Kiel legi la taksonomion | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||
| Biologia klasado | ||||||||||||
| ||||||||||||
 | ||||||||||||
Aliaj Vikimediaj projektoj | ||||||||||||


_Edit.jpg.webp)

.jpg.webp)
Anuroj[1] (science: Anura el antikva greka an-, sen + oura, vosto, aŭ senvostaj amfibioj) estas la biologia ordo de karnovoraj mallongkorpaj senvostaj bestoj en la klaso amfibioj kiu inkluzivas ranojn kaj bufojn. Tamen, ne estas klara scienca distingo inter ranoj kaj bufoj. Unu familio de anuroj la diskolanguloj, havas ecojn de ambaŭ ranoj kaj bufoj.
La plej antikva fosilia "pra-rano" nome Triadobatrachus aperis en la komenco de Triaso de Madagaskaro, sed datado per molekula horloĝo sugestas ke ties originoj povas etendi plue reen al Permio, antaŭ 265 milionoj da jaroj. Ranoj estas amplekse distribuataj, game el tropikoj al ĉearktaj regionoj, sed la plej granda koncentro de specidiverseco estas en tropikaj pluvarbaroj. Estas proksimume 4 800 registritaj specioj, ĉirkaŭkalkule 85% el vivantaj amfibiaj specioj. Ili estas ankaŭ unu el la kvin plej diversaj ordoj de vertebruloj.
La korpokonsisto de plenkreska rano estas ĝenerale karakterizata de fortika korpo, elstaregaj okuloj, fenda lango, membroj falditaj suben, kaj foresto de vosto ĉe plenkreskuloj. Krom vivi en nesala akvo kaj sur seka tero, la plenkreskuloj de kelkaj specioj estas adaptitaj al loĝado subtera aŭ en arboj. La haŭto de ranoj estas glandenhava, kun sekrecioj gamaj el malbongustegaj al venenaj. Verukenhavaj specioj de ranoj tendencas esti nomataj bufoj sed la distingo inter ranoj kaj bufoj estas bazita sur neformalaj nomigaj konvencioj koncentrataj al la verukoj pli ol al taksonomio aŭ al evolua historio; kelkaj bufoj estas pli proksime rilataj al ranoj ol aliaj bufoj. La haŭto de ranoj varias en koloro el bone-kamuflitaj makulecaj bruna, griza kaj verda al tre koloraj modeloj de bril-ruĝa aŭ -flava kaj nigra por averti pri veneneco kaj predantoforigo.
Ranoj tipe demetas siajn ovojn en akvo. El la ovoj eliras larvoj nomataj ranidoj kiuj havas vostojn kaj internajn brankojn. Ili havas tre specializitajn asprajn buŝopartojn taŭgajn por herbovoraj, ĉiomanĝantaj aŭ planktonvorulaj dietoj. La vivociklo estas kompletita kiam ili metamorfozas al plenkreskuloj. Kelkaj specioj demetas ovojn surteren aŭ rekte eksteriras al stadio de ranido. Plenkreskaj ranoj ĝenerale havas karnovoran dieton konsistantan el malgrandaj senvertebruloj, sed ekzistas ankaŭ ĉiomanĝantaj specioj kaj kelkaj manĝas fruktojn. Ranoj estas tre efikaj je konverto de tio kion ili manĝas al korpomaso. Ili krome estas tre grava manĝofonto por predantoj kaj parto de la dinamiko de manĝoreto de multaj el la ekosistemoj de la mondo. La haŭto estas duontralasebla, kio faras ilin vundeblaj pro senakviĝo, kaj tiele ili ĉu vivas en humidaj lokoj aŭ havas specialajn adaptaĵojn por elteni sekajn habitatojn. Ranoj produktas ampleksan gamon de voĉoj, ĉefe dum la reprodukta sezono, kaj montras multajn diferencajn tipojn de kompleksaj kutimoj por allogi partnerojn, por forigi predantojn kaj ĝenerale por survivi.
Ranoj estas valoraj kiel manĝo de homoj kaj ankaŭ havas multajn kulturajn rolojn en literaturo, simbolismo kaj religio. Ranaj populacioj malpliiĝis grave el la 1950-aj jaroj. Pli ol unu triono da specioj estas konsiderataj minacataj de formorto kaj ĉirkaŭ cent dudek ŝajne iĝis formortintaj ekde la 1980-aj jaroj.[2] La nombro de disformaĵoj inter ranoj estas malpliiĝanta kaj ĵuse konata funga malsano, nome Ĥitridjomikozo, disvastiĝis tra la tuta mondo. Konservobiologoj laboras por kompreni la kaŭzojn de tiuj problemoj kaj por solvi ilin.
Taksonomio
Ĉirkaŭ 88% el la specioj de amfibioj estas klasitaj en la ordo Anura.[3] Tio inkludas ĉirkaŭ 4,810 speciojn en 33 familioj, el kiuj la Leptodaktiledoj (1,100 specioj), Hiledoj (800 specioj) kaj Ranedoj (750 specioj) estas la plej riĉaj en specioj.[3]
_tight_crop.jpg.webp)
La uzado de komunaj nomoj "rano" kaj "bufo" ne havas taksonomian justigon. El klasiga vidpunkto, ĉiuj membroj de la ordo Anura estas ranoj, sed nur membroj de la familio Bufonedoj estas konsiderataj "veraj bufoj". La uzado de la termino "rano" en komunaj nomoj kutime referencas al specioj kiuj estas akvaj aŭ duon-akvaj kaj havas mildajn, malsekajn haŭtojn; la termino "bufo" ĝenerale referencas al specioj kiuj estas surteraj kun sekaj, verukhavaj haŭtoj.[4][5] Estas nombraj esceptoj al tiu ordigo. La Eŭropa fajrzona bufo (Bombina bombina) havas iom verukhavan haŭton kaj preferas akvecan habitaton[6] dum la Panama ora rano (Atelopus zeteki) estas en la bufa familio Bufonedoj kaj havas mildan haŭton.[7]
La grupo de Anura inkludas ĉiujn modernajn ranojn kaj ajnajn fosiliajn speciojn kiuj kongruas ene de la anura difino. La karakteroj de anuraj plenkreskuloj inkludas: 9 aŭ malmultajn antaŭsakrajn vertebrojn, la esto de urostilaj formitaj aŭ fuziitaj vertebroj, sen vosto, longa kaj antaŭendekliva ilio, pli mallongaj antaŭaj membroj ol malantaŭaj, radiuso kaj ulno fuziitaj, tibio kaj fibulo fuziitaj, longecaj taloj, foresto de antaŭfronta osto, esto de hioida plato, malsupra makzelo sendenta (kun la escepto de Gastrotheca guentheri) konsistanta el tri paroj de ostoj (angulospleniaj, dentaj, kaj mentomekeliaj, el kiuj tiu lasta paro forestas ĉe la klado Pipoidea),[8] nesubtenita lango, limfaj spacoj subhaŭtaj, kaj muskolo, nome protractor lentis, ligita al la lenso de la okulo.[9] La anuraj larvoj aŭ ranidoj havas unusolan centran spiran spirtruon kaj buŝopartojn konsistantaj el keratinecaj bekoj kaj dentaĵoj.[9]

Ranoj kaj bufoj estas ĝenerale klasitaj en tri subordojn: Malnovbatrakoj, kiuj inkludas kvar familiojn de praranoj; Mezbatrakoj, kiuj inkludas kvin familiojn de pli evoluciitaj intermezaj ranoj; kaj Novbatrakoj, laŭ multe la plej granda grupo, kiu enhavas la ceterajn 24 familiojn de modernaj ranoj, inkludante plej oftajn speciojn tra la tuta mondo. La subordo de Novbatrakoj estas siavice dividita en du superfamilioj nome Hyloidea kaj Ranoidea.[10] Tiu klasigo estas bazita sur trajtoj de morfologio kiaj la nombro de vertebroj, la strukturo de la brusta skapolo, kaj la morfologio de ranidoj. Kvankam tiu klasigo estas tre malferme akceptita, la rilataro inter familioj de ranoj estas ankoraŭ studota.[11]
Kelkaj specioj de anuroj pretas hibridiĝi. Por ekzemplo, la manĝebla rano (Pelophylax esculentus) estas hibrido inter Pelophylax lessonae kaj la ridorano (P. ridibundus).[12] La fajrventraj bufoj Bombina bombina kaj la B. variegata estas similaj laŭ inklino al hibridiĝo. Tiuj estas malpli fekundaj ol ties gepatroj, kio kondukas al hibrida zono kie la hibridoj estas hegemoniaj.[13]
Biologia klasado



Estas proksimume 5 000 specioj nune konataj de anuroj. La vivantaj anuroj estas ofte dividitaj al tri subordoj.
- Subordo Malnovbatrakoj aŭ Archaeobatrachia - 4 familioj, 6 genroj, 28 specioj
- Subordo Mezbatrakoj aŭ Mesobatrachia - 6 familioj, 20 genroj, 168 specioj;
- Subordo Novbatrakoj aŭ Neobatrachia - 19 familioj, 310 genroj, 4688 specioj
- Artroleptedoj
- Braĥicefaledoj
- Bufonedoj (Bufonidae), inter ili la genro bufoj
- Centrolenedoj
- Dendrobatedoj, inter ili ekzemple la specio Dendrobates azureus
- Heleofrinedoj
- Hemisotedoj
- Hiledoj (Hylidae)
- Hiperoliedoj
- Leptodaktiledoj
- Manteledoj
- Mikrohiledoj
- Mjobatraĥedoj
- Ranedoj (Ranidae), inter ili la genro ranoj
- Rakoforedoj
- Rinodermatedoj
- Soglosedoj
- Krome Brachycephaloidea
- Craugastoridae
- inter kiuj Pristimantis gretathunbergae
- Craugastoridae
- Telmatobiedoj
- Titikaka rano, Telmatobius culeus
- Telmatobiedoj
Morfologio kaj fiziologio
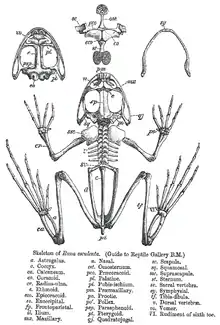
Ranoj ne havas voston, escepte kiam larvoj, kaj plej havas longajn malantaŭajn krurojn, longajn maleolajn ostojn, membranhavajn fingrojn, senungajn, grandajn okulojn, kaj mildan aŭ verukhavan haŭtaĵon. Ili havas mallongajn vertebrarojn, kun ne pli ol 10 liberaj vertebroj kaj fuziitaj vostostoj.[14] Kiel ĉe aliaj amfibioj, oksigeno povas pasi tra ties tre trapaseblaj haŭtoj. Tiu unika trajto permesas ilin resti en lokoj sen aliro al la aero, spirante tra siaj haŭtoj.[15] La ripoj estas malbone disvolvigitaj, kaj tiele la pulmoj estas plenigitaj per la buŝa pumpado kaj rano senigita el siaj pulmoj povas elteni siajn korpofunkciojn sen ili.[15] Por ke la haŭto utilas kiel spira organo, ĝi devas resti humida. Tio faras ke ranoj tuŝeblas de variaj substancoj kiujn ili povas traafi en la medio, kelkaj el kiuj povas esti venenaj kaj povas dissolviĝi en akvofilmo kaj povas pasi al ties sangofluon. Tio povas esti unu el la kaŭzoj de la tutmonda malpliiĝo en ranopopulacioj.[16][17][18][19]
Ranoj gamas laŭgrande el la ĵus malkovrita 7.7 mm Paedophryne amauensis de Papuo-Nov-Gvineo[20] al la 300 mm goliata rano (Conraua goliath) de Kameruno. La haŭto pendas malfikse sur la korpo pro la manko de malfiksa konektohisto. Ranoj havas tri okul-harajn membranojn: unu estas travidebla por protekti la okulojn subakve, kaj du varias el travideblaj al netravideblaj. Ili havas timpanon en ĉiu flanko de ties kapoj kiu utilas al aŭdado kaj, en kelkaj specioj, estas kovrataj de haŭtaĵo. Veraj bufoj tute ne havas dentojn, sed plej ranoj havas ilin, specife pedicelajn dentojn en kiuj la krono estas separata el la radiko per fibra histo. Tiuj estas ĉe la bordo de supra makzelo kaj ankaŭ estas vomeraj dentoj sur la palato de ties buŝoj. Ne estas dentoj en la malsupra makzelo kaj ranoj kutime englutas sian manĝon kiel tuto. La dentoj estas ĉefe uzataj por kapti la predon kaj teni ĝin surloke ĝis engluto, procezo helpata de la retirigo de la okuloj en la kapo.[21] La afrika taŭrorano (Pyxicephalus), kiu predas relative grandajn animalojn kiaj musoj kaj aliaj ranoj, havas konusformajn ostoprojekciojn nomitajn odontoidaj procezoj ĉe la antaŭo de la malsupra makzelo kio funkcias kiel dentoj.[3]

Piedoj kaj kruroj
La strukturo de la piedoj kaj kruroj ege varias inter ranospecioj, dependante parte ĉu ili loĝas ĉefe surgrunde, en akvo, en arboj aŭ en nestotruoj. Ranoj devas kapabli moviĝi rapide tra sia medio por kapti predojn kaj fuĝi el predantoj, kaj nombraj adaptoj helpas ilin fari tion. Plej ranoj estas bonaj saltantoj aŭ posteuloj de prauloj kiuj estis tio, kaj multo de muskol-skeleta morfologio modifiĝis tiucele. La tibio, fibulo, kaj tarsoj fuziiĝis en unusola, forta osto, ĉar havas la radiuson kaj la ulnon en la antaŭaj kruroj (kiuj povas absorbi la frapon de surteriĝo). La metatarsoj plilongiĝis por aldoni al la kruro longon kaj permesi al la rano peliĝi kontraŭ la grundo dum pli longa tempoperiodo en la eksalto. La ilio plilongiĝis kaj formis movevlan artikon kun la sakro kiu, ĉe specialistaj saltantoj kiaj ranedoj kaj hiledoj, funkcias kiel aldona krurartiko por havi plian povon en la saltoj. La vostovertebroj fuziiĝis en urostilo kiu estas retirita ene de la pelvo. Tio permesas ke la forto estu transpasata el la kruroj al la korpo dum salto.[14]

(Rana temporaria)

La muskola sistemo simile modifiĝis. La malantaŭaj kruroj de praranoj supozeble enhavis parojn de muskoloj kiuj estus agintaj male (unu muskolo por faldi la genuon, alia muskolo por etendigi ĝin), kiel estas vidata en plej el aliaj krurhavaj animaloj. Tamen, ĉe modernaj ranoj, preskaŭ ĉiuj muskoloj estis modifitaj por kontribui al la agado salti, kun nur kelkaj malgrandaj muskoloj restantaj por revenigi la kruron reen al la starta loko kaj elteni la sintenon. Ankaŭ la muskoloj estis ege pligrandigitaj, kaj la ĉefaj krurojmuskoloj respondecas pro ĉirkaŭ 17% el la totala amaso de la rano.[22]
Multaj ranoj havas membranhavajn piedojn kaj la grado de membraneco estas rekte proporcia al la kvanto de tempo kiun la specio pasas en akvo.[23] La komplete akvoloĝanta Afrika nana rano (specioj de genro Hymenochirus) havas tute membranhavajn fingrojn, dum tiuj de la arborano de White (Litoria caerulea), nome arboloĝanta specio, estas nur kvarone aŭ duone membranhavaj.[24]
La arboranoj havas kusenetojn en la finoj de siaj fingrojn por helpi alkroĉado al vertikalaj surfacoj. Tiuj ne estas suĉokusenetoj, anstataŭe la surfaco konsistas el kolonaj ĉeloj kun ebenaj pintoj kun malgrandaj malplenaĵoj inter ili lubrikitaj de mukaj glandoj. Kiam la ranoj aplikas premon, la ĉeloj algluiĝas al la malregulaĵoj de la surfaco kaj la alkroĉo estas tenita pere de surfaca tensio. Tio permesas la ranon grimpi sur mildaj surfacoj, sed la sistemo ne funkcias efike kiam la kusenetoj estas tro malsekaj.[25]
En multaj arboranoj, malgranda "intermetita strukturo" en ĉiu fingro pliigas la surfacan areon tuŝantan la subtavolon. Krome, ĉar saltado tra arboj povas esti danĝera, multaj arboranoj havas taliajn artikojn kiuj permesas kaj saltadon kaj piediradon. Kelkaj ranoj kiuj loĝas alte en arboj eĉ posedas prilaboratan gradon de membraneco inter siaj fingroj. Tio permesas ke la ranoj "paraŝutas" aŭ faras kontrolatan glitadon el unu loko en la kanopeo al alia.[26]
Grundoloĝantaj ranoj ĝenerale ne havas la adaptaĵojn de akvoloĝantaj kaj arboremaj ranoj. Plej havas pli malgrandajn fingrokusenojn, se estas io tia, kaj malmultan membranecon. Kelkaj nestotruaj ranoj kiaj la Kouĉa skafiopo (Scaphiopus couchii) havas klapecan fingretendaĵon ĉe malantaŭaj piedoj, nome tuberaĵon el keratino ofte referencata kiel ŝpato, kio helpas ili nestotrui.[27]
Foje dum la stadio de ranidoj, unu el la disvolviĝantaj malantaŭaj kruroj estas manĝata de predanto kiel ekzemple libela nimfo. En kelkaj okazoj, plena kruro ankoraŭ kreskas, sed en aliaj ne, kvankam la rano povas ankoraŭ vivi sian normalan vivodaŭron kun nur tri kruroj. Foje, parazita platvermo (Ribeiroia ondatrae) elfosas en la malantaŭo de ranido, kio kaŭzas rearanĝon de la kruraj ĝermoĉeloj kaj la rano povas disvolvigi kroman kruron aŭ eĉ du.[28]

Haŭto
La haŭto de rano estas protekta, havas spiran funkcion, povas absorbi akvon kaj helpas kontrolon la korpotemperaturon. Ĝi havas multajn glandojn, partikularle sur la kapo kaj la dorso, kiuj ofte eksudas malbongustajn kaj venenajn substancojn. La sekrecio estas ofte glueca kaj helpas teni la haŭton malseka, protektas kontraŭ la eniro de ŝimoj kaj bakterioj, kaj faras la animalon glitiga kaj pli kapabla fuĝi el predantoj.[29] La haŭto estas mudo|mudata post kelkaj semajnoj. Ĝi kutime elŝiriĝas en la mezo de la dorso kaj tra la ventro, kaj la rano eligas siajn brakojn kaj krurojn. La mudata haŭtaĵo estas poste movigita al la kapo kie ĝi estas rapide manĝata.[30]
Estante malvarmsangaj, ranoj adoptas taŭgajn kondutajn modelojn por reguligi sian temperaturon. Por plivarmiĝi, ili povas moviĝi al la suno aŭ al varma surfaco; se ili trovarmiĝas, ili povas moviĝi en la ombro aŭ adoptante sintenon kiu eksponas la minimuman areon de haŭto al la aero. Tiu sinteno estas uzata ankaŭ por eviti akvoperdon kaj inkludas ke la rano kaŭras ĉe la subtavolo kun siaj manoj kaj piedoj metitajn sub siaj mentono kaj korpo.[31] La koloro de la haŭto de rano estas uzata por termoregulado. En malvarmetaj marĉokondiĉoj, la koloro estos pli malhela ol dum varma seka tago. La griza saŭmonesta arborano (Chiromantis xerampelina) kapablas eĉ turniĝi blanka por minimumi la ŝancon trovarmiĝi.[32]
Multaj ranoj kapablas absorbi akvon kaj oksigenon rekte tra la haŭto, speciale ĉirkaŭ la pelva areo, sed la permeableco de la rana haŭto povas rezulti ankaŭ en akvoperdo. Glandoj situas sur la tuta korpo eksudas mukon kiu helpas teni la haŭton malseka kaj malpliigas vaporigon. Kelkaj glandoj en la manoj kaj brusto de maskloj estas specializitaj en produktado de gluecaj sekrecioj por helpi en ampleksumo. Similaj glandoj en arboranoj produktas gluecan substancon sur la gluigaj diskoj de la piedoj. Kelkaj arboranoj malpliigas akvoperdon per havado de akvimuna tavolo de haŭto, kaj kelkaj sudamerikaj specioj kovras sian haŭton per vakseca sekrecio. Aliaj ranoj havas adoptitajn kutimojn por konservi akvon, inklude eston noktula kaj ripozado en akvo-konserva situacio. Kelkaj ranoj povas ankaŭ ripozi en grandaj grupoj el kiu ĉiu rano premiĝas kontraŭ siaj najbaroj. Tio malpliigas la kvanton de haŭto eksponita al la aero aŭ al seka surfaco, kaj tiele malpliigas la akvoperdon.[31] La Vudhaŭsa bufo (Bufo woodhousii), se havas aliron al akvo post estado en seka loko, sidas en malprofundaĵo por reakviĝi.[33] La masklo de harrano aŭ triĥobatraĥo (Trichobatrachus robustus) havas dermajn papilojn projekciantaj el ties malsupra dorso kaj femuroj, kio havigas al ĝi krispan aspekton. Ili enhavas sangotubetojn kaj oni supozas ke ili pliigas la areon de haŭto disponebla por spirado.[34]
Kelkaj specioj havas ostecajn platojn enmetitajn en sia haŭto, nome trajto kiu ŝajne evoluis sendepende kelkajn fojojn.[35] En kelkaj aliaj specioj, la haŭto ĉe la pinto de la kapo estas kompaktigitaj kaj la konekta histo de la dermo estas kun-ostigita kun la ostoj de la kranio (eksostozo).[36][37]
Kamuflado estas ofta defenda meĥanismo ĉe ranoj. Plej kamuflaj ranoj estas noktemaj; dumtage ili serĉas sintenon kie ili povas miksiĝi en la fono kaj resti sendetekte. Kelkaj ranoj havas kapablon ŝanĝi koloron, sed tio estas kutim limigita al malgranda gamo de koloroj. Por ekzemplo, la arborano de White (Litoria caerulea) varias inter helverda kaj bruneca laŭ la temperaturo, kaj la Pacifika arborano (Pseudacris regilla) havas verdajn kaj brunajn morfojn, senmakule aŭ makule, kaj ŝanĝas koloron depende el la jarepoko kaj ĝenerala fonkoloro.[38] Trajtoj kiaj verukoj kaj haŭtofaldoj estas kutime ĉe grundoloĝantaj ranoj, ĉe kiuj milda haŭto ne havigas tiom efikan kamufladon. Kelkaj ranoj ŝanĝas koloron inter nokto kaj tago, ĉar lumo kaj malsekeco stimulas la pigmentoĉelojn kaj faras ke ili etendiĝu aŭ kuntiriĝu.[15]

Spirado kaj cirkulado
La haŭto de rano estas permeabla al oksigeno kaj al karbona duoksido, same kiel al akvo. Estas sangotubetoj ĉe la surfaco de la haŭto kaj kiam rano estas subakvo, oksigeno disiĝas rekte en la sango. Kiam ne subakve, rano spiras per procezo konata kiel buŝa pumpado. Ties pulmoj estas similaj al tiuj de homoj, sed la brustomuskoloj ne intervenas en spirado, kaj ne ekzistas ripoj aŭ diafragmo kiu helpu movigu la aeron enen kaj eksteren. Anstataŭe, ĝi ŝvelas sian gorĝon kaj enmetas aeron tra la naztruoj, kio en multaj specioj povas fermiĝi per valvoj. Kiam la suba muŝo estas kumpremita, aero estas pelita en la pulmoj.[39] La tute akva KAlimanta barburulo (Barbourula kalimantanensis) estas la unua rano konata kiu tute ne havas pulmojn.[40]
Ranoj havas tri-ĉambrajn korojn, trajto kiun ili kunhavas kun lacertoj.[41] Oksigenenhava sango el pulmoj kaj sen-oksigena sango el la histoj de spirado eniras en la koron tra separataj aŭrikloj. Kiam tiuj ĉambroj kuntiriĝas, la du sangorojoj pasas en komunan ventriklon antaŭ esti pumpita tra spirala valvo al la taŭga tubeto, nome aorto por oksigenenhava sango kaj pulmarterio por sen-oksigena sango. La ventriklo estas parte dividita en mallarĝaj kavaĵoj kiuj malgrandigas la mikson de la du sangotipoj. Tiuj trajtoj permesas al ranoj havi pli altan metabolan proporcion kaj esti pli aktivaj ol alimaniere estus ebla.[41]
Kelkaj specioj de ranoj havas adaptaĵojn kiuj permesas ilin survivi en oksigenmanka akvo. La rano de la Lago Titikaka (Telmatobius culeus) estas unu tia specio kaj havas faldecan haŭton kiu pliigas sian surfacan areon por faciligi la gasan interŝanĝon. Ĝi normale ne faras uzadon de siaj rudimentaj pulmoj sed ĝi foje plialtigas kaj malaltigas sian korporitmon en la lagofundo por pliigi la fluadon de akvo ĉirkaŭ ĝi.[42]

Digesto kaj ekskrecio
Ranoj havas makzelajn dentojn laŭlonge de siaj supraj makzeloj kiuj estas uzataj por teni manĝaĵon antaŭ engluti ĝin. Tiuj dentoj estas tre malfortaj, kaj ne povas esti uzataj por maĉado aŭ kaptado kaj damaĝo de lertaj predoj. Anstataŭe, la rano uzas sian gluecan, fendan langon por kapti muŝojn kaj aliajn malgrandajn moviĝantajn predojn. La lango normale kuŝas volvita en la buŝo, libera ĉe la dorso kaj ligita al la makzelo frunte. Ĝi povas esti elpafita kaj retirita je granda rapido.[23] Kelkaj ranoj ne havas langon kaj metas manĝaĵon en siaj buŝoj nur per siaj manoj.[23] La okuloj helpas en la englutado de manĝaĵoj ĉar ili povas esti retiritaj tra truoj en la kranion kaj helpas peli manĝaĵojn suben al la gorĝo.[23] La manĝaĵo poste moviĝas tra la ezofago en la stomakon kie la digestaj enzimoj estas aldonitaj kaj ĝi estas kirlita. Ĝi poste sekvas al la malgranda intesto (duodeno kaj ileo) kie plej de la digesto okazas. Pankreata suko el la pankreato, kaj la galo, produktita de la hepato kaj stokita en la gal-veziko, estas sekreciataj en la malgranda intesto, kie la fluaĵoj digestigas la manĝaĵon kaj la nutraĵoj estas absorbitaj. La manĝoruboj pasas al la granda intesto kie la troa akvo estas forigita kaj la restaĵoj estas pasataj tra la kloako.[43]
Kvankam adaptitaj al la sutera vivo, ranoj similas al la nesalakvaj fiŝoj en sia nekapablo konservi la korpakvon efektive. Kiam ili estas surtere, multa akvo perdiĝas pro evaporado tra la haŭto. La ekskrecia sistemo estas simila al tiu de mamuloj kaj estas du renoj kiuj forigas nitrogenajn produktojn el la sango. Ranoj produktas grandajn kvantojn de dissolvita urino por forigi la venenajn produktojn el la renaj tubetoj.[44] La nitrogeno estas ekskreciata kiel amoniako fare de ranidoj kaj akvaj ranoj sed ĉefe kiel ureo, malpli venena produkto, fare de plej surteraj plenkreskuloj. Kelkaj specioj de arborano kun malmulta aliro al akvo ekskrecias la eĉ malpli venenan ureatan acidon.[44] La urino passs laŭlonge de paraj ureteroj al la urina veziko el kiu ĝi estas forigita periode al la kloako. Ĉiuj korporubaĵoj eliras el la korpo tra la kloako kiu finas en kloaka elirejo.[45]
Reprodukta sistemo

Ĉe la maskla rano, la du testikoj estas ligitaj al la renoj kaj spermo pasas al la renoj tra fajnaj tuboj nome spermo-duktoj aŭ eferentaj duktoj. Ĝi poste vetura tra la ureteroj, kiuj estas logike konataj kiel urinogenitaj duktoj. Ne estas peniso, kaj la spermo estas eligita el la kloako rekte sur la ovojn kiam la ino demetas ilin. La ovarioj de la ranino estas apud la renoj kaj la ovoj pasas laŭ paro de ovoduktoj kaj tra la kloako al la ekstero.[45]
Kiam la ranoj pariĝas, la masklo grimpas surdorse de la ino kaj per siaj antaŭaj kruroj ĉirkaŭas ŝian korpon, ĉu malantaŭ ŝiaj antaŭaj kruroj aŭ ĝuste antaŭ ŝiaj malantaŭaj kruroj. Tiu sinteno estas nomita ampleksumo kaj povas pludaŭri eĉ dum kelkaj tagoj.[46] La maskla rano havas kelkajn hormon-dependajn duarangajn seksajn karakterojn. Tiuj povas esti la disvolvigo de specialaj kusenetoj sur siaj dikfingroj en la reprodukta sezono, por doni al li firman kaptokapablon.[47] La brakumo de la maskla rano dum la ampleksumo stimulas la inon ovodemeti, kutime volvita en gelateno, kiel frajo.[45] En multaj specioj la masklo estas pli malgranda kaj svelta ol la ino. Maskloj havas voĉajn kordojn kaj faras gamon de kvakoj, partikulare en la reprodukta sezono, kaj en kelkaj specioj ili havas ankaŭ voĉosakojn por amplifi la sonon.[45]
Nervosistemo
La ranoj havas tre disvolvigitan nervosistemon kiu konsistas el cerbo, spina kordo kaj nervoj. Multaj partoj de la rana cerbo korespondas kun tiuj de homoj. Ĝi konsistas el du flarloboj, du cerbaj hemisferoj, pineala glando, du vidloboj, cerbeto kaj mjela plilongigo. Muskolara kunordigo kaj sinteno estas kontrolata de la cerbeto, kaj la mjela plilongigo regulas spiradon, digeston kaj aliajn aŭtomatajn funkciojn.[45] La relativa grando de la cerbeto en ranoj estas multe pli malgranda ol ĝi estas en homoj. Ranoj havas dek parojn de cerbaj nervoj kiuj pasas informaron el la ekstero rekte al la cerbo, kaj dek parojn de spinaj nervoj kiuj pasas informaron el la pintoj de la cerbo tra la spina kordo.[45] Kontraste, ĉiuj amniotoj (mamuloj, birdoj kaj reptilioj) havas dekdu parojn de kraniaj nervoj.[48]

Vidkapablo
La okuloj de plej ranoj estas situaj ĉe ĉiu flanko de la kapo ĉe la pinto kaj projekcias eksteren kiel duonsferaj elstaraĵoj. Ili havigas duokulan vidkapablon al kampo de 100° antaŭe kaj al totala vidkampo de preskaŭ 360°.[49] Ili povas estis la nura parto de por la cetero enakva rano kiu estas eksterakve. Ĉiu okulo havas fermeblajn suprajn kaj malsuprajn palpebrojn kaj niktiton kiu havigas plian protekton, speciale kiam la rano naĝas.[50] Membroj de la akva familio de Pipedoj havas la okulojn situaj ĉe la kapopinto, loko pli bone taŭga por detektado de predo en la supra akvo.[49] La irisoj montras gamon de koloroj kaj la pupiloj gamon de formoj. La komuna bufo (Bufo bufo) havas orkolorajn irisojn kaj horizontalajn fendecajn pupilojn, la ruĝokula arborano (Agalychnis callidryas) havas vertikalajn fendopupilojn, la venenaj sagetranoj havas malhelajn irisojn, la fajrventraj bufoj (specioj de la genro Bombina) havas triangulajn pupilojn kaj la tomatoranoj (specioj de la genro Dyscophus) havas cirklajn pupilojn. La irisoj de la suda bufo (Anaxyrus terrestris) havas bildomodelon kiu celas miksiĝi kun la ĉirkaŭa kamufla haŭto.[50]
La aldistanca vidkapablo de rano estas pli bone ol ties deproksima vidkapablo. Alvokantaj ranoj tuj silentiĝos kima ili vidas entrudulon aŭ eĉ nur moviĝantan ombron, sed ju pli proksima esta objekto, malplej facile ĝi estas vidata.[50] Kiam rano pafas sian langon por kapti insekton ĝi estas reagante al malgranda moviĝanta objekton kiun ĝi ne povas vidi bone kaj ĝi devas bone lokigi antaŭfare precize ĉar iliaj okuloj fermiĝas kiam la lango estas etendita.[23] Ĉu rano vidas kolore estas debatata sed oni montris ke ĝi respondas pozitive al la blua lumo, eble ĉar tiu koloro estas asocia kun akvejoj kiuj povas havigi rifuĝon kiam la rano sentas sin minacita.[51]
Aŭdkapablo
Ranoj povas aŭdi kaj en aero kaj sub akvo. Ili ne havas eksterajn orelojn; la orel-membranoj estas rekte eksponitaj aŭ povas esti kovrataj per tavolo de haŭto kaj estas videblaj kiel cirkla areo ĝuste malantaŭ la okulo. La grando kaj distanco el la orelmembranoj rilatas al la frekvenco kaj ondolongo je kiu la rano alvokas. En kelkaj specioj kiaj la taŭrorano, la grando de la membrano indikas la sekson de la rano; maskloj havas membranojn kiuj estas pli grandaj ol ties okuloj dum ĉe inoj, la okuloj kaj membranoj estas multe samgrandaj.[52] Bruo kaŭzas ke la membrano vibras kaj la sono estas transigita al la meza kaj interna orelo. La meza orelo enhavas duoncirklajn kanalojn kiuj helpas kontroli ekvilibron kaj orientadon. En la interna orelo, la aŭdoharaj ĉeloj estas aranĝitaj en du areoj de la koĥleo, nome la bazaj papiloj kaj la amfibiaj papiloj. La unuaj detektas altajn frekvencojn kaj la lasta malaltajn frekvencojn.[53] Ĉar la koĥleo estas mallonga, ranoj uzas elektran meĥanismon por etendi siajn kampojn de aŭdeblajn frekvencojn kaj helpi diskriminacii diferencajn sonojn.[54] Tiu aranĝo ebligas la detektadon de teritoriaj kaj reproduktaj alvokoj de la samspecianoj. En kelkaj specioj kiuj loĝas en aridaj regionoj, la sono de ŝtormo aŭ forta pluvo povas eligi ilin el dormanta stato.[53] Rano povas esti alarmita pro neatendita bruo sed ĝi kutime faras nenian agon ĝis ĝi lokigis la fonton de la sono pervide.[52]
Alvoko
.jpg.webp)
La alvoko aŭ kvako de rano estas unika de ĝia specio. Ranoj kreas tiun sonon per pasado de aero tra la laringo en la gorĝon. En plej el la ranalvokoj, la sono estas amplifita de unu aŭ pliaj voĉosakoj, membranoj de haŭto sub la gorĝo aŭ ĉe la angulo de la buŝo, kiuj etendas dum la amplifado de la alvoko. Kelkaj ranalvokoj estas tiom laŭtaj ke ili povas esti aŭdataj ĝis el unu mejlon for.[55]
Ranoj de la genroj Heleioporus kaj Neobatrachus ne havas voĉosakojn sed povas tamen produkti laŭtan alvokon. Ties buŝa kavaĵo estas pligrandigita kaj kupolforma, kaj funkcias kiel resonejo kiu amplifas la sonon. Specioj de ranoj kiuj ne havas voĉosakojn kaj kiuj ne havas laŭtan alvokon tendencas loĝi en areoj ĉe konstanta bruo, fluanta akvo. Ili bezonas uzi alternativajn rimedojn por komuniki. La marborda vostorano (Ascaphus truei) loĝas en montorojoj de Nordameriko kaj ne havas voĉon.[56]
La ĉefa tialo por alvoko estas ebligi ke la masklaj ranoj allogu partneron. Maskloj povas alvoki individue aŭ povas esti ĥoro de sonoj kie nombraj maskloj koincidas en reproduktejoj. Inoj de multaj ranospecioj, kiaj la ordinara arborano (Polypedates leucomystax), respondas al la masklalvokoj, kio funkcias por plifortigi la reproduktan aktivecon en reprodukta kolonio.[57] Raninoj preferas masklojn kiuj produktas sonojn de pli granda intenseco kaj pli malalta frekvenco, atributoj kiuj elstaras el ranaro. La tialo por tio ŝajne estas montri la propran forton, la masklo montras sian bonfarton por produkti pli bonkvalitajn idarojn.[58]
Diferenca alvoko estas elsendata de masklo aŭ nericevema ino kiam ŝi estas rajdata de masklo. Tiu estas distinga ĉirpada sono kaj estas akompanata de vibrado de la korpo.[59] Arboranoj kaj kelkaj ne-akvaj specioj havas pluvalvokon kiun ili faras surbaze de humideco antaŭ pluvo.[59] Multaj specioj havas ankaŭ teritorian alvokon kiu estas uzata por forigi aliajn masklojn. Ĉiuj el tiuj alvokoj estas elsendataj havante fermitan buŝon.[59] Danĝeralvoko, elsendata de kelkaj ranoj kiam ili estas en danĝero, estas produktita havante malfermitan buŝon rezulte en pli altatona alvoko. Ĝi estas tipe uzata kiam la rano estas kaptata de predanto kaj povas utili por forigi aŭ disorientigi la atakanton tiele ke ĝi liberigas la ranon.[59]
Multaj specioj de ranoj havas profundajn alvokojn. La kveko de la amerika Taŭra rano (Rana catesbiana) estas foje srkibita anglalingve kiel "jug o' rum".[60] La Pacifika arborano (Pseudacris regilla) produktas la onomatopean "ribbit" ofte aŭdata en filmoj.[61] Aliaj literumadoj de ranalvokoj estas "brekekekex koax koax", nome alvoko de Ridorano (Pelophylax ridibundus) en La ranoj, de antikva greka komedia dramo de Aristofano.[62]
Torporo

Dum tre malfacilaj vivkondiĉoj, kelkaj ranoj eniras al stato de torporo kaj restas neaktivaj dum monatoj. En pli malvarmaj regionoj, multaj specioj de ranoj vintrodormas vintre. Tiuj kiuj loĝas surtere kiel la Amerika bufo (Bufo americanus) elfosas truon kaj faras vintrumejon en kiu kuŝas dormetanta. Alia, malpli efika elfosanto, trovas fendon aŭ enterigas sin en sekaj folioj. Akvaj specioj kiaj la Taŭra rano (Rana catesbeiana) normale sinkas en la fundo de flako kie ili kuŝas, duonenmetitaj en koto sed ankoraŭ kapablas aliri al la oksigeno dissolvita en akvo. Ties metabolo malrapidiĝas kaj ili vivas el siaj energirezervoj. Kelkaj ranoj povas eĉ survivi frostigitaj. Glacikristaloj formiĝas sub la haŭto kaj en la korpokavaĵo sed la esencaj organoj estas protektataj el frostado per alta koncentrado de glukozo. Ŝajne senviva, frostigitarano povas re-ekspiradi kaj la koro povas restarti kiam la kondiĉoj revarmiĝas.[63]
Aliflanke, la stria truorano (Cyclorana alboguttata) regule torpore somerumas dum la varma, seka sezono en Aŭstralio, survivante en dormanta stato sen aliro al manĝo aŭ akvo dum naŭ aŭ dek monatoj de la jaro. Ĝi nestotruas subgrunde kaj buliĝas ene de protekta kokono formata de sia eligita haŭtaĵo. Esploristoj de la Universitato de Kvinslando troviĝis ke dum tiu torpora somerumado, la metabolo de la rano estas ŝanĝita kaj la operacia efikeco de la mitokondrio pliiĝas. Tio signifas ke la limigita kvanto de energio disponebla por la komateca rano estas uzata en pli efika maniero. Tiu surviva meĥanismo estas utila nur por animaloj kiuj restas komplete nekonsciaj dum etenda tempoperiodo kaj kies energiaj postuloj estas malaltaj ĉar ili estas malvarmsangaj kaj ne bezonas generi varmon.[64] Alia esplorado montris ke, por havigi tiujn energiajn postulojn, muskoloj atrofias, sed la muskoloj de la antaŭaj kruroj prefere ne estas tuŝitaj de tio.[65] Oni trovis ranojn kiuj havas suprajn kritikajn temperaturojn de ĉirkaŭ 41 gradojn Celsius.[66]
Movmanieroj
Vivohistorio

La plimulto de anuroj serĉas akvon por tie demeti sian frajon kaj la larvoj vivas enakve ĝis sia metamorfozo. La subfamilio Brachycephaloidea demetas sur tero ovojn, ene de kiuj la larvoj evoluas rekte en ranetojn, kaj almenaŭ unu specio (Limnonectes larvaepartus) estas vivnaska.
Defendado
Distribuado kaj konservostatuso
Homa uzado
En homa kulturo
En Esperanto aperis
- Frank van Hertrooij, Verdaj Akrobatoj ĉe la blogo Scivolemo, 31-a de januaro 2021
Vidu ankaŭ
Bildaro




Referencoj
- ↑ anuroj en vortaro.net
- ↑ Stuart, S. N.; Chanson, J. S.; Cox, N. A.; Young, B. E.; Rodrigues, A. S. L.; Fischman, D. L.; Waller, R. W. (2004). "Status and trends of amphibian declines and extinctions worldwide". Science 306 (5702): 1783–1786. doi:10.1126/science.1103538. PMID 15486254.
- 1 2 3 Pough, F. H.. (2003) Herpetology: Third Edition. Benjamin Cummings. ISBN 0-13-100849-8.
- ↑ Cannatella, David C. (1997). "Salientia". Tree of Life Web Project. Alirita la 9an de Junio 2016.
- ↑ Badger, D.. (1995) Frogs. Airlife Publishing. ISBN 1-85310-740-9.
- ↑ Kuzmin, Sergius L. (1999-09-29). Bombina bombina. AmphibiaWeb. University of California, Berkeley. Alirita 2012-06-15.
- ↑ Atelopus zeteki. IUCN Red List of Threatened Species. Version 2012.1 (2010). Alirita 2012-08-02.
- ↑ Duellman, William E. Biology of Amphibians el Google Books
- 1 2 Cannatella, David (2008-01-11). Anura. Tree of Life web project. Arkivita el la originalo je 2012-10-19. Alirita 2012-08-08.
- ↑ (1993) “The major clades of frogs”, Herpetological Monographs 7, p. 94–117. doi:10.2307/1466954.
- ↑ (2005) “Systematic review of the frog family Hylidae, with special reference to Hylinae: Phylogenetic analysis and revision”, Bulletin of the American Museum of Natural History 294, p. 1–240. DOI 10.1206/0003-0090(2005)294[0001:SROTFF]2.0.CO;2
- ↑ Kuzmin, S. L. (1999-11-10). Pelophylax esculentus. Alirita 2012-10-12.
- ↑ Köhler, S. (2003). Mechanisms for partial reproductive isolation in a Bombina hybrid zone in Romania. Dissertation for thesis. Alirita 2012-06-05.
- 1 2 Flam, F. (1995) Finding earliest true frog will help paleontologists understand how frog evolved its jumping ability. Knight Ridder/Tribune News Service via HighBeam Research. Arkivita el la originalo je 2013-05-13. Alirita 2012-06-10. Arkivita kopio. Arkivita el la originalo je 2013-05-13. Alirita 2017-03-08.
- 1 2 3 Burton, Maurice. (1972) The Observer's Book of British Wild Animals. Frederick Warne & Co, p. 204–209. ISBN 0-7232-1503-0.
- ↑ (2004) “The impact of insecticides and herbicides on the biodiversity and productivity of aquatic communities”, Ecological Applications 15 (2), p. 618–627. doi:10.1890/03-5342.
- ↑ (2005) “The lethal impact of Roundup on aquatic and terrestrial amphibians”, Ecological Applications 15 (4), p. 1118–1124. doi:10.1890/04-1291.
- ↑ (2002) “Hermaphroditic, demasculinized frogs after exposure to the herbicide atrazine at low ecologically relevant doses”, Proceedings of the National Academy of Sciences of the United States of America 99 (8), p. 5476–5480. doi:10.1073/pnas.082121499.
- ↑ (2007) “Aquatic eutrophication promotes pathogenic infection in amphibians”, Proceedings of the National Academy of Sciences of the United States of America 104 (40), p. 15781–15786. doi:10.1073/pnas.0707763104.
- ↑ "Tiny frog claimed as world's smallest vertebrate", 2012-01-12. Kontrolita 2012-09-28.
- ↑ (2004-03-15) “Contribution of eye retraction to swallowing performance in the northern leopard frog, Rana pipiens”, Journal of Experimental Biology 207 (Pt 8), p. 1361–1368. doi:10.1242/jeb.00885.
- ↑ Minott, Kevin (2010-05-15). "How frogs jump". National Geographic. [www.komverse.com/2010/05/15/how-frogs-jump-nationalgeographic/] Alirita la 17an de Marto 2017.
- 1 2 3 4 5 Tesler, P. (1999). The amazing adaptable frog. Exploratorium:: The museum of science, art and human perception. Alirita 2012-06-04.
- ↑ Vincent, L. (2001) Litoria caerulea. James Cook University. Arkivita el la originalo je 2004-04-22. Alirita 2012-08-03. Arkivigite je 2005-07-24 per la retarkivo Wayback Machine Arkivita kopio. Arkivita el la originalo je 2004-04-22. Alirita 2017-03-09.
- ↑ (1980) “Toe pad morphology and mechanisms of sticking in frogs”, Biological Journal of the Linnean Society 13 (3), p. 199–216. doi:10.1111/j.1095-8312.1980.tb00082.x.
- ↑ (2002) “New and poorly known parachuting frogs (Rhacophoridae : Rhacophorus) from Sumatra and Java”, Herpetological Monographs 16, p. 46–92. doi=10.1655/0733-1347(2002)016[0046:NAPKPF]2.0.CO;2
- ↑ Couch's spadefoot (Scaphiopus couchi). Arizona-Sonora Desert Museum. Alirita 2012-08-03.
- ↑ Walker, M., "Legless frogs mystery solved", BBC News, 2009-06-25.
- ↑ Stebbins, Robert C.. (1995) A Natural History of Amphibians. Princeton University Press, p. 10–14. ISBN 0-691-03281-5.
- ↑ (1932) “Notes on feeding and molting in frogs”, The American Naturalist 66 (707), p. 530–540. doi:10.1086/280458.
- 1 2 Badger, D.. (1995) Frogs. Airlife Publishing. ISBN 1-85310-740-9.
- ↑ Smyth, H. R.. (1962) Amphibians and Their Ways. Macmillan. ISBN 0-02-612190-5.
- ↑ Dickerson, M. C.. (1969) The Frog Book: North American Frogs and Toads. Dover Publications. ISBN 978-0-486-21973-8.
- ↑ Blackburn, D. C. (2002-11-14). Trichobatrachus robustus. AmphibiaWeb. Alirita 2012-08-18.
- ↑ (1985) “Osteoderms in Anurans”, Journal of Herpetology 18 (3), p. 313–328. doi:10.2307/1564085.
- ↑ Vitt, Laurie J.. (2013) Herpetology: An Introductory Biology of Amphibians and Reptiles. Academic Press. ISBN 9780123869203.
- ↑ (2005) “Head co-ossification, phragmosis and defence in the casque-headed tree frog Corythomantis greeningi”, Journal of Zoology 265 (1), p. 1–8. doi:10.1017/S0952836904005953.
- ↑ (2003) “Fixed green and brown color morphs and a novel color‐changing morph of the Pacific tree frog Hyla regilla”, The American Naturalist 162 (4), p. 461–473. doi:10.1086/378253.
- ↑ Kimball, John (2010) Vertebrate Lungs: Frog Lungs. Kimball's Biology Pages. Arkivita el la originalo je 2012-08-19. Alirita 2012-07-09. Arkivita kopio. Arkivita el la originalo je 2012-08-19. Alirita 2017-03-10.
- ↑ Boisvert, Adam (2007-10-23). Barbourula kalimantanensis. AmphibiaWeb. University of California, Berkeley. Alirita 2012-07-09.
- 1 2 Kimball, John (2010). Animal Circulatory Systems: Three Chambers: The Frog and Lizard. Kimball's Biology Pages. Alirita 2012-07-09.
- ↑ Lee, Deborah (2010-04-23). Telmatobius culeus. AmphibiaWeb. University of California, Berkeley. Alirita 2012-07-09.
- ↑ Frog Digestive System. TutorVista.com (2010). Arkivita el la originalo je 2013-02-04. Alirita 2012-08-04. Arkivita kopio. Arkivita el la originalo je 2013-02-04. Alirita 2017-03-10.
- 1 2 Dorit, R. L.. (1991) Zoology. Saunders College Publishing. ISBN 0-03-030504-7.
- 1 2 3 4 5 6 Frog's internal systems. TutorVista.com (2010). Arkivita el la originalo je 2012-04-22. Alirita 2012-06-04. Arkivita kopio. Arkivita el la originalo je 2012-04-22. Alirita 2017-03-10.
- ↑ Dorota (2006). Life cycle of a frog. Frogland. Alirita 2012-08-04.
- ↑ Hormones, sex accessory structures, and secondary sexual characteristics in amphibians. Hormones and Reproduction of Vertebrates – Vol 2: Amphibians, p. 83–98. Arkivita el la originalo je 2013-05-14. Alirita 2012-08-04. Arkivita kopio. Arkivita el la originalo je 2013-05-14. Alirita 2017-03-11.
- ↑ Amniota. Tree of Life Web Project (2012). Alirita 2012-08-04. Arkivita kopio. Arkivita el la originalo je 2012-02-04. Alirita 2017-03-11.
- 1 2 Howard, Ian, P.. (1995) Binocular Vision and Stereopsis. Oxford University Press. ISBN 0195084764.
- 1 2 3 Badger, David. (1995) Frogs. Airlife Publishing Ltd, p. 31–35. ISBN 1-85310-740-9.
- ↑ Muntz, W. R. A.. (1964) Vision in frogs. W. H. Freeman.
- 1 2 Badger, David. (1995) Frogs. Airlife Publishing. ISBN 1-85310-740-9.
- 1 2 Stebbins, Robert C.. (1995) A Natural History of Amphibians. Princeton University Press, p. 67–69. ISBN 0-691-03281-5.
- ↑ (1998) “Electrical properties of frog saccular hair cells: distortion by enzymatic dissociation”, Journal of Neuroscience 18 (8), p. 2962–2973.
- ↑ Bullfrog. Ohio Department of Natural Resources. Arkivita el la originalo je 2012-08-18. Alirita 2012-06-19. Arkivita kopio. Arkivita el la originalo je 2012-08-18. Alirita 2017-03-14.
- ↑ Nafis, Gary (2012). Ascaphus truei: Coastal Tailed Frog. California Herps. Alirita 2012-06-19.
- ↑ (1997) “Communication signals and sexual selection in amphibians”, Current Science (PDF) 72, p. 923–927.
- ↑ Gerhardt, H. C. (1994). “The evolution of vocalization in frogs and toads”, Annual Review of Ecology and Systematics 25, p. 293–324. doi:10.1146/annurev.es.25.110194.001453.
- 1 2 3 4 Badger, David. (1995) Frogs. Airlife Publishing Ltd, p. 39–44. ISBN 1-85310-740-9.
- ↑ Hilton, Bill Jr.. (1986) Jug-o-Rum: Call of the Amorous Bullfrog 1. Hilton Pond Center for Piedmont Natural History.
- ↑ . The RRRRRRRRiveting Life of Tree Frogs (Februaro 2005). Arkivita el la originalo je 2012-03-09. Alirita 2012-08-04. Arkivita kopio. Arkivita el la originalo je 2012-03-09. Alirita 2017-03-14.
- ↑ Aristophanes The Frogs. Arkivita el la originalo je 2012-05-13. Alirita 2012-06-19. Arkivita kopio. Arkivita el la originalo je 2012-05-13. Alirita 2017-03-14.
- ↑ Emmer, Rick (1997-11-24). How do frogs survive winter? Why don't they freeze to death?. Scientific American. Alirita 2012-06-15.
- ↑ (2009) “Metabolic depression during aestivation in Cyclorana alboguttata”, Comparative Biochemistry and Physiology – Part A: Molecular & Integrative Physiology 154 (4), p. 557–563. doi:10.1016/j.cbpa.2009.09.001.
- ↑ (2005) “Lessons from an estivating frog: sparing muscle protein despite starvation and disuse”, AJP: Regulatory, Integrative and Comparative Physiology 290 (3), p. R836–R843. doi:10.1152/ajpregu.00380.2005.
- ↑ Wilmer, Pat. (2009) Environmental Physiology of Animals. Wiley, p. 188. ISBN 9781405107242.
Eksteraj ligiloj
- http://www-itg.lbl.gov/ITG.hm.pg.docs/Whole.Frog/Whole.Frog.html
- http://raysweb.net/specialplaces/pages/frogsdecline.html Malapero de ranoj
- http://www.thelilypad.org Arkivigite je 2022-02-14 per la retarkivo Wayback Machine
- En tiu ĉi artikolo estas uzita traduko de teksto el la artikolo Frog en la angla Vikipedio.