Japanische Spitzmulle
Die Japanischen Spitzmulle (Urotrichini) sind eine Gattungsgruppe aus der Familie der Maulwürfe (Talpidae). Sie stellen zwei Arten, die in Japan endemisch verbreitet sind. Es handelt sich um kleine Vertreter der Maulwürfe, die äußerlich mit ihrer langen Schnauze und dem langen Schwanz eher an Spitzmäuse erinnern. Die Gliedmaßen sind deutlich schlank gestaltet und eignen sich weniger gut zum Graben. Dementsprechend verbringen die Tiere einen Teil ihrer aktiven Zeit an der Erdoberfläche. Dennoch graben sie teilweise oberflächennahe Tunnel und Gänge. Die Nahrung setzt sich aus Insekten und Regenwürmern zusammen. Die Fortpflanzung erfolgt überwiegend im Frühjahr. Häufig nutzen die Tiere Laub- und Nadelwälder sowie teils offene Landschaften in Tief- und höheren Gebirgslagen. Die Tribus der Japanischen Spitzmulle erhielt im Jahr 1883 ihre wissenschaftliche Definition, bestand aber damals noch aus weiteren Mitgliedern. Erst im Verlauf des 20. Jahrhunderts wurde sie auf die beiden japanischen Arten reduziert. Teilweise gliederte man sie auch in die Neuweltmaulwürfe mit ein. Genetische Analysen zeigten jedoch auf, dass es sich bei den Japanischen Spitzmullen um eine eigenständige Gruppe handelt, die mit anderen Spitzmullen Ostasiens und Nordamerikas näher verwandt ist. Die Fossilgeschichte der Japanischen Spitzmulle ist recht umfangreich und reicht mit mehreren ausgestorbenen Formen bis in den Übergang vom Eozän zum Oligozän vor rund 34 Millionen Jahren zurück. Der Großteil des Fundmaterials stammt aus Europa und den angrenzenden Gebieten Asiens. Erst im Mittelpleistozän lassen sich die heutigen Arten in Japan fassen.
| Japanische Spitzmulle | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Japanischer Spitzmull (Urotrichus talpoides) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Urotrichini | ||||||||||||
| Dobson, 1883 |
Merkmale
Habitus


Die Japanischen Spitzmulle repräsentieren kleine Vertreter der Maulwürfe. Ihre Kopf-Rumpf-Länge reicht von 7,0 bis 8,4 cm beim True-Spitzmull (Dymecodon pilirostris) bis zu 8,9 bis 10,4 cm beim Japanischen Spitzmull (Urotrichus talpoides). Der Schwanz wird 2,7 bis 4,4 cm lang. Er nimmt bei ersterer Form 43 bis 60 % der Länge des restlichen Körpers ein und ist dadurch verhältnismäßig länger als bei letzterer, bei der der Schwanz nur 34 bis 39 % ausmacht. Das Gewicht variiert insgesamt zwischen 8 und 28,5 g. Im Körperbau weisen die Japanischen Spitzmulle Ähnlichkeiten mit dem der Spitzmäuse auf. Sie heben sich dadurch von den grabenden Maulwürfen durch eine verlängerte Schnauze und den vergleichsweise langen und zudem dicken Schwanz hervor. Die Vorder- und Hinterfüße sind schmal, die vorderen haben nur kleine, wenig gebogene Grabkrallen. Die Gliedmaßen sind relativ lang und können abweichend von den grabenden Maulwürfen unter den Körper gestellt werden. Übereinstimmend mit den anderen Maulwürfen sind die Augen sehr klein, während die Ohren im Fell verborgen bleiben. Das Fell selbst ist dicht, allerdings weniger samtartig als das anderer Maulwurfsarten. Es kann eine grauschwarze, schwärzlich braune bis hin zu einer kastanienbraunen Farbgebung aufweisen. Lange Leithaare machen rund 1 bis 1,3 % des Fells auf und treten dadurch häufiger auf als bei den Eigentlichen Maulwürfen (Talpini).[1][2][3]
Schädel- und Gebissmerkmale
Der Schädel der Japanischen Spitzmulle ist wie bei den anderen Maulwürfen auch konisch gestaltet mit einem relativ breiten Hirnschädel und einem spitz zulaufenden Rostrum. Der Bereich an der Orbita zeigt sich zumeist mehr oder weniger deutlich eingeschnürt. Allgemein ist der Schädel leicht gebaut.[2] Am Mittelohr treten keine Spezialisierungen in Form vergrößerter Gehörknöchelchen auf. Höchstwahrscheinlich steht dies mit einer Wahrnehmung von Tönen im höheren Frequenzbereich in Verbindung.[4][5]
| Gattung | Zahnformel | Zahnanzahl |
|---|---|---|
| Dymecodon | oder | 38 |
| Urotrichus | oder | 36 |
| Neurotrichus | 36 | |
| Scaptonyx | 42 | |






Das Gebiss der Japanischen Maulwürfe ist stärker reduziert als im Vergleich zu den Eigentlichen Maulwürfen (Talpini) oder einigen Formen der Neuweltmaulwürfe (Scalopini). Es besteht beim True-Spitzmull aus 38, beim Japanischen Spitzmull aus 36 Zähnen. Gegenüber ersterer Art hat sich bei letzterer ein weiterer Zahn im Unterkiefer zurückgebildet. In der oberen Zahnreihe ist der erste Schneidezahn sehr groß ausgebildet, dem dann mehrere kleinere einspitzige Zähne folgen. Ähnliches kann zur unteren Zahnreihe gesagt werden. Die Molaren weisen mehrere spitze Haupthöcker auf. Bei denen des Oberkiefers sind sie dilambdodont (W-förmig) angeordnet.[6][2] Bezüglich der genauen numerischen Position der einzelnen einspitzigen Zähne besteht eine Unsicherheit, so dass zwei verschiedenen Zahnformeln angeführt werden. Die Unterschiede resultieren aus der Diskussion, welcher der einspitzigen Zähne reduziert wurde. Unter Berücksichtigung der Tatsache, dass der Mittelkieferknochen der Säugetiere alleinig die Schneidezähne trägt, befindet sich die Sutur zwischen diesem und dem Oberkiefer zwischen dem fünften und sechsten Zahn vor dem vordersten Molaren, ausgehend von jeweils sieben „Vor-Molaren-Zähnen“ beim True-Spitzmull und beim Japanischen Spitzmull. Demnach wäre der fünfte „Vor-Molaren-Zahn“ der Eckzahn und es verbleiben zwei obere Schneidezähne (sechster und siebenter „Vor-Molaren-Zahn“) im Mittelkieferknochen. In dieser Ansicht bestehen im Oberkiefer vier Prämolaren zwischen dem Eckzahn und dem ersten Molar, während höchstwahrscheinlich der dritte Schneidezahn, also der dritte von vorn gezählte Zahn, verlustig ging.[7] Andere Autoren sehen jedoch einen der Prämolaren als nicht erneut ausgebildet an, vermutet wird der zweite oder dritte, so dass das obere Dauergebiss drei Schneidezähne und drei Prämolaren aufweisen sollte.[8] Für die untere Zahnreihe ergäbe sich dann die numerische Position aus der Lage der einzelnen Zähne zu den jeweiligen Entsprechungen in der oberen Zahnreihe, wobei nach übereinstimmender Meinung der vorderste Schneidezahn generell fehlt. Neben der unterschiedlichen Zahnanzahl beim Japanischen Spitzmull und beim True-Spitzmull lassen sich auch noch Abweichungen in der Höhe der Zahnkronen der Mahlzähne aufzeigen. Bei ersterem sind sie hochkronig (hypsodont), bei letzterem niederkronig (brachyodont).[9] Allen Japanischen Spitzmullen ist abweichend von den Eigentlichen Maulwürfen und übereinstimmend mit den Neuweltmaulwürfen ein Zahnwechsel vom Milch- zum Dauergebiss zu eigen.[3]
Skelettmerkmale
Im postcranialen Skelettbau, vor allem der Vordergliedmaßen, bestehen teils deutliche Unterschiede zwischen den Japanischen Spitzmullen und den Vertretern der grabenden Maulwürfe, was auf eine geringere Grabbefähigung ersterer hinweist. Das Schlüsselbein ist ähnlich wie beim Amerikanischen Spitzmull und beim Langschwanzmnaulwurf nicht so stark gestaucht wie im Vergleich zu den Eigentlichen Maulwürfen und den Neuweltmaulwürfen. Das Schulterblatt weist im Unterschied zu den beiden letztgenannten und übereinstimmend mit ersteren beiden noch ein Metacromion auf, ein Fortsatz, der am Acromion ansetzt und einen Teil des Musculus trapezius aufnimmt. Zusätzlich ist der Oberarmknochen schlanker und die Knochen des Unterarms (Elle und Speiche) sind länger als bei den grabenden Maulwürfen.[10][11][3] Typischerweise ist bei den Japanischen Spitzmullen der Präpollex („Vordaumen“), auch Os falciforme genannt, sehr klein. Bei den grabenden Maulwürfen verbreitert er durch seine enorme Größe seitlich die Handfläche.[12][13]
Genetische Merkmale
| Gattung | Chromosomensatz | Fundamentale Anzahl | X-Chromosom | Y-Chromosom |
|---|---|---|---|---|
| Dymecodon | 2n = 34 | FNa = 62 | metazentrisch | fleckenförmig |
| Urotrichus | 2n = 34 | FNa = 64 | metazentrisch | fleckenförmig |
| Neurotrichus | 2n = 38 | FNa = 72 | metazentrisch | fleckenförmig |
| Scaptonyx | 2n = 34 | FNa = 64 | metazentrisch | fleckenförmig |
Sowohl der Japanische Spitzmull als auch der True-Spitzmull verfügen über einen diploiden Chromosomensatz von 2n = 34. Die Anzahl der Arme der Autosomenpaare beträgt bei ersterer Art 64, bei letzterer 62. Insgesamt zeigt sich der Chromosomensatz beider Vertreter sehr homogen aufgebaut. Das erweist sich unter anderem durch ein sehr großes, metazentrisches Chromosom 1 und ein Chromosom 5 mit zusätzlicher Einschnürung, das X-Chromosom ist jeweils metazentrisch, das Y-Chromosom fleckenförmig. Unterschiede bestehen unter anderem im Chromosom 15, das einerseits acrozentrisch wie beim True-Spitzmull oder subtelozentrisch wie beim Japanischen Spitzmull sein kann. Vergleichbar zu den Japanischen Spitzmullen ist der Chromosomensatz des auf dem ostasiatischen Festland vorkommenden Langschwanzmaulwurf (Scaptomyx) als engen Verwandten, dessen Chromosom 15 allerdings eine metazentrische Form besitzt. Generell zeichnen sich die Maulwürfe durch einen sehr konservativen Bau des Chromosomensatzes aus. Die ursprüngliche Version wird ebenfalls mit 2n = 34 angenommen. Dieser war wohl auch den gemeinsamen Vorfahren der Japanischen Spitzmulle zu eigen, in Nachfolgenden kam es dann nur zu einzelnen Veränderungen. Innerhalb des Japanischen Spitzmulls ist eine Besonderheit am Chromosom 14 zu verzeichnen, da sich hier zwei cytogentische Gruppen abzeichnen: eine westliche Form mit einem subtelozentrischen Chromosom 14 und eine östliche mit einem metazentrischen. Beim True-Spitzmull ist das entsprechende Chromosom subtelozentrisch.[14][15][16][17][18]
Verbreitung und Lebensraum
Die Japanischen Spitzmulle sind endemisch in Japan verbreitet. Beide Arten bewohnen die südlichen drei Hauptinseln Honshū, Shikoku und Kyūshū, der Japanische Spitzmull kommt zusätzlich auf einigen vorgelagerten Inseln vor. Sie treten sowohl sympatrisch als auch parapatrisch auf. Der Japanische Spitzmull bevorzugt vor allem tiefere Lagen mit Laubwäldern, der True-Spitzmull ist hingegen häufig in höheren Gebirgsregionen und in Nadelwäldern anzutreffen. Dadurch ist sein Vorkommen deutlich fleckenhafter als das des Japanischen Spitzmulls. Darüber hinaus werden auch Buschlandschaften und offene Grasgebiete besiedelt, gelegentlich auch Bereiche mit steinigerem Untergrund.[19][3]
Lebensweise
Die Lebensweise der Japanischen Spitzmulle ist nur punktuell gut untersucht. Ihr Körperbau eignet sich weniger gut für eine grabende Lebensweise. Die Tiere finden sich daher öfter an der Oberfläche als andere Maulwurfsarten. Dennoch legen sie Pfade an, die nur gering in den Untergrund eingetieft sind oder dichte verrottenden Laubfalllagen am Erdboden queren. Die Tunnel und Pfade dienen vornehmlich der Suche nach Nahrung. Diese besteht vorwiegend aus Wirbellosen, teilweise aber auch aus pflanzlichem Material. Einen großen Anteil machen Insekten und Regenwürmer aus, darüber hinaus bereichern Spinnen, Hundertfüßer und andere den Speiseplan. Beim Japanischen Spitzmull ist bekannt, dass er über variierende Jagdmethoden verfügt, die an die jeweiligen Bedingungen der Erdoberfläche beziehungsweise der unterirdischen Gänge angepasst sind.[20][21] Über die Fortpflanzung ist nur wenig bekannt. Die Paarung findet im Frühjahr statt, beim Japanischen Spitzmull kann auch eine herbstliche Phase auftreten. Zu Beginn der Fortpflanzungszeit schwellen die Hoden der Männchen an. Die Weibchen tragen bis zu sechs Embryonen aus. Die Dauer der Trächtigkeit ist unbekannt, wird aber auf rund vier Wochen geschätzt. Ebenso lange währt wohl auch die Saugphase. Das Höchstalter der Tiere beträgt schätzungsweise zwei bis drei Jahre.[22][23] Abweichend von mehreren Arten der Eigentlichen Maulwürfe und zusätzlich vom Amerikanischen Spitzmull treten bei den Weibchen der Japanischen Spitzmulle keine Hermaphroditen auf.[24][25][19][3]
Systematik
Innere Systematik der Maulwürfe nach He et al. 2016[26]
|
Die Japanischen Spitzmulle sind eine Tribus aus der Familie der Maulwürfe (Talpidae) und der Ordnung der Insektenfresser (Eulipotyphla). Die Gruppe vereint nur wenig an eine grabende Lebensweise angepasste Maulwürfe, deren beide heute bekannten Vertreter in Japan verbreitet sind. Ihr Skelettbau ist dadurch noch nicht so stark überprägt, außerdem bestehen durch den äußerlich eher an Spitzmäuse erinnernden Körperbau phänotypisch größere Unterschiede zu den grabenden Angehörigen der Maulwürfe wie den Eigentlichen Maulwürfen (Talpini) und den Neuweltmaulwürfen (Scalopini). Innerhalb der Familie stellen der Amerikanische Spitzmull (Neurotrichini) und der Langschwanzmaulwurf (Scaptonychini) die nächsten Verwandten dar, die eine ähnliche Lebensweise wie die Japanischen Spitzmulle verfolgen. Die drei Gruppen formen eine monophyletische Einheit.[26] Einige Systematiken gliedern die beiden letztgenannten teilweise auch in die Urotrichini mit ein.[3] Unterschiede zwischen den Urotrichini, den Neurotrichini und den Scaptonychini finden sich jedoch vor allem im Gebissaufbau.[8][27][28] Gemäß molekulargenetischen Untersuchungen erfolgte die Trennung der drei Gruppen von den anderen Linien der Maulwürfe bereits im Oberen Eozän vor rund 37 Millionen Jahren. Eine stärkere Aufspaltung der verschiedenen Spitzmulle setzte im Zeitraum von vor 30 bis 27 Millionen Jahren im Oligozän ein. Die gemeinsame Einheit der Spitzmull-Vertreter steht einer Klade gegenüber, die sich aus den Eigentlichen Maulwürfen, den Desmanen (Desmanini) und dem Sternmull (Condylurini) zusammensetzt. Übergeordnet gehören alle genannten Maulwurfslinien der Unterfamilie der Altweltmaulwürfe (Talpinae) an, die zusätzlich noch die Neuweltmaulwürfe beinhalten. Die Altweltmaulwürfe umfassen sowohl unterirdisch lebende als auch an eine semi-aquatische Lebensweise angepasste Tiere aus Eurasien und aus Nordamerika. Ihre Schwestergruppe bilden die Spitzmausmaulwürfe (Uropsilinae) des östlichen Asiens.[26]
In der Regel werden zwei rezente Arten verteilt auf ebenso viele Gattungen unterschieden. Dadurch setzt sich die Tribus der Japanischen Spitzmulle folgendermaßen zusammen:[28][26]
- Tribus: Japanische Spitzmulle (Urotrichini Dobson, 1883)
- True-Spitzmull (Dymecodon True, 1886); höhere Gebirgslagen auf den Inseln Honshū, Shikoku und Kyūshū
- Japanischer Spitzmull (Urotrichus Temminck, 1841); Tiefländer auf den Inseln Honshū, Shikoku und Kyūshū
Der True-Spitzmull gilt aufgrund einiger anatomischer Besonderheiten gegenüber dem Japanischen Spitzmull, zu nennen wären die höhere Zahnanzahl und die niederkronigen hinteren Zähne, als die stammesgeschichtlich ältere Form.[9] Abweichend von der hier vorgestellten Gliederung der Japanischen Spitzmulle vereint der achte Band des Standardwerkes Handbook of the Mammals of the World aus dem Jahr 2018 unter den Urotrichini zusätzlich noch den Langschwanzmaulwurf und den Amerikanischen Spitzmull. Hierbei werden mit den Urotrichina und den Scaptonychina zwei Untertriben ausgehalten, von denen erstere den Langschwanzmaulwurf, letztere neben den eigentlichen Japanischen Spitzmullen auch den Amerikanischen Spitzmull aufnimmt.[3]
Neben den heute noch bestehenden Formen sind auch mehrere ausgestorbene Vertreter beschrieben worden:
- Desmanodon Engesser, 1980
- Myxomygale Filhol, 1890
- Paratalpa Lavocat, 1951
- Percymygale Hugueney & Maridet, 2017
- Pseudoparatalpa Lopatin, 1999
- Tenuibrachiatum Ziegler, 2003
Der Verweis von Desmanodon zu den Japanischen Spitzmullen ist nicht ganz eindeutig,[29] da einige Autoren dieses auch mit unsicherer Stellung innerhalb der Familie angeben.[30][31] Ähnlich verhält es sich mit Desmanella, das mitunter der Tribus zugerechnet,[30] von dem überwiegenden Teil der Wissenschaftler jedoch als den Spitzmausmaulwürfen zugehörig betrachtet wird.[32][33][34] Für Pseudoparatalpa ist auch eine Gleichsetzung mit Paratalpa denkbar.[30] Darüber hinaus gibt es noch weitere Formen, die aber weitgehend als synonym zu bereits bestehenden betrachtet werden müssen. Hierzu gehören unter anderem Teutonotalpa, das im Jahr 1974 von J. Howard Hutchison benannt worden war,[35] aber identisch mit Paratalpa ist,[36] ebenso wie Palurotrichus, zurückgehend auf Reinhard Ziegler 1985,[37] mit Myxomygale übereinstimmt.[38][36]
Forschungsgeschichte
Als erster westlicher Forscher berichtete Coenraad Jacob Temminck über spitzmausartige Maulwürfe in Japan. Auf ihn geht auch die wissenschaftliche Erstbeschreibung des Japanischen Spitzmulls zurück, die er im Jahr 1841 veröffentlichte und in der er die Gattungsbezeichnung Urotrichus einführte.[39] Gut 45 Jahre später benannte dann Frederick W. True den True-Spitzmull als weiteren Vertreter, setzte ihn aber mit der Gattung Dymecodon vom Japanischen Spitzmull deutlich ab.[40] Bereits drei Jahre zuvor, im Jahr 1883, hatte George Edward Dobson eine längere Abhandlung über Maulwürfe veröffentlicht, eingebunden in ein mehrbändiges Werk zu den Insektenfressern. Hierin versuchte sich Dobson an einer Gliederung der Familie der Maulwürfe und stellte dabei mehrere Untergruppen auf. Als Unterscheidungsmerkmale zog er die variierende Gebissstruktur und weitere anatomische Charakteristika heran. Hierbei definierte Dobson eine taxonomische Einheit namens Urotrichi, welche sich durch einen schlanken Oberarmknochen, ein ebensolches Schlüsselbein sowie durch moderat breite Hände ohne ausgeprägten Präpollex („Vordaumen“) hervorhob. Innerhalb seiner Urotrichi ordnete Dobson sowohl den Japanischen Spitzmull als auch den Amerikanischen Spitzmull und den Langschwanzmaulwurf ein. Letztere beiden sah er als näher zueinander stehend an gegenüber ersteren.[6]
Die Eigenständigkeit der Tribus der Japanischen Spitzmulle wurde dann folgend teils angezweifelt. Oldfield Thomas verwies im Jahr 1912 die Japanischen Spitzmulle zu den Neuweltmaulwürfen als einer der von ihm angenommenen fünf Unterfamilien der Maulwürfe, hierin aber als abgetrennte Tribus der Urotrichini zu den Scalopini. Teilweise beruht die Eingliederung in die Neuweltmaulwürfe auf einem ähnlichen Gebissaufbau mit einem vergrößerten vorderen oberen Schneidezahn, was den Eigentlichen Maulwürfen fehlt.[41] Thomas' Gliederung wurde zum Teil von George Gaylord Simpson im Jahr 1945 in seiner generellen Taxonomie der Säugetiere adaptiert, ohne aber die tribale Aufgliederung zu übernehmen.[42] Sie findet sich dann auch in John Reeves Ellermans und Terrence C. S. Morrison-Scotts Katalogwerk zur asiatischen Säugetierfauna aus dem Jahr 1966 wieder.[43] Erst ein Jahr später stellte Leigh Van Valen eine neue Aufteilung der Maulwürfe vor und trennte dabei die Neuweltmaulwürfe und die Japanischen Spitzmulle wieder voneinander. Beide führte er auf tribaler Ebene innerhalb der Altweltmaulwürfe. Im gleichen Zug gliederte Van Valen den Langschwanzmaulwurf aus den Japanischen Maulwürfen aus und kreierte für ihn die eigene Tribus der Scaptonychini. Seinen Schritt begründete er mit dem abweichenden Gebissaufbau beim Langschwanzmaulwurf.[44] Eine ähnliche Argumentation zog Rainer Hutterer im Jahr 2005 heran, um im Rahmen des Werkes Mammal Species of the World den Amerikanischen Spitzmull ebenfalls aus den Japanischen Spitzmullen herauszulösen und mit ihm die Tribus der Neurotrichini zu begründen. Damit verblieben innerhalb der Japanischen Spitzmulle lediglich die beiden heutigen Arten Japans.[28]
In der Regel wurden die heutigen Spitzmulle als relativ eng verwandt betrachtet, was Dobson schon bei seiner Aufstellung der Tribus der Urotrichini unterstrich. Bei der Herauslösung des Amerikanischen Spitzmulles aus der Tribus im Jahr 2005 gab Hutterer neben den Gebissmerkmalen noch molekulargenetische Ergebnisse als Begründung an. Diese waren zwei Jahr zuvor von Akio Shinohara und Kollegen veröffentlicht worden und zeigten keine unmittelbar nähere Beziehung der Spitzmulle zueinander an. Vielmehr standen die Japanischen Spitzmulle den Desmanen und den Eigentlichen Maulwürfen näher, der Amerikanische Spitzmull den Neuweltmaulwürfen. Den Langschwanzmaulwurf berücksichtigte die Studie nicht.[45] Im folgenden Jahr bezog Shinohara letzteren mit ein, im Resultat war die Verwandtschaft der Spitzmulle aber eher ambivalent.[46] Gleichzeitig durchgeführte anatomische Untersuchungen konnten die ersten genetischen Analysen weitgehend bestätigen, so dass die Ähnlichkeit der Spitzmulle weitgehend als konvergente Entwicklung aufgefasst wurde.[47][11] Eine genetische Studie aus dem Jahr 2016 hatte erstmals alle damals bekannten Gattungsvertreter der Maulwürfe zur Basis. Dadurch konnte die Zugehörigkeit der Spitzmulle zueinander bestätigt werden. Die Zuordnung in verschiedenen Triben blieb hierbei erhalten.[26] Ein ähnliches Ergebnis legte eine anatomische Studie vor, in der neben den heutigen Vertretern auch zahlreiche fossile Formen eingebunden waren.[48]
Stammesgeschichte
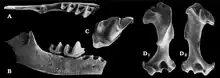
Fossilvertreter der Tribus der Urotrichini lassen sich bis in das Paläogen zurückverfolgen. Der überwiegende Teil des Fundmaterials ist aus Eurasien bekannt. Als älteste Vertreter können Percymygale und Myxomygale angesehen werden. Erstere Form kam aus verschiedenen Karstspalten entlang der Donau, etwa von Möhren in der Nähe von Treuchtlingen, im südwestlichen Deutschland zu Tage. Sie trat vom ausgehenden Eozän vor 34 Millionen Jahren durch das gesamte Oligozän bis zum Unteren Miozän hinweg auf. Zu den spätesten Nachweisen gehören jene von Merkur aus dem nordwestlichen Tschechien. Möglicherweise lassen sich auch einige isolierte Zähen von der Isle of Wight dieser Gattung zuordnen. Das Fundmaterial umfasst Gebissreste und Gliedmaßenknochen, bei denen sich der Oberarmknochen durch einen leichten und schlanken Bau auszeichnet. Ursprünglich war Percymygale in Myxomygale integriert, wurde aber im Jahr 2018 unter anderem aufgrund eines umfangreicheren vorderen Gebisses als eigenständige Form ausgegliedert.[33][49][50] Myxomygale selbst war bereits im Jahr 1890 durch Henry Filhol anhand eines Unterkiefers aus den Phosphatgruben von Quercy im Südwesten Frankreichs wissenschaftlich eingeführt worden.[51] Dieser Erstfund stellt gleichzeitig mit einer Einstufung in das Obere Oligozän einen der ältesten Belegen der Gattung dar. Lediglich ein weiterer Unterkiefer aus den Cyrenamergeln bei Offenbach am Main weist ein noch geringfügig höheres Alter auf.[52] Teilweise wird aber auch die Gattung Oreotalpa, die mit einer Stellung im Oberen Eozän den bisher frühesten Nachweis der Maulwürfe in Nordamerika repräsentiert, zu Myxomygale verwiesen,[49] was bisher aber noch nicht verifiziert werden konnte.[50] Nachfolgend ist Myxomygale überwiegend für das Miozän verzeichnet Ihr können heute zahlreiche Reste aus mehreren Lokalitäten in Mittel- und Westeuropa zugeschrieben werden. Zu den bekanntesten gehören wiederum mehrere Karstpalten der Fränkischen Alb im südlichen Deutschland, darunter Petersbuch. Allein die Spalte Petersbuch 28, die sich vor rund 18 Millionen Jahren füllte, erbrachte rund 115 Ober- und Unterkieferfragmente zuzüglich von 50 isolierten Zähnen, des Weiteren auch mehr als 230 Oberarmknochen und rund ein Dutzend weitere Gliedmaßenreste. Bei der wenige Millionen Jahre jüngeren Spalte Petersbuch 10 sind es immerhin noch rund ein halbes Dutzend Gebiss- und Beinfragmente.[30][53] Vergleichbar zu Percymygale repräsentiert Myxomygale einen kleinen Maulwurf mit schlanken Gliedmaßen.[37][36][30][50]
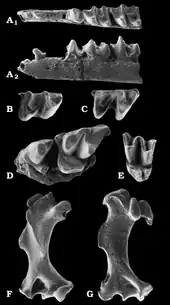
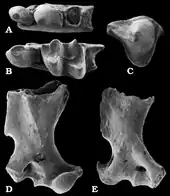
Aus den Karstspalten der Fränkischen Alb kamen zusätzlich noch Fossilreste anderer Angehöriger der Urotrichini zum Vorschein. Tenuibrachiatum, benannt im Jahr 2003 durch Reinhard Ziegler, wurde bisher weitgehend nur aus der Spalte Petersbuch 31 dokumentiert, ihr Alter entspricht der von Petersbuch 10. Es liegen über ein Dutzend Gebiss- und Zahnreste vor, ein besonderes Charakteristikum stellt die reduzierte Anzahl an vorderen Zähnen im Unterkiefer dar. Gemäß des wissenschaftlichen Namens waren auch bei Tenuibrachiatum die Vordergliedmaßen grazil ausgeprägt, belegt an fast einem Dutzend Oberarmknochen.[30] Ähnlich verhält es sich mit Paratalpa. Die Form ist unter anderem mit einigen Zähnen aus der oberoligozänen Fossillagerstätte Rott bei Bonn[54] und mit zahlreichen Gebissfragmenten aus untermiozänen Schichten bei Ulm und dem nahe gelegenen Eggingen registriert.[38][36] Die Gattung war recht weit verbreitet und wurde unter anderem aus Cournon im westlichen Frankreich berichtet, der Typuslokalität. Sie verschwand aber im Unteren Miozän wieder. Ersetzt wurde Paratalpa dann durch Desmanodon. Beide Vertreter sind anhand des Gebisses kaum zu unterscheiden, weswegen sie teils als synonym angesehen wurden. Bezüglich des Oberarmknochens ergeben sich aber deutliche Unterschiede, da dieser bei Desmanodon durch eine langgestreckte Knochenleiste am Schaft (Tuberculum teres) deutlich spezialisierter war als bei Paratalpa und dadurch eine gewisse Grabbefähigung vermuten lässt. Anzunehmen ist, dass die Form eher trockene Landschaften besiedelte. Aus den süddeutschen Karstspaltenfundstellen wie Petersbuch liegen daher nur einige wenige Funde vor. Dagegen ist Desmanodon aus Fundstellen wie Forsthard, Rembach oder Rauscheröd in Niederbayern ebenso wie aus Kleineisenbach sowie Giggenhausen nördlich von München häufiger belegt, letztere beiden Lokalitäten lieferten nahezu 50 Gebiss- und Zahnfunde sowie Gliedmaßenknochen.[55][30][56] Auch sind Einzelzähne aus dem Tagebau von Oberdorf bei Graz dokumentiert.[57] Von diesen mitteleuropäischen Fundstellen reicht das Nachweisgebiet von Desmanodon über den südalpinen und südosteuropäischen Raum bis nach Anatolien, von wo aus den Lokalitäten Yeni Eskihisar und Sariçay das Belegmaterial der Gattung in Form von Zähnen und einem Oberarmknochen stammt. Des Weiteren ist Desmanodon auch auf der Iberischen Halbinsel registriert. Schwerpunkt des zeitlichen Auftretens bildet das Untere Miozän, im Mittleren Miozän verschwand die Gattung wieder aus weiten Teilen Europas Jedoch zeigen mehrere Zahnfund aus Borský Svätý Jur in der Slowakei ein Überleben bis in das Obere Miozän an.[58][29][31][59][60]
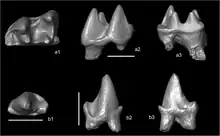
Abseits der zahlreichen europäischen und westasiatischen Funde liegen Fossilreste zusätzlich aus dem zentralen Asien vor. Myxomygale konnte nördlich des Aralsees in Kasachstan aus der untermiozänen Aral-Formation dokumentiert werden, der Nachweis beruht aber nur auf wenigen isolierten Zähnen. Die gleiche Gesteinseinheit barg einzelne Unterkieferreste von Pseudoparatalpa, bei denen als auffälliges Kennzeichen der Eckzahn fehlt.[61][62][63] Einige isolierte Zähne sind des Weiteren aus der Fundstelle Tagay am Westufer der Insel Olchon im Baikalsee überliefert. Sie werden zu Desmanodon gestellt und bilden den ersten Beleg der Form außerhalb des westeurasischen Fundgebietes. Das Material datiert in den Übergang vom Unteren zum Mittleren Miozän.[64]
Neben einigen kontroversen Funden der Gattung Urotrichus aus dem Oberen Miozän Europas[65] sind die heutigen Vertreter der Japanischen Spitzmulle erstmals im Verlauf des Mittelpleistozäns fossil fassbar. Funde liegen vor allem aus verschiedenen Höhlen auf der japanischen Hauptinsel Honshū vor. Zu nennen wäre als einer der ältesten Nachweise die Ikumo-Höhle in der Präfektur Yamaguchi im äußersten Südwesten der Insel. Ähnlich den heutigen Gegebenheiten treten der Japanische Spitzmull und der True-Spitzmull zum Teil gemeinsam auf.[66][67] Angenommen wird, dass die Vorfahren der heutigen Arten im Verlauf des Pliozäns von Ostasien kommend über Landbrücken die japanische Inselwelt erreichten. Eventuell war die Gattung Dymecodon ursprünglich über weite Bereiche des heutigen Japans verbreitet, wurde aber mit den Ankunft beziehungsweise Herausdifferenzierung von Urotrichus in höhere Gebirgslagen abgedrängt.[68]
Bedrohung und Schutz
Beide Arten der Japanischen Spitzmulle sind laut IUCN in ihrem Bestand „nicht gefährdet“ (least concern). Als Begründung gibt die Naturschutzorganisation die jeweils weite Verbreitung und Häufigkeit an. Größere Bedrohung sind nicht bekannt. Sowohl der Japanische Spitzmull als auch der True-Spitzmull kommen in mehreren Schutzgebieten vor.[69][70]
Literatur
- Hisashi Abe: Classification and biology of Japanese Insectivora (Mammalia). I. Studies on variation and classification. Journal of the Faculty of Agriculture, Hokkaido University 55, 1967, S. 191–265 ()
- Hisashi Abe: Classification and biology of Japanese Insectivora (Mammalia). II. Biological aspects. Journal of the Faculty of Agriculture, Hokkaido University 55 (4), 1968, S. 429–458 ()
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 602–603) ISBN 978-84-16728-08-4
- Ronald M. Nowak: Walker's Mammals of the World. 2 Bände. 6. Auflage. The Johns Hopkins University Press, Baltimore MD u. a. 1999, ISBN 0-8018-5789-9
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. A taxonomic and geographic Reference. 2 Bände. 3. Auflage. The Johns Hopkins University Press, Baltimore MD 2005, ISBN 0-8018-8221-4
Einzelnachweise
- Naohiko Sagara: Hair Types of Japanese Insectivora, with Special Reference to the Presence of Straight Guard Hairs. Journal of the Mammalogical Society of Japan 11 (1/2), 1986, S. 57–64 ()
- Hisashi Abe: Classification and biology of Japanese Insectivora (Mammalia). I. Studies on variation and classification. Journal of the Faculty of Agriculture, Hokkaido University 55, 1967, S. 191–265 ()
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 602) ISBN 978-84-16728-08-4
- Matthew J. Mason: Evolution of the Middle Ear Apparatus in Talpid Moles. Journal of Morphology 267, 2006, S. 678–695
- Daisuke Koyabu, Misato Hosojima und Hideki Endo: Into the dark: patterns of middle ear adaptations in subterranean eulipotyphlan mammals. Royal Society Open Science 4, 2017, S. 170608, doi:10.1098/rsos.170608
- George Edward Dobson: A Monograph of the Insectivora, systematic and anatomical. Part II. London, 1883, S. 126–172 ()
- Yoshiharu Imaizumi und Kinziro Kubota: Numerical identification of teeth in Japanese shrew-moles, Urotrichus talpoides and Dymecodon pilirostris. The Bulletin of Tokyo Medical and Dental University 25 (2), 1978, S. 91–99 ()
- Alan C. Ziegler: Dental homologies and possible relationships of recent Talpidae. Journal of Mammalogy 52 (1), 1971, S. 50–68
- G. Sansalone: Evolution of hypsodonty reveals a long-standing ecological separation in the Japanese shrew-moles. Journal of Zoology 297, 2015, S. 146–155
- Berry Campbell: The shoulder anatomy of the moles. A study in phylogeny and adaptation. The American Journal of Anatomy 64, 1939, S. 1–39
- Marcelo R. Sánchez-Villagra, Inés Horovitz und Masaharu Motokawa: A comprehensive morphological analysis of talpid moles (Mammalia) phylogenetic relationships. Cladistics 22, 2006, S. 59–88
- Marcelo R. Sánchez-Villagra und Peter R. Menke: The mole’s thumb – evolution of the hand skeleton in talpids (Mammalia). Zoology 108, 2005, S. 3–12
- Christian Mitgutsch, Michael K. Richardson, Rafael Jiménez, José E. Martin, Peter Kondrashov, Merijn A. G. de Bakker und Marcelo R. Sánchez-Villagra: Circumventing the polydactyly ‘constraint’: The mole’s ‚thumb‘. Biology Letters 8, 2011, S. 74–77, doi:10.1098/rsbl.2011.0494
- Kimiyuki Tsuchiya: Karyological investigations of Japanese mammals. Honyurui Kagaku 33, 1976, S. 53–59 (japanisch) ()
- Shin-ichiro Kawada und Yoshitaka Obara: Reconsideration of the Karyological Relationship between Two Japanese Species of Shrew-moles, Dymecodon pilirostrisand Urotrichus talpoides. Zoological Science 16 (1),1999, S. 167–174
- Masashi Harada, Akiro Ando, Kimiyuki Tsuchiya und Kazuhiro Koyasu: Geographical Variations in Chromosomes of the Greater Japanese Shrew-Mole, Urotrichus talpoides (Mammalia: Insectivora). Zoological Science 18, 2001, S. 433–442
- A. Nakata, A. Yoshimura, M. Kuro-o und Y. Obara: Karyotypic evolution and organization of the highly repetitive DNA sequences in the Japanese shrew-moles, Dymecodon pilirostris and Urotrichus talpoides. Cytogenetic and Genome Research 111, 2005, S. 152–158, doi:10.1159/000086385
- Shin-ichiro Kawada, Song Li, Ying-Xiang Wang, Orin B. Mock, Sen-ichi Oda und Kevin M. Campbell: Karyotype evolution of shrew moles (Soricomorpha: Talpidae). Journal of Mammalogy 89 (6), 2008, S. 1428–1434
- Hisashi Abe: Classification and biology of Japanese Insectivora (Mammalia). II. Biological aspects. Journal of the Faculty of Agriculture, Hokkaido University 55 (4), 1968, S. 429–458 ()
- Yoshiharu Imaizumi: Hunting Methods in Relation to Hunting Situations in Japanese Shrew-mole, Urotrichus talpoides. Annotations Zoologicae Japonensis 51 (4), 1978, S. 245–253
- Yoshiharu Imaizumi: Hunting Methods in Relation to Hunting Situations in Japanese Shrew-mole, Urotrichus talpoides II. Detection of the Earthworm "Head". Annotations Zoologicae Japonensis 52 (3), 1979, S. 212–224
- Hideaki Usuki: Studies of the Shrew Mole (Urotrichus talpoides) IV. Seasonal reproductive conditions and some reflections on breeding cycle. Journal of the Mammalian Society of Japan 4 (1), 1968, S. 7–11
- Nobuo Ishii: Reproductive Activity of the Japanese Shrew-mole, Urotrichus talpoides Temminck. Journal of the Mammalian Society of Japan 9, 1982, S. 25–36
- F. David Carmona, Masaharu Motokawa, Masayoshi Tokita, Kimiyuki Tsuchiya, Rafael Jiménez und Marcelo Sánchez-Villagra: The Evolution of Female Mole Ovotestes Evidences High Plasticity of Mammalian Gonad Development. Journal of Experimental Zoology (Mol Dev Evol) 310 B, 2008, S. 259–266
- Adriane Watkins Sinclair, Stephen Glickman, Kenneth Catania, Akio Shinohara, Lawrence Baskin und Gerald R. Cunha: Comparative morphology of the penis and clitoris in four species of moles (Talpidae). Journal of Experimental Zoology (Mol Dev Evol) 328B, 2017, S. 275–294, doi:10.1002/jez.b.22732
- Kai He, Akio Shinohara, Kristofer M. Helgen, Mark S. Springer, Xue-Long Jiang und Kevin L. Campbell: Talpid Mole Phylogeny Unites Shrew Moles and Illuminates Overlooked Cryptic Species Diversity. Molecular Biology and Evolution 34 (1), 2016, S. 78–87
- Gerhard Storch und Shuding Qiu: The Neogene mammalian faunas of Ertemte and Harr Obo in Inner Mongolia (Nei Mongol), China. 2. Moles – Insectivora: Talpidae. Senckenbergiana lethaea 64, 1983, S. 89–127
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. A taxonomic and geographic Reference. 2 Bände. 3. Auflage. Johns Hopkins University Press, Baltimore MD 2005, ISBN 0-8018-8221-4 ()
- Lars W. van den Hoek Ostende: Insectivore faunas from the Lower Miocene of Anatolia. Part 4: The genus Desmenodon (Talpidae) with the description of a news pecies from the Lower Miocene of Spain. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen C 100 (1/2), 1974, S. 27–65
- Reinhard Ziegler: Moles (Talpidae) from the late Middle Miocene of South Germany. Acta Palaeontologica Polonica 48 (4), 2003, S. 617–648
- Jérôme Prieto, Lars W. va der Hoek Ostende und János Hír: The Middle Miocene insectivores from Sámsonháza 3 (Hungary, Nógrád County): Biostratigraphical and palaeoenvironmental notes near to the Middle Miocene Cooling. Bulletin of Geosciences 87 (2), 2012, S. 227–240
- Lars W. van den Hoek Ostende: Insectivore faunas from the Lower Miocene of Anatolia — Part 5: Talpidae. Scripta Geologica 122, 2001, S. 1–45
- Lars W. van den Hoek Ostende und Oldrich Fejfar: Erinaceidae and Talpidae (Erinaceomorpha, Soricomorpha, Mammalia) from the Lower Miocene of Merkur-Nord (Czech Republic, MN 3). Beiträge zur Paläontologie 30, 2006, S. 175–203
- Antonio García-Alix, Marc Furió, Raef Minwer-Barakat, Elvira Martín Suárez und Matthijs Freudenthal: Environmental control on the biographical distribution of Desmanella (Soricomorpha, Mammalia) in the Miocene of the Iberian Peninsula. Palaeontology 54 (4), 2011, S. 753–762
- J. Howard Hutchison: Notes on type specimens of European Miocene Talpidae and a tentative classification of Old World Tertiary Talpidae (Insectivora: Mammalia). Geobios 7 (3), 1974, S. 211–256
- Reinhard Ziegler: Talpidae (Insectivora, Mammalia) aus dem Oberoligozän und Untermiozän Süddeutschlands. Stuttgarter Beiträge zur Paläontologie Serie B 167, 1990, S. 1–80
- Reinhard Ziegler: Talpiden (Mammalia, Insectívora) aus dem Orleanium und Astaracium Bayerns. Mitteilungen der Bayerischen Staatssammlung, Paläontologie und historische Geologie 25, 1985, S. 131–175
- Lars W. van den Hoek Ostende: The Talpidae (Insectivora, Mammalia) of Eggingen Mittelhart (Baden-Württemberg, F.R.G.) with special reference to the Paratalpa-Desmanodon lineage. Stuttgarter Beiträge zur Naturkunde Serie B 152, 1989, S. 1–29
- Coenraad Jacob Temminck: Eenige geslachten van Zoogdieren, een deel der Fauna van Japan uitmakende. Het Instituut, von Verslagen en mededeelingen uitgegeven Tür de vier klassen van het Koninklijk Nederlandsch Instituut van Wetenschappen, Letterkunde en Schoone Kunsten über den jare 1841, S. 208–216 ()
- Frederick W. True: Description of a new genus and species of mole, Dymecodon pilirostris, from Japan. Proceedings of the United States National Museum 9, 1886, S. 97–98 ()
- Oldfield Thomas: On the collection of mammals from the Tsin-ling mountains, Central China, presented by Mr. G. Fenwick Owen to the National Museum. Annals and Magazine of Natural History 8 (10), 1912, S. 395–403 ()
- George Gaylord Simpson: The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History 85, 1945, S. 1–350 (S. 52–53)
- J. R. Ellerman und T. C. S. Morrison-Scott: Checklist of Palaearctic and Indian Mammals 1758 to 1946. Zweite Ausgabe, London, 1966, S. 1–809 (S. 33–34) ()
- Leigh van Valen: New Paleocene Insectivores and Insectivore classification. Bulletin of the American Museum of Natural History 135, 1967, S. 217–284
- Akio Shinohara, Kevin L. Campbell und Hitoshi Suzuki: Molecular phylogenetic relationships of moles, shrew moles, and desmans from the new and old worlds. Molecular Phylogenetics and Evolution 27, 2003, S. 247–258, doi:10.1016/S1055-7903(02)00416-5
- Akio Shinohara, Hitoshi Suzuki, Kimiyuki Tsuchiya, Ya-Ping Zhang, Jing Luo, Xue-Long Jiang, Ying-Xiang Wang und Kevin L. Campbell: Evolution and Biogeography of Talpid Moles from Continental East Asia and the Japanese Islands Inferred from Mitochondrial and Nuclear Gene Sequences. Zoological Science 21, 2004, S. 1177–1185
- Masaharu Motokawa: Phylogenetic relationships within the family talpidae. Journal of Zoology 263, 2004, S. 147–157
- Achim H. Schwermann und Richard S. Thompson: Extraordinarily preserved talpids (Mammalia, Lipotyphla) and the evolution of fossoriality. Journal of Vertebrate Paleontology 35 (4), 2015, S. e934828, doi:10.1080/02724634.2014.934828
- Reinhard Ziegler: Moles (Talpidae, Mammalia) from Early Oligocene karstic fissure fillings in South Germany. Geobios 45, 2012, S. 501–513, doi:10.1016/j.geobios.2011.11.017
- Marguerite Hugueney und Olivier Maridet: Evolution of Oligo-Miocene talpids (Mammalia, Talpidae) in Europe: focus on the genera Myxomygale and Percymygale n. gen. Historical Biology 30 (1/2), 2018, S. 267–275, doi:10.1080/08912963.2017.1282477
- Henry Filhol: Description d’un nouveau genre d’insectivore provenant des dépôts de phosphate de chaux du Quercy. Bulletin de la Société philomathique de Paris 8 (11), 1890, S. 176–177 ()
- Reinhard Ziegler und Gerhard Storch: Mammals from the Cyrena Beds of Offenbach (Hesse) – Biostratigraphic correlation. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 248 (3), 2008, S. 267–278
- Johannes Klietmann, Doris Nagel, Michael Rummel und Lars W. van den Hoek Ostende: A gap in digging: the Talpidae of Petersbuch 28 (Germany, Early Miocene). Paläontologische Zeitschrift 89, 2015, S. 563–592, doi:10.1007/s12542-014-0228-2
- Thomas Mörs: Die Säugetiere der oberoligozänen Fossillagerstätte Rott bei Bonn (Rheinland). Decheniana 149, 1996, S. 205–232
- Reinhard Ziegler und Volker Fahlbusch: Kleinsäuger-Faunen aus der basalen Oberen Süßwasser-Molasse Niederbayerns. Zitteliana 14, 1986, S. 3–58 ()
- Jérôme Prieto: The Middle Miocene mole Desmanodon crocheti sp. nov. (Talpidae, Mammalia): the last representative of the genus in the North Alpine foreland basin. Paläontologische Zeitschrift 84, 2010, S. 217–225, doi:10.1007/s12542-009-0038-0
- Reinhard Ziegler: Marsupialia, Insectivora und Chiroptera (Mammalia). Annalen des Naturhistorischen Museums in Wien. Serie A für Mineralogie und Petrographie, Geologie und Paläontologie, Anthropologie und Prähistorie 99, 1997, S. 43–97
- Florentin Cailleux, Lars W. van den Hoek Ostende und Peter Joniak: The Late Miocene Talpidae (Eulipotyphla, Mammalia) from the Pannonian Region, Slovakia. Journal of Paleontology 98 (1), 2024, S. 128–151, doi:10.1017/jpa.2023.95
- Froukje N. van der Sar, Roos van Glabbeek, Wilma Wessels, Zoran Marković und Hans de Bruijn: Insectivores and marsupials from the upper Oligocene of Banovići (Bosnia and Herzegovina). Journal of Vertebrate Paleontology 37 (6), 2017, S. e1368529, doi:10.1080/02724634.2017.1368529
- Vicente D. Crespo, Rafael Marquina-Blascob, Francisco Javier Ruiz-Sánchez und Plini Montoya: An unusual insectivore assemblage from the early Miocene of southwestern Europe: The talpids and dimylids from the Ribesalbes–Alcora Basin (Spain). Comptes Rendus Palevol 18, 2019, S. 407–416, doi:10.1016/j.crpv.2019.03.003
- A. V. Lopatin: Oligocene and Early Miocene Insectivores (Mammalia) from Western Kazakhstan. Paleontological Journal 33 (2), 1999, S. 182–191
- A. V. Lopatin: Early Miocene small mammals from the North Aral Region (Kazakhstan) with special reference to their biostratigraphic significance. Paleontological Journal 38 (suppl 3), 2004, S. S217–S323
- Oleg G. Bendukidze, Hans de Bruijn und Lars W. van der Hoek Ostende: A revision of Late Oligocene associations of small mammals from the Aral Formation (Kazakhstan) in the National Museum of Georgia, Tbilissi. Palaeodiversity 2, 2009, S. 343–377
- Leonid L. Voyta, Thomas Mörs und Margarita A. Erbajeva: Erinaceomorpha and Soricomorpha (Mammalia) of the Miocene Tagay fauna (Olkhon Island, Lake Baikal, Eastern Siberia): A preliminary report. Palaeobiodiversity and Palaeoenvironments 102, 2022, S. 897–914, doi:10.1007/s12549-022-00557-9
- Reinhard Ziegler: Insectivores (Lipotyphla) and bats (Chiroptera) from the Late Miocene of Austria. Annalen des Naturhistorischen Museums zu Wien 107 A, 2006, S. 93–196
- Yoshinari Kawamura: Quaternary Rodent Faunas in the Japanese Islands (Part 1). Memoirs of the Faculty of Science, Kyoto University, Series of Geology and Mineralogy 53 (1/2), 1988, S. 31–348 ()
- Yoshinari Kawamura: Quaternary Mammalian Faunas in the Japanese Islands. The Quaternary Research 30 (2), 1991, S. 213–220 ()
- Akio Shinohara, Kevin L. Campbell und Hitoshi Suzuki: An evolutionary view on the Japanese talpids based on nucleotide sequences. Mammal Study 30 (suppl. 1), 2005, S. S19–S24, [[doi:10.3106/1348-6160(2005)30[S19:AEVOTJ]2.0.CO;2]]
- D. Laginha Pinto Correia: Dymecodon pilirostris. The IUCN Red List of Threatened Species 2016. e.T41488A22323252 (); zuletzt aufgerufen am 29. Oktober 2022
- F. Cassola: Urotrichus talpoides. The IUCN Red List of Threatened Species 2016. e.T41489A22323186. (); zuletzt aufgerufen am 29. Oktober 2022
Weblinks
- Urotrichus talpoides in der Roten Liste gefährdeter Arten der IUCN 2016. Eingestellt von: F. Cassola, 2016. Abgerufen am 29. Oktober 2022.
- Dymecodon pilirostris in der Roten Liste gefährdeter Arten der IUCN 2016. Eingestellt von: D. Laginha Pinto Correia, 2016. Abgerufen am 29. Oktober 2022.