Vertoviridae
Vertoviridae ist eine vom International Committee on Taxonomy of Viruses (ICTV) im März 2022 neu eingerichtete Familie von Archaeenviren innerhalb der Klasse Caudoviricetes. Sie ist innerhalb dieser Klasse bisher ohne nähere Zuordnung (Stand 29. Februar 2024).[1][2]
| Vertoviridae | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|

ChaoS9-Partikel und Phylogenetischer | ||||||||||||||
| Systematik | ||||||||||||||
| ||||||||||||||
| Taxonomische Merkmale | ||||||||||||||
| ||||||||||||||
| Wissenschaftlicher Name | ||||||||||||||
| Vertoviridae | ||||||||||||||
| Links | ||||||||||||||
|


Die Zugehörigkeit zu den Caudoviricetes und ihre halophilen Archaeenwirte klassifizieren die Vertoviridae nicht-taxonomisch einerseits als arTVs (archaeal tailed viruses), andererseits als Haloviren.
Beschreibung
Morphologie
Die Virionen (Viruspartikel) der Vertoviridae haben den für die Klasse Caudoviricetes typischen Kopf-Schwanz-Aufbau; es gibt keine Virushülle. Der Kopf, ein ikosaedrisches Kapsid ist im Durchmesser etwa 64 nm groß. Der Schwanz ist kontraktil, er misst etwa 170 nm an Länge und 18 nm an Durchmesser, es gibt kurzen Schwanzfasern.[3][4]
Wirte
Die Wirte der Vertoviridae sind verschiedene Mitglieder der Euryarchaeota-Ordnung Halobacteriales (synonym: Natrialbales, Haloferacales),[5] insbesondere:
Genom und Proteom

Drei Mitglieder der Familie wurden bis 2023 sequenziert:
- phiCh1: Probe aus Österreich, 1996 entnommen, Veröffentlichung 1997 (Angela Witte et al.)[10][11]
- phiH: Probe aus Deutschland, 1981 entnommen, Veröffentlichung 2018 (Mike Dyall-Smith et al.)[12][13]
- ChaoS9: Probe aus Deutschland, 2007 entnommen, Veröffentlichung 2019 (Mike Dyall-Smith et al.)[6][7]
Das Genom der Vertoviridae ist unsegmentiert (monopartit), es ist im Allgemeinen ein lineares dsDNA-Molekül mit einer Größe von ca. 59 kbp (Kilobasenpaaren), es kodiert etwa 97 Proteine. Das Genom ist endständig (terminal) redundant,[A. 1] zudem gibt es zirkuläre Permutationen[15] (circular permutations, vergleiche Madisaviridae, Saparoviridae und die Zobellviridae-Gattung Citrovirus).[3]
Das Genom der Vertoviridae kodiert u. a. eine HNH-Endonuklease,[16][17][A. 2][10][12][6]
Das Genom von ChaoS9 hat einen G+C-Gehalt von 65,3 % und vorhergesagt 85 kodierende Sequenzen (coding regions, coding sequences, CDS)[A. 3] es kodiert eine tRNA (für Arginin).[7] Das Genom von phiH (Variante phiH1) hat einen G+C-Gehalt von (je nach Autor) 63.7–64,0 % und vorhergesagt 97 CDS.[11]
Replikationszyklus
Die Replikation erfolgt im Zytoplasma der Wirtszelle (prokaryotischer Wirt). Sie umfasst folgende Schritte:[3]
- Adsorption: Das Virion heftet sich über seine Schwanzfasern an die Zielzelle an.
- Ausstoß der viralen DNA in das Zytoplasma der Wirtszelle durch Kontraktion der Schwanzscheide.
- Transkription und Translation der „frühen“ Gene.
- Synthese linearer Kopien der Virus-DNA als Concatamer. Es folgt die Transkription und Translation der „späten“ Gene.
- Zusammenbau (Assembly) eines leeren Prokapsids und Verpackung des Genoms.
- Zusammenbau der Schwanzfasern und des Schwanzes. Fertigstellung der Virionen.
- Die reife Virionen werden schließlich durch Lyse aus der Wirtszelle freigesetzt.
Die Transkription von Genen erfolgt durch Operons; die frühe Transkription wird durch Antisense-mRNA des Virus reguliert.[3]
Systematik
Die Systematik der Vertoviridae gemäß International Committee on Taxonomy of Viruses (ICTV) ist wie folgt (Stand 29. Februar 2024):[18][4][2][7]
Familie Vertoviridae[19] (ursprünglich als „F9-Gruppe“ bezeichnet)
- Gattung Chaovirus (ursprünglich „F9G2“, ausgegliedert aus Gattung Myohalovirus)
- Spezies Chaovirus ChaoS9
- Stamm Halobacterium-Phage ChaoS9 [Halovirus ChaoS9] – Genomlänge 55.145 bp[7]
- Spezies Chaovirus ChaoS9
- Gattung Myohalovirus[20] (früher Phihlikevirus, phiH-like viruses, ursprünglich „F9G1“)
- Spezies Myohalovirus phiCh1 (früher Natrialba virus PhiCh1)
- Stamm Natrialba-Phage PhiCh1 [Bakteriophage Phi-Ch1, Natronobacterium magadii phage ΦCh1, φCh1] – Genomlänge 58.487 bp[7][11][21][22]
- Spezies Myohalovirus phiH (früher Halobacterium virus phiH)
- Stamm Halobacterium-Phage phiH [Bakteriophage phi-H] mit Variante phiH1 [ΦH1] – Genomlänge 58.072 bp[12][13]
- Stamm Halobacterium-Phage Hs1 (englisch Halobacterium phage Hs1)[23]
- Spezies Myohalovirus phiCh1 (früher Natrialba virus PhiCh1)
Etymologie
Der Name der Familie Vertoviridae leitet sich ab von lateinisch verto ‚ich wende‘ (von vertere ‚wenden‘);[24] dies bezieht sich auf den für die Viren dieser Familie charakteristischen invertierbaren Schwanzfaserlocus, die Endung ‚-viridae‘ bezeichnet Virusfamilien.[1][4][2]
- Der Gattungsname Chaovirus, eine Zusammenziehung (Kofferwort) von ChaoS9 virus, leitet sich ab vom Namen des ersten isolierten Virus dieser Gattung, ChaoS9.[4]
- Das Art-Epitheton ChaoS9 ist der Name des Referenzstammes, der seinerseits ein Kofferwort ist aus Caudovirus of haloarchaeal origin und dem Stamm S9 der von diesem Virus parasitierten Archaeenart Halobacterium salinarum ist.[4]
- Der Gattungsname Myohalovirus leitet sich ab von einer Zusammenziehung der beiden nicht-taxonomischen Klassifizierungen dieser Gattung, den Myoviren (Morphotyp) und Haloviren (Habitat).[4][7]
- Das Art-Epitheton phiCh1 verweist auf den Referenzstamm dieser Art, phi-Ch1.
- Auch das Art-Epitheton phiH leitet sich ab vom Referenzstamm der Spezies, phi-H.
Hs1
Die erste Beschreibung eines Archaeenvirus erfolgte 1974 durch Terje Torsvik und Ian D. Dundas in einem Nature-Artikel mit dem Titel „Bacteriophage of Halobacterium salinarum“ („Bakteriophage von Halobacterium salinarum“)[25][26] Dieses Virus mit dem Akronym Hs1[27][28][29] infiziert die Haloarchaeen-Spezies Halobacterium salinarum und hat signifikante Ähnlichkeit mit dem Halobacterium-Phagen phiH (ΦH) und ist vom Morphotyp der Myoviren, es muss daher derselben Virusspezies Myohalovirus phiH angehören.[27][12]
ChaoS9
ChaoS9 ist der Referenzstamm der Virenspezies Chaovirus ChaoS9.
ChaoS9-Genom
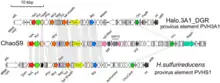
Auf das ChaoS9 stieß man, nachdem eine große Kultur von Archaeen des Stamms Halobacterium salinarum S9 spontan zusammengebrochen war. Wie sich herausstellte, war sie von dem Virus infiziert und lysiert worden.[6][7] Die Isolation des ChaoS9 genannten Virus aus dem Kulturlysat ergab eine homogene Population von Partikeln mit der Kopf-Schwanz-Morphologie der Caudoviricetes. Das Virus-Genom erwies sich als eine lineare, teilweise redundante und zirkulär permutierte dsDNA (Doppelstrang-DNA) mit einer Länge von 55.145 bp und einem G+C-Anteil von 65,3 %, nach Vorhersage 85 kodierenden Sequenzen (CDS) sowie einem tRNA-Gen für die Aminosäure Arginin (wie oben beschrieben).[7]
Der linke Arm des Virus-Genoms (Position 0–28 kbp) kodiert für Proteine, die in ihrer Sequenz denen damals bereits bekannten und (teil-)sequenzierten Caudoviricetes-Mitgliedern ähnlich waren; am ähnlichsten waren die Myohaloviren phiH1 (Wirt: ebenfalls Halobacterium salinarum) und phiCh1 (Wirt: die verwandte Spezies Natrialba magadii). Die ChaoS9-Gene für den Schwanz der Viruspartikel ähneln denen von phiH1 und phiCh1, insbesondere trägt ChaoS9 auch ein Schwanzfaser-Genmodul, das den invertierbaren Modulen dieser beiden Viren ähnelt. Alle drei Viren werden daher vom Morphotyp des Schwanzes her als Myoviren klassifiziert. Interessanterweise ist jedoch das Hauptkapsidprotein Mcp von ChaoS9 dem von Haloarcula hispanica tailed virus 1 (HHTV-1, aus der Caudoviricetes-Familie Madisaviridae, Morphotyp: Siphoviren) am ähnlichsten. Mit ChaoS9 verwandte Provirus-Elemente zeigten ähnlich die größte Übereinstimmung Proteinen für den Zusammenbau des Virus-Schwanzes, wichen aber im Vergleich der Proteine für die Kopf-Assembly ab.[7]
Der rechte Arm (Position 29–55 kbp) von ChaoS9 kodiert für Proteine, die an der DNA-Replikation beteiligt sind; die anderen Proteine weisen wenig Ähnlichkeit mit denen von phiH1, phiCh1 oder Provirus-Elementen auf – die meisten von ihnen konnten 2019 keiner bekannten Funktion zugeordnet werden.[7] Während die Schwanzgenregion von ChaoS9 den beiden anderen Viren ähnlich ist, gilt das weder für die Gene zum Zusammenbau des Kopfes auf dem linken Genomarms noch für den größten Teil des rechten Arms (mit Replikations- und Lyse-Genen). Insbesondere sind das Hauptkapsidprotein, die große Untereinheit der Terminase und die Portalproteine von ChaoS9 alle nicht mit denen von phiH1 und phiCh1 verwandt.[7]
Evolutionärer Ursprung von ChaoS9
Dieses ausgeprägte Muster von Ähnlichkeiten bzw. Unterschieden des ChaoS9-Genoms mit den Genomen von phiH1 und phiCh1 deutet auf eine Evolutionsgeschichte mit (mindestens einem) großen Rekombinationsereignis als plausibelste Erklärung hin: Danach hat ein Rekombinationsereignis das Morphogenesemodul für den ersetzt hat, während das für den Schwanz unverändert blieb. Durch diese Rekombination ist ChaoS9 als ein chimäres Virus entstanden, wobei die Grenze zwischen den mutmaßlichen Genen für den Kopfverschluss (head closure, hco) und das Halsprotein (neck protein, nep) liegt: Der (maßgebliche) Schwanz ist zwar vom Morphotyp der Podoviren, aber der Kopf ist vom Morphotyp der Siphoviren. In der Literatur zu den Caudoviricetes sind Beispiele keine Seltenheit, bei denen die Module für die Zusammensetzung von Kopf und Schwanz von verschiedenen Virustypen abzustammen scheinen; wie etwa:[7]
- Gordonia spp. Phage Kita,[30][31]
- Xylella-fastidiosa-Phage Xfas53,[32][33]
- Enterobacteria-Phage phi80,[34][35]
- Salmonella-Typhimurium-Phage Gifsy-2.[36][35]
Durch den Umstand, dass bei den Caudoviricetes zunächst Kopf und Schwanz als separate Strukturen gebildet und anschließend zusammengefügt werden, ist die Wahrscheinlichkeit dafür, dass solche Rekombinationsereignisse lebensfähige Nachkommen hervorbringen, erhöht.[7]
ΦCh1
ΦCh1 (auch phiCh1 geschrieben) ist der Referenzstamm der Virenspezies Myohalovirus phiCh1.
Isolierung und Virus-Wirt-Beziehung von ΦCh1
ΦCh1 infiziert bestimmte Stämme der haloalkalophilen (Salz und hohe pH-Werte liebenden) Archaeenart Natrialba magadii (veraltet: Natronobacterium magadii)[8][9] aus der Ordnung Halobacteriales (synonym: Natrialbales).[5] und ist ein (im Allgemeinen) temperentes Virus und kommt als Provirus (Prophage) im Genom seines Wirts integriert vor.[11]
Die Beobachtung, dass Batch-Kulturen von N. magadii (Referenzstamm DSM:3394 alias MS3[8][9]) spontan lysiert wurden, führte zur erstmaligen Isolierung des Virus ΦCh1, für das dies Archaeenart als Wirt spezifisch ist. Zum Nachweis der Infektiosität von ΦCh1 durch Witte et al. (1997) wurde der von Viren (inklusive Proviren) befreite Stamm N. magadii L13 verwendet. ΦCh1 ist das erste Virus, das für die Gattung Natrialba (bzw. Natronobacterium, wie ursprünglich bezeichnet) beschrieben wurde.[11]
Eine erneute Infektion des Stammes N. magadii [DSM:3394] mit dem isolierten Virus ΦCh1 war nicht möglich, was auf ein Art „Immunsystem“ schließen lässt, wie es beispielsweise auch für die lysogenen Stämme R1–3 und R1–24 von Halobacterium salinarum beschrieben wurde (Stolt und Zillig, 1992). N. magadii wurde durch wiederholtes Subkultivieren und Testen einzelner Kolonien durch Infektion mit dem ΦCh1 von lysogenen Viren „geheilt“ (d. h. es wurde eine Linie erzeugt, die frei von diesen Viren und Proviren ist). Auf diese Weise wurde ein nicht lysogener Stamm von N. magadii isoliert, der als L13 bezeichnet wird. (Witte et al., 1997). Als Ergebnis kann der Stamm L13 kann mit ΦCh1 infiziert werden, was sowohl zur Lyse der infizierten Zellen als auch zur Lysogenisierung (Einlagerung ins Genom als Provirus) führt.[11]
Das Virus bildet trübe Plaques auf dem Zellrasen auf Agarplatten („Petrischalen“). Aus einer einzelnen Plaque wurde eine virenproduzierende Kolonie isoliert, die den Namen L11 erhielt und in der Studie von Witte et al. (1997) verwendet wurde. Da die Zelllyse nur in stationären Kulturen beobachtet wurde, scheint sie wachstumsphasenabhängig zu sein.[11]
Genom und Wirts-RNA von ΦCh1
Die Morphologie seiner Virionen (Viruspartikel) ist ähnlich wie bei anderen die Archaeenart Halobacterium infizierendem Caudoviricetes-Mitgliedern vom Morphotyp der Myoviren. In Natriumchlorid-Lösungen unter 2 ᴍ (mol/ℓ) verliert das Virus seine morphologische Stabilität und Infektiosität.[11]
Die Virionen von ΦCh1 enthalten sowohl doppelsträngige, lineare DNA mit einer Länge von ca. 55 kbp (Kilobasenpaaren) als auch mehrere RNA-Typen mit etwa 80 bis 700 Nukleotiden Länge. Hybridisierungsversuche von markierten Fragmenten dieser virion-assoziierten RNA mit der DNA des Genoms von N. magadii und von ΦCh1 selbst zeigten, dass diese RNA wirtskodiert ist. ΦCh1 war zum Zeitpunkt der Erstbeschreibung 1997 das einzige unter den bekannten Archaeenviren, dessen reife Partikel sowohl (eigene) DNA als auch (Wirts-)RNA enthalten.[11][A. 4]
Im Gegensatz zur DNA von N. magadii ist die DNA von ΦCh1 zumindest teilweise methyliert. Offenbar kodiert ΦCh1 entweder seine eigene Methylase oder es aktiviert eine stille (unter normalen Umständen nicht exprimierte) Methylase seines Wirts.[11]
Anmerkungen
- Endständig redundante DNA ist DNA, die an jedem Ende wiederholte Sequenzen, so genannte terminale Repeats, enthält. Wie z. B. bei Escherichia-Virus T4 werden mit diesen Sequenzen die Enden der linearen DNA verbunden um für die Replikation vorübergehend eine zyklische DNA zu bilden. Der Begriff wurde erstmals 1964 von Michael London geprägt.[14]
- siehe auch Homing-Endonuklease.
- vergleiche Offener Leserahmen (open reading frame, ORF);
- Die Partikel der Arenaviridae enthalten Ribosomen ihres Wirts und damit ebenfalls (ribosomale) Wirts-RNA, sie haben als RNA-Viren aber selbst ein RNA-Genom.
Einzelnachweise
- ICTV: Family: Vertoviridaee. 2022 Release, MSL #38.
- Mart Krupovic: Family: Vertoviridaee (interim Report). Juni 2023.
- SIB: Vertoviridae. Auf: ViralZone.
- Ying Liu, Tatiana A. Demina, Simon Roux, Pakorn Aiewsakun, Darius Kazlauskas, Peter Simmonds, David Prangishvili, Hanna M. Oksanen, Mart Krupovic: Create three new orders and 14 new families in the class Caudoviricetes (Duplodnaviria, Uroviricota) for classification of archaeal tailed viruses (zip:docx), PDF (via Universität Helsinki). Vorschlag 2021.001A vom Oktober 2020.
-
LPSN: Order Halobacteriales Grant and Larsen 1989. Synonyms:
- Natrialbales Gupta et al. 2015
- Haloferacales Gupta et al. 2015
- NCBI Nucleotide: Halobacterium phage ChaoS9.
- Mike Dyall-Smith, Peter Palm, Gerhard Wanner, Angela Witte, Dieter Oesterhelt, Friedhelm Pfeiffer: Halobacterium salinarum virus ChaoS9, a Novel Halovirus Related to PhiH1 and PhiCh1. In: MDPI: Genes, Band 10, Nr. 3, 1. März 2019, Special Issue Genetics of Halophilic Microorganisms, S. 194; doi:10.3390/genes10030194 (englisch).
- LPSN: Species Natrialba magadii (Tindall et al. 1984) Kamekura et al. 1997. Synonyms: Natronobacterium magadii Tindall et al. 1984 – homotypic synonym.
- NCBI Taxonomy Browser: Natrialba magadii, Details: Natrialba magadii (Tindall et al. 1984) Kamekura et al. 1997 (species); homotypic aynonym: Natronobacterium magadii Tindall et al. 1984; type strain: ATCC:43099, DSM:3394, JCM:8861, …MS3,… .
- NCBI Taxonomy Browser: Natrialba phage PhiCh1, equivalent: Bacteriophage Phi-Ch1, … (no rank). Nucleotide: NC_054954.1: Natrialba phage PhiCh1, complete genome.
- Angela Witte, Ulrike Baranyi, Reinhard Klein, Michael Sulzner, Cheng Luo, Gerhard Wanner, Detlev H. Krüger, Werner Lubitz: Characterization of Natronobacterium magadii phage ΦCh1, a unique archaeal phage containing DNA and RNA. In: Molecular Microbiology, Band 23, Nr. 3, Januar/Februar 1997, S. 603–616; doi:10.1046/j.1365-2958.1997.d01-1879.x, PMID 9044293, ResearchGate:247656818, Epub 31. Oktober 2003 (englisch).
- NCBI Taxonomy Browser: Halobacterium phage phiH, equivalent: Bacteriophage phi-H, … (no rank). Nucleotide: txid169684[Organism:noexp]: Halobacterium phage phiH; NC_052650.1: Halobacterium phage phiH variant phiH1, complete genome.
- Mike Dyall-Smith, Felicitas Pfeifer, Angela Witte, Dieter Oesterhelt, Friedhelm Pfeiffer: Complete Genome Sequence of the Model Halovirus PhiH1 (ΦH1). In: MDPI: Genes, Band 9 Nr. 10, 12. Oktober 2018, Special Issue Genetics of Halophilic Microorganisms, S. 493; doi:10.3390/genes9100493, PMID 30322017, PMC 6210493 (freier Volltext) (englisch).
- M. Madigan, J. Martinko (Hrsg.): Brock Biology of Microorganisms. 11th Auflage. Prentice Hall, 2006, ISBN 0-13-144329-1 (englisch).
- Spencer Bliven, Andreas Prlić: Circular Permutation in Proteins. In: PLoS Computational Biology, Band 8, Nr. 3, 29. März 2012, S. e1002445; doi:10.1371/journal.pcbi.1002445.
- InterPro: IPR002711: HNH endonuclease.
- Smriti Kala, Nichole Cumby, Paul D. Sadowski, Batool Zafar Hyder, Voula Kanelis, Alan R. Davidson, Karen L. Maxwell: HNH proteins are a widespread component of phage DNA packaging machines. In: PNAS, Band 111, Nr. 16, 22. April 2014, S. 6022–6027; doi:10.1073/pnas.1320952111, PMID 24711378, PMC 4000778 (freier Volltext), Epub 7. April 2014 (englisch).
- ICTV: Taxonomy Browser.
- NCBI Taxonomy Browser: Vertoviridae.
- SIB: Myohalovirus. Auf: ViralZone.
- Reinhard Klein, Ulrike Baranyi, N. Rössler, Barbara Greineder, H. Scholz, Angela Witte: Natrialba magadii virus φCh1: first complete nucleotide sequence and functional organization of a virus infecting a haloalkaliphilic archaeon. In: Molecular Microbiology, Band 45, Nr. 3, 25. Juli 2002, S. 851–863; doi:10.1046/j.1365-2958.2002.03064.x (englisch).
- M. Iro, Reinhard Klein, B. Galos, Ulrike Baranyi, N. Rössler, Angela Witte: The lysogenic region of virus φCh1: identification of a repressor-operator system and determination of its activity in halophilic Archaea. In: Extremophiles, Band 11, 19. Februar 2007, S. 383–396; doi:10.1007/s00792-006-0040-3, Epub 23. November 2006 (englisch).
- Ana Senčilo, Elina Roine: A Glimpse of the genomic diversity of haloarchaeal tailed viruses. In: Frontiers in Microbiology, Band 5, Nr. 84, 12. März 2014; Reihe: Proceedings of Halophiles 2013: The international congress on halophilic microorganisms; doi:10.3389/fmicb.2014.00084, PMC 3950731 (freier Volltext), PMID 24659986 (englisch).
- verto. Lateinwörterbuch (frag-caesar.de).
- Stephen T. Abedon, Kelly L. Murray: Archaeal viruses, not archaeal phages: an archaeological dig. In: Archaea. 2013. Jahrgang, 7. April 2013, S. 251245, doi:10.1155/2013/251245, PMID 23653528, PMC 3638648 (freier Volltext) – (englisch).
- Terje Torsvik, Ian D. Dundas: Bacteriophage of Halobacterium salinarium. In: Nature, Band 248, S. 680–681, 19. April 1974; doi:10.1038/248680a0 (englisch).
- Sen-Lin Tang, Stewart Nuttall, Mike Dyall-Smith: Haloviruses HF1 and HF2: evidence for a recent and large recombination event. In: J Bacteriol. 186. Jahrgang, Nr. 9, Mai 2004, S. 2810–2817, doi:10.1128/JB.186.9.2810-2817.2004, PMID 15090523, PMC 387818 (freier Volltext) – (englisch).
- Tatiana A. Demina, Hanna M. Oksanen: Euryarchaeal Viruses. In: Academic Press: Encyclopedia of Virology, 4. Auflage, 2021, Band 4, S. 368–379; doi:10.1016/B978-0-12-809633-8.20989-8, Abstract (englisch).
- Terje Torsvik, Ian D. Dundas: Persisting Phage Infection in Halobacterium salinarium str. 1. In: Journal of General Virology, Band 47, Nr. 1, 1. März 1980; doi:10.1099/0022-1317-47-1-29 (englisch).
- NCBI Taxonomy Browser: Gordonia phage Kita (no rank). Nymphadoravirus kita (Nymbaxtervirinae).
- Welkin H. Pope, Travis N. Mavrich, Rebecca A. Garlena, Carlos A. Guerrero-Bustamante, Deborah Jacobs-Sera, Matthew T. Montgomery, Daniel A. Russell, Marcie H. Warner, Graham F. Hatfull, et al.: Bacteriophages of Gordonia spp. Display a Spectrum of Diversity and Genetic Relationships., In: ASM Journals: mBio, Band 8, Nr. 4, 15. August 2017, S. e01069-17; doi:10.1128/mBio.01069-17, PMID 28811342, PMC 5559632 (freier Volltext) (englisch). Siehe insbes. Fig. 4
- NCBI Taxonomy Browser: Xylella phage Xfas53, equivalent: Xylella fastidiosa phage Xfas53 (species).
- Elizabeth J. Summer, Christopher J. Enderle, Stephen J. Ahern, Jason J. Gill, Cruz P. Torres, David N. Appel, Mark C. Black, Ry Young, Carlos F. Gonzalez: Genomic and biological analysis of phage Xfas53 and related prophages of Xylella fastidiosa. In: ASM Journals: Journal of Bacteriology, Band 192, Nr. 1, Januar 2010, S. 179–190; doi:10.1128/JB.01174-09, PMID 19897657, PMC 2798268 (freier Volltext) (englisch).
- NCBI Taxonomy Browser: Enterobacteria phage phi80, equivalent: Enterobacteria phage Lula (species).
- Julianne H. Grose, Sherwood R. Casjens: Understanding the enormous diversity of bacteriophages: the tailed phages that infect the bacterial family Enterobacteriaceae. In: Virology, Band 468–470, November 2014, S. 421–443; doi:10.1016/j.virol.2014.08.024, PMID 25240328, PMC 4301999 (freier Volltext) (englisch).
- NCBI Taxonomy Browser: Salmonella typhimurium phage Gifsy-2 (species).
Anm.: Der Phage hat (im Wesentlichen) den Morphotyp der Siphoviren[35] und gehört damit zu den Caudoviricetes.