Velociraptor
Velociraptor (lat. velox ‚schnell‘, raptor ‚Räuber‘[2]) ist eine Gattung theropoder Dinosaurier, die vor etwa 85 bis 76 Millionen Jahren in der späten Kreidezeit lebte.[3] Velociraptor zählt zu den Dromaeosauridae. Momentan werden zwei Arten anerkannt: Die Typusart Velociraptor mongoliensis, deren fossile Knochen in der Mongolei und der Inneren Mongolei (China) gefunden wurden, sowie die 2008 beschriebene, bisher nur durch Schädelmaterial aus der Inneren Mongolei bekannte Art Velociraptor osmolskae.
| Velociraptor | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelettrekonstruktion von Velociraptor | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oberkreide (spätes Santonium bis mittleres Campanium)[1] | ||||||||||||
| 85,2 bis 76,4 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Velociraptor | ||||||||||||
| Osborn, 1924 | ||||||||||||
| Arten | ||||||||||||
| ||||||||||||
Mit über einem Dutzend entdeckten fossilen Skeletten ist Velociraptor einer der am besten untersuchten Dromaeosauriden. Ein besonders populäres Fossil zeigt Velociraptor mit einem Protoceratops; beide Tiere starben augenscheinlich während eines Kampfes.
Mit einer Länge von bis zu zwei Metern war Velociraptor ein mittelgroßer Vertreter der Dromaeosauridae. Es handelte sich um einen auf zwei Beinen laufenden, gefiederten Fleischfresser mit einem langen, steifen Schwanz und einer auffällig vergrößerten, sichelartigen Kralle an der zweiten Zehe des Hinterfußes. Diese Sichelkralle wurde vermutlich bei der Jagd verwendet. Ein Unterscheidungsmerkmal zu anderen Dromaeosauriden ist die lange, flache und leicht nach oben gebogene Schnauze.
Der breiten Öffentlichkeit bekannt geworden ist Velociraptor insbesondere wegen seiner von künstlerischer Freiheit geprägten Rolle als intelligentes menschenfressendes Raubtier in der Jurassic-Park-Filmreihe, auch wenn dort die Größe und Form der Tiere eher der von Deinonychus und Utahraptor entsprechen.
Merkmale


Velociraptor war ein mittelgroßer Dromaeosauride mit einer Länge von bis zu 2,07 Metern, einer Hüfthöhe von 50 Zentimetern und einem geschätzten Gewicht von bis zu 15 Kilogramm.[4] Der Schädel war bis zu 25 Zentimeter lang und wies eine im Vergleich zu anderen Dromaeosauriden dünne, flache und lange Schnauze auf: So machte die Schnauze 60 % der Gesamtlänge des Schädels aus, zeigte von oben betrachtet jedoch nur ein Drittel der Breite des hinteren Schädels. Der vordere Abschnitt der Schnauze war leicht nach oben gebogen, was zu einer konkaven Schnauzenoberseite und einer konvexen Unterseite führte. Die Augen waren nach vorne gerichtet, was ein gutes räumliches Sehen ermöglichte. Sein räumliches Blickfeld war etwa 55° breit und 10° hoch; das sind mit Eulen vergleichbare Werte.[5] Jede Kieferseite war mit 26 bis 28 in größeren Abständen stehenden Zähnen besetzt, welche an der hinteren Schneidekante stärker gesägt waren als an der vorderen – das kann als Anpassung an das Greifen schneller Beute gedeutet werden.[2][6]
Das Handskelett ähnelte in seiner Struktur und Flexibilität dem der Vögel und zeigte drei stark gekrümmte Krallen. Der zweite Finger war der längste Finger, der erste Finger der kürzeste. Aufgrund der Struktur der Handwurzelknochen war die Pronation (Einwärtsdrehung) des Handgelenks unmöglich. Dies zwang Velociraptor dazu, seine Hände stets mit der Handflächenseite nach innen zu tragen.[7] Velociraptor lief wie andere Dromaeosauriden ausschließlich auf dem dritten und vierten Zeh. Der erste Zeh war wie bei anderen Theropoden reduziert („Wolfskralle“) und berührte den Boden nicht. Auch der stark modifizierte zweite Zeh wurde über dem Boden gehalten: Er trug eine auffällig große, bis zu 6,5 Zentimeter lange, sichelförmige Kralle, ein typisches Merkmal der Dromaeosauridae. Sie wurde vermutlich als Waffe bei der Jagd eingesetzt.[8][9] Neuere biomechanische Studien zeigen auf, dass sie auch zum Klettern geeignet war.[10]
Extrem verlängerte Knochenfortsätze zur Verbindung der Wirbel (Präzygapophysen) an den Wirbelbögen, verlängerte Chevron-Knochen an der Unterseite der Wirbel sowie verknöcherte Sehnen versteiften den Schwanz des Velociraptor. Die Präzygapophysen waren ab dem zehnten Schwanzwirbel ausgebildet und erstreckten sich in rutenartigen Bündeln, in Abhängigkeit von ihrer Position vier bis zehn Wirbel überspringend, nach vorn. Diese Versteifung schränkte vertikale Bewegungen des Schwanzes stark ein, auch wenn ein Fund mit „S“-förmig gekurvtem Schwanz mit intakten Präzygapophysen vermuten lässt, dass der Schwanz eine beträchtliche horizontale Beweglichkeit hatte. Die Anpassungen des Schwanzes waren vermutlich vorteilhaft, um in Kurven mit hoher Geschwindigkeit Balance und Stabilität zu wahren.[8][9] Eine biomechanische Studie errechnete eine Maximalgeschwindigkeit von 38,9 Kilometern pro Stunde.[11]
Im Jahre 2007 wurden an einem gut erhaltenen Vorderarmknochen von Velociraptor Ansätze für Federkiele gefunden, womit eine Befiederung bei Velociraptor bestätigt ist.[12]
Geschichte


Eine Expedition des American Museum of Natural History (AMNH) in die mongolische Gobi-Wüste entdeckte im Jahr 1922 das erste bekannte Velociraptor-Fossil: Einen zerdrückten, aber kompletten Schädel zusammen mit einer der vergrößerten Krallen des zweiten Zehs und Fragmenten der Fußknochen (Exemplarnummer AMNH 6515). 1924 erfolgte die Erstbeschreibung durch den Museumsleiter Henry Fairfield Osborn als Velociraptor mongoliensis.[2] Der Gattungsname Velociraptor bedeutet so viel wie „schneller Räuber“ (lat.: velox – „schnell“, raptor – „Räuber“), während das Art-Epitheton mongoliensis auf das Land Mongolei hinweist, wo die Fossilien gefunden wurden. Früher im selben Jahr nannte Osborn die neue Spezies in einem Presseartikel „Ovoraptor djadochtari“ (nicht zu verwechseln mit Oviraptor); da dieser Name jedoch weder in einem wissenschaftlichen Magazin noch durch eine formale Beschreibung veröffentlicht wurde, gilt er nach den Internationalen Regeln für die Zoologische Nomenklatur (ICZN) als Nomen nudum („nackter Name“), weshalb der Name Velociraptor Priorität genießt.[13]
Während nordamerikanischen Forschern im Kalten Krieg keine Einreise in die kommunistische Mongolei gestattet war, entdeckten sowjetische, polnische und mongolische Forscher weitere Skelette. So wurde 1971 das berühmte Fossil der „Kämpfenden Dinosaurier“ von einer polnisch-mongolischen Forschergruppe entdeckt (Exemplarnummer GIN 100/25), welches einen Velociraptor während eines Kampfes mit einem Protoceratops zeigt.[8][14][15] Es gilt als Nationalschatz der Mongolei, wurde jedoch im Jahr 2000 dem American Museum of Natural History in New York für eine temporäre Ausstellung geliehen.[16] Heute steht es im National Museum of Natural History in Ulaanbaatar.[17][18]
Zwischen 1988 und 1990 entdeckte ein chinesisch-kanadisches Team Überreste von Velociraptor in Nordchina.[19] Amerikanische Wissenschaftler kehrten 1990 in die Mongolei zurück und so entdeckte ein mongolisch-amerikanisches Team mehrere gut erhaltene Skelette.[9][20]
1999 entdeckte ein chinesisch-belgisches Team einen Oberkiefer und ein Tränenbein, die zwar zu Velociraptor gehörten, jedoch untypisch für die Typusart Velociraptor mongoliensis sind. Forscher um Pascal Godefroit beschrieben diesen Fund 2008 als Holotypus einer neuen Art, Velociraptor osmolskae.[3] Sie unterscheidet sich von Velociraptor mongoliensis vor allem durch ein vergrößertes Promaxillarfenster, eine Schädelöffnung (Schädelfenster) im Oberkiefer, die sich mittig etwa auf gleicher Höhe zwischen Augenhöhle und Nasenöffnung befindet.[3] Das Art-Epitheton osmolskae ehrt die polnische Paläontologin Halszka Osmólska, welche kurz vor der Erstbeschreibung verstarb.[21]
Fundorte
Die meisten Fossilien von Velociraptor mongoliensis wurden in der Djadochta-Formation gefunden, sowohl im mongolischen Aimag Ömnö-Gobi als auch in der chinesischen Inneren Mongolei. Bislang noch nicht einer der beiden Arten zugeordnete Fundstücke von Velociraptor stammen auch aus der etwas jüngeren Barun-Goyot-Formation der Mongolei.[22] Diese geologischen Formationen werden auf die Oberkreide (spätes Santonium bis mittleres Campanium) datiert und sind damit etwa 85 bis 76 Millionen Jahre alt.[23][24]

Velociraptor-Funde stammen aus vielen der berühmten und ergiebigsten Fundstellen der Djadochta-Formation: So stammt das Typusexemplar aus Bajandsag oder „Flaming Cliffs“ (auch als Shabarakh Usu bekannt)[2], während die „Kämpfenden Dinosaurier“ in der Turig-Fundstelle (Tugrugeen Shireh) entdeckt wurden.[15] Auch bekannte Fundstellen der Barun-Goyot-Formation, Khulsan und Khermeen Tsav, enthielten Fossilien, die eventuell zu Velociraptor gehörten.[25] Zähne und fragmentarische Knochenreste, die juvenilen Velociraptor mongoliensis zugeschrieben werden, stammen aus der Bayan-Mahandu-Formation der Inneren Mongolei.[19] Dort wurde auch das Schädelmaterial von Velociraptor osmolskae gefunden.[3]
Paläoökologie
Alle Sedimentgesteinsschichten mit Velociraptor-Fossilien zeigen Hinweise auf ein trockenes Klima zur Zeit der Ablagerung. Die Landschaft war durch saisonal austrocknende Gewässer und Sanddünen geprägt, und das Klima glich dem der heutigen wechselfeuchten Tropen. Während der Ablagerung der jüngeren Barun-Goyot-Formation herrschte ein etwas feuchteres Klima als zur Zeit der Ablagerung der Djadochta-Formation.[24] Die Körperhaltung einiger kompletter Skelette und deren Erhaltung in strukturlosen Sandsteinablagerungen lässt vermuten, dass diese Exemplare im Verlauf der häufig auftretenden Sandstürme lebendig begraben wurden.[3] Velociraptor lebte zusammen mit Ceratopsiern wie Protoceratops, Udanoceratops, Ankylosauriden wie Pinacosaurus sowie verschiedenen Theropoden wie Oviraptoriden, Troodontiden, Alvarezsauriden und anderen Dromaeosauriden wie beispielsweise Adasaurus.[22] Viele Dinosaurier-Gattungen kommen sowohl in der Djadochta-Formation und der Barun-Goyot-Formation als auch in der Bayan-Mandahu-Formation vor, wobei die Vertreter aus verschiedenen Formationen oftmals unterschiedlichen Arten zugerechnet werden. Beispielsweise bestand die Djadochta-Fauna aus Velociraptor mongoliensis, Protoceratops andrewsi, und Pinacosaurus grangeri, während die Bayan-Mandahu-Fauna die Arten Velociraptor osmolskae, Protoceratops hellenikorhinus und Pinacosaurus mephistocephalus beinhaltete. Diese Unterschiede in der Artzusammensetzung könnten auf eine natürliche Barriere, welche die zwei geographisch eng benachbarten Formationen voneinander trennte, oder auf einen leichten Unterschied in ihrem stratigraphischen Alter zurückzuführen sein.[3]
Systematik
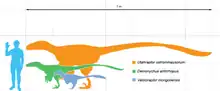
Velociraptor wird zur Unterfamilie Velociraptorinae gezählt, einer fortschrittlichen (abgeleiteten) Untergruppe der Familie Dromaeosauridae. In der phylogenetischen Systematik (Kladistik) werden die Velociraptorinae meist als „alle Dromaeosauriden, die näher mit Velociraptor als mit Dromaeosaurus verwandt sind,“ definiert. Während anfangs Velociraptor als einzige Gattung innerhalb der Velociraptorinae eingeordnet wurde, fügten einige spätere Analysen weitere Gattungen – meistens Deinonychus und Saurornitholestes – hinzu.[8][26] Die Verwandtschaftsbeziehungen innerhalb der Dromaeosauridae bleiben jedoch umstritten. So entwirft eine aktuelle kladistische Analyse von Forschern um Mark Norell (2006) eine Gattung Velociraptorinae, die Velociraptor, Deinonychus, Tsaagan sowie Saurornitholestes mit einschließt.[27] Eine weitere Analyse von Longrich und Currie (2009) ordnet den Velociraptorinae die Gattungen Velociraptor, Itemirus, Tsaagan und Adasaurus zu, wobei Itemirus als die am engsten mit Velociraptor verwandte Gattung betrachtet wird.[28] Csiki und Kollegen (2010) kommen zu dem Schluss, dass der erst kürzlich entdeckte Balaur der engste Verwandte von Velociraptor war.[29]
Es folgt ein aktuelles Klassifikationsbeispiel (vereinfacht nach Longrich und Currie, 2009):[28]
| Dromaeosauridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
In der Vergangenheit vermuteten einige Forscher, dass es sich bei einigen anderen Dromaeosauriden wie Deinonychus antirrhopus und Saurornitholestes langstoni um Arten von Velociraptor handelte. Demnach wären die Gattungen Deinonychus und Saurornitholestes mit Velociraptor identisch gewesen – da jedoch Velociraptor der zuerst vergebene Name ist, behält er Priorität. Somit wurden diese Arten in Velociraptor antirrhopus und Velociraptor langstoni umbenannt.[4] Heute sind lediglich Velociraptor mongoliensis[6][7][30] und Velociraptor osmolskae[3] anerkannte Arten von Velociraptor.
Bei der Erstbeschreibung im Jahr 1924 wurde Velociraptor in die Familie Megalosauridae gestellt, der damals fast alle bekannten fleischfressenden Dinosaurier zugeschrieben wurden.[2] Die Megalosauridae und insbesondere Megalosaurus wurden lange als sogenannte „Papierkorb-Taxa“ verwendet, denen eine Vielzahl von Funden offensichtlich nicht näher miteinander verwandter Theropoden zugeschrieben wurden (polyphyletisch). In der Folgezeit schrieben verschiedene Autoren Velociraptor unter anderem den Compsognathidae und den Coeluridae zu.[31] Erst seit 1969 wird Velociraptor zusammen mit Dromaeosaurus und Deinonychus innerhalb der Familie Dromaeosauridae klassifiziert.[32]
Die Dromaeosauridae bilden zusammen mit den Troodontidae die Gruppe Deinonychosauria, welche wiederum zu den Maniraptora gezählt wird. Häufig werden die Deinonychosauria als Schwestergruppe der Vögel betrachtet.[33][34] Einige Forscher vermuten gar, dass die Dromaeosauridae die Vögel als Teilgruppe einschließt, dass also Vögel von urtümlichen Mitgliedern der Dromaeosauridae, die noch nicht den Vögel zuzurechnen sind („Nichtvogeldromaeosauriden“), abstammen.[35] Verschiedene Forscher vermuten, dass es sich bei den Dromaeosauriden inklusive Velociraptor tatsächlich um eine Gruppe innerhalb der Vögel handelte, die sekundär flugunfähig geworden ist.[36][7]
Paläobiologie

Jagdverhalten
Die charakteristische Sichelkralle des zweiten Zehs der Dromaeosauriden wurde in der Vergangenheit häufig als Vorrichtung zum Aufschlitzen großer Beutetiere interpretiert.[37] Manning und Kollegen (2005) führten Versuche mit einem hydraulischen Robotermodell des Fußes von Deinonychus an der Bauchdecke eines Schweins durch und zeigten, dass die Sichelkralle der Dromaeosauriden lediglich kleine und rundliche Einstichlöcher hinterließ, jedoch nicht in der Lage war, die Bauchdecke aufzuschlitzen. Diese Forscher vermuten, dass die Sichelkralle als eine Art Steigeisen fungiert haben könnte, mit der sich die Dromaeosauriden auf großen Beutetieren festhalten konnten, während Hände und Zähne dem Beutetier zahlreiche Verletzungen zuführten.[38]
.jpg.webp)
Direkte Hinweise auf das Jagdverhalten von Velociraptor gibt das Fossil der „kämpfenden Dinosaurier“, welches einen Velociraptor mongoliensis inmitten eines Kampfes mit einem Protoceratops andrewsi zeigt: So liegt die linke Hand von Velociraptor direkt hinter dem Wangenhorn von Protoceratops, während letzterer den rechten Unterarm von Velociraptor mit seinem Schnabel festhält. Der rechte Fuß des Velociraptor scheint unter dem liegenden Körper des Protoceratops eingeschlossen zu sein. Der linke Fuß ist mit nach unten gerichteter Sichelkralle in die Halsregion des Protoceratops ausgestreckt. Das lässt Carpenter (1998) vermuten, dass die Sichelkralle nicht zum Aufschlitzen, sondern zum gezielten Durchstechen der Luftröhre oder der Halsadern eingesetzt wurde.[39]
Die Sedimentstrukturen, in denen die beiden Skelette erhalten sind, wurden ursprünglich als Ablagerungen eines Flussdeltas interpretiert, woraus Barsbold (1974) folgerte, dass die Tiere ertranken.[15] Spätere Untersuchungen zeigten jedoch, dass es sich um äolische (durch Wind entstandene) Ablagerungen von Sanddünen handelte. So wurden die Tiere möglicherweise während des Kampfes plötzlich von Sand lebend begraben – entweder von einer kollabierenden Düne oder von einem Sandsturm. Carpenter (1998) merkt jedoch an, dass sowohl Vorder- als auch Hinterbeine des Protoceratops-Exemplars fehlen, was der Hypothese eines Lebendbegräbnisses widerspricht. Zur Erklärung dieses Befundes schlägt er ein anderes Szenario vor: So könnte der Velociraptor den Protoceratops getötet haben, wobei sein rechtes Bein unter dem Kadaver des Protoceratops eingeklemmt wurde, so dass der Velociraptor sich nicht mehr befreien konnte und starb. Aasfresser könnten die Beine des Protoceratops fortgetragen haben, bevor beide Kadaver von Flugsand bedeckt wurden.[39]

Der verwandte Dromaeosauride Deinonychus könnte in Gruppen gelebt und gejagt haben. Entsprechende Hinweise stammen aus zwei Steinbrüchen, die jeweils die Überreste mehrerer Deinonychus zusammen mit den Überresten des großen Pflanzenfressers Tenontosaurus beherbergten.[40][41] Den einzigen unzweifelhaften Hinweis auf ein Leben in Gruppen bei Dromaeosauriden stellen sechs nebeneinander verlaufende, versteinerte Fußspuren einer größeren Dromaeosauriden-Spezies aus China dar; Hinweise auf ein Jagen in Gruppen bietet dieser Fund jedoch nicht.[42] Obwohl viele isoliert vorgefundene Velociraptor-Funde bekannt sind, wurde kein Individuum in Verbindung mit einem anderen Individuum gefunden. Somit gibt es nur begrenzte Nachweise für ein Leben in Gruppen bei Dromaeosauriden und keinen einzigen für Velociraptor spezifischen Nachweis, obwohl Velociraptor in den populären Medien, beispielsweise im Film Jurassic Park, regelmäßig als in Gruppen lebend und jagend dargestellt wird.[30]
Nekrophagie
2008 fanden Paläontologen in der Bayan-Mahandu-Formation Zähne, die vermutlich zu Velociraptor gehören, um einen mit Bissspuren versehenen Kieferknochen, der vermutlich von Protoceratops stammt.[43] Dieser Fund wird als Beweis für den Konsum von Aas (Nekrophagie) durch Velociraptor gedeutet, da dieser von einem frisch getöteten Protoceratops zuerst andere Körperteile gefressen hätte.[43][44] Ebenso wurde bestätigt, dass Protoceratops eines der Beutetiere von Velociraptor war.[43]
Stoffwechsel
Velociraptor war wohl bis zu einem gewissen Grad warmblütig (endotherm), die Wachstumsrate der Knochen von Dromaeosauriden deutet jedoch auf einen im Vergleich zu Säugetieren und Vögeln geringeren Stoffwechsel (Metabolismus) hin.[7] Weitere Hinweise auf Endothermie geben der Bau des Atmungsapparats und zusätzlich die Tatsache, dass ein wechselwarmer (ektothermer) Stoffwechsel wohl nicht in der Lage war, die schnelle, bipede Fortbewegung eines Velociraptor über einen längeren Zeitraum zu unterstützen.[45]
Federn

Fossilien von ursprünglichen Dromaeosauriden zeigen eine umfangreiche Befiederung sowie voll entwickelte Flügel[46], was für die Paläontologen Anlass zu Untersuchungen war, ob auch Velociraptor gefiedert gewesen sein könnte. 2007 wurden an einer Elle von Velociraptor Ansätze für zwölf Federkiele der Armschwingen nachgewiesen, womit eine Befiederung letztendlich bestätigt wurde.[12]
Die Federn könnten bei einer Form des Ausdrucksverhaltens etwa vor der Paarung eine Rolle gespielt oder das Ausbrüten der Eier wie bei den heutigen Vögeln unterstützt haben. Auch die Stabilisierung des Laufens bei hoher Geschwindigkeit könnte einen Vorteil gebracht und die Evolution des Gefieders gefördert haben.[12]
Velociraptor in der Populärkultur
Velociraptor wurde bekannt durch seine Rolle als grausamer Jäger im Roman DinoPark von Michael Crichton und in dessen Verfilmung Jurassic Park durch den Regisseur Steven Spielberg. Die „Raptoren“ nehmen die Rollen der Hauptantagonisten ein, welche die menschlichen Protagonisten durch einen Dinosaurierzoo jagen. Sie wurden ursprünglich an Deinonychus angelehnt, einen größeren Verwandten von Velociraptor, der damals vom Paläontologen Gregory S. Paul als Velociraptor antirrhopus bezeichnet wurde.[4] Eine Figur in Michael Crichtons Roman sagt ebenfalls, dass „…Deinonychus zu den Velociraptoren“ gehöre. Das zeigt, dass Crichton die Klassifizierung von Gregory Paul nutzte, auch wenn sie durchgehend als Velociraptor mongoliensis bezeichnet werden.[47]
Aus dramaturgischen Gründen wurden die „Raptoren“ im Film deutlich vergrößert und die Form der Schnauze verändert.[48][49] Ebenso wurden Bau und Stellung der Arme verändert, und die Schwänze sehr kurz und flexibel dargestellt, was klar der Anatomie der Fossilien widerspricht. Ebenso auffällig ist die fehlende Befiederung in Roman und Film, obwohl diese laut des wissenschaftlichen Beraters des Films Jack Horner bereits bekannt war. Er gibt als Grund die damals noch limitierten Möglichkeiten der Computertechnologie und dramaturgische Gründe für die Veränderung an.[50] In Jurassic Park III wurden die „Raptoren“ mit Federn am Kopf und im Nacken dargestellt, welche nicht der eigentlichen Befiederung von Dromaeosauriden entsprechen.[12] In einer Szene in Jurassic Park III wird ebenfalls erwähnt, dass die Intelligenz der „Raptoren“ diejenige von Delfinen, Walen und einigen Primaten übersteige. Dies ist jedoch aufgrund des Gehirnvolumens höchst unwahrscheinlich.[51]
Durch den Erfolg von Jurassic Park wurde Velociraptor einer der bekanntesten Dinosaurier der Populärkultur. Er kommt in vielen Spielzeugsortimenten, Videospielen, Fernsehserien, Filmen und Dokumentationen vor. 2011 brachte die britische Indierockband Kasabian ein Album unter dem Titel Velociraptor heraus, auf dem Gitarrist Sergio Pizzorno in aggressiver Haltung mit Federn abgebildet ist, so wie sie dem Saurier zugeschrieben werden. Auf dem Album befindet sich ein Stück desselben Namens.[52]
Ferner dient Velociraptor der 1995 gegründeten Profi-Basketballmannschaft von Toronto, den Toronto Raptors, als Namensvorbild. Zudem ist das Maskottchen, das ursprünglich auch das Teamlogo zierte, offensichtlich an Velociraptor angelehnt.
In der Zeichentrick-Serie Extreme Dinosaurs von 1997 sind die drei Hauptantagonisten Velociraptoren. Sie und die anderen in der Serie vorkommenden Dinosaurier sind humanoid bzw. anthropomorph, indem sie z. B. alle ungefähr gleich groß und nur wenig größer als Menschen sind, Kleidung tragen und futuristische Werkzeuge und Fortbewegungsmittel nutzen.
Im US-amerikanischen Freizeitpark Island of Adventure basiert eine Achterbahn auf Velociraptoren. Die Bahn hört auf dem Namen VelociCoaster und handelt von dem Film Jurassic World.
Das erste befiederte Modell eines Velociraptors ist in der Ausstellung Dinosaurier – Die Urzeit lebt! im LWL-Museum für Naturkunde in Münster zu sehen.[53]
Weblinks
Einzelnachweise
- Gregory S. Paul: The Princeton Field Guide To Dinosaurs. Princeton University Press, Princeton NJ u. a. 2010, ISBN 978-0-691-13720-9, S. 137, Online.
- Henry Fairfield Osborn: Three new Theropoda, Protoceratops zone, central Mongolia (= American Museum Novitates. Nr. 144, ISSN 0003-0082 = Publications of the Asiatic Expeditions of the American Museum of Natural History. Contribution. Nr. 32). The American Museum of Natural History, New York NY 1924, online.
- Pascal Godefroit, Philip J. Currie, Li Hong, Shang Chang Yong, Dong Zhi-Ming: A new species of Velociraptor (Dinosauria: Dromaeosauridae) from the Upper Cretaceous of northern China. Bd. 28, Nr. 2, 2008, ISSN 0272-4634, S. 432–438, doi:10.1671/0272-4634(2008)28[432:ANSOVD]2.0.CO;2.
- Gregory S. Paul: Predatory Dinosaurs of the World. A complete and illustrated Guide. Simon and Schuster, New York NY u. a. 1988, ISBN 0-671-61946-2, S. 464 ff.
- Kent A. Stevens: Binocular vision in theropod dinosaurs. In: Journal of Vertebrate Paleontology. Bd. 26, Nr. 2, 2006, S. 321–330, doi:10.1671/0272-4634(2006)26[321:BVITD]2.0.CO;2.
- Rinchen Barsbold, Halszka Osmólska: The skull of Velociraptor (Theropoda) from the Late Cretaceous of Mongolia. In: Acta Palaeontologica Polonica. Bd. 44, Nr. 2, 1999, ISSN 0567-7920, S. 189–219, online.
- Gregory S. Paul: Dinosaurs of the Air. The Evolution and Loss of Flight in Dinosaurs and Birds. Johns Hopkins University Press, Baltimore MD u. a. 2002, ISBN 978-0-8018-6763-7.
- Ринченгийн Барсболд: Хищные динозавры мела Монголии. Совместная Советско-Монгольская палеонтологическая экспедиция (= Труды. 19). Наука, Москва 1983, S. 5–119 (In englischer Sprache: Rinchen Barsbold: Carnivorous dinosaurs from the Cretaceous of Mongolia. The Joint Soviet-Mongolian Paleontological Expedition (= Transaction of the Joint Soviet-Mongolian Paleontological Expedition. Bd. 19). Selbstverlag des Verfassers, Berkeley CA 1983, 5–119, online).
- Mark A. Norell, Peter J. Makovicky: Important features of the dromaeosaurid skeleton II: information from newly collected specimens of Velociraptor mongoliensis (= American Museum Novitates. Nr. 3282). The American Museum of Natural History, New York NY 1999, online.
- Phillip L. Manning, Lee Margetts, Mark R. Johnson, Philip J. Withers, William I. Sellers, Peter L. Falkingham, Paul M. Mummery, Paul M. Barrett, David R. Raymont: Biomechanics of Dromaeosaurid Dinosaur Claws: Application of X-Ray Microtomography, Nanoindentation, and Finite Element Analysis. In: The Anatomical Record. Bd. 292, Nr. 9, 2009, ISSN 1932-8486, S. 1397–1405, doi:10.1002/ar.20986.
- William Irvin Sellers, Phillip Lars Manning: Estimating dinosaur maximum running speeds using evolutionary robotics. In: Proceedings of the Royal Society. Series B: Biological Sciences. Bd. 274, Nr. 1626, 2007, ISSN 0950-1193, S. 2711–2716, doi:10.1098/rspb.2007.0846.
- Alan H. Turner, Peter J. Makovicky, Mark A. Norell: Feather Quill Knobs in the Dinosaur Velociraptor. Bd. 317, Nr. 5845, 2007, S. 1721, doi:10.1126/science.1145076, PMID 17885130.
- Henry Fairfield Osborn: The Discovery of an Unknown Continent. In: Natural History. Bd. 24, Nr. 2, 1924, ISSN 0028-0712, S. 133–149, Digitalisat.
- Zofia Kielan-Jaworowska, Rinchen Barsbold: Narrative of the Polish-Mongolian Paleontological Expeditions. In: Palaeontologia Polonica. Bd. 27, 1972, ISSN 0078-8562, S. 5–13.
- Rinchen Barsbold: Saurornithoididae, a new family of theropod dinosaurs from Central Asia and North America. In: Palaeontologia Polonica. Bd. 30, 1974, ISSN 0078-8562, S. 5–22.
- American Museum of Natural History: Fighting Dinosaurs: New Discoveries from Mongolia: Exhibition Highlights. 29. Oktober 2000, abgerufen am 1. August 2014.
- Ch. Sumiyabazar: Mongolia's fighting dinosaurs in der UB-Post (englischsprachige, mongolische Zeitschrift) vom 14. Februar 2006
- Foto der Fighting Dinosaurs aus dem National Museum of National History
- Tomasz Jerzykiewicz, Philip J. Currie, David A. Eberth, Paul A. Johnston, Emlyn H. Koster, Jia-Jian Zheng: Djadokhta Formation correlative strata in Chinese Inner Mongolia: an overview of the stratigraphy, sedimentary geology, and paleontology and comparisons with the type locality in the pre-Altai Gobi. In: Canadian Journal of Earth Sciences. Bd. 30, Nr. 10, 1993, ISSN 0008-4077, S. 2180–2195. doi:10.1139/e93-190.
- Mark A. Norell, Peter J. Makovicky: Important features of the dromaeosaur skeleton: information from a new specimen (= American Museum Novitates. Nr. 3215). The American Museum of Natural History, New York NY 1997, online.
- Nachruf der Society of Vertebrate Palaeontology (Memento vom 27. Oktober 2010 im Internet Archive)
- David B. Weishampel, Paul M. Barrett, Rodolfo A. Coria, Jean Le Loueff, Xing Xu, Xijin Zhao, Ashok Sahni, Emily M. P. Gomani, Christopher N. Noto: Dinosaur distribution. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 517–606.
- Felix M. Gradstein, James G. Ogg, Alan G. Smith (Hrsg.): A Geologic Time Scale 2004. Cambridge University Press, Cambridge 2005, ISBN 0-521-78142-6.
- Tomasz Jerzykiewicz, Dale A. Russell: Late Mesozoic stratigraphy and vertebrates of the Gobi Basin. In: Cretaceous Research. Bd. 12, Nr. 4, 1991, ISSN 0195-6671, S. 345–377, doi:10.1016/0195-6671(91)90015-5.
- Halszka Osmólska: Barun Goyot Formation. In: Philip J. Currie, Kevin Padian (Hrsg.): Encyclopedia of Dinosaurs. Academic Press, San Diego CA u. a. 1997, ISBN 0-12-226810-5, S. 41.
- Philip J. Currie: New information on the anatomy and relationships of Dromaeosaurus albertensis (Dinosauria: Theropoda). In: Journal of Vertebrate Paleontology. Bd. 15, Nr. 3, 1995, S. 576–591, doi:10.1080/02724634.1995.10011250.
- James M. Clark, Alan H. Turner, Peter J. Makovicky, Rinchen Barsbold, Timothy Rowe: A new dromaeosaurid theropod from Ukhaa Tolgod (Omnogov, Mongolia) (= American Museum Novitates. Nr. 3545). The American Museum of Natural History, New York NY 2006, online.
- Nicolas R. Longrich, Philip J. Currie: A microraptorine (Dinosauria – Dromaeosauridae) from the late Cretaecous of North Aermica. In: Proceedings of the National Academy of Sciences. Bd. 106, Nr. 13, 2009, S. 5002–5007, doi:10.1073/pnas.0811664106.
- Zoltán Csiki, Mátyás Vremir, Stephen L. Brusatte, Mark A. Norell: An aberrant island-dwelling theropod dinosaur from the Late Cretaceous of Romania. In: Proceedings of the National Academy of Sciences. Bd. 107, Nr. 35, 2010, S. 15357–15361, doi:10.1073/pnas.1006970107.
- Mark A. Norell, Peter J. Makovicky: Dromaeosauridae. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 196–209.
- Velociraptor. In: The Paleobiology Database. Abgerufen am 1. August 2014.
- Edwin H. Colbert, Dale A. Russell: The small Cretaceous dinosaur Dromaeosaurus (= American Museum Novitates. Nr. 2380). The American Museum of Natural History, New York NY 1969, Digitalisat.
- Sunny H. Hwang, Mark A. Norell, Ji Qiang, Gao Keqin: New specimens of Microraptor zhaoianus (Theropoda, Dromaeosauridae) from northeastern China. American Museum novitates (= American Museum Novitates. Nr. 3381). The American Museum of Natural History, New York NY 2002, Digitalisat.
- Alan H. Turner, Diego Pol, Julia A. Clarke, Gregory M. Erickson, Mark A. Norell: A Basal Dromaeosaurid and Size Evolution Preceding Avian Flight. In: Science. Bd. 317, Nr. 5843, 2007, S. 1378–1381, doi:10.1126/science.1144066.
- Mark A. Norell, James M. Clark, Peter J. Makovicky: Phylogenetic relationships among coelurosaurian theropods. In: Jacques Gauthier, Lawrence F. Gall (Hrsg.): New Perspectives on the Origin and early Evolution of Birds. Proceedings of the International Symposium in Honor of John H. Ostrom. February 13–14, 1999. New Haven, Connecticut. Peabody Museum of Natural History – Yale University, New Haven CT 2001, ISBN 0-912532-57-2, S. 49–67, online (PDF; 72,38 KB).
- Larry D. Martin: A basal archosaurian origin for birds. In: Acta Zoologica Sinica. Bd. 50, Nr. 6, 2004, ISSN 0001-7302, S. 978–990.
- John H. Ostrom: Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana (= Bulletin of the Peabody Museum of Natural History. Bd. 30, ISSN 0079-032X). Peabody Museum of Natural History – Yale University, New Haven CI 1969.
- Phillip L. Manning, David Payne, John Pennicott, Paul M. Barrett, Roland A. Ennos: Dinosaur killer claws or climbing crampons? In: Biology Letters. Bd. 2, Nr. 1, 2005, ISSN 1744-9561, S. 110–112, doi:10.1098/rsbl.2005.0395.
- Ken Carpenter: Evidence of predatory behavior by theropod dinosaurs. In: Gaia. Revista de Geociências. Bd. 15, 1998, ISSN 0871-5424, S. 135–144.
- W. Desmond Maxwell, John H. Ostrom: Taphonomy and paleobiological implications of Tenontosaurus-Deinonychus associations. In: Journal of Vertebrate Paleontology. Bd. 15, Nr. 4, 1995, S. 707–712, doi:10.1080/02724634.1995.10011256.
- Daniel L. Brinkman, Richard L. Cifelli, Nicholas J. Czaplewski: First occurrence of Deinonychus antirrhopus (Dinosauria: Theropoda) from the Antlers Formation (Lower Cretaceous: Aptian-Albian) of Oklahoma (= Oklahoma Geological Survey Bulletin. Bd. 146, ISSN 0078-4389). University of Oklahoma, Norman OK 1998, Digitalisat (PDF; 2,13 MB).
- Rihui Li, Martin G. Lockley, Peter J. Makovicky, Masaki Matsukawa, Mark A. Norell, Jerald D. Harris, Mingwei Liu: Behavioral and faunal implications of Early Cretaceous deinonychosaur trackways from China. In: Die Naturwissenschaften. Bd. 95, Nr. 3, 2008, S. 185–191, doi:10.1007/s00114-007-0310-7, PMID 17952398.
- David Hone, Jonah Choiniere, Corwin Sullivan, Xing Xu, Michael Pittman, Qingwei Tan: New evidence for a trophic relationship between the dinosaurs Velociraptor and Protoceratops. In: Palaeogeography, Palaeoclimatology, Palaeoecology. Bd. 291, Nr. 3/4, 2010, ISSN 0031-0182, S. 488–492, doi:10.1016/j.palaeo.2010.03.028.
- Matt Walker: Fossil find shows Velociraptor eating another dinosaur In: BBC Earth News, 6. April 2010. Abgerufen am 1. August 2014
- Herman Pontzer, Vivian Allen, John R. Hutchinson: Biomechanics of Running Indicates Endothermy in Bipedal Dinosaurs. In: PLOS ONE. Bd. 4, Nr. 11, 2009, e7783, doi:10.1371/journal.pone.0007783, (Dazu: Corrections. In: PLoS One. Bd. 4, Nr. 12, 2009, doi:10.1371/annotation/635e46fc-4be3-4f42-ad5c-ee3a276cd24f).
- Xing Xu, Zhonghe Zhou, Xiaolin Wang, Xuewen Kuang, Fucheng Zhang, Xiangke Du: Four-winged dinosaurs from China. In: Nature. Bd. 421, Nr. 6921, 2003, S. 335–340, doi:10.1038/nature01342, PMID 12540892.
- Michael Crichton: Jurassic Park. A Novel. Alfred A. Knopf, New York, NY 1990, ISBN 0-394-58816-9, S. 117.
- Jody Duncan: The Winston Effect. The Art and History of Stan Winston Studio. Titan Books, London 2006, ISBN 1-84576-365-3, S. 175.
- Robert T. Bakker: Raptor Red. Bantam Books, New York NY 1995, ISBN 0-553-57561-9, S. 4.
- Paläontologe über „Jurassic Park“: „Hauptgegner war die Dramaturgie“. Abgerufen am 8. Mai 2023.
- Hans C. E. Larsson, Paul C. Sereno, Jeffrey A. Wilson: Forebrain enlargement among nonavian theropod dinosaurs. In: Journal of Vertebrate Paleontology. Bd. 20, Nr. 3, 2000, S. 615–618, doi:10.1671/0272-4634(2000)020[0615:FEANTD]2.0.CO;2.
- Kasabian – The official site for news about the band Kasabian (Memento vom 8. März 2012 im Internet Archive).
- Erstes befiedertes Velociraptor-Modell in Deutschland - Mitteilung 19.08.14. Abgerufen am 30. Januar 2023.