Wasserpotential
Das Wasserpotential ist ein Begriff, der in der Pflanzenphysiologie verwendet wird, um die Verfügbarkeit von Wasser in einem System (z. B. Pflanzengewebe, Boden oder Luft) zu charakterisieren. Er wird zur Beschreibung der Wasseraufnahme und des Wassertransports in Pflanzen verwendet und mit oder (griechischer Buchstabe Psi) bezeichnet. Differenzen bzw. Gradienten des Wasserpotentials treiben den Wassertransport an, wobei das Wasser vom Ort mit dem höheren zum Ort mit dem niedrigeren Potential fließt (z. B. Boden – Wurzel, Blatt – Luft).[1][2][3][4]


Aus physikalischer Sicht ist das Wasserpotential das chemische Potential des Wassers, skaliert auf Einheiten des Drucks. Es ist ein Maß für die Arbeit, die geleistet werden muss, um (bei konstantem Druck und konstanter Temperatur) ein Einheitsvolumen Wasser aus einem Referenzzustand dem System zuzuführen.[3] Da Wasserpotentiale in der Natur in der Regel negative Werte annehmen, hat das höhere Potential (im Sinne der größeren Zahl inkl. Vorzeichen) den kleineren Zahlenwert, und umgekehrt.[1] So wird zum Beispiel Wasser von einer Stelle mit einem Wasserpotential von (völlig gesättigt) zu einer anderen Stelle mit einem Wasserpotential von fließen.


In älterer Literatur wird gelegentlich der Begriff Saugkraft als Triebkraft für den Wassertransport in Pflanzen verwendet. Dieser Begriff ist weniger präzise, nicht einheitlich definiert und verhält sich anschaulich gesprochen umgekehrt zum Wasserpotential: Je niedriger das Wasserpotential, desto höher die Saugkraft und umgekehrt.[5][4][6] Ein verwandtes Konzept ist das Hydraulische Potential in der Bodenkunde. Im Unterschied zum Wasserpotential werden dabei osmotische Effekte außer Acht gelassen, sodass es sich zur Beschreibung von Transportprozessen, bei denen gelöste Stoffe ungehindert mit dem Wasser transportiert werden (z. B. Versickerung), eignet.
Definition
Das Wasserpotential wird über das chemische Potential von Wasser definiert:[1][2][3][7]

- .

Hierbei ist das chemische Potential in einem Standardzustand, üblicherweise reines Wasser bei Atmosphärendruck auf einer festgelegten Referenzhöhe, und das Molvolumen von reinem flüssigem Wasser. Die Größenart des Wasserpotentials ist damit die Energie pro Volumen, was gleichbedeutend mit Druck ist. Als Maßeinheit wird meist Megapascal () verwendet.



Zerlegung in Teilpotentiale
Wasserpotential in der flüssigen Phase
Das Wasserpotential wird häufig in eine Summe von Teilpotentialen zerlegt, welche unterschiedliche physikalische Effekte beschreiben. In der flüssigen Phase (bzw. in mit wässriger Lösung gefüllten porösen Medien) gilt dabei:[1][2][8][9]

mit dem Osmotischen Potential , dem Druckpotential , dem Matrixpotential und dem Gravitationspotential .




Das Osmotische Potential (auch: Lösungspotential) ist von der Stoffmengenkonzentration gelöster Stoffe, insbesondere Salzen, abhängig. Es ist gleich dem Negativen des osmotischen Drucks , d. h. . In hinreichend verdünnten Lösungen gilt das Van-’t-Hoff’sche Gesetz:[10][1][2]



wobei die Gesamtkonzentration der gelösten Stoffe, die universelle Gaskonstante und die absolute Temperatur ist. Das osmotische Potential ist bei verdünnten Lösungen eine kolligative Eigenschaft. In Böden in Trockengebieten und in den Marschen kann das osmotische Potential besonders ausgeprägt negativ sein.



Das Druckpotential berücksichtigt den Effekt des Drucks innerhalb des beobachteten Systems. Als Referenzdruck wird meist der Atmosphärendruck () verwendet. Dieser Term ist wegen des Turgors vor allem im Protoplast bzw. Symplast von Pflanzenzellen relevant. Negative Werte treten beispielsweise im Xylem auf. Der hydrostatische Druck spielt in der Bodenkunde unterhalb der Grundwasseroberfläche eine Rolle.




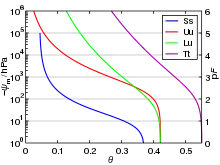


Das Matrixpotential (auch: Kapillarpotential) umfasst alle Oberflächeneffekte, mit denen Wasser von einem porösen Medium (Boden oder Zellwand bzw. Apoplast) festgehalten wird, sowie kolloidale Effekte. Das Matrixpotential ist umso stärker negativ, je feiner die Poren sind (im Boden: je feinkörniger er strukturiert ist). Wenn die Poren langsam austrocknen, steigt der Betrag des Matrixpotentials an, bis nur noch das nicht mobilisierbare Totwasser in den feinsten Poren vorhanden ist. In der Bodenkunde entspricht das Matrixpotential der Bodenwasserspannung mit umgekehrtem Vorzeichen: . Der Zusammenhang zwischen Matrixpotential und relativem Wassergehalt im Boden wird als Wasserspannungskurve dargestellt und häufig durch die Van-Genuchten-Gleichung empirisch beschrieben:[12][11]



wobei , , , und empirisch zu bestimmende Parameter sind.





Das Gravitationspotential beschreibt den Einfluss der Lageenergie auf das Wasser. Mit der Dichte flüssigen Wassers , der Erdbeschleunigung und der Höhe relativ zum Referenzzustand gilt . Das Gravitationspotential ist dann relevant, wenn Wasser über große Höhen (mehrere Meter) transportiert wird, beispielsweise in Bäumen.




Eingesetzt ergibt sich:

Wasserpotential in der Gasphase
Für das Wasserpotential in der Gasphase, insbesondere also in Luft, gilt:[2]

Dabei ist wieder die universelle Gaskonstante, die absolute Temperatur und das Molvolumen von flüssigem Wasser; ist der Partialdruck des Wasserdampfs und der Sättigungsdampfdruck von Wasser bei der gegebenen Temperatur.[Anm. 1] Da der Quotient gleich der relativen Luftfeuchtigkeit ist, lässt sich dies auch schreiben als:






Typische Werte
Reines Wasser auf Referenzhöhe und bei Atmosphärendruck besitzt definitionsgemäß das Wasserpotential . Gut bewässerte, salzarme Böden haben leicht negatives Wasserpotential von etwa .[13] Am permanenten Welkepunkt hat ein Boden ein Wasserpotential im Bereich bis .[9] Im gut bewässerten Zustand herrscht in Wurzeln ein Wasserpotential von bis .[13] In Blättern von krautigen Pflanzen liegt das Wasserpotential normalerweise zwischen und , bei Bäumen und Sträuchern können die Werte stärker negativ ausfallen, bis etwa .[1][13] In Blättern von Pflanzen, die an stark arides Klima angepasst sind, sind sogar Werte unter möglich.[1] Die Umgebungsluft hat bei und einer Luftfeuchtigkeit von gemäß obiger Formel für die Gasphase ein Wasserpotential von .[13] Meerwasser hat aufgrund der osmotischen Effekte der gelösten Salze ein Wasserpotential von etwa .[14]












Wasserfluss durch eine Membran
Die Wasserpotentialdifferenz stellt die Triebkraft für den Wasserfluss durch eine ideale, d. h. für gelöste Stoffe vollkommen undurchlässige Membran (oder sonstige Grenzfläche) dar. Die Fließgeschwindigkeit bzw. volumetrische Wasserflussdichte (in ) lässt sich in diesem Fall ausdrücken als[15][16]



wobei die Differenz der Wasserpotentiale zwischen den beiden Seiten der Membran und die hydraulische Leitfähigkeit (in ) ist. Für die gute Wasserleitfähigkeit vieler Biomembranen sind Aquaporine in entscheidender Weise verantwortlich.[Anm. 2]



Für reale Membranen, die für gelöste Stoffe teilweise durchlässig sind, verringert sich der Beitrag des osmotischen Potentials zum Wassertransport. Man kann die Fähigkeit einer Membran oder Grenzfläche, gelöste Stoffe zurückzuhalten, in einem Reflexionskoeffizienten (mit ) zusammenfassen und erhält



wobei das hydraulische Potential bezeichnet. Im Grenzfall einer ideal reflektierenden Membran () ergibt sich obige Gleichung mit der Wasserpotentialdifferenz als Triebkraft. Im Grenzfall einer Membran, die gelöste Stoffe ungehindert mit dem Wasser hindurchlässt (), ergibt sich die Differenz des hydraulischen Potentials als Triebkraft anstelle der des Wasserpotentials. Der Reflexionskoeffizient ist abhängig von der Art der gelösten Stoffe (Größe, elektrische Ladung, Polarität) sowie von der Art und dem Zustand der Membran.



Experimentelle Bestimmung
Messung in Pflanzengeweben
Abhängig von dem zu untersuchenden Pflanzengewebe kommen unterschiedliche Messverfahren zum Einsatz. Meistens werden einzelne Teilpotentiale getrennt gemessen und anschließend die Summe berechnet.[Anm. 3]
Das Wasserpotential im Xylemsaft eines Blatts (oder Sprosses) kann beispielsweise gemessen werden, indem man dieses in eine sogenannte Scholander-Bombe einspannt. Dies ist eine Kammer, aus der nur der abgetrennte Stiel des Blattes herausragt. Erhöht man den Luftdruck in der Kammer ausgehend vom Atmosphärendruck so weit, bis Xylemsaft an der Schnittfläche auszutreten beginnt, so ist diese Druckdifferenz ein Maß für das zuvor im Xylem herrschende (negative) Druckpotential . Misst man bei einer weiteren Druckerhöhung das Volumen an austretendem Saft in Abhängigkeit vom Druck, so kann man aus der sich ergebenden Druck-Volumen-Kurve auch das osmotische Potential ermitteln.[2][17] Da das Matrixpotential im Xylem keine große Rolle spielt () und das Gravitationspotential falls nötig leicht aus der Höhe des Blattes berechnet werden kann (), kann man so durch Addition das Wasserpotential im Xylem bestimmen.



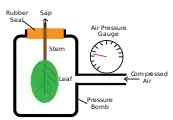 Funktionsprinzip der Scholander-Bombe
Funktionsprinzip der Scholander-Bombe Scholander-Bombe
Scholander-Bombe Bei einem bestimmten Druck in der Kammer tritt der Xylemsaft auf der Schnittfläche hervor.
Bei einem bestimmten Druck in der Kammer tritt der Xylemsaft auf der Schnittfläche hervor.
Die hydraulischen Eigenschaften einer einzelnen Zelle lassen sich im sogenannten Höfler-Diagramm darstellen. Dazu wird das Wasserpotential der Zellwand durch tränken in einer Lösung mit bekannten osmotischen Eigenschaften sukzessive variiert und das sich im Gleichgewicht einstellende Zellvolumen gemessen. Zur experimentellen Bestimmung des osmotischen Potentials kann die Gefrierpunkterniedrigung genutzt werden, da beides kolligative Eigenschaften sind. Daneben kann das Druckpotential bei manchen Zellen direkt durch Anstechen der Vacuole mit einer Drucksonde gemessen werden.[2][17]
Messung im Boden
Das Wasserpotential eines Bodens kann mit Hilfe von Gipsblock-Elektroden direkt gemessen werden. Dabei wird ein Gipsblock in den Boden eingebaut und die elektrische Leitfähigkeit innerhalb des Blocks gemessen:
- Bei hohem Wassergehalt des Bodens (niedriger Betrag des Wasserpotentials) sind viele der Poren des Blocks mit Wasser gefüllt und leiten den Strom besser.
- Bei niedrigem Wassergehalt des Bodens (hoher Betrag des Wasserpotentials, d. h. jedoch stärker negative Zahl) sind wenige der Poren des Blocks mit Wasser gefüllt und leiten den Strom schlechter.
Mit Hilfe eines Tensiometers kann das Matrixpotential bestimmt werden. Dieses entspricht in salzarmen Böden im Wesentlichen dem Wasserpotential, da hier das Osmotische Potential vernachlässigbar ist.[18]
Historische Entwicklung
Erforschung des Wassertransports in Pflanzen

Bis ins 19. Jahrhundert hinein wurde der Wassertransport in Pflanzen, wie viele andere Prozesse in der belebten Natur, in der Tradition von Aristoteles’ De anima mit einer den Lebewesen eigenen Seele begründet. Grundlegend für das moderne Verständnis des Wassertransports in Pflanzen und für die Entwicklung des Begriffs des Wasserpotentials waren die Erkenntnisse zur Diffusion und Osmose in der zweiten Hälfte des 19. Jahrhunderts durch Adolf Fick, Moritz Traube, Hugo de Vries und Wilhelm Pfeffer (vgl. Pfeffersche Zelle). Pfeffer soll um 1900 bereits gelegentlich Potentialkonzepte verwendet haben. Nachdem klar wurde, dass der osmotische Druck alleine nicht ausreicht, um alle Transportvorgänge zu beschreiben, entstanden bis Mitte des 20. Jahrhunderts unterschiedliche, konkurrierende Begriffe (Saugkraft, Hydratur, …), mit denen versucht wurde, die relevanten Triebkräfte zusammenzufassen. Ab der Mitte des 20. Jahrhunderts hat sich im Zuge verbesserter Messverfahren der Begriff des Wasserpotentials als fundamentales Konzept etabliert.[20]
Tang und Wang kritisierten 1941 die unscharfe und uneinheitliche Begriffsbildung und schlugen vor, den Wasserstatus von pflanzlichen und tierischen Zellen mit Begriffen der Thermodynamik, insbesondere dem chemischen Potential, zu beschreiben.[21] Der Begriff des Wasserpotentials soll von Schofield geprägt worden sein.[22] Diesen Begriff griff Owen 1952 auf, als er nachwies, dass Weizensamen auch unter extrem trockenen Bedingungen zur Wasseraufnahme und Keimung im Stande sind und dies zu erklären versuchte.[23] Der Begriff des Wasserpotentials wurde schließlich von Slatyer und Taylor verbreitet:[4]
“An alternative method of expression is in terms of the chemical potential of water, which is identical with the partial molal free energy. This procedure was adopted by Day (1942) who referred to it as ‘moisture potential.’ Schofield, cited by Owen (1952), suggested the term ‘water potential’ for the same function, and more recently the 1959 UNESCO conference on plant water relations in arid regions favourably considered a terminology based on an evaluation of this function.”
„Eine alternative Methode dies auszudrücken ist vom chemischen Potential von Wasser auszugehen, welches identisch mit der partiellen molalen Freien Energie ist. Diese Vorgehensweise wurde von Day (1942) benutzt, der sie als ‚Feuchtigkeitspotential‘ bezeichnete. Schofield, den Owen (1952) zitierte, schlug den Begriff ‚Wasserpotential‘ für dieselbe Funktion vor und erst kürzlich, 1959, hat sich die UNESCO-Konferenz zu Pflanzenwasserthemen in ariden Regionen für eine Terminologie basierend auf der Betrachtung dieser Funktion ausgesprochen.“
Allerdings verwendet Slatyer in diesem Artikel für das Wasserpotential noch die Größenart des chemischen Potentials und argumentiert gegen die Verwendung von der Größenart des Drucks.[22]
Im Strasburger – Lehrbuch der Botanik wird das Wasserpotential erstmals in der 31. Auflage von 1978 verwendet.[7] Bis einschließlich der 30. Auflage von 1971 wurde der ältere, weniger scharf definierte Begriff Saugkraft benutzt.[5] Während die Definition des Wasserpotentials als das chemische Potential von Wasser bezogen auf sein Molvolumen in modernen Lehrbüchern[1][2][3][7] üblich ist, kann dies in äquivalenter Weise auch verbal ausgehend von der Freien Enthalpie ausgedrückt werden (so beispielsweise bei Richter[4]). Für die verschiedenen Teilpotentiale sind nach wie vor unterschiedliche Konventionen hinsichtlich der Notation im Gebrauch.[24][2]
Verwandte Begriffe in Nachbarwissenschaften
Die Begriffe Freie Enthalpie (gelegentlich auch Gibbs Potential genannt) und Chemisches Potential wurden in den 1870er Jahren von Josiah Willard Gibbs in der Thermodynamik eingeführt.[25] Edgar Buckingham, der 1900 bereits ein Thermodynamik-Lehrbuch verfasst hatte, führte 1907 in seiner Arbeit Studies on the Movement of Soil Moisture verwandte Potentialkonzepte in der Bodenkunde ein, wobei er von einer Definition über zu leistende Arbeit ausging.[26] Wassertransportprozesse im Boden, bei denen gelöste Stoffe ungehindert mit dem Wasser transportiert werden, werden heutzutage mit dem Hydraulischen Potential beschrieben, welches verglichen mit dem Wasserpotential das osmotische Potential außer Acht lässt.[18][27]
Dass verwandte Potentiale in unterschiedlichen Wissenschaften auf unterschiedliche Größen bezogen und damit in unterschiedlichen Größenarten angegeben werden, hat sowohl historische wie auch praktische Gründe. In der chemischen Thermodynamik ist das Mol die natürliche Bezugsgröße, weshalb das chemische Potential in gemessen wird. Für die Pflanzenphysiologie ist der Druck eine sinnvolle Größenart, weil in Pflanzenzellen die Druckkomponente (Turgor) einen beträchtlichen Beitrag zum Potential leistet und auch osmotische Eigenschaften traditionell als osmotische Drücke angegeben werden. Deshalb wird hier als Bezugsgröße das Volumen gewählt, was auf das Wasserpotential in Einheiten des Drucks (; veraltet auch , , oder ) führt. In der Bodenkunde kommen verschiedene Bezugsgrößen zum Einsatz, wobei dort die Höhe einer Wassersäule ein besonders anschauliches Maß darstellt, welches unmittelbar mit Messungen im Wasserstandsrohr korrespondiert.[18][27][Anm. 4]






Literatur
- Lincoln Taiz, Eduardo Zeiger, Ian Max Møller, Angus Murphy (Hrsg.): Plant Physiology and Development. 6. Auflage. Sinauer Associates, Sunderland, Massachusetts, USA 2014, ISBN 978-1-60535-255-8, Chapter 3 Water and Plant Cells und Chapter 4 Water Balance of Plants, S. 83–118 (Englischsprachige Originalausgabe, Autorin Chapter 3 & 4: N. Michele Holbrook).
Bzw. deutschsprachige Übersetzung:
Lincoln Taiz, Eduardo Zeiger (Hrsg.): Physiologie der Pflanzen. 1. Auflage. Spektrum Akademischer Verlag, Heidelberg/ Berlin 2000, ISBN 3-8274-0537-8, Kapitel 3 Der Wasserhaushalt der pflanzlichen Zellen und Kapitel 4 Der Wasserhaushalt der Pflanzen, S. 59–100. - Peter Schopfer, Axel Brennecke: Pflanzenphysiologie. Begründet von Hans Mohr. 6. Auflage. Elsevier, Spektrum Akademischer Verlag, München 2006, ISBN 3-8274-1561-6, Kapitel 3 Die Zelle als energetisches System und Kapitel 13 Ferntransport von Wasser und anorganischen Ionen, S. 47–70, 311–331.
- Paul J. Kramer, John S. Boyer: Water relations in plant and soil. Academic Press, San Diego 1995, ISBN 0-12-425060-2.
- Park S. Nobel: Physicochemical and environmental plant physiology. 4. Auflage. Elsevier Academic Press, Amsterdam/ London 2009, ISBN 978-0-12-374143-1, Chapter 2 Water, Chapter 8 Leaves and Fluxes, Chapter 9 Plants and Fluxes, S. 45–100, 365–506.
Weblinks
- Plant Physiology and Development, Sixth Edition – Companion Website. Web Topics and Essays. Lincoln Taiz, Eduardo Zeiger, Ian Max Møller, Angus Murphy, 2015, abgerufen am 23. Juli 2015 (englisch, Webbasiertes Zusatzmaterial zu gleichnamigem Lehrbuch, Sinauer Associates, frei zugänglich).
Darin insbesondere:- N. Michele Holbrook u. a.: Web Topics zu Chapter 3: Water and Plant Cells. 2015, abgerufen am 20. Juli 2015 (englisch).
- N. Michele Holbrook u. a.: Web Topics und Essays zu Chapter 4: Water Balance of Plants. 2015, abgerufen am 20. Juli 2015 (englisch).
Einzelnachweise
- N. Michele Holbrook (Kapitel-Autorin Chapter 3 & 4): Plant Physiology. Hrsg.: Lincoln Taiz, Eduardo Zeiger. 6. Auflage. Sinauer Associates, Sunderland, Massachusetts, USA 2014, ISBN 978-1-60535-255-8, Chapter 3, Sections Water Potential, Water Potential of Plant Cells, Cell Wall and Membrane Properties, Plant Water Status, and Summary, S. 89–98.
- Peter Schopfer, Axel Brennecke: Pflanzenphysiologie. Begründet von Hans Mohr. 6. Auflage. Elsevier, Spektrum Akademischer Verlag, München 2006, ISBN 3-8274-1561-6, Abschnitt 3.5: Chemisches Potential von Wasser und 3.6 Anwendungen des Wasserpotentialkonzepts auf den Wasserzustand der Zelle, S. 51–61.
- Paul J. Kramer, John S. Boyer: Water relations in plant and soil. Academic Press, San Diego 1995, ISBN 0-12-425060-2, Chapter 2: Functions and Properties of Water, Subsection Chemical Potential of Water, S. 35–37.
- Gerhard Richter: Stoffwechselphysiologie der Pflanzen. Physiologie und Biochemie des Primär- und Sekundärstoffwechsels. 5. Auflage. Georg Thieme Verlag, Stuttgart 1988, ISBN 3-13-442005-8, 2. Kapitel, Abschnitt 1 Wasserhaushalt, S. 31–51.
- Dietrich von Denffer u. a.: Lehrbuch der Botanik für Hochschulen. Begründet von Eduard Strasburger u. a. 30. Auflage. Gustav Fischer Verlag, Stuttgart 1971, ISBN 3-437-20050-X, Zweiter Teil: Physiologie, Erster Abschnitt: Physiologie des Stoffwechsels, II. Das Wasser, S. 205–223 (Ab der nachfolgenden 31. Auflage von 1978 wird der Wassertransport nicht mehr mit der Saugkraft, sondern mit dem Wasserpotential beschrieben. Der Begriff „Saugkraft“ kommt im Fließtext lediglich noch bildlich zum Einsatz. Spätestens in der 37. Auflage von 2014 kommt „Saugkraft“ gar nicht mehr vor (Volltextsuche im PDF, doi:10.1007/978-3-642-54435-4).).
- Ulrich Kutschera: Kurzes Lehrbuch der Pflanzenphysiologie. Quelle und Meyer, Wiesbaden 1995, ISBN 3-8252-1861-9, Kapitel 4: Wasserhaushalt der Pflanzenzelle: Diffusion, Osmose, Wasserpotential, S. 60.
- Peter Sitte: Lehrbuch der Botanik für Hochschulen. Begründet von Eduard Strasburger. 33. Auflage. G. Fischer, Stuttgart / Jena / New York 1991, ISBN 978-3-437-20447-0, Zweiter Teil Physiologie, IV Nährstoffe und ihr Umsatz in Pflanzen, B Wasserhaushalt, S. 321–337 (Diese Definition des Wasserpotentials über das chemische Potential findet sich erstmals in der 31. Auflage von 1978. In älteren Ausgaben, konkret in der 29. Auflage von 1967 und der 30. Auflage von 1971, kommt „Wasserpotential“ nicht vor; stattdessen wird der Begriff „Saugkraft“ verwendet. In neueren Ausgaben, konkret in der 36. Auflage von 2008 und der 37. Auflage von 2014, wird zwar mit dem „Wasserpotential“ gerechnet, eine Definition fehlt aber.).
- Paul J. Kramer, John S. Boyer: Water relations in plant and soil. Academic Press, San Diego 1995, ISBN 0-12-425060-2, Chapter 3: Cell Water Relations, Section Water Status, S. 49–53.
- Paul J. Kramer, John S. Boyer: Water relations in plant and soil. Academic Press, San Diego 1995, ISBN 0-12-425060-2, Chapter 4: Soil and Water, Section Soil water terminology, S. 89–93.
- Gerd Wedler, Hans-Joachim Freund: Physikalische Chemie. 6., vollst. überarb. und aktualisierte Auflage. Wiley-VCH Verlag, Weinheim 2012, ISBN 978-3-527-32909-0, Abschnitt Osmotischer Druck in 2.5.6 Phasengleichgewichte in Zweikomponentensystemen zwischen einer Mischphase und einer reinen Phase, S. 348–352 (Gleichung (2.5-79)).
- Ad-hoc-AG Boden: Verknüpfungsregel 1.18 – Parameter für das Modell einer stetigen Funktion der θ(ψ)-Beziehung. (PDF; 242 kB) Staatlichen Geologischen Dienste und BGR, 17. September 2004, archiviert vom (nicht mehr online verfügbar) am 4. März 2016; abgerufen am 29. Juli 2015.
- Karl-Heinrich Hartge, Rainer Horn: Einführung in die Bodenphysik. 3. Auflage. Ferdinand Enke Verlag, Stuttgart 1999, ISBN 3-432-89683-2, Abschnitt 4.6 Wasserspannung und Wassergehalt, insbesondere Unterabschnitt 4.6.5 Mathematische Beschreibung der Wasserspannungskurve, S. 130–137.
- Peter Schopfer, Axel Brennecke: Pflanzenphysiologie. Begründet von Hans Mohr. 6. Auflage. Elsevier, Spektrum Akademischer Verlag, München 2006, ISBN 3-8274-1561-6, Kapitel 13: Ferntransport von Wasser und anorganischen Ionen, S. 311–331 (insbes. Abb. 13.2 auf S. 313).
- Joachim W. Kadereit, Christian Körner, Benedikt Kost, Uwe Sonnewald: Strasburger − Lehrbuch der Pflanzenwissenschaften. Begründet von E. Strasburger, F. Noll, H. Schenk, A. F. W. Schimper. 37. Auflage. Springer Spektrum, Berlin/ Heidelberg 2014, ISBN 978-3-642-54435-4, Unterabschnitt 19.1.2.4 Mineralsalze als Standortfaktoren, S. 345, doi:10.1007/978-3-642-54435-4.
- N. Michele Holbrook: Web Topic 3.9: Understanding Hydraulic Conductivity. Plant Physiology and Development, Sixth Edition. Lincoln Taiz, Eduardo Zeiger, 2015, abgerufen am 21. Juli 2015 (englisch, Webbasiertes Zusatzmaterial zum gleichnamigen Lehrbuch, Sinauer Associates, frei zugänglich).
- Paul J. Kramer, John S. Boyer: Water relations in plant and soil. Academic Press, San Diego 1995, ISBN 0-12-425060-2, Chapter 3: Cell Water Relations, Section Water Transport, S. 63–68.
- N. Michele Holbrook: Web Topic 3.6: Measuring Water Potential. Plant Physiology and Development, Sixth Edition. Lincoln Taiz, Eduardo Zeiger, 2015, abgerufen am 20. Juli 2015 (englisch, Webbasiertes Zusatzmaterial zum gleichnamigen Lehrbuch, Sinauer Associates, frei zugänglich).
- Hans-Peter Blume u. a.: Lehrbuch der Bodenkunde. Begründet von Fritz Scheffer und Paul Schachtschabel. 16. Auflage. Spektrum Akademischer Verlag, Heidelberg 2010, ISBN 978-3-8274-2251-4, Abschnitt 6.4 Bodenwasser (insbes. Unterabschnitt 6.4.2.1 Potenziale) und Unterabschnitte 4.2.2 Wasser und Atmosphäre, 9.2.2 Wasserbewegungen im System Boden–Pflanze–Atmosphäre und 9.2.3 Wasserverbrauch und Pflanzenertrag, S. 220–249, 103, 384–388, doi:10.1007/978-3-8274-2251-4.
- Wilhelm Pfeffer: Osmotische Untersuchungen. Studien zur Zellmechanik. 2. Auflage. Wilh. Engelmann, Leipzig 1921, S. 5 (unveränderte Auflage des Erstdrucks von 1877).
- Paul J. Kramer, John S. Boyer: Water relations in plant and soil. Academic Press, San Diego 1995, ISBN 0-12-425060-2, Chapter 1: Historical Review, S. 1–15.
- P. S. Tang, J. S. Wang: A Thermodynamic Formulation of the Water Relations in an Isolated Living Cell. In: The Journal of Physical Chemistry. Band 45, Nr. 3, 1941, S. 443–453, doi:10.1021/j150408a010.
- R. O. Slatyer: Absorption of water by plants. In: The Botanical Review. Band 26, Nr. 3. Springer-Verlag, 1960, ISSN 0006-8101, S. 331–392, doi:10.1007/BF02860807.
- P. C. Owen: The Relation of Germination of Wheat to Water Potential. In: Journal of Experimental Botany. Band 3, Nr. 2, 1952, S. 188–203, doi:10.1093/jxb/3.2.188.
P. C. Owen: The Relation of Water Absorption by Wheat Seeds to Water Potential. In: Journal of Experimental Botany. Band 3, Nr. 9. Oxford University Press, 1952, ISSN 0022-0957, S. 276–290. - N. Michele Holbrook: Web Topic 3.3: Alternative Conventions for Components of Water Potential. Plant Physiology and Development, Sixth Edition. Lincoln Taiz, Eduardo Zeiger, 2015, abgerufen am 22. Juli 2015 (englisch, Webbasiertes Zusatzmaterial zum gleichnamigen Lehrbuch, Sinauer Associates, frei zugänglich).
- Gerd Wedler, Hans-Joachim Freund: Physikalische Chemie. 6., vollst. überarb. und aktualisierte Auflage. Wiley-VCH Verlag, Weinheim 2012, ISBN 978-3-527-32909-0, Zeittafel in vorderer Umschlagsklappe und Abschnitt 2.3, S. 301–324.
- T. N. Narasimhan: Central Ideas of Buckingham (1907): A Century Later. In: Vadose Zone Journal. Band 6, Nr. 4, 2007, ISSN 1539-1663, S. 687–693, doi:10.2136/vzj2007.0080.
- Karl-Heinrich Hartge, Rainer Horn: Einführung in die Bodenphysik. 3. Auflage. Ferdinand Enke Verlag, Stuttgart 1999, ISBN 3-432-89683-2, Abschnitt 4.4 Potential des Bodenwassers, insbes. Tabelle 4.1, S. 119–128.
Anmerkungen
- In deutlich geringerem Maß hängt der Sättigungsdampfdruck auch vom Gesamtdruck ab. Für biologische Anwendungen ist dies vernachlässigbar, da diese Abhängigkeit schwach ist und der Druck in der Gasphase üblicherweise nahe am Atmosphärendruck ist.
- Werte von in Pflanzen liegen im Bereich (Paul J. Kramer, John S. Boyer: Water relations in plant and soil. Academic Press, San Diego 1995, ISBN 0-12-425060-2, Chapter 3: Cell Water Relations, Table 3.1, S. 66–67.).
- Das Gravitationspotential kann, falls es überhaupt für die jeweilige Fragestellung relevant ist, immer leicht aus dem Höhenunterschied berechnet werden.
- Zu den Formeln für das „Wasserpotential“ bei
Hans-Peter Blume u. a.: Lehrbuch der Bodenkunde. Begründet von Fritz Scheffer und Paul Schachtschabel. 16. Auflage. Spektrum Akademischer Verlag, Heidelberg 2010, ISBN 978-3-8274-2251-4, doi:10.1007/978-3-8274-2251-4 (Gleichung (Gl. 6.4.7) auf S. 225). und
Karl-Heinrich Hartge, Rainer Horn: Einführung in die Bodenphysik. 3. Auflage. Ferdinand Enke Verlag, Stuttgart 1999, ISBN 3-432-89683-2 (S. 126 Mitte).
ist anzumerken, dass diese weder eine Definition noch eine allgemeine Berechnungsgrundlage darstellen, sondern nur als Rechenanleitung für das Wasserpotential im Boden in unmittelbarer Wurzelnähe zu verstehen sind. Vielmehr entspricht dem Wasserpotential der pflanzenphysiologischen Literatur das, was bei Scheffer/Schachtschabel/Blume ((Gl. 6.4.5) auf S. 224) und bei Hartge/Horn (S. 122 oben) als „Gesamtpotential“ bezeichnet wird.

