas atribuataj al malsamaj formoj kaj grandecoj de "atomoj" (odoraj molekuloj en la moderna kompreno) kiuj stimulas la flaran organon. Moderna manifestacio de tiu teorio estis la klonado de flaraj ricevantaj proteinoj fare de Linda B. Buck kaj Richard Axel (kiuj ricevis Nobelpremion en 2004) kaj posta parigo de odoraj molekuloj al specifaj ricevantaj proteinoj. Ĉiu odora ricevanta molekulo rekonas nur specifan molekulan trajton aŭ klason de odoraj molekuloj. Mamuloj havas ĉirkaŭ mil genojn kiuj kodas por odorricevo.[7] El la genoj kiuj kodas por odoraj ricevantoj, nur frakcio povas funkcii. Homoj havas ege malpli da aktivaj odoraj ricevantaj genoj ol aliaj primatoj kaj aliaj mamuloj.[8] En mamuloj, ĉiu flara ricevanta neŭrono esprimas nur unu funkcikapablan odoran ricevanton.[9] Odoraj ricevantaj nervaj ĉeloj funkcias kiel ŝlosilo-serura sistemo: Se la aerportataj molekuloj de certa kemiaĵo povas bone sidi en la seruro, la nerva ĉelo respondas. Ekzistas, nuntempe, nombro da konkurencaj teorioj koncerne al la mekanismo de odorkodigo kaj percepto. Laŭ la forma teorio, ĉiu ricevanto sensas trajton de la odora molekulo. Malforta-forma teorio, nomita odotopa teorio, sugestas ke malsamaj ricevantoj trovas nur malgrandajn pecojn de molekuloj kaj ĉi tiuj minimumaj enigoj kombiniĝas formi pli grandan flaran percepton (simile al kiel la vida percepto konstruiĝas el malpli grandaj, informo-malriĉaj sensoj, kombinitaj kaj rafinitaj por krei detalan entutan percepton).[Citaĵo bezonata] Alternativa teorio, la ''vibra teorio'' proponita de Luca Turin, postulatas ke odoraj ricevantoj sensas la frekvencojn de vibroj de odoraj molekuloj en la transruĝa gamo de kvantuma tunelado.[10][11] Tamen, la kondutaj prognozoj de ĉi tiu teorio estas dubindaj.[12] Neniu teorio kontentige klarigas flaran percepton.
Ĉefa flara sistemo
Vertebruloj sensas odorojn per olfaktaj sensaj neŭronoj en la olfakta epitelio. La olfakta epitelio konsistas el almenaŭ ses morfologie kaj biokemie malsamaj ĉelaj tipoj.[13] La proporcio de olfakta epitelio kompare al spira epitelio (ne nervizita) donas indikon de flara sensiveco de la besto. Homoj havas ĉirkaŭ 10 cm2 de olfakta epitelio, dum iuj hundoj havas 170 cm2. Hunda olfakta epitelio estas ankaŭ konsiderinde pli dense nervizita, kun centoble pli da ricevantoj por kvadrata centimetro.[14]
Molekuloj de odorantoj pasante tra la supera naza konko de la nazaj trairejoj solviĝas en la interna tegaĵo de muko de la supera parto de la kavo kaj estas sensataj de flaraj ricevantoj sur la dendritoj de la flarsensaj neŭronoj. Tio ĉi povas okazi per difuzado aŭ per ligado de la odoranto al odorantligaj proteinoj. La muko kovranta la epitelion enhavas mukopolisakaridojn, salojn, enzimojn kaj antikorpojn (ĉi tiuj estas tre gravaj, ĉar la flaraj neŭronoj provizas rektan trairon por infekto pasi al la cerbo). Ĉi tiu muko agas kiel solvento por odoraj molekuloj, fluas konstante kaj estas anstataŭigita post proksimume ĉiu 10 minuta periodo.
En insektoj, odoroj estas sensataj de flarsensaj neŭronoj en la kemiosensaj sensiloj, kiuj ĉeestas en insektaj antenoj, palpiloj kaj tarsoj, sed ankaŭ sur aliaj partoj de la insekta korpo. Odorantoj penetras en la kutiklajn porojn de kemiosensaj sensiloj kaj kontaktas insektajn odorligajn proteinojn (OBPs) aŭ Kemiosensajn proteinojn (CSPs), antaŭ ol aktivigi la sensajn neŭronojn.
Ricevanta neŭrono
La ligado de la ligando (odoranta molekulo) al la ricevanto kondukas al aga potencialo en la ricevanta neŭrono, tra dua mesaĝista vojo, dependante de la organismo. En mamuloj, la odorantoj stimulas adenilatan ciklazon sintezi cAMP tra G-proteino nomita Golfo. cAMP, kiu estas la dua mesaĝisto ĉi tie, malfermas ciklan nukleotid-ŝaltitan jonkanalon (CNG), produktante enfluon de katjonoj (plejparte Ca2+ kun kelkaj Na+) en la ĉelon, iomete despolarizante ĝin. La Ca2+ siavice malfermas Ca2+-aktivigitan kloridan kanalon, kaŭzante elfluadon de Cl-, plue despolarizante la ĉelon kaj ekkaŭzante agan potencialon. Ca2+ estas tiam elpremata tra natrio-kalcio interŝanĝanto. Kalcio-kalmodulin-komplekso ankaŭ agas malhelpi la ligadon de cAMP al la cAMP-dependa kanalo, kaj tiel kontribuante al olfakta adapto. Ĉi tiu mekanismo de transmetado estas iom unika, en tio ke cAMP funkcias rekte ligante al la jona kanalo prefere ol per aktivigo de proteina kinazo-A. Ĝi estas simila al la transmetada mekanismo por lumsensiloj, en kiu la dua mesaĝisto cGMP funkcias rekte ligante al jonaj kanaloj, kiu sugestas ke eble unu el ĉi tiuj ricevantoj evolue adaptis al la alia. Estas ankaŭ konsiderindaj similecoj en la tuja procesado de stimuloj per latera inhibado. Oni povas mezuri la mezan aktivecon de la ricevantaj neŭronoj per pluraj metodoj. En vertebruloj, respondoj al odoro estas mezureblaj per elektro-olfaktogramo aŭ per kalcia bildigo de ricevantaj neŭronaj finaĵoj en la olfakta ampolo. En insektoj, oni povas fari elektroantenogramon aŭ ankaŭ kalcian bildigon ene de la olfakta ampolo.
Olfaktampolaj projekcioj
 Olfaktaj sensaj neŭronoj projekcias aksonojn al la cerbo ene de la olfakta nervo, (cerba nervo I). Ĉi tiuj nervaj fibroj, sen mjelinaj ingoj, pasas al la olfakta ampolo de la cerbo tra perforaĵo en la kribreca plato, kiu siavice projekcias olfaktan informon al la olfakta kortiko kaj aliaj areoj.[15] La aksonoj de la olfaktaj ricevantoj konverĝas en la ekstera tavolo de la olfakta ampolo ene de malgrandaj (~50 mikrometrojn laŭ diametro) strukturoj nomitaj glomeruloj. Mitralaj ĉeloj, kiuj troviĝas en la interna tavolo de la olfakta ampolo, formas sinapsojn kun la aksonoj de la sensaj neŭronoj en glomeruloj kaj sendas la informon pri la odoro al aliaj partoj de la olfakta sistemo, kie multaj signaloj povas esti procezataj formi sintezitan olfaktan percepton. Granda grado de konverĝo okazas, kun dudekkvinmil aksonoj sinapsantaj ĉe ĉirkaŭ dudek kvin mitralaj ĉeloj kaj kun ĉiu de ĉi tiuj mitralaj ĉeloj projekciantaj al multoblaj glomeruloj. Mitralaj ĉeloj ankaŭ projekcias al periglomerulaj ĉeloj kaj granulaj ĉeloj kiu inhibas la mitralajn ĉelojn kiuj ĉirkaŭas ĝin (flanka inhibo). Granulaj ĉeloj ankaŭ efektivigas inhibon kaj eksciton de mitralaj ĉeloj tra vojoj de centrifugaj fibroj kaj la antaŭaj olfaktaj nukleoj. Neŭromodulantoj, kiel Acetilkolino, Serotonino kaj Norepinefrino, ĉiuj sendas aksonojn al la olfakta ampolo kaj estis implikitaj en pliiga modulado, skema apartigo kaj memoraj funkcioj, respektive.[16][17][18]
Olfaktaj sensaj neŭronoj projekcias aksonojn al la cerbo ene de la olfakta nervo, (cerba nervo I). Ĉi tiuj nervaj fibroj, sen mjelinaj ingoj, pasas al la olfakta ampolo de la cerbo tra perforaĵo en la kribreca plato, kiu siavice projekcias olfaktan informon al la olfakta kortiko kaj aliaj areoj.[15] La aksonoj de la olfaktaj ricevantoj konverĝas en la ekstera tavolo de la olfakta ampolo ene de malgrandaj (~50 mikrometrojn laŭ diametro) strukturoj nomitaj glomeruloj. Mitralaj ĉeloj, kiuj troviĝas en la interna tavolo de la olfakta ampolo, formas sinapsojn kun la aksonoj de la sensaj neŭronoj en glomeruloj kaj sendas la informon pri la odoro al aliaj partoj de la olfakta sistemo, kie multaj signaloj povas esti procezataj formi sintezitan olfaktan percepton. Granda grado de konverĝo okazas, kun dudekkvinmil aksonoj sinapsantaj ĉe ĉirkaŭ dudek kvin mitralaj ĉeloj kaj kun ĉiu de ĉi tiuj mitralaj ĉeloj projekciantaj al multoblaj glomeruloj. Mitralaj ĉeloj ankaŭ projekcias al periglomerulaj ĉeloj kaj granulaj ĉeloj kiu inhibas la mitralajn ĉelojn kiuj ĉirkaŭas ĝin (flanka inhibo). Granulaj ĉeloj ankaŭ efektivigas inhibon kaj eksciton de mitralaj ĉeloj tra vojoj de centrifugaj fibroj kaj la antaŭaj olfaktaj nukleoj. Neŭromodulantoj, kiel Acetilkolino, Serotonino kaj Norepinefrino, ĉiuj sendas aksonojn al la olfakta ampolo kaj estis implikitaj en pliiga modulado, skema apartigo kaj memoraj funkcioj, respektive.[16][17][18]
La mitralaj ĉeloj lasas la olfaktan ampolon en la flanka olfakta traktuso, kiu sinapsas sur kvin ĉefaj regionoj de la cerebro: la antaŭa olfakta nukleo, la olfakta tubereto, la amigdalo, la piriforma kortiko kaj la entorina kortiko. La antaŭa olfakta nukleo projekcias, tra la antaŭa komisuro, al la kontralatera olfakta ampolo, inhibi ĝin. La piriforma kortiko havas du gravajn dividaĵojn kun anatome klaraj organizoj kaj funkcioj. La antaŭa piriforma kortiko (APC) estas pli bone asociita kun la mezurado de la kemia strukturo de la odorantaj molekuloj kaj dum la posta piriforma kortiko (PPC) estas plej bone sciita pro ĝia forta rolo klasifiki odorojn kaj taksi similecojn inter odoroj (ekz. menta, ligna, kaj citrusa estas odoroj kiuj estas distingeblaj per la PPC malgraŭ ke ili estas tre diversaj kemiaĵoj kaj en koncentriĝo-sendependa maniero).[19] La piriforma kortiko projekcias al la meza dorsa nukleo de la talamo, kiu tiam projekcias al la orbitofronta kortiko. La orbitofronta kortiko efektivigas konscian percepton de la odoro. La tritavola piriforma kortiko projekcias al nombro da talamaj kaj hipotalamaj nukleoj, la hipokampo kaj amigdalo kaj la orbitofronta kortiko sed ĝia funkcio estas plejparte nekonata. La entorina kortiko projekcias al la amigdalo kaj okupiĝas en emociaj kaj aŭtonomaj respondoj al odoro. Ĝi ankaŭ projekcias al la hipokampo kaj okupiĝas pri instigo kaj memoro. Odora informo konserviĝas en longatempa memoro kaj havas fortajn konektojn al emocia memoro. Tio ĉi estas eble pro proksimaj anatomiaj ligoj de la olfakta sistemo al la limba sistemo kaj hipokampo, areoj de la cerbo kiuj estas okupitaj en emocio kaj loka memoroj, respektive.
Pro tio ke iu ajn ricevanto estas respondemaj al diversaj odorantoj kaj ekzistas multa konverĝo ĉe la nivelo de la olfakta ampolo, ŝajnas strange ke homoj kapablas distingi tiom multe da malsamaj odoroj. Ŝajnas ke devas okazi tre kompleksa formo de procezado; tamen, kiel oni povas montri, dum multaj neŭronoj en la olfakta ampolo (kaj eĉ la piriforma kortiko kaj amigdalo) estas respondemaj al multaj malsamaj odoroj, duono de la neŭronoj en la orbitofronta kortiko estas respondema al nur unu odoro kaj la aliaj al nur kelkaj. Estis montrite per mikroelektrodaj studoj ke ĉiu individua odoro donas apartan specifan spacan mapon de ekscito en la olfakta ampolo. Eblas ke, per spaca kodado, la cerbo kapablas distingi specifajn odorojn. Tamen, oni devas ankaŭ konsideri tempan kodiĝon. Dum tempo, la spacaj mapoj ŝanĝiĝas, eĉ por unu aparta odoro kaj la cerbo devas esti kapabla procezi ĉi tiujn detalojn ankaŭ.
Enigoj de la du naztruoj havas apartajn enigojn al la cerbo kun la rezulto ke eblas por homoj sperti perceptan rivalecon en la olfakta senso parenca al tio de duokula rivaleco kiam estas du malsamaj enigoj en la du naztruoj.[20]
En insektoj, odoroj estas sensataj de sensiloj kiuj troviĝas sur la antenoj kaj makzela palpilo kaj unue traktataj de la antena lobo (analoge al la olfakta ampolo) kaj poste de la agarikaj korpoj kaj latera korno.
Akcesora flara sistemo
Multaj bestoj, inkluzive plejmultajn mamulojn kaj rampulojn, sed ne homoj, havas du apartajn olfaktajn sistemojn: ĉefan olfaktan sistemon, kiu sensas volatilajn stimulojn, kaj akcesoran olfaktan sistemon, kiu sensas fluidajn stimulojn. Konduta pruvo sugestas ke ĉi tiuj fluidfazaj stimuloj ofte funkcias kiel feromonoj, kvankam feromonoj ankaŭ povas esti sensataj de la ĉefa olfakta sistemo. En la akcesora olfakta sistemo, stimuloj estas sensataj de la vomeronaza organo, kiu troviĝas en la vomero, inter la nazo kaj la buŝo. Serpentoj uzas ĝin por flari predon, eligante sian langon kaj tuŝante ĝin al la organo. Kelkaj mamuloj faras flemenreagon (germane flehmen) por direkti stimulojn al ĉi tiu organo.
La sensaj ricevantoj de la akcesora olfakta sistemo troviĝas en la vomeronaza organo. Kiel en la ĉefa olfakta sistemo, la aksonoj de ĉi tiuj sensaj neŭronoj projekcias de la vomeronaza organo al la akcesora olfakta ampolo, kiu en la muso troviĝas sur la dorsa-posta parto de la ĉefa olfakta ampolo. Diference al en la ĉefa olfakta sistemo, la aksonoj kiuj lasas la akcesoran olfaktan ampolon ne projekcias al la kortiko de la cerbo sed prefere al celoj en la amigdalo kaj lita nukleo de la nerva fibro stria terminalis kaj de tie al la hipotalamo, kie ili povas influi agresan kaj pariĝa kondutojn.
Homa flara sistemo
En virinoj, la flarsenso estas plej forta ĉirkaŭ la tempo de ovolado, signife pli forta ol dum aliaj fazoj de la menstrua ciklo kaj pli forta ol la senso en viroj.[21]
La MHC-genoj (nomitaj HLA en homoj) estas grupo de genoj en multaj bestoj kaj grava por la imuna sistemo; ĝenerale, ido de gepatroj kun malsamaj MHC-genoj havas pli fortan imunan sistemon. Fiŝoj, musoj kaj virinoj kapablas flari iun econ de la MHC-genoj de eblaj sekspartneroj kaj preferas partnerojn kun MHC-genoj malsamaj al la propraj.[22][23]
Homoj povas sensi individuojn kiu estas sango-rilatitaj (patrinaj/patroj kaj infanoj sed ne edzoj kaj edzinoj) per olfakto.[24] Patrinoj povas identigi per korpa odoro siajn biologiajn infanojn sed ne iliajn vicinfanojn. Antaŭadoleskaj infanoj povas olfakte sensi siajn plenajn gefratojn sed ne duongefratojn aŭ vicgefratojn kaj tio ĉi povus klarigi evitadon de incesto kaj la Westermarck-efikon.[25] Funkcia bildigo montras ke ĉi tiu flara parenceca malkaŝa procezo okupas la fronta-tempiostan kuniĝon, la insulan kortikon kaj la dorsomedialan prefrontan kortikon sed ne la primaran aŭ duarangan olfaktajn kortikojn aŭ la rilatajn piriforman kortikon aŭ orbitofrontan kortikon.[26]
En insektoj, odoroj estas sensataj de flarsensaj neŭronoj en la kemiosensaj sensiloj, kiuj ĉeestas en insektaj antenoj, palpiloj kaj tarsoj, sed ankaŭ sur aliaj partoj de la insekta korpo. Odorantoj penetras en la kutiklajn porojn de kemiosensaj sensiloj kaj kontaktas insektajn odorligajn proteinojn (OBPs) aŭ Kemiosensajn proteinojn (CSPs), antaŭ ol aktivigi la sensajn neŭronojn.
Olfakta kodigo kaj percepto
Kiel olfakta informo kodiĝas en la cerbo por ebligi konvenan percepton estas ankoraŭ esplorata kaj la procezo ne estas tute komprenita. Kiam odoro estas sensata de ricevantoj, la ricevantoj iusence dispecigas la odoron kaj tiam la cerbo rekonstruas la odoron por identigi kaj percepti.[27] La odoro ligiĝas al ricevantoj kiuj rekonas nur specifan funkcian grupon aŭ trajton, de la odoro. Tial la kemia karaktero de la odoro estas grava.[28]
Liginte la odoron, la ricevanto aktiviĝas kaj sendas signalon al la glomeruloj.[28] Ĉiu glomerulo ricevas signalojn de multaj ricevantoj kiuj sensas similajn odorajn trajtojn. Ĉar multaj ricevantaj tipoj aktiviĝas pro la diversaj kemiaj trajtoj de la odoro, pluraj glomeruloj aktiviĝas. Ĉiuj signaloj el la glomeruloj tiam sendiĝas al la cerbo, kie la kombinaĵo de glomerulaj aktivigoj kodos la malsamajn kemiajn trajtojn de la odoro. La cerbo tiam esence kunmetas la pecojn de la aktiviga skemo por identigi kaj percepti la odoranton.[28] Ĉi tiu distribuita kodado permesas al la cerbo sensi specifajn odorojn en miksaĵo de multaj apartaj fonaj odoroj.[29]
Ĝenerala ideo estas, ke la aranĝo de cerbaj strukturoj respondas al fizikaj trajtoj de stimuloj (nome topografia kodigo) kaj similaj analogioj estas faritaj en olfakto pri konceptoj kiel la aranĝo rilata al kemiaj trajtoj (nomita kemiotopio) aŭ perceptaj trajtoj.[30] Dum kemiotopio estas tre polemikiga koncepto, ekzistas plua pruvo por percepta informo efektivigita en la spacaj dimensioj de olfaktaj retoj.[30][31]
Kvankam komuna saĝo kaj laika literaturo, bazitaj sur impresionismaj eltrovoj en la 1920aj jaroj, longe prezentis homan olfakton esti kapabla distingi inter proksimume 10,000 unikaj odoroj, lastatempa esplorado sugestis ke la ordinara individuo kapablas distingi inter pli ol unu triliono da unikaj odoroj.[32] Esploristoj en la plej ĝisdata studo notis ke ĉi tiu takso estas "konservativa" kaj ke kelkaj testatoj — testoj inkludis elprovi la psikofizikajn respondojn al kombinaĵoj de pli ol 128 unikaj odormolekuloj kun kombinaĵoj kunmetitaj el ĝis 30 malsamaj komponantaj molekuloj — eble kapablas deĉifri inter mil trilionoj da odorantoj, dum ili taksis ke ilia plej malbona faranto povas distingi inter 80 milionoj da odoroj.[33] La aŭtoroj de la studo konkludis, "Tio ĉi estas ege pli ol antaŭaj taksoj de distingeblaj olfaktaj stimuloj. Ĝi montras ke la homa flara sistemo, kun ĝiaj centoj da malsamaj flaraj ricevantoj, ege superas la aliajn sensojn laŭ la nombro de fizike malsamaj stimuloj kiujn ĝi povas distingi."[34] Tamen, la aŭtoroj ankaŭ notis ke la kapableco distingi inter odoroj ne estas analoga al la kapablo konsekvence identigi ilin kaj ke eksperimentatoj ne estas tipe kapablaj identigi individuajn odorajn stimulantojn el la odoroj kiujn la esploristoj kunmetis de multaj odoraj molekuloj. La studon forte kritikis sciencisto Markus Meister, kiu skribis ke "ekstravagancaj asertoj estas bazitaj sur eraroj pri matematika logiko".[35][36] La logikon de ĉi tiu referaĵo laŭvice kritikis denove la aŭtoroj de la origina referaĵo.[37]
Genetiko de olfakto
Diversaj homoj flaras diversajn odorojn kaj plejmultaj diferencoj estas kaŭzitaj de genetikaj diferencoj.[38] Kvankam odorantaj ricevantaj genoj konsistigas iun el la grandaj genaj familioj en la homa genaro, nur manpleno da genoj estas ligitaj konklude al specifaj odoroj. Ekzemple, la odoranta ricevanto OR5A1 kaj ĝiaj genetikaj variantoj (aleloj) estas responsaj por nia kapablo (aŭ malkapablo) flari β-iononon, grava aromo en manĝaĵoj kaj trinkaĵoj.[39] Simile, la odoranta ricevanto OR2J3 rilatas al la kapablo sensi la gresosimilan odoron, cis-3-hexen-1-olo.[40] La prefero (aŭ malŝato) al koriandro estas ligita al la olfakta ricevanto OR6A2.[41]
Interagoj de olfakto kun aliaj sensoj
Olfakto kaj gustumado
Olfaktaj, gustumaj kaj triĝeminaj ricevantoj (ankaŭ nomita kemestezo) kune kontribuas al gusto. La homa lango povas distingi inter nur kvin apartaj kvalitoj de gusto, dum la nazo povas distingi inter enormaj nombroj da substancoj, eĉ en etaj kvantoj. Estas dum elspirado kiam okazas la olfakta kontribuo al gusto, kontraste al tio de vera flarado, kiu okazas dum la enspira fazo.[42] La flara sistemo estas la nura homa senso kiu preteriras la talamon kaj konektiĝas rekte al la antaŭcerbo.[43]
Olfakto kaj aŭdado
Oni montris ke olfakta kaj sona informoj konverĝas en la flaraj tuberetoj de ronĝuloj.[44] Oni proponis ke ĉi tiu neŭra konverĝo estigas percepton nomitan angle smound (odor-sono).[45] Dum gustumo rezultas el interagoj inter odoro kaj gusto, odor-sono eble rezultas el interagoj inter odoro kaj sono.
Kvantigi olfakton en industrio
Sciencistoj elpensis metodojn por kvantigi la intensecon de odoroj, precipe por analizi malagrablajn aŭ abomenajn odorojn liberigitajn de industria fonto en komunumon. Ek de la 1800-aj jaroj, industriaj landoj renkontis okazojn kie la proksimeco de industria fonto aŭ rubodeponejo produktis malfavorajn reagojn ĉe apudaj loĝantoj pri aerportata odoro. La baza teorio de odoranalizo estas mezuri tioman diluon per "pura" aero, kiu estas postulita por fari la samplon neapartigebla de la "pura" aŭ referenca normo. Pro tio ke ĉiu persono perceptas odoron malsame, odorpanelo konsistanta de pluraj homoj estas kunmetita, ĉiu ekflaranta la saman samplon de diluita aero. Portebla olfaktometro estas utiligebla por mezuri la kvanton de odoro.
Olfakto en plantoj kaj bestoj
La ĉiroj de plantoj estas tre sensivaj al aerportataj volatilaj organikaj kombinaĵoj. Parazitoj, ekzemple kuskuto, utiligas tion serĉante siajn preferatajn gastigantojn.[46] La emano de volatilaj kombinaĵoj estas mezurebla kiam foliaro estas paŝtata de bestoj. Minacitaj plantoj povas tiam defendi sin per kemiaĵoj, ekzemple movante taninajn kombinaĵojn al iliaj folioj. (Vidu ĉe Planta percepto).
La graveco kaj sensiveco de flarado varias inter malsamaj organismoj; plejmultaj mamuloj havas bonan flarsenson, dum plejmultaj birdoj ne, krom la procelarioformaj birdoj (ekz., petreloj kaj albatrosoj), certaj specioj de vulturoj kaj la kivioj. Inter mamuloj, la flarsenso estas evoluinta en la karnomanĝuloj kaj hufuloj, kiuj ĉiam devas esti konsciaj, unu pri la alia, kaj ĉe tiuj kiuj serĉas per odoro por ilia manĝaĵo, kiel talpoj.
Kalkuloj sugestantaj plian aŭ malplian sensivecon en diversaj specioj reflektas eksperimentajn trovojn de la reagoj de bestoj spertantaj aromojn en konataj ekstremaj diluoj. Ĉi tiuj estas, tial, bazitaj sur perceptoj de ĉi tiuj bestoj, anstataŭ nur naza funkcio. Tio estas, la odoro de la cerbo-rekonantaj centroj devas reagi al la stimulo sensata, por ke la besto montru respondon al la odoro testata. Oni taksis ke hundoj ĝenerale havas flarsenson proksimume cent mil al milionfoje pli akuta ol tiu de homo.[Citaĵo bezonata] Tion ĉi ne signifas ke ili estas superverŝitaj de odoroj kiujn homaj nazoj povas sensi; anstataŭe, tio signifas ke ili povas percepti molekulan ĉeeston kiam ĝi estas multe pli diluita en la aero.
Flarhundoj kiel grupo povas flari unu- ĝis dekmilion-foje pli akute ol homo, kaj spurhundoj, kiuj havas la plej akutan flarsenson de ajnaj hundoj,[citaĵo bezonata] havas nazon dek- al centmilion-foje pli sensiva ol homa nazo. Ili estis breditaj por la specifa celo spuri homojn kaj povas sensi odoran spuron kelkajn tagojn malnovan. La dua-plej-sensiva nazo estas tiu de la vertagoj, kiuj estis breditaj por spuri kaj ĉasi kuniklojn kaj aliajn malgrandajn bestojn.
Ursoj, ekzemple la bruna urso en partoj de Nordameriko, havas flarsenson sep fojojn pli akuta ol tio de la ĉashundoj, nepre utila por trovi subterajn manĝaĵon. Uzante siajn longajn ungojn, ursoj fosadas profundajn tranĉeojn serĉante tunelantajn bestojn kaj nestojn kaj ankaŭ radikojn, bulbojn kaj insektojn. Ursoj povas sensi la odoron de manĝaĵo for de ĝis 30 kilometroj. Pro ilia grandeco, ili ofte postpredas nove mortigitajn bestojn, forpelante la predantojn (inkluzive luparojn kaj homajn ĉasistojn).
La flarsenso estas malpli evoluigita en la katarinaj primatoj kaj mankas ĉe cetacoj, kiuj kompensas per bone evoluigita gustosenso.[Citaĵo bezonata] En kelkaj strepsirinoj, kiel la ruĝventra lemuro, odoraj glandoj troviĝas sur la kapo. En multaj specioj, olfakto estas tre agordita al feromonoj; maskla silkraŭpa moteo, ekzemple, povas sensi ununuran molekulon de bombikolo.
Ankaŭ fiŝoj havas bone evoluigitan flarsenson, kvankam ili loĝas en akva medio. Salmo utiligas sian flarsenson identigi kaj reveni al sia hejma rivereto. Katfiŝo uzas sian flarsenson identigi alian individuan katfiŝon kaj daŭrigi socian hierarkion. Multaj fiŝoj uzas la flarsenson por identigi eblajn partnerojn aŭ por atentigi al la ĉeesto de manĝaĵo.
Eviti endogamion
Pro tio ke endogamio (samsangeco) estas malutila, ĝi emas esti evitata. En la hejma muso, la ĉefa urina proteina (MUP) genfasko provizas tre polimorfan odoran signalon de genetika identeco kiu ŝajne subtenas la kapablon rekoni parencojn kaj eviti endogamion. Pro tio okazas malpli da pariĝoj inter musoj kiuj havas la samajn MUP haplotipojn ol oni atendus se okazus hazardaj pariĝoj.[47]
Vidu ankaŭ
Malfunkcio de flarado
- Anosmio perdo de la flarsenso
- Disosmio odoroj ŝajnas diferencaj ol la atendataj aŭ memoritaj
- Hiperosmio nekutime akuta flarsenso
- Hipoosmio reduktita sensiveco al odoroj
Aliaj temoj
- Nazo
- Vomeronaza organo
- Kemestezo
- Elektronika nazo
- Maŝino olfakto
- Naza administro flara translokigo
- Odoro
- Olfaktometro
- Olfakta eninga glio
- Flara laceco
- Fantosmio
- Odortransloka ilo
- Vibra Teorio de Olfakto
- Evoluo de olfakto
Referencoj
- ↑ (September 2015) “Structure–odour relationships reviewed in the postgenomic era”, Flavour and Fragrance Journal 30 (5), p. 342–361. doi:10.1002/ffj.3249.
- ↑ Charles Linsmeiser. (2011) “Sensation and Perception”, Psychology. Worth Publishers, p. 166–171. ISBN 978-1-4292-3719-2.
- ↑ Hussain A, Saraiva LR, Korsching SI (2009). “Positive Darwinian selection and the birth of an olfactory receptor clade in teleosts”, PNAS 106 (11), p. 4313–8. doi:10.1073/pnas.0803229106.
- ↑ . Taste, Smell, and Touch: Lecture Notes (1999).
- ↑ (2012-10-19) “Better Smelling Through Genetics: Mammalian Odor Perception”, Current Opinion in Neurobiology 18 (4), p. 364–369. doi:10.1016/j.conb.2008.09.020.
- ↑ G. A. Kimble. (1985) Topics in the History of Psychology, Volume 1. L. Erlbaum Associates.
- ↑ Buck L, Axel R (April 1991). “A novel multigene family may encode odorant receptors: a molecular basis for odor recognition”, Cell 65 (1), p. 175–87. doi:10.1016/0092-8674(91)90418-X.
- ↑ Gilad Y, Man O, Pääbo S, Lancet D (March 2003). “Human specific loss of olfactory receptor genes”, PNAS 100 (6), p. 3324–7. doi:10.1073/pnas.0535697100.
- ↑ Pinel, John P.J. (2006) Biopsychology.
- ↑ Turin L (December 1996). “A spectroscopic mechanism for primary olfactory reception”, Chemical senses 21 (6), p. 773–91. doi:10.1093/chemse/21.6.773.
- ↑ Turin L (June 2002). “A method for the calculation of odor character from molecular structure”, Journal of Theoretical Biology 216 (3), p. 367–85. doi:10.1006/jtbi.2001.2504.
- ↑ Keller A, Vosshall LB (April 2004). “A psychophysical test of the vibration theory of olfaction”, Nature Neuroscience 7 (4), p. 337–8. doi:10.1038/nn1215.
- ↑ Doty, R. L. (2001).
- ↑ Bear, Connors and Paradiso, Mark, Barry and Michael. (2007) Neuroscience: Exploring the Brain. Usono: Lippincott Williams & Wilkins, p. 265–275.
- ↑ Morris, H., & Schaeffer, J. P. (1953).
- ↑ Rothermel Markus (2014). “Cholinergic Inputs from Basal Forebrain Add an Excitatory Bias to Odor Coding in the Olfactory Bulb”, J. Neurosci. 34 (13), p. 4654–4664. doi:10.1523/JNEUROSCI.5026-13.2014.
- ↑ Kapoor Vikrant (2016). “Activation of raphe nuclei triggers rapid and distinct effects on parallel olfactory bulb output channels”, Nat. Neurosci. 19, p. 813–4. doi:10.1038/nn.4219.
- ↑ Shea Stephen D. (2008). “Noradrenergic Induction of Odor-Specific Neural Habituation and Olfactory Memories”, J. Neurosci. 28 (42), p. 10711–10719. doi:10.1523/JNEUROSCI.3853-08.2008.
- ↑ Margot C (July 2009). “A noseful of objects”, Nat. Neurosci. 12 (7), p. 813–4. doi:10.1038/nn0709-813.
- ↑ Zhou W, Chen D (2009). “Binaral rivalry between the nostrils and in the cortex”, Curr Biol 19 (18), p. 1561–5. doi:10.1016/j.cub.2009.07.052.
- ↑ Navarrete-Palacios E, Hudson R, Reyes-Guerrero G, Guevara-Guzmán R (July 2003). “Lower olfactory threshold during the ovulatory phase of the menstrual cycle”, Biological Psychology 63 (3), p. 269–79. doi:10.1016/S0301-0511(03)00076-0.
- ↑ Boehm T, Zufall F (February 2006). “MHC peptides and the sensory evaluation of genotype”, Trends in Neurosciences 29 (2), p. 100–7. doi:10.1016/j.tins.2005.11.006.
- ↑ Santos PS, Schinemann JA, Gabardo J, Bicalho Mda G (April 2005). “New evidence that the MHC influences odor perception in humans: a study with 58 Southern Brazilian students”, Hormones and Behavior 47 (4), p. 384–8. doi:10.1016/j.yhbeh.2004.11.005.
- ↑ Porter RH, Cernoch JM, Balogh RD (1985). “Odor signatures and kin recognition”, Physiol Behav 34 (3), p. 445–8. doi:10.1016/0031-9384(85)90210-0.
- ↑ Weisfeld GE, Czilli T, Phillips KA, Gall JA, Lichtman CM (July 2003). “Possible olfaction-based mechanisms in human kin recognition and inbreeding avoidance.”, Journal of experimental child psychology 85 (3), p. 279–95. doi:10.1016/S0022-0965(03)00061-4.
- ↑ Lundström JN, Boyle JA, Zatorre RJ, Jones-Gotman M (August 2009). “The neuronal substrates of human olfactory based kin recognition”, Human Brain Mapping 30 (8), p. 2571–80. doi:10.1002/hbm.20686.
- ↑ Wilson DA (June 2001). “Receptive fields in the rat piriform cortex.”, Chemical senses 26 (5), p. 577–84. doi:10.1093/chemse/26.5.577.
- 1 2 3 Leon M, Johnson BA (2003). “Olfactory coding in the mammalian olfactory bulb”, Brain Res. Brain Res. Rev. 42 (1), p. 23–32. doi:10.1016/S0165-0173(03)00142-5.
- ↑ Rokni, D. (2014). “An olfactory cocktail party: figure-ground segregation of odorants in rodents”, Nature Neuroscience 17 (9), p. 1225–1232. doi:10.1038/nn.3775.
- 1 2 Auffarth, B. (2013). “Understanding smell -- the olfactory stimulus problem”, Neuroscience & Biobehavioral Reviews 37 (8), p. 1667–1679. doi:10.1016/j.neubiorev.2013.06.009.
- ↑ Soucy, Edward R. (2009). “Precision and diversity in an odor map on the olfactory bulb”, Nature Neuroscience 12.2 (2), p. 210–220. doi:10.1038/nn.2262.
- ↑ Helen Briggs. "Nose can detect one trillion odours", 20-a de marto 2014, paĝoj 23 March 2014.
- ↑ Sarah C.P. Williams. "Human Nose Can Detect a Trillion Smells", 20-a de marto 2014. Kontrolita 23-a de marto 2014.
- ↑ (21-a de marto 2014) “Humans Can Discriminate More than 1 million Olfactory Stimuli”, Science 343 (6177), p. 1370–1372. doi:10.1126/science.1249168.
- ↑ (2014) “Can Humans Really Discriminate 1 Trillion Odors?”, ArXiv arXiv:1411.0165.
- ↑ Meister, M. (2015). “On the dimensionality of odor space”, eLife 4. doi:10.7554/eLife.07865.
- ↑ (2015) “On the dimensionality of olfactory space”, BioRxiv. doi:10.1101/022103.
- ↑ Howgego, J. (2013) Sense for scents traced down to genes.
- ↑ Jaeger SR, McRae JF, Bava CM, Beresford MK, Hunter D, Jia Y, Chheang SL, Jin D, Peng M, Gamble JC, Atkinson KR, Axten LG, Paisley AG, Tooman L, Pineau B, Rouse SA, Newcomb RD (2013). “A Mendelian Trait for Olfactory Sensitivity Affects Odor Experience and Food Selection”, Current Biology 23 (16), p. 1601–1605. doi:10.1016/j.cub.2013.07.030.
- ↑ McRae JF, Mainland JD, Jaeger SR, Adipietro KA, Matsunami H, Newcomb RD (2012). “Genetic Variation in the Odorant Receptor OR2J3 is Associated with the Ability to Detect the "Grassy" Smelling Odor, cis-3-hexen-1-ol”, Chemical Senses 37 (7), p. 585–593. doi:10.1093/chemse/bjs049.
- ↑ Callaway, E. (2012) taste of coriander linked to genetic variants Nature News, 12 Sep 2012
- ↑ Masaoka Y, Satoh H, Akai L, Homma I (2010). “Expiration: The moment we experience retronasal olfaction in flavor”, Neurosci Lett. 473 (2), p. 92–96. doi:10.1016/j.neulet.2010.02.024.
- ↑ Annual Review of Psychology (February 2001), 52 (1), pg. 423-452,Richard L. Doty
- ↑ Wesson DW, Wilson DA (2010). “Smelling Sounds: Olfactory-auditory convergence in the olfactory tubercle”, J Neurosci 30 (8), p. 3013–1021. doi:10.1523/JNEUROSCI.6003-09.2010.
- ↑ Peeples, Lynne. Scientific American, ''Making scents of sounds'' Lynne Peeples, 23 February 2010, accessed 25 February 2010. Scientificamerican.com. Alirita 2012-12-30.
- ↑ Fountain, Henry, "This Plant Has the Sense of Smell (Loves Tomatoes, Hates Wheat)", The New York Times, 3-a de oktobro 2006.
- ↑ (2007) “The genetic basis of inbreeding avoidance in house mice”, Curr. Biol. 17 (23), p. 2061–6. doi:10.1016/j.cub.2007.10.041.
Plua legado
- Gordon Shepherd Neurogastronomy: How the Brain Creates Flavor and Why It Matters New York : Columbia University Press, 2012 ISBN 978-0-231-15910-4
Eksteraj ligiloj
- Mammalian Odor Perception through Genetics
- Research on Interesting Questions About Smells Arkivigite je 2010-01-22 per la retarkivo Wayback Machine
- Insect Olfaction of Plant Odour
- Smells and Odours - How Smell Works at thenakedscientists.com
- Olfaction at cf.ac.uk
- Structure-odor relations: a modern perspective at flexitral.com (PDF)
- Chirality & Odour Perception at leffingwell.com
- ScienceDaily Artille 08/03/2006, Quick -- What's That Smell? Time Needed To Identify Odors Reveals Much About Olfaction at sciencedaily.com
- Scents and Emotions Linked by Learning, Brown Study Shows at brown.edu.com
- Sense of Smell Institute at senseofsmell.org Arkivigite je 2007-06-12 per la retarkivo Wayback Machine. Research arm of international fragrance industry's The Fragrance Foundation
- Olfactory Systems Laboratory at Boston University Arkivigite je 2008-06-28 per la retarkivo Wayback Machine
- Smells Database
- Olfaction and Gustation, Neuroscience Online (electronic neuroscience textbook by UT Houston Medical School)