Zahntroste
Die Zahntroste (Odontites) sind eine Pflanzengattung innerhalb der Familie der Sommerwurzgewächse (Orobanchaceae). Alle Zahntroste leben halbparasitisch, indem sie mit Saugorganen (Haustorien) in die Wurzeln benachbarter Pflanzenexemplare einwachsen und über diese Verbindung von ihnen Wasser und Nährstoffe erhalten. Das Hauptverbreitungsgebiet der etwa 26 Arten liegt im westlichen Mittelmeerraum, viele Arten kommen als Endemiten in einem sehr begrenzten Gebiet vor, nur drei Arten besiedeln ein größeres Gebiet.
| Zahntroste | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Gelber Zahntrost (Odontites luteus) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Odontites | ||||||||||||
| Ludw. |
Beschreibung
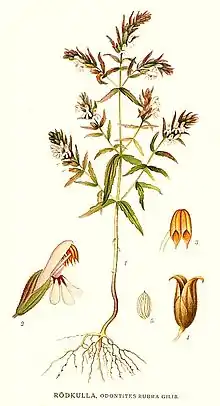

Vegetative Merkmale
Zahntrost-Arten sind vorwiegend einjährige,[1] krautige Pflanzen,[2] wenige Arten sind kurzlebig ausdauernd und verholzend. Über Wurzelhaustorien können sie benachbarte Pflanzen parasitieren. Die meisten Vertreter wachsen aufrecht, nur wenige Arten wie Odontites corsicus, Odontites maroccanus und Odontites violaceus bilden auf dem Boden aufliegende Triebe, an denen nur die Blütenstände aufrecht wachsen.
Die gegenständig[1] am Stängel verteilt angeordneten Laubblätter sind sitzend. In der Knospe sind die Hälften der Blattspreite eingerollt (revolut). Die Blattspreiten sind ungeteilt. Da viele Arten die Blätter nur teilweise entrollen, ist der Blattrand oftmals nach unten gebogen. Etwa zwei Drittel der Arten besitzen schmal-lanzettliche bis linealische Blattspreiten, die durch die umgebogenen Blattränder oftmals heidekrautartig wirken. Die restlichen Arten haben breitere Blattspreiten, die lanzettlich bis oval-lanzettlich sind. Der Blattrand ist bei den meisten Arten ganzrandig oder nur mit ein bis drei kleinen Zähnen besetzt, nur die Arten der Odontites vernus-Gruppe, sowie Odontites cebennensis und Odontites lanceolatus besitzen Blätter mit vier bis acht deutlich ausgeprägten Zähnen.
Zahntrost-Arten sind sowohl mit einfachen als auch mit drüsigen Trichomen behaart. Die einfachen Trichome sind einzellig und mehr oder weniger starrborstig; je nach Art variieren Länge, Anordnung und Dichte. Die drüsigen Trichome treten in verschiedenen Formen auf. Neben den bei allen Arten auftretenden und taxonomisch unbedeutenden Trichomen des schildförmigen Drüsen- beziehungsweise Kurzdrüsen-Typs sind die verschiedenen Formen der Stieldrüsen-Trichome (langgestielte Köpfchentrichome) ein wichtiges Merkmal zur taxonomischen Einteilung der Gattung. So können diese fehlen, wenigzellig und breit elliptisch (lanceolatus-Typ), vielzellig und kugelig-elliptisch (pyrenaeus-Typ) oder vielzellig und kegelförmig (viscosus-Typ) sein. Die Länge der Stieldrüsen liegt meist zwischen 150 und 700 μm, selten sind sie auch nur 50 µm lang. Die schildförmigen Drüsen sind flach gebaut aus einem einzelligen Stiel, der ins Epidermisgewebe versenkt ist, und einem flach gewölbten Schild aus schmalen, parallel angeordneten Zellen. Die Kurzdrüsen sind 20 bis 90 µm lang, ihr Köpfchen besteht aus ein bis vier Zellen.
Das Wurzelsystem besteht nur aus wenigen, sehr großen Hauptwurzeln, von denen ein schwach ausgeprägtes Netz aus feineren Wurzeln ausgeht. Das Xylem der Wurzeln ist sehr gut ausgebildet, im Gegensatz zum Phloem. Wurzelhaare finden sich meist nur an jungen Sämlingen sowie in der unmittelbaren Umgebung der Haustorien.[3]
Die mehrjährigen Arten Odontites squarrosus, Odontites bocconii und Odontites linkii werden meist nur etwa vier Jahre alt. Durch jährlich an der Basis austreibende, teilweise sterile Seitentriebe entsteht eine strauchige Wuchsform. Das Holz dieser Arten ist im Querschnitt zerstreut- bis halbringporig, wobei die Poren in lockeren, radialen Reihen angeordnet sind. Durch die Bildung mehrreihiger, nur wenige Poren aufweisende Lagen dickwandigerer Spätholzzellen sind Jahresringe auszumachen. Die Holzstrahlen bestehen aus radial kurzen Zellen, die in vielen Reihen langgestreckt und in wenigen Reihen quadratisch geformt sind. Die Holzstrahlen sind meist von drei bis fünf Reihen Scheidezellen umgeben.
Blütenstände und Blüten

In den vielblütigen, stark verzweigten, traubigen, Blütenständen enden die Hauptachsen bei den Zahntrost-Arten nicht in einer Blüte (Polytelie). Je nach Art sind die Blühzonen der Blütenstände kurz, kompakt und dichtblütig oder bereits zur Blütezeit locker und gestreckt. Oftmals verlängert sich während der Fruchtreifung der Blütenstand erheblich, so dass er aus langgestreckten, starren und leicht verholzten Achsen besteht. Die Blütenstände stehen sowohl an den Haupt- als auch an den Seitensprossen, wobei der Hauptblütenstand mit einer basalen, bis zu 22 Knoten umfassenden Hemmzone beginnt. In dieser Hemmzone stehen die sogenannten Interkalarblätter (laubblattartige Deckblätter stark gehemmter, nicht aufblühender Blüten). Die Deckblätter werden zur Spitze des Blütenstands hin kleiner und sind stets kürzer als die Laubblätter; die Übergangszone zwischen Laub- und Deckblättern ist je nach Art unterschiedlich stark ausgeprägt. Die Blüten stehen meist paarweise in den Knoten des Blütenstandes. Sie blühen bei den meisten Arten der Reihe nach von unten nach oben auf, bei einigen Arten konnte jedoch die umgekehrte Reihenfolge festgestellt werden.
Die Blüten sind zygomorph und innerhalb der Gattung sehr formenreich: die Organe treten oftmals in vielfältig abgewandelten Formen auf. Die Länge der Blüten variiert zwischen 6 und 12 Millimetern. Der Kelch besteht aus vier verwachsenen Kelchblättern, die zwei Lippen mit insgesamt vier ganzrandigen, breit- bis schmaldreieckigen Zipfeln bilden. Der Kelch ist häufig mit Stieldrüsen besetzt, die sich oft, wie auch der gesamte Kelch, nach der Blütezeit vergrößern. Die aus fünf verwachsenen Kronblättern bestehende Krone ist gelb oder rötlich-purpurfarben, wobei von beiden Farbgruppen unterschiedliche Farbtöne und Helligkeiten auftreten. Die Krone besteht aus einer mehr oder weniger gebogenen Kronröhre sowie einem zweilippigen Kronsaum. Die aus zwei Kronblattzipfeln bestehende Oberlippe bildet einen flachen Helm. Die ausgebreitete Unterlippe ist dreizipfelig und die seitlichen Kronlappen sind ganzrandig, der mittlere Kronlappen ist ausgerandet[2].
Die vier Staubblätter treten in zwei unterschiedlichen Formen auf, wobei die beiden unteren länger sind als die beiden oberen. Sie werden entweder vom Oberlippenhelm verdeckt oder stehen mehr oder weniger weit über die Krone hinaus. Bei allen Arten außer der Odontites luteus-Gruppe und Odontites corsicus neigen sich die Staubblätter zusammen, so dass die Staubbeutel an den oberen stumpfen Enden durch schraubig gedrehte Haare miteinander verfilzt sind. Die unteren Enden sind grannig zugespitzt. Die Pollenkörner sind bei fast allen Arten einheitlich gebaut: im Äquatorialquerschnitt sind sie fast dreieckig, die Pollenkornwand (Exine) ist zwischen den Keimspalten (Mesocolpi) stark verdünnt. An den Polen sowie an den Rändern der Colpen ist das Retikulum weitmaschiger als an den Mesocolpen. Nur Odontites hollianus, Odontites corsicus und Odontites rigidifolius weichen mehr oder weniger stark von diesem Aufbau ab. Der zweiblättrige Fruchtknoten enthält vier bis vierzig Samenanlagen, die schräg von der Plazenta herabhängen. Die Narbe ist köpfchenförmig.
Früchte und Samen
Die Früchte der Zahntrost-Arten sind in der oberen Hälfte oder im oberen Drittel steifhaarige Kapselfrüchte. Sie öffnen sich fachspaltig, indem die Fruchtblätter entlang der Mittelrippe von oben nach unten aufreißen. Die größten Kapselfrüchte hat Odontites hollianus mit etwa 9,1 Millimeter Länge, die kürzesten Odontites viscosus und Odontites maroccanus mit jeweils etwa 3,5 Millimeter Länge. Die Früchte enthalten viele Samen.[2] Die Samen sind spindelförmig, 1,2 bis 2,8 Millimeter lang und gerippt. Die Flügel der Samen sind quer-gerillt.[2]
Ökologie
Entwicklung
Die Samen keimen im Frühjahr nach einer obligatorischen Winterruhe (Kälteinduktion). Die Keimung erfolgt epigäisch. Die kleinen Keimblätter sind bis etwa 1,5 Millimeter lang; sie sind elliptisch und ganzrandig. In den Blattachseln am Hauptspross bilden sich im Laufe der Entwicklung Seitenäste, die sich noch ein weiteres Mal verzweigen können. Wie der Hauptspross bilden auch sie traubige Blütenstände. Vor oder spätestens zur Blütezeit werfen die meisten Arten der Gattung ihre Laubblätter ab, die Tragblätter werden meist später abgeworfen, so dass die Früchte an den völlig blattlosen Sprossen stehen.
Parasitismus
Berührt eine Zahntrost-Wurzel die Wurzel einer anderen Pflanze, so versucht sie über einfache Kontaktorgane, die Haustorien, eine Verbindung zu dieser Wurzel aufzubauen. Nur selten werden auch andere Pflanzenteile wie beispielsweise oberirdische Sprossachsen oder Rhizome befallen[4]. Bei erfolgreichem Eindringen der Haustorien in den Wirt wird eine Verbindung zwischen dem Xylem beider Pflanzen hergestellt. Über diese Verbindung entziehen die Zahntroste ihren Wirten Wasser sowie darin gelöste organische Verbindungen und anorganische Nährionen. Voraussetzung für den Nährstofftransport in Richtung Zahntrost ist eine hohe Wasserabgabe durch die Blätter. Dies wird durch die Drüsen, die bei fast allen Zahntrost-Arten zahlreich auftreten, durch Guttation und vor allem tagsüber auch durch Transpiration erreicht[3]. Nur bei Arten wie dem Gelben Zahntrost (Odontites luteus), die an besonders sonnenreichen Standorten wachsen, sind weniger Drüsen vorhanden.
Innerhalb der Sommerwurzgewächse wird der Parasitismus der Zahntroste als eine sehr einfache Form angesehen. Die Wurzelsysteme sind nur wenig umgebildet, die Haustorien sind als einfach aufgebaute Sekundärhaustorien (die also noch nicht zu Beginn der Sämlingsentwicklung entstehen) ausgeprägt. Zahntroste sind nicht wirtsspezifisch, das heißt, sie befallen nahezu alle Pflanzen in ihrer Nähe. Es kommt dabei häufig vor, dass sie auch Pflanzen der eigenen Art befallen. Die Wirtspflanzen werden durch den Befall offensichtlich nicht beeinträchtigt.[5] Da die Pflanzen eigenes Chlorophyll besitzen, können sie sich auch ohne Wirt (autotroph) entwickeln (Fakultativer Parasitismus). Vitalität, Grad der Verzweigung und auch die Fertilität sind jedoch stark von den parasitierten Wirten abhängig; rein autotroph wachsende Pflanzen sind deutlich schwächer und können oftmals keine Samen ausbilden. Besonders Gräser sind als Wirtspflanzen geeignet, da sie durch ihre zahlreichen Faserwurzeln für die Haustorien gute Angriffsmöglichkeiten bieten.
Bestäubung
Innerhalb der Gattung lassen sich zwei Typen von Bestäubungsmechanismen unterscheiden. Beim homogamen und zwingend autogamen (selbstbefruchtenden) Typ, der beispielsweise bei Odontites powellii, Odontites corsicus und Odontites vulcanicus auftritt, findet bereits in der Knospe eine Selbstbefruchtung statt (Kleistogamie). Die Staubbeutel springen schon vor dem Aufblühen auf; der dann noch zurückgekrümmte Griffel liegt so, dass sich die Narbe genau zwischen den Staubbeuteln befindet und somit bestäubt wird. In der geöffneten Blüte ragt der Griffel meist nicht aus der Blüte heraus, sondern ist im Helm verborgen.
Der protogyne (vorweibliche) Typ fördert meist die Fremdbestäubung: Der empfangsbereite Griffel steht schon aus der noch nicht geöffneten Knospe heraus, während die Staubblätter im Inneren noch geschlossen sind. Diese springen frühestens zwei Tage später auf, wenn die Blüte bereits aufgeblüht ist. Von diesem Bestäubungstyp existieren verschiedene Varianten, von denen einige die nachträgliche Selbstbestäubung ermöglichen (schwache Protogynie), andere diese jedoch komplett verhindern (starke Protogynie). Je nach Aufblühreihenfolge wird eine Befruchtung durch Pollen des gleichen Blütenstandes (Geitonogamie, bei Aufblühfolge von unten nach oben) oder durch Pollen von anderen Blütenständen oder Individuen (Xenogamie, bei Aufblühfolge von oben nach unten) gefördert.
Meist sind Bienen und Hummeln (Apiformes) die Bestäuber der protogynen Blüten (Melittophilie). Je nach Größe der Blüten unterscheiden sich die vorwiegenden Bestäuber: An den kleineren Blüten bis etwa 7 Millimetern Länge sind hauptsächlich Honigbienen (Apis) zu beobachten, während die größeren Blüten mit über 9 mm Länge fast ausschließlich von Hummeln (Bombus) besucht werden. Die Zahntrost-Sägehornbiene hat sich auf den überwiegenden Besuch von Sommerwurzgewächsen spezialisiert (Oligolektie), neben den Zahntrosten sammelt sie gelegentlich auch an Augentrosten (Euphrasia) und Wachtelweizen (Melampyrum) Nektar.[6]
Ausbreitung
Die Samen der Zahntroste sind nicht an eine besondere Form der Ausbreitung angepasst und werden daher meist nur in unmittelbarer Umgebung der Pflanze ausgestreut. Obwohl sie sehr klein sind, ist das Gewicht der Samen für eine Windausbreitung (Anemochorie) zu hoch. Auch eine Verschleppung durch Tiere (Epizoochorie) ist sehr unwahrscheinlich, da die Samen nicht geflügelt sind, wie beispielsweise die der Bartsia, und so nicht an glatten und feuchten Oberflächen kleben bleiben. Nur für wenige Arten kommt eine begrenzte Verbreitung durch Mitschleppen in Heu in Frage; hierzu zählen vor allem die verschiedenen Sippen der Odontites vernus-Gruppe.
Fraßfeinde
Die Raupen der Spannerart Perizoma bifaciata ernähren sich oligophag (nur wenige Nahrungspflanzen akzeptierend) unter anderem von den unreifen Samen des Gelben Zahntrostes (Odentites luteus) und des Frühlings-Zahntrostes (Odontites vernus).[7] Zudem wurden die Raupen der Gammaeule (Autographa gamma) aus der Familie der Eulenfalter (Noctuidae) am Frühlings-Zahntrost (Odontites vernus) gefunden.[8]
Verbreitung und Standorte

Die meisten Arten der Gattung sind im westlichen Mittelmeerraum heimisch. Zwei Schwerpunktgebiete mit jeweils sechs vorkommenden Arten liegen in Nordwestafrika, eines im mittleren Atlasgebirge in Marokko, ein weiteres in Ostalgerien und Tunesien. Ein weiterer Schwerpunkt mit insgesamt fünf Arten liegt im Nordosten Spaniens, vier der dort vorkommenden Arten sind endemisch. Dieser Lokalendemismus ist innerhalb der Gattung weit verbreitet, viele Arten sind nur von kleinen Beständen in Gebirgen oder auf Inseln bekannt, etwa ein Drittel kommt nur noch in Restpopulationen vor und ist vom Aussterben bedroht. Nur drei Arten haben ein größeres Verbreitungsgebiet: Odontites viscosus mit seinem Verbreitungsschwerpunkt am Südhang der Pyrenäen reicht im Norden bis an die West- und Zentralalpen; der Gelbe Zahntrost (Odontites luteus) besiedelt in Mitteleuropa auch die sommertrockensten Gebiete und reicht vom Nordosten Spaniens bis zur Wolgaplatte und verstreut bis zum Kaukasus, zur Krim und dem östlichen Taurusgebirge; der Frühlings-Zahntrost (Odontites vernus) reicht nördlich bis nach Südskandinavien und über weite Teile der gemäßigten Gebiete Eurasiens bis nach Ostasien.
Die Standorte liegen in Höhenlagen zwischen 0 und 2500 Metern. Die meisten Arten sind an trockene, oftmals felsig-steinige Trespen- und Steppenrasen oder lichte Stein-Eichen-, Flaum-Eichen- und Kiefernwälder angepasst. Im Gegensatz dazu kommt der Frühlings-Zahntrost (Odontites vernus) hauptsächlich in frischen bis nassen und nährstoffreichen Rasengesellschaften vor und kann sogar die Salzwiesen der Ost- und Nordseeküsten besiedeln. Fast alle Arten haben einen sehr hohen Lichtbedarf, nur wenige, beispielsweise Odontites viscosus, Odontites cebennensis und Odontites powellii, wachsen auch an halbschattigen Standorten.
Systematik
Äußere Systematik
Die Gattung Odontites gehört zur Tribus Rhinantheae innerhalb der Familie der Orobanchaceae. Die Verwandtschaftsbeziehungen innerhalb der Tribus Rhinantheae, sind noch nicht eindeutig geklärt und werden kontrovers diskutiert.
Die erste molekularbiologische Untersuchung, in die eine Zahntrost-Art einbezogen war, wurde erst 2006 von Jonathan Bennett und Sarah Mathews veröffentlicht.[9] Das folgende Kladogramm zeigt die auf Grundlage dieser Untersuchung vermuteten Verwandtschaftsverhältnisse (vereinfacht nach[9]). Dabei konnten Vertreter der Gattungen Bartsia und der Schuppenwurzen (Lathraea) zum Teil nicht eindeutig eingeordnet werden, die Gattungen Bartsia und Parentucellia ließen sich ebenfalls nicht eindeutig unterscheiden. Die gelegentlich zu Odontites gezählten Gattungen Macrosyringion, Odontitella, Bornmuellerantha und Bartsiella sowie die wahrscheinlich ebenfalls nahe verwandten Gattungen Hedbergia und Nothobartsia wurden von Bennet et al. 2006 noch nicht in molekulargenetische Untersuchungen einbezogen.[9]
| |||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
Die Verwandtschaftsbeziehungen etwas besser abklären konnten Scheunert et al. 2012, doch blieben noch viele Fragen offen.[10]
Innere Systematik
Die Gattung Odontites umfasst nach der Monographie von Markus Bolliger (1996) folgende 26 Arten. Einige phänetisch sehr ähnliche Arten wurden in dieser Arbeit zu Artengruppen zusammengefasst; andere zuvor als Art beschriebene Taxa wurden wiederum nur noch mit dem Status einer Unterart anerkannt.
 Habitus, gegenständige Laubblätter und Blüten von Odontites hollianus  Habitus von Odontites longiflorus (Syn.: Macrosyringion longiflorum) im Habitat
|

Taxonomie, Botanische Geschichte und Etymologie
Der Gattungsname Odontites geht auf Plinius den Älteren zurück. Er benennt eine Pflanzenart, die wahrscheinlich dieser Gattung angehört, mit dem Namen odontītis, was wahrscheinlich vom griechischen Wort όδούς (odous für Zahn; Mehrzahl: Odontes) abgeleitet ist. Zum Teil geht man davon aus, dass diese Namensgebung auf die angebliche Wirksamkeit dieser Pflanzenart gegen Zahnschmerzen zurückzuführen ist. Wahrscheinlicher ist aber, dass sich der Name auf die zahnförmig gezackten Ränder der Kelchblätter bezieht.[16] Andererseits ist es auch denkbar, dass die Analogie zwischen den Zähnen der Blätter und den Zähnen des Menschen dazu führte, dass der Pflanze eine entsprechende Wirksamkeit zugesprochen wurde.[17] Die Anwendung des bitter schmeckenden, als Herba Euphrasiae rubra bezeichneten Extraktes wurde jedoch schon Anfang des 19. Jahrhunderts als „lange außer Gebrauch“ bezeichnet.[18] Auch zur Linderung von Menstruationsbeschwerden soll diese Pflanzenart eingesetzt worden sein; dazu wurden zerdrückte Pflanzen in die Schuhe gelegt.[19][20]
Die Gattung Odontites wurde 1757 durch Christian Gottlieb Ludwig aufgestellt.[21] Typusart ist Odontites vulgaris Moench. Zwar veröffentlichte Carl von Linné bereits 1753 einige der heute zu den Zahntrosten gehörenden Arten, er ordnete sie jedoch den Augentrosten (Euphrasia) zu.[22] In der Folge wurden die Arten immer wieder entweder als eigenständige Gattung geführt, teilweise wurden einzelne Artengruppen als eigene Gattungen ausgegliedert, oder aber die Arten wurden wahlweise den Augentrosten oder den Bartsia zugerechnet. Grund hierfür ist die morphologische Eigenständigkeit vieler Zahntrost-Arten, die eine Platzierung zwischen den beiden großen Gattungen Euphrasia und Bartsia nahelegt. Auch für die Gliederung innerhalb der Gattung wurden verschiedene Konzepte vorgestellt, von denen jedoch keines als allgemein anerkannt gilt.
Synonyme für Odontites Ludw. sind: Odontites sect. Orthantha Benth., Dispermotheca Beauverd, Macrosyringion Rothm., Odontitella Rothm., Orthantha (Benth.) A.Kern., Orthanthella Rauschert.[11]
Eine erste Unterteilung der Gattung in die Sektionen Lasiopera, Orthantha und Euodontites stammt von George Bentham aus dem Jahr 1846.[23] Später weicht er jedoch selbst von diesem Konzept ab und ordnet 1876 die Arten der Gattung Bartsia zu, wobei er nur noch die Sektionen Orthantha und Euodontites verwendet.[24] 1888 gliedert Anton Kerner von Marilaun die Arten um Odontites luteus in die Gattung Orthantha aus. Dieses Konzept wurde 1891 auch von Richard Wettstein und später von dessen Schüler Josef Hoffmann übernommen. Hoffmann veröffentlichte ab 1897 unter dem Titel „Beitrag zur Kenntniss der Gattung Odontites“ eine mehrteilige Monographie über die ihm bekannten europäischen Arten der Gattung. Seine Unterteilung innerhalb der Gattung in fünf unbenannte Gruppen ist vor allem durch Merkmale der Blüten begründet.[25] Die Artengruppe um Odontites viscosus wurde 1911 von Gustave Beauverd als Gattung Dispermotheca ausgegliedert, die Orthantha erkennt er mit dem Status einer Untergattung von Odontites an. Zudem schlägt er vor, die Art Odontites aucheri in eine eigene Gattung oder eine eigene Sektion zu stellen, nimmt aber diese Umordnung selbst nicht vor. Die bisher letzte vorgenommene Einteilung der Arten innerhalb der Gattung stammt von Werner Rothmaler aus dem Jahr 1943, der die Zahntroste in die Sektionen Euodontites, Orthantha und Dispermotheca unterteilt. Gleichzeitig setzt er den Vorschlag Beauverds um und gliedert Odontites aucheri in die von ihm neu aufgestellte Gattung Bornmuellerantha ein. Zudem errichtet er die Gattungen Macrosyringion und Odontitella und gliedert damit jeweils zwei weitere Arten aus Odontites aus.[26] Erst 1996 erfolgt eine weitere monographische Behandlung der Gattung durch Markus Bolliger. Er verzichtet dabei auf eine Einteilung der Gattung in Sektionen, erkennt aber die von Rothmaler aufgestellten Gattungen an und gliedert Odontites rameauanus in eine weitere eigenständige und monotypische Gattung Bartsiella aus.
Unabhängig davon, welches Gattungskonzept genutzt wurde, wurden die Arten der heutigen Gattung Odontites traditionell der Familie der Braunwurzgewächse (Scrophulariaceae) zugerechnet. Raffaello Bellini stellte 1907 eine Unterteilung der Familie auf, in der er alle parasitischen von den nicht-parasitischen Gattungen trennte. Die parasitischen Gattungen, so auch Odontites, ordnete er der Unterfamilie Rhinanthoideae zu. Die natürliche Aufstellung dieser Unterfamilie wurde in der Folge zunächst vor allem durch morphologische, später auch durch molekularbiologische Untersuchungen bestätigt. Seit den frühen 1990er Jahren zeigte sich jedoch immer mehr, dass das klassische Familienkonzept der Braunwurzgewächse keine natürlichen Verwandtschaftsverhältnisse abbildete. Infolgedessen wurde zunächst 1999 von Nelson D. Young et al. vorgeschlagen, die Gattungen der bisherigen Unterfamilie Rhinathoideae der Familie der Sommerwurzgewächse (Orobanchaceae) zuzuordnen. Diese Einordnung wurde 2001 von Richard Olmstead et al. durchgeführt und 2003 auch in die Systematik der Bedecktsamer nach APG II übernommen.[27][28][29]
Quellen
Einzelnachweise
Die Informationen dieses Artikels entstammen zum größten Teil den unter Literatur angegebenen Quellen, darüber hinaus werden folgende Quellen zitiert:
- Datenblatt in Flora Vascular.
- Zhi-Yun Zhang, Nikolai N. Tzvelev: Odontites Ludwig, S. 96 - textgleich online wie gedrucktes Werk, In: Z. Y. Wu, P. H. Raven (Hrsg.): Flora of China, Band 18: Scrophulariaceae through Gesneriaceae. Science Press, Peking und Missouri Botanical Garden Press, St. Louis, 1998, ISBN 0-915279-55-X.
- R. N. Govier et al.: Hemiparasitic Nutrition in Angiosperms. II: Root Haustoria and Leaf Glands of Odontites verna (Bell.) Dum. and their Relevance to the Abstraction of Solutes from the Host. In: New Phytologist, Band 67, 1968. S. 963–972.
- Hans Christian Weber: Über Wirtspflanzen und Parasitismus einiger mitteleuropäischer Rhinanthoideae (Scrophulariaceae). In: Plant Systematics and Evolution, Band 125, 1976, S. 97–107. doi:10.1007/BF00986775
- Hans Christian Weber: Zur Evolution des Parasitismus bei den Scrophulariaceae und Orobanchaceae. In: Plant Systematics and Evolution, Band 136, 1980, S. 217–232. doi:10.1007/BF01004627
- Denis Michez, Connal Eardley: Monographic revision of the bee genus Melitta Kirby 1802 (Hymenoptera: Apoidea: Melittidae). In: Ann. soc. entomol. Fr. (n.s.), Band 43, Heft 4, 2007. S. 379–440. PDF; 6,6 MB
- Axel Hausmann: Sterrhinae. In A. Hausmann (Hrsg.): The Geometrid Moths of Europe 2. Apollo Books, Stenstrup 2004, ISBN 87-88757-37-4.
- Vergleiche HOSTS - a Database of the World's Lepidopteran Hostplants, Abfrage vom 27. September 2008.
- Jonathan R. Bennet, Sarah Mathews: Phylogeny of the parasitic plant family Orobanchaceae inferred from Phytochrome A., In: American Journal of Botany, Band 93, Nummer 7, 2006. S. 1039–1051. Abstract.
- Agnes Scheunert, Andreas Fleischmann, Catalina Olano-Marín, Christian Bräuchler, Günther Heubl: Phylogeny of tribe Rhinantheae (Orobanchaceae) with a focus on biogeography, cytology and re-examination of generic concepts. In: Taxon, Volume 61, Issue 6, 2012, S. 1269–1285. JSTOR:24389112
- Karol Marhold, 2011: Scrophulariaceae: Datenblatt Odontites In: Euro+Med Plantbase - the information resource for Euro-Mediterranean plant diversity.
- Datenblatt Odontites bei POWO = Plants of the World Online von Board of Trustees of the Royal Botanic Gardens, Kew: Kew Science.
- Ridha El Mokni, Houcine Sebei, Mohamed Hédi El Aoun: Rediscovery of a rare North African endemic Odontites (Orobanchaceae): first record and variability from Tunisia., In: International Journal of Advanced Research, Volume 3, Issue 2, 2015, S. 376–382. Volltext-PDF.
- Salvatore Brullo, Valeria Tomaselli, Robert Philipp Wagensommer: A new species of Odontites (Orobanchaceae) from southern Italy, In: Phytotaxa, Volume 213, Issue, 3, 17. Juni 2015, S. 271–281. doi:10.11646/phytotaxa.213.3.7
- Odontites im Germplasm Resources Information Network (GRIN), USDA, ARS, National Genetic Resources Program. National Germplasm Resources Laboratory, Beltsville, Maryland. Abgerufen am 4. August 2015.
- Helmut Genaust: Etymologisches Wörterbuch der botanischen Pflanzennamen. 3., vollständig überarbeitete und erweiterte Auflage. Birkhäuser, Basel/Boston/Berlin 1996, ISBN 3-7643-2390-6 (Nachdruck ISBN 3-937872-16-7).
- Martin Krampen: Pflanzenlesebuch: Pflanzenstudium – Pflanzennutzung – Pflanzenpoesie. Georg Olms Verlag, 1994, ISBN 978-3-487-09829-6.
- Carl Ferdinand Gräfe: Encyclopädisches Wörterbuch der medicinischen Wissenschaften. Band 11. Verlag von Veit et Comp., 1834 (eingeschränkte Vorschau in der Google-Buchsuche).
- August Johann Georg Carl Batsch: Versuch einer Anleitung zur Kenntniss und Geschichte der Pflanzen für academische Vorlesungen entworfen und mit den nöthigsten Abbildungen versehen. 2. Teil. Johann Jacob Gebauer, 1788 (eingeschränkte Vorschau in der Google-Buchsuche).
- Heinrich Gottfried Mattuschka: Flora Silesiaca oder Verzeichniß der in Schlesien wildwachsenden Pflanzen, 2. Teil, Verlag Wilhelm Gottlieb Korn, Breslau und Leipzig, 1777.
- Christian Gottlieb Ludwig: Institutiones Historico-Physicae Regni Vegetabilis, Verlag J. F. Gleditsch, Leipzig, 1757.
- Linné beschrieb Euphrasia odontites (= Odontites vulgaris), Euphrasia linifolia, Euphrasia lutea (beide = Odontites luteus) und Euphrasia viscosa (= Odontites viscosus); vergleiche: Carl von Linné: Species Plantarum, Verlag Lars Salvius, Stockholm, 1753. Seite 604f.
- George Bentham: Odontites. In: Alphonse de Candolle (Hrsg.): Prodromus systematis naturalis regni vegetabilis, Band 10, 1846. S. 549–552. eingescannt bei botanicus.org.
- George Bentham: Genera plantarum: ad exemplaria imprimis in Herberiis Kewensibus servata definita, Band 2, Teil 2, Verlage Reeve & Co., Williams & Norgate, London 1876. eingescannt bei botanicus.org.
- Josef Hoffmann: Beitrag zur Kenntniss der Gattung Odontites. In: Österreichische Botanische Zeitschrift, 47. Jahrgang, Nummer 4, April 1897, S. 113–117, doi:10.1007/BF01795149 (zobodat.at [PDF]).
- Werner Rothmaler: Die Aufspaltung von Odontites Hall. ex. Zinn. In: Mitteilungen des Thüringischen Botanischen Vereins, Neue Folge, Heft 50: Festschrift zum 80. Geburtstag von Joseph Bornmüller. Verlag Gebr. Knabe KG, Weimar, 1943. Seiten 224–230.
- Nelson D. Young et al.: The evolution of parasitism in the Scrophulariaceae/Orobanchaceae: plastid gene sequences refute an evolutionary transition series. In: Annals of the Missouri Botanical Garden, Band 86, 1999, S. 876–893.eingescannt bei botanicus.org.
- Richard Olmstead et al.: Disintegration of the Scrophulariaceae. In: American Journal of Botany, Band 88, Heft 2, 2001. S. 348–361.
- Angiosperm Phylogeny Group: An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II. In: Botanical Journal of the Linnean Society, Band 141. S. 399–436. doi:10.1046/j.1095-8339.2003.t01-1-00158.x
Literatur
- Markus Bolliger: Monographie der Gattung Odontites (Scrophulariaceae) sowie der verwandten Gattungen Macrosyringion, Odontitella, Bornmuellerantha und Bartsiella. In: Willdenowia: Annals of the Botanic Garden and Botanical Museum Berlin-Dahlem, Band 26, 1996. S. 37–168. (Online: Teil 1 (PDF; 2,4 MB), Teil 2 (PDF; 1,2 MB), Teil 3 (PDF; 1,2 MB), Teil 4; PDF; 2,1 MB)
- Agnes Scheunert, Andreas Fleischmann, Catalina Olano-Marín, Christian Bräuchler, Günther Heubl: Phylogeny of tribe Rhinantheae (Orobanchaceae) with a focus on biogeography, cytology and re-examination of generic concepts. In: Taxon, Volume 61, Issue 6, 2012, S. 1269–1285. JSTOR:24389112
- Daniel M. Joel, Jonathan Gressel, Lytton J. Musselman: Parasitic Orobanchaceae: Parasitic Mechanisms and Control Strategies. Springer Science & Business Media, 2013, ISBN 978-3-642-38146-1, S. 513 (eingeschränkte Vorschau in der Google-Buchsuche).
Weblinks
- Suche nach „Odontites“ in der Roten Liste gefährdeter Arten der IUCN.