Spitzmausmaulwürfe
Die Spitzmausmaulwürfe (Uropsilus), manchmal auch Ohrenspitzmaus-Maulwürfe, sind eine Säugetiergattung aus der Familie der Maulwürfe (Talpidae). Sie bilden gleichzeitig die monotypische Unterfamilie der Uropsilinae, die allen anderen Maulwürfen im Stammbaum gegenübersteht. In ihrem äußeren Erscheinungsbild ähneln die Tiere eher den Spitzmäusen als den anderen Mitgliedern der Familie. Dadurch zeichnet sie ein langer Körper, ein ebensolcher Schwanz und eine spitze Schnauze aus. Die Augen sind analog zu den anderen Maulwürfen verkleinert, allerdings ragen die Ohren sichtbar aus dem Fell. Im Skelettbau bestehen deutliche Unterschiede zu den Maulwürfen, was unter anderem am Schultergürtel und den langen, schlanken Knochen der Vordergliedmaßen ersichtlich ist. Die Tiere zeigen dadurch keine Anpassungen an eine grabende Lebensweise. Bevorzugt bewegen sie sich am Erdboden fort, Informationen über ihr Verhalten sind aber rar. Das hauptsächliche Verbreitungsgebiet der Spitzmausmaulwürfe umfasst das zentrale und südwestliche Ostasien sowie das nördliche Südostasien, wo sie bewaldete gebirgige Regionen bewohnen. In ihrer stammesgeschichtlichen Vergangenheit waren sie aber weit über Eurasien verbreitet und kamen auch in Nordamerika vor. Fossilfunde reichen bis in das ausgehende Oligozän zurück, umfassen in der Regel aber lediglich Schädel- und Gebissteile sowie einzelne Elemente der Extremitäten. Die Gattung wurde im Jahr 1871 wissenschaftlich eingeführt, zwölf Jahre später folgte die Benennung der Unterfamilie. Heute nicht mehr anerkannt, wurden die Spitzmausmaulwürfe forschungsgeschichtlich teilweise in drei Gattungen aufgespalten. Gegenwärtig sind zehn Arten anerkannt, genetische Untersuchungen verweisen jedoch auf die Existenz von zahlreichen kryptischen Arten. Über die Bestandsgefährdungen der meisten Vertreter ist kaum etwas bekannt.
| Spitzmausmaulwürfe | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

drei Arten der Spitzmausmaulwürfe: A: Uropsilus huanggangensis, B. Chinesischer Spitzmausmaulwurf (Uropsilus gracilis), C: Dabieshan-Spitzmausmaulwurf (Uropsilus dabieshanensis) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name der Unterfamilie | ||||||||||||
| Uropsilinae | ||||||||||||
| Dobson, 1883 | ||||||||||||
| Wissenschaftlicher Name der Gattung | ||||||||||||
| Uropsilus | ||||||||||||
| Milne-Edwards, 1871 |
Merkmale
Habitus
Die Spitzmausmaulwürfe gehören zu den kleinsten bekannten Maulwürfen. Die Größe zwischen den einzelnen Arten ist nur wenig variabel. In der Regel beträgt die Kopf-Rumpf-Länge 6,5 bis 8,3 cm, hinzu kommt ein 5,0 bis 7,2 cm langer Schwanz. Die Schwanzlänge entspricht somit gut 75 bis 95 % der Länge des restlichen Körpers. Das Gewicht variiert von 5,9 bis 11,2 g, Angaben liegen aber nicht zu allen Arten vor. Der Körperbau der Spitzmausmaulwürfe ist untypisch für Maulwürfe und weist Ähnlichkeiten zu dem der Spitzmäuse auf. Markante Kennzeichen finden sich daher in dem gestreckten Körper, den Kopf mit spitzer, beweglicher Nase und mit äußerlich sichtbaren Ohrmuscheln sowie in dem langen Schwanz. Entsprechend der anderen Maulwürfe ist der Hals sehr kurz, zudem sind die Augen stark zurückgebildet. Die Vorder- und Hinterfüße sind schmal und eignen sich mit ihren schlanken Krallen nicht zum Graben. Das Fell ist am Rücken häufig graubraun bis schwärzlich gefärbt, die Unterseite erscheint bei einigen Arten heller. Die Füße und der Schwanz sind kaum behaart, sondern mit Schuppen versehen, bei letzterem zumeist in ringförmiger Anordnung.[1][2]
Schädel- und Gebissmerkmale

Im Schädelbau ähneln die Spitzmausmaulwürfe weitgehend den Japanischen Spitzmullen (Urotrichini). Der Jochbogen ist vollständig entwickelt, aber sehr schlank. Er wölbt sich hoch nach oben auf, zum Oberkiefer hin verläuft er dadurch abwärts steigend und weniger horizontal wie bei den anderen Maulwürfen. Am vorderen Bogenansatz ragt ein zusätzlicher Fortsatz auf, der als Ankerstelle des Massetermuskels fungiert. Das Foramen lacrimale wird ähnlich groß wie das Foramen infraorbitale, beide sitzen direkt übereinander. An der Schädelbasis sind die Paukenblasen geöffnet. Die Gehörknöchelchen des Mittelohrs zeigen sich normal entwickelt und weisen keine Vergrößerungen auf. Dies wird in der Regel mit einem Hören im höheren Frequenzbereich verbunden.[3][4][5][2]
| Gattung | Zahnformel | Zahnanzahl |
|---|---|---|
| U. andersoni | 38 | |
| U. atronates, U. dabieshanensis, U. fansipanensis, U. gracilis, U. huanggangensis, U. investigator, U. nivatus | 38 | |
| U. aequodonenia | 36 | |
| U. soricipes | 34 |




Das Gebiss ist sehr variabel aufgebaut, was dazu führte, dass die Spitzmausmaulwürfe in der Vergangenheit auf verschiedene Gattungen aufgeteilt wurden. Die Zahnanzahl reicht von 34 bis 38. Gegenüber dem standardisierten vollständigen Gebiss der Höheren Säugetiere ist jenes der Spitzmausmaulwürfe reduziert, wobei der Verlust an Zähnen überwiegend den Bereich der Schneidezähne und Prämolaren betrifft. In der Regel sind in der oberen Zahnreihe sechs bis sieben, in der unteren fünf bis sechs Zähne vor den Molaren ausgebildet. Der zurückgebildete Zahn im hinteren Gebissabschnitt entspricht vermutlich in der Position dem ursprünglich dritten Prämolaren, im vorderen Gebiss ist jeweils der dritte Schneidezahn nicht mehr erhalten, unter Umständen fehlt auch der erste. Die Mahlzähne ähneln weitgehend denen der anderen Maulwürfe. Die Kauflächen werden durch mehrere spitze Höckerchen charakterisiert, die an den oberen Mahlzähnen dilambdodont (W-förmig) angeordnet sind. Abweichend von anderen Maulwürfen tritt am letzten Höckerchen ein Wulst auf, der dem hinteren Zahnrand einen eingedellten und nicht geraden Verlauf verleiht. Als Besonderheit kommt es bei den Spitzmausmaulwürfen zu einem Wechsel vom Milchgebiss zum dauerhaften Gebiss, was ansonsten nur bei den Neuweltmaulwürfen (Scalopiniu) und den Japanischen Spitzmullen (Urotrichini) bekannt ist.[3][6][2]
Skelettmerkmale
Im Skelettbau weichen die Spitzmausmaulwürfe am deutlichsten von allen anderen Maulwürfen ab. Auffallend ist dies vor allem am Schultergürtel, der unterhalb des siebenten Halswirbels ansetzt, während er bei den restlichen Formen mit Ausnahme der Desmane (Desmanini) deutlich höher liegt. Die Flügel des Brustbeinkopfes laden weit aus und sind nicht so eng wie bei den übrigen Maulwürfen. Das Schlüsselbein lässt noch den typischen S-förmigen Verlauf erkennen, wie er bei den Spitzmäusen vorkommt, zeigt sich aber etwas verkürzt. Allerdings ist es nicht so stark gestaucht wie bei anderen Maulwürfen, bei denen es im Extremfall würfelförmig sein kann. Im Gegensatz zu diesen gelenkt das Schlüsselbein sowohl mit dem Schulterblatt als auch mit dem Oberarmknochen, während es bei den restlichen Mitgliedern der Familie nur mit letzterem in Verbindung steht. Am Schulterblatt tritt ein deutliches Metacromion auf (ein Fortsatz am Acromion), der sonst reduziert ist. Der Oberarmknochen, aufgrund der Grabetätigkeit der massivste Langknochen bei den Maulwürfen, ist bei den Spitzmausmaulwürfen lang und schlank. Es fehlen die zumeist wuchtigen Gelenkenden ebenso wie die ausladende Pectoralleiste am oberen Ende als markante Muskelansatzstellen. Gleiches gilt für die Elle, deren oberer Gelenkfortsatz, das Olecranon eher kurz gestaltet und nicht so lang ausgezogen ist wie bei den meisten Maulwürfen. Die Hand der Spitzmausmaulwürfe verfügt über fünf Strahlen. Ein sogenannter Präpollex („Vordaumen“), auch Os falciforme, der bei den grabenden Maulwürfen als verknöchertes Sesambein die Handfläche seitlich verbreitert, ist bei den Spitzmausmaulwürfen nicht ausgebildet. die Endphanlangen, an denen die Krallen ansetzen, sind glatt und nicht eingekerbt. Am Becken artikulieren die beiden Schambeine nicht über die Schambeinfuge. Außerdem besteht kein Kontakt zwischen dem Darmbein und dem Kreuzbein. Generell besteht die Wirbelsäule aus 7 Hals-, 13 Brust-, 6 Lenden-, 6 Kreuzbein- und bis zu 20 Schwanzwirbeln.[7][4][8][9][2]
Genetische Merkmale
| Gattung | Chromosomensatz | Fundamentale Anzahl | X-Chromosom | Y-Chromosom |
|---|---|---|---|---|
| U. andersoni | 2n = 34 | FNa = 52 | metazentrisch | fleckenförmig |
| U. nivatus | 2n = 34 | FNa = 46 | metazentrisch | fleckenförmig |
| U. soricipes | 2n = 36 | FNa = 54 | metazentrisch | fleckenförmig |
| andere Arten | ? | ? | ? | ? |

Cytogenetische Studien fanden bisher nur bei wenigen Vertretern der Spitzmausmaulwürfe Anwendung. Bei den untersuchten Formen wurde ein diploider Chromosomensatz von 2n = 34 bis 36 und eine fundamentale Anzahl, also die Anzahl der Arme der Autosomenpaare, von 46 bis 54 festgestellt. Der Chromosomensatz der Spitzmausmaulwürfe entspricht weitgehend dem der meisten anderen analysierten Maulwürfe, die gesamte Bandbreite reicht aber hier von 2n = 32 bis 48. Es wird angenommen, dass der Wert von 2n = 34 dem der ursprünglichen Maulwürfe entspricht. Das X-Chromosom ist metazentrisch und groß, das Y-Chromosom hingegen fleckenförmig und klein. Letzteres spiegelt einen generellen Trend der Maulwürfe wider, bei dem das männliche Geschlechtschromosom zu einer Miniaturisierung neigt. Bei den Spitzmausmaulwürfen ist das Y-Chromosom allerdings durchschnittlich größer, was möglicherweise auf den ursprünglichen Entwicklungsstand der Gruppe hindeutet.[10][11][12] Das Mitogenom konnte für fast alle Arten erfasst werden. Es besteht aus 16.519 bis 16.575 Basenpaaren.[13][14][15]
Verbreitung und Lebensraum
Die Spitzmausmaulwürfe sind in Ost- und Südostasien verbreitet. Das hauptsächliche Vorkommen findet sich im zentralen und südwestlichen China mit den Provinzen Sichuan und Yunnan sowie angrenzend auch Hubei, Anhui und Jiangxi.[16] Des Weiteren wurde ein Angehöriger auch im nördlichen Vietnam dokumentiert.[17] Einzelne Nachweise liegen aus dem nördlichen Myanmar vor, doch ist hier unklar, welchem Vertreter sie zuzuweisen sind. Die meisten Arten haben nur ein kleines, fleckenhaftes Auftreten. Lediglich der Chinesische (Uropsilus gracilis) und der Sichuan-Spitzmausmaulwurf (Uropsilus soricipes) bewohnen ausgedehntere Regionen. Der Lebensraum der Tiere besteht überwiegend aus Wäldern in mittleren und höheren Gebirgsregionen von 1250 bis 4500 m Seehöhe.[1][2]
Lebensweise
Über die Lebensweise der Spitzmausmaulwürfe ist wenig bekannt. Ihr Körperbau eignet sich nicht zum Graben unterirdischer Gänge. Sie leben daher hauptsächlich am Boden und suchen im Laubabfall nach Nahrung. Diese dürfte aus wirbellosen Tieren bestehen.[1][2]
Systematik
Innere Systematik der Maulwürfe nach He et al. 2016[18]
|
Die Spitzmausmaulwürfe sind eine Gattung innerhalb der Familie der Maulwürfe (Talpidae). Sie bilden zudem als momentan einziges rezentes Mitglied eine eigene Unterfamilie, die Uropsilinae, die damit monotypisch ist. Die Uropsilinae gelten als relativ urtümliche Maulwürfe, die sich über ein spitzmausartiges Erscheinungsbild mit langem Schwanz und sichtbaren Ohren charakterisieren. Ihre seitlich gepressten Krallen sind nur wenig zum Graben geeignet, weswegen die Tiere weitgehend oberirdisch leben.[2] Sie weichen dadurch von den meisten anderen Maulwürfen ab und nehmen aus systematischer Sicht eine Sonderrolle innerhalb der Familie ein. Diese drückt sich darin aus, dass sie das Schwestertaxon aller übrigen Mitglieder der Familie darstellen, welche wiederum in der Unterfamilie der Altweltmaulwürfe zusammengefasst werden. Molekulargenetische Untersuchungen ergaben eine Abspaltung der Spitzmausmaulwürfe von den anderen Linien der Maulwürfe bereits im Mittleren Eozän vor rund 47 Millionen Jahren. Eine stärkere Diversifizierung der Gattung selbst setzte aber erst im Übergang vom Miozän zum Pliozän vor etwa 6,8 Millionen Jahren ein.[14][18][15][19][20]
Die Unterfamilie enthält eine Gattung mit momentan zehn Arten. Daraus ergibt sich folgende Gliederung:[21][2][20][17][16]
- Unterfamilie: Uropsilinae Dobson, 1883
- Uropsilus Milne-Edwards, 1871
- Gleichzahn-Spitzmausmaulwurf (Uropsilus aequodonenia Liu, Liu, Sun, Guo, Fan & Murphy, 2013); Sichuan
- Anderson-Spitzmausmaulwurf (Uropsilus andersoni (Thomas, 1911)); Sichuan
- Schwarzrücken-Spitzmausmaulwurf (Uropsilus atronates (Allen, 1923)); Yunnan
- Dabieshan-Spitzmausmaulwurf (Uropsilus dabieshanensis Hu, Xu, Zhang, Liu, Liao, Yang, Sun, Shi, Ban, Li, Liu & Zhang , 2021); Anhui
- Uropsilus fansipanensis Bui, Okabe, Le, Nguyen & Motokawa, 2023; Lao Cai, Vietnam
- Chinesischer Spitzmausmaulwurf (Uropsilus gracilis (Thomas, 1911)); Chongqing, Sichuan, Guizhou und Yunnan
- Uropsilus huanggangensis Ren, Xu, Li, Yao, Fang, Khanal, Cheng, Zeng, Jiang & Chen, 2023; Jiangxi
- Yunnan-Spitzmausmaulwurf (Uropsilus investigator (Thomas, 1922)); Yunnan
- Yulong-Spitzmausmaulwurf (Uropsilus nivatus (Allen, 1923)); Yunnan
- Sichuan-Spitzmausmaulwurf (Uropsilus soricipes Milne-Edwards, 1871); Sichuan
Innere Systematik der Spitzmausmaulwürfe nach Ren et al. 2023[16]
|
Zuzüglich zu diesen anerkannten Arten belegen molekulargenetische Analysen, dass die Gattung noch weitere kryptische Arten einschließt. Das bisher untersuchte Erbgut wies neben den bekannten Vertretern noch wenigstens acht monophyletische Linien nach, die alle mögliche Arten darstellen. Die Diversifizierung der verschiedenen Linien erfolgte in einem Zeitraum zwischen 4,34 und 1,25 Millionen Jahren, also vom Unteren Pliozän bis zum Unteren Pleistozän. Die bisherige Unterscheidung der Spitzmausmaulwürfe basiert auf Abweichungen in der Zahnformel und Größenvariationen der oberen Prämolaren. Die vermuteten neuen Arten zeigen in diesen Merkmalen jedoch keine Abweichungen, sondern liegen in der Variationsbreite der bereits beschriebenen Formen, hauptsächlich des Chinesischen, aber auch des Sichuan- und des Yunnan-Spitzmausmaulwurfs. Daher müssen weitere Untersuchungen vorgenommen werden, um die möglichen Arten auf morphologischem Weg von den bekannten Vertretern zu unterscheiden.[22][18][15][19]
Darüber hinaus sind neben der heute bestehenden Gattung Uropsilus noch mehrere Fossilformen belegt:[4][23][24]
- Desmanella Engesser, 1972
- Mystipterus Hall, 1930
- Theratiskos van den Hoek Ostende, 2001
Ursprünglich galt Desmanella als Vertreter der Desmane (Desmanini), allerdings spricht die Ausprägung eines funktionalen Milchgebisses eher für eine Stellung bei den Spitztmausmaulwürfen. Einige Autoren zählen zusätzlich Asthenoscapter und Mygatalpa zu den Spitzmausmaulwürfen, überwiegend werden diese aber zu den Desmanen verwiesen.[23][24] Mystipterus wurde zuerst im Jahr 1930 als Fledertier angesehen und später zu den Spitzmäusen verschoben, bevor Leigh Van Valen 1967 den Bezug zu den Maulwürfen erkannte und J. Howard Hutchison die Form im Jahr darauf in die Spitzmausmaulwürfe eingliederte. Sie schließt Mydecodon als Synonym mit ein.[25][4]
Forschungsgeschichte


Die westliche Welt erfuhr erstmals im Jahr 1871 von der Existenz der Spitzmausmaulwürfe. In diesem Jahr führte Alphonse Milne-Edwards den Sichuan-Spitzmausmaulwurf ein und etablierte zudem die Gattung Uropsilus. Letztere sah er als Übergangsform zwischen den Japanischen Spitzmullen (Urotrichini) und den Spitzmäusen an. Im Unterschied zu den äußerlich teils ähnlichen Japanischen Spitzmullen hob Milne-Edwards den weitgehend unbehaarten Schwanz hervor, worauf sich auch der Gattungsname bezieht. Er bildet sich aus den griechischen Wörtern οὐρά (ura) für „Schwanz“ und ψῑλός (psilos) für „kahl“.[26][27] Gut zwölf Jahre später benannte George Edward Dobson die Unterfamilie der Uropsilinae als höhere taxonomische Einheit. Er führte sie allerdings unter Uropsili. In einer Übersichtstabelle setzte er die Spitzmausmaulwürfe von allen anderen damals bekannten Maulwürfen durch die vorhandenen Ohrmuscheln und die schlanken Füße mit ihren nicht eingekerbten Endzehen ab.[3] Die heute korrekte Bezeichnung als Unterfamilie der Uropsilinae wurde von Oldfield Thomas im Jahr 1912 erstmals verwendet. Er stellte die Spitzmausmaulwürfe hierbei als eine von fünf Unterfamilien der Maulwürfe heraus.[28]
Nach Milne-Edwards initialer Erstbeschreibung kamen erst zu Beginn des 20. Jahrhunderts weitere Arten hinzu. Das betrifft den Chinesischen Spitzmausmaulwurf (Uropsilus gracilis) und den Anderson-Spitzmausmaulwurf (Uropsilus andersoni), die beide im Jahr 1911 von Oldfield Thomas eingeführt wurden. Alle drei damals bekannten Arten zeigten nur wenige morphologische Varianz, zeichneten sich aber weitgehend durch eine andere Gebissstruktur aus. So besitzt der Sichuan-Maulwurf insgesamt 34 Zähne, der Chinesische und der Anderson-Spitzmausmaulwurf haben hingegen 38, die aber unterschiedlich angeordnet sind. Oldfield Thomas sah sich dadurch veranlasst, die Gattung Uropsilus aufzuteilen. Er behielt den Sichuan-Spitzmausmaulwurf als Nominatform in der Gattung Uropsilus, verwies aber den Chinesischen Spitzmausmaulwurf zur Gattung Nasillus und den Anderson-Spitzmausmaulwurf zur Gattung Rhynchonax. Das neue System stellte Thomas im Jahr 1911 vor, beschrieb es aber erst im Jahr darauf detaillierter. Die beiden neuen Gattungen unterschieden sich dadurch, dass bei Nasillus im Gegensatz zu Rhynchonax im Unterkiefer ein Schneidezahn weniger vorkommt, dafür aber ein Prämolar zusätzlich auftritt. Abweichend zu Uropsilus verfügen beide Gattungen neben einem weiteren, variablen Zahn im Unterkiefer noch über einen vierten Prämolaren im Oberkiefer.[29][30] Die Unterteilung wurde eine Zeitlang aufrechterhalten, so durch Thomas selbst 1922 mit der Beschreibung des Yunnan-Spitzmausmaulwurfs (Uropsilus investigator),[31] aber etwa auch durch Glover Morrill Allen, der im Folgejahr den Schwarzrücken-Spitzmausmaulwurf (Uropsilus atronates) und den Yulong-Spitzmausmaulwurf (Uropsilus nivatus) etablierte.[32][33] Auch Wilfred Hudson Osgood gehört dazu. In einer Studie aus dem Jahr 1938 deckte er aber bereits Inkonsistenzen bezüglich der Gebissstruktur auf, die vor allem durch Allens Beschreibung der beiden neuen Arten entstanden waren.[34]
Einen gänzlich anderen Weg gingen John Reeves Ellerman und Terence Charles Stuart Morrison-Scott in den 1950er und 1960er Jahren. Sie vereinten alle Spitzmausmaulwürfe unter dem Sichuan-Spitzmausmaulwurf und hoben damit die Unterteilung in mehrere Arten und Gattungen auf. Als Begründung stellten sie die geringe äußerliche Unterscheidbarkeit heraus, während sie die abweichende Zahnanzahl als innerartliche und individuelle Variabilität einschätzten. Innerhalb des Sichuan-Spitzmausmaulwurfs unterschieden Ellermann und Morrison-Scott insgesamt fünf Unterarten: neben der Nominatform zusätzlich den Chinesischen, den Anderson-, den Yunnan- und den Yulong-Spitzmausmaulwurf.[35] Mitte der 1980er Jahre führte Robert S. Hoffmann eine Revision der Spitzmausmaulwürfe durch und kam zu dem Schluss, dass die Spitzmausmaulwürfe doch vielfältiger sind, als von Ellermann und Morrison-Scott postuliert. Dies betraf nicht nur die individuelle Gebissvariabilität, da seine Untersuchungen zufolge Zahnanomalien insgesamt sehr selten auftraten, sondern schloss auch die große geographische und damit einhergehend die ökologische Bandbreite mit ein. Hoffmann teilte die Spitzmausmaulwürfe daher wieder in drei Arten auf, namentlich erkannte er den Sichuan-, den Chinesischen und den Anderson-Spitzmausmaulwurf an. Alle anderen Formen sah er weitgehend als Unterarten des Chinesischen Spitzmausmaulwurfs an.[36] Später kam noch der Yunnan-Spitzmausmaulwurf hinzu. Die neue Systematik fand Einzug in das Standardwerk Mammal Species of the World aus dem Jahr 2005 und wurde mehrfach rezipiert.[37] Erst genetische Untersuchungen beginnend in den 2000er Jahren deckten dann eine deutlich artenreichere Aufteilung der Spitzmausmaulwürfe auf, was mehrere kryptische Arten mit einschloss.[14][18][15][19][20]
Stammesgeschichte
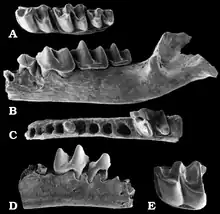
Die Spitzmausmaulwürfe sind heute zumeist auf gebirgige Refugien im östlichen und südöstlichen Asien beschränkt, in ihrer stammesgeschichtlichen Vergangenheit war die Gruppe weit über Eurasien verbreitet und kam auch in Nordamerika vor. Nachweise reichen bis in das ausgehende Oligozän und beginnende Miozän zurück. Die bedeutendste Gattung dieser frühen Zeit bildet Desmanella, eine sehr kleine Form mit dem vollständigen Gebiss der Höheren Säugetiere. Ihr werden über ein Dutzend Arten zugewiesen.[38][39][40] Die frühesten Belege stammen aus Mitteleuropa. In den Übergangsbereich vom Oligozän zum Miozän vor etwa 23 Millionen Jahren fallen einige isolierte Zähne von Eggingen bei Ulm und aus der Blätterkohle der Fossillagerstätte Rott bei Bonn.[41][42] Weitere frühe Funde wurden neben Gebissresten aus Merkur-Nord bei Tuchořice in Tschechien[40] unter anderem in verschiedenen Spaltenfüllungen bei Petersbuch in der Fränkischen Alb getätigt. Zu nennen ist etwa die Spalte 28, deren Inhalt auf ein Alter von rund 18 Millionen Jahre datiert. Sie erbrachte allein über 360 Gebissreste und isolierte Zähne sowie über ein Dutzend Oberarmknochen. Unter den Zähnen befand sich auch der seltene Beleg von Teilen des Milchgebisses.[43][24] Nur wenig jünger lässt sich die Spalte 2 mit gut 40 Überresten aus Zähnen, Unterkieferfragmenten und Gliedmaßenelementen einordnen.[44] Auch die Spalten 6, 18 und 31 enthielten Zahn- und Gebissfragmente von Desmanella, diese gehören aber bereits dem Mittleren Miozän an.[45] Im nordalpinen Gebiet lässt sich die Gattung auch für das weitere Miozän dokumentieren. Von Bedeutung sind hier rund ein halbes Dutzend Zähne aus den mittelmiozänen Süßwasserkalken von Anwil bei Basel, die Burkart Engesser im Jahr 1972 zur Aufstellung von Desmanella verwendete.[46] Deutlich jüngere Funde repräsentieren jene aus Dorn-Dürkheim in Rheinhessen mit einem Alter von etwa 8,5 Millionen Jahren.[38] In einem ähnlichen zeitlichen Zusammenhang stehen einige Zähne aus Eichkogel oder Götzendorf in der Umgebung von Wien[47][48] ebenso wie von verschiedenen Fundstellen wie Studienka und Šalgovce in der Slowakei.[49] Das beginnende Pliozän vor etwa 5 Millionen Jahren markieren wiederum Zahnfunde aus Podlesice im südlichen Polen.[50]
Außerhalb des mitteleuropäischen Raumes ist Desmanella ebenfalls recht häufig belegt. Eine sehr reichhaltige Fundregion liegt im westlichen Teil des Kontinentes, insbesondere auf der Iberischen Halbinsel. Hier decken Lokalitäten unter anderem aus dem Ribesalbes-Alcora-Becken oder bei Concud nahe Teruel, beide im östlichen Spanien, beziehungsweise dem Granada-Becken im Süden des Land einen Großteil des Zeitraums des Miozäns ab. Wie in Mitteleuropa wurden hier überwiegend Einzelzähne und Gebissfragmente dokumentiert.[51][52][53] Der jüngste Nachweis aus dem westlichen Europa und möglicherweise der Gattung ist bisher von Balaruc im Massiv von La Gardiole im südlichen Frankreich dokumentiert, der dem ausgehenden Pliozän angehört. Auffallend hierbei ist, dass das Gebiss bei den späten Vertretern offensichtlich schon um einen Schneidezahn reduziert war.[54] Des Weiteren wurde Desmanella für das südöstliche Europa berichtet. Neben einigen Funden aus Rumänien[39] ist vor allem das Material aus Pikermi in der Region Attika von Bedeutung, welches in das Obere Miozän datiert. Weiter östlich schließen sich einige Funde aus Kleinasien an, so aus Sari Cay, Eskihisar und Amasya in der Türkei. Die Altersdaten reichen vom Mittleren bis zum Oberen Miozän. In Ostasien, dem eigentlichen Verbreitungsgebiet der heutigen Spitzmausmaulwürfe, barg unter anderem die Tunggur-Formation in der zentralen Inneren Mongolei Reste von Desmanella. Ihr Bildungszeitraum entspricht dem Mittleren Miozän. Etwas älter dürften einige wenige Einzelfunde aus der regional nahe gelegenen Aoerban-Formation sein, die dem unteren Abschnitt der Gesteinseinheit entstammen. Die Fundgebiete liegen aber außerhalb des Vorkommens der rezenten Arten.[38][55]
Parallel zu dem sehr frühen Erscheinen von Desmanella im mitteleuropäischen Raum lässt sich im Unteren Miozän in Kleinasien die Gattung Theratiskos nachweisen. Der kleine Spitzmausmaulwurf unterscheidet sich von der erstgenannten Form in einigen Zahnmerkmalen und einem noch schlankeren Oberarmknochen, teilt mit ihr jedoch das vollständige Gebiss der Höheren Säugetiere. Seine Überreste wurden an verschiedenen Fundstellen im zentralen Anatolien aufgefunden. Einzelne Zähne und Kieferfragmente sowie Teile des Bewegungsapparates sind dadurch aus Keseköy, Harami und Gökler bekannt. Das Material von Keseköy, verschiedene Unterkiefer- und Oberarmfragmente, diente Lars W. van den Hoek Ostende im Jahr 2001 zur Etablierung der Gattung.[23][56] Als wahrscheinlich weiteres Nachweisgebiet für Theratiskos ist Zentralasien mit der Aral-Formation nördlich des Aralsees in Kasachstan zu werten. Die im Unteren Miozän entstandene Gesteinseinheit erbrachte bisher aber nur einzelne Zähne.[57][58] Abseits dieser Fundgebiete repräsentiert ein Mahlzahn aus der Ramnagar-Formation in den Siwaliks höchstwahrscheinlich einen Vertreter der Spitzmausmaulwürfe und gehört damit zum ersten Nachweis der Gruppe auf dem Indischen Subkontinent. Er datiert mit Alterswerten von 13,9 bis 13,0 Millionen Jahren in das Mittlere Miozän.[59]
Fossil traten die Spitzmausmaulwürfe auch in Nordamerika auf. Sie werden der Gattung Mystipterus zugewiesen. Das Typusmaterial und der forschungsgeschichtlich älteste Fossilnachweis wurde in der Esmeralda-Formation im Fish Lake Valley im US-Bundesstaat Nevada geborgen und besteht aus einem Unterkieferteil mit einem einzigen Zahn. Später kamen noch weitere Kieferfragmente hinzu. Der hingegen stammesgeschichtlich älteste Fund wurde mit einem Unterkieferbruchstück mit erhaltener hinterer Bezahnung aus der Brule-Formation im südwestlichen North Dakota zu Tage gefördert.[60] Zusätzliche Reste in Form einiger weniger Unterkieferfragmente entstammen der Pawnee-Creek-Formation im Logan County von Colorado.[61] Deutlich umfangreicher sind die Fossilfunde aus dem Quartz Basin in Oregon, die neben Schädelresten und Zähnen auch Elemente des Bewegungsapparates mit einschließen. Weitere Belege sind mit einem fragmentierten Unterkiefer aus der Harrison-Formation im Cherry County von Nebraska bekannt.[62] Mystipterus trat vom Oligozän bis zum Oberen Miozän auf. Abweichend von den eurasischen Fossilformen besaß die Gattung durch den Verlust eines Prämolaren bereits ein reduziertes Gebiss.[4]
Bedrohung und Schutz
Gegenwärtig werden von der IUCN nur vier Arten der Spitzmausmaulwürfe geführt. Von diesen gelten der Sichuan- und der Chinesische Spitzmausmaulwurf aufgrund ihrer vermuteten weiten Verbreitung als nicht gefährdet. Den Anderson- und den Yunnan-Spitzmausmaulwurf weist die Naturschutzorganisation aufgrund fehlender Informationen keiner Gefährdungskategorie zu. Die meisten Arten sind in Naturschutzgebieten präsent.[63][2]
Literatur
- Robert S. Hoffmann und Darrin Lunde: Order Soricomorpha – Shrews and Moles. In: Andrew T. Smith und Yan Xie (Hrsg.): A Guide to the Mammals of China. Princeton University Press, 2008, S. 326–327 ISBN 978-0-691-09984-2
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 ISBN 978-84-16728-08-4
- Ronald M. Nowak: Walker's Mammals of the World. 2 Bände. 6. Auflage. The Johns Hopkins University Press, Baltimore MD u. a. 1999, ISBN 0-8018-5789-9
Einzelnachweise
- Robert S. Hoffmann und Darrin Lunde: Order Soricomorpha – Shrews and Moles. In: Andrew T. Smith und Yan Xie (Hrsg.): A Guide to the Mammals of China. Princeton University Press, 2008, S. 326–327 ISBN 978-0-691-09984-2
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 597) ISBN 978-84-16728-08-4
- George Edward Dobson: A Monograph of the Insectivora, systematic and anatomical. Part II. London, 1883, S. 126–172 ()
- J. Howard Hutchison: Fossil Talpidae (Insectivora, Mammalia) from the Later Tertiary of Oregon. Bulletin of the Museum of Natural History University of Oregon 11, 1968, S. 1–117 (S. 36–46)
- Matthew J. Mason: Evolution of the Middle Ear Apparatus in Talpid Moles. Journal of Morphology 267, 2006, S. 678–695
- Alan C. Ziegler: Dental homologies and possible relationships of recent Talpidae. Journal of Mammalogy 52 (1), 1971, S. 50–68
- B. Campbell: The shoulder anatomy of the moles. A study in phylogeny and adaptation. American Journal of Anatomy 64, 1939, S. 1–39
- Marcelo R. Sánchez-Villagra und Peter R. Menke: The mole’s thumb – evolution of the hand skeleton in talpids (Mammalia). Zoology 108, 2005, S. 3–12
- Marcelo R. Sánchez-Villagra, Inés Horovitz und Masaharu Motokawa: A comprehensive morphological analysis of talpid moles (Mammalia) phylogenetic relationships. Cladistics 22, 2006, S. 59–88
- S. Kawada, S. Li, Y. Wang und S. Oda: Karyological study of Nasillus gracilis (Insectivora, Talpidae, Uropsilinae). Mammalian Biology 71 (2), 2006, S. 115–119, doi:10.1016/j.mambio.2005.11.006
- Masaharu Motokawa, Yi Wu und Masashi Harada: Karyotypes of Six Soricomorph Species from Emei Shan, Sichuan Province, China. Zoological Science 26 (11), 2009, S. 791–797, doi: 10.2108/zsj.26.791
- Svetlana V. Pavlova, Vladimir S. Lebedev, Vasily D. Yakushov, Yongke Zhu, Yun Fang, Yue‑Hua Sun und Boris I. Sheftel: High diversity of small insectivorous mammals on Qinghai–Tibet Plateau and first description of karyotype for four endemics of China. Scientific Reports 11, 2021, S. 24496, doi:10.1038/s41598-021-03809-4
- Feiyun Tu, Shaoying Liu, Yang Liu, Zhiyu Sun, Yonghua Yin, Chaochao Yan, Lu Lu, Bisong Yue und Xiuyue Zhang: Complete mitogenome of Chinese shrew mole Uropsilus soricipes (Milne-Edwards, 1871) (Mammalia: Talpidae) and genetic structure of the species in the Jiajin Mountains (China). Journal of Natural History 48 (23/24), 2014, S. 1467–1483, doi:10.1080/00222933.2013.842012
- Feiyun Tu. Zhenxin Fan, Robert W. Murphy, Shunde Chen, Xiuyue Zhang, Chaochao Yan, Yang Liu, Zhiyu Sun, Jianrong Fu, Shaoying Liu und Bisong Yue: Molecular phylogenetic relationships among Asiatic shrewlike moles inferred from the complete mitogenomes. Journal of Zoological Systematics and Evolutionary Research 53 (2), 2015, S. 155–160, doi:10.1111/jzs.12081
- Yu Xu, Yunting Hu und Feiyun Tu: Mitogenome of a cryptic species within Uropsilus and divergence time estimation. Mitochondrial DNA Part B: Resources 2 (2), 2017, S. 685–686
- Xueyang Ren, Yifan Xu, Yixian Li, Hongfeng Yao, Yi Fang, Laxman Khanal, Lin Cheng, Wei Zeng, Xuelong Jiang und Zhongzheng Chen: A new species of shrew moles, genus Uropsilus Milne-Edwards, 1871 (Mammalia, Eulipotyphla, Talpidae), from the Wuyi Mountains, Jiangxi Province, eastern China. ZooKeys 1186, 2023, S. 25–46, doi:10.3897/zookeys.1186.111592
- Hai Thuan Bui, Shinya Okabe, Linh Tu Hoang Le, Ngan Thi Nguyen und Masahara Motokawa: A new shrew mole species of the genus Uropsilus (Eulipotyphla: Talpidae) from northwestern Vietnam. Zootaxa 5339 (1), 2023, S. 59–78, doi:10.11646/zootaxa.5339.1.3
- Kai He, Akio Shinohara, Kristofer M. Helgen, Mark S. Springer, Xue-Long Jiang und Kevin L. Campbell: Talpid Mole Phylogeny Unites Shrew Moles and Illuminates Overlooked Cryptic Species Diversity. Molecular Biology and Evolution 34 (1), 2016, S. 78–87
- Tao Wan, Kai He, Wei Jin, Shao‐Ying Liu, Zhong‐Zheng Chen, Bin Zhang, Robert W. Murphy und Xue‐Long Jiang: Climate Niche Conservatism and Complex Topography Illuminate the Cryptic Diversification of Asian Shrew‐like Moles. Journal of Biogeography 45 (10), 2018, S. 2400–2414, doi:10.1111/jbi.13401
- Ting-Li Hu, Zhen Xu, Heng Zhang, Ying-Xun Liu, Rui Liao, Guang-Dao Yang, Ruo-Lei Sun, Jie Shi, Qian Ban, Chun-Lin Li, Shao-Ying Liu und Bao-Wei Zhang: Description of a new species of the genus Uropsilus (Eulipotyphla: Talpidae: Uropsilinae) from the Dabie Mountains, Anhui, Eastern China. Zoological Research 42 (3), 2021, S. 294–299, doi:10.24272/j.issn.2095-8137.2020.266
- Liu Yang, Liu Shaoying, Sun Zhiyu, Guo Peng, Fan Zhenxin und Robert W. Murphy: A new species of Uropsilus (Talpidae: Uropsilinae) from Sichuan, China. Acta Theriologica Sinica 3 (2), 2013, S. 113–122
- Tao Wan, Kai He und Xue-Long Jiang: Multilocus phylogeny and cryptic diversity in Asian shrew-like moles (Uropsilus, Talpidae): implications for taxonomy and conservation. BMC Evolutionary Biology 13, 2013, S. 232 ()
- Lars W. van den Hoek Ostende: Insectivore faunas from the Lower Miocene of Anatolia – Part 5: Talpidae. Scripta Geologica 122, 2001, S. 1–35
- Johannes Klietmann, Doris Nagel, Michael Rummel und Lars W. van den Hoek Ostende: A gap in digging: the Talpidae of Petersbuch 28 (Germany, Early Miocene). Paläontologische Zeitschrift 89, 2015, S. 563–592, doi:10.1007/s12542-014-0228-2
- Leigh Van Valen: New Paleocene Insectivores and Insectivore classification. Bulletin of the American Museum of Natural History 135, 1967, S. 217–284
- Alphonse Milne-Edwards in Armand David: Rapport adressé a MM les professeurs-administrateurs du Muséum d'Histoire Naturelle. Nouvelles archives du Muséum d'histoire naturelle 7, 1871, S. 75–100 (S. 92) ()
- Alphonse Milne-Edwards: Mémoir sur la faune mammalogique du Tibet Oriental, et principalement de la principauté de Moupin. In: Henri Milne-Edwards und Alphonse Milne-Edwards (Hrsg.): Recherches pour servir à l'histoire naturelle des mammifères: comprenant des considérations sur la classification de ces animaux. Paris, 1868–1872, Tome Premier – Texte. S. 231–379 (S. 272–277) (), Tome Second – Atlas. Tafel 40 (Abb. 1) und Tafel 40A (Abb. 1) ()
- Oldfield Thomas: On the collection of mammals from the Tsin-ling mountains, Central China, presented by Mr. G. Fenwick Owen to the National Museum. Annals and Magazine of Natural History 8 (10), 1912, S. 395–403 ()
- Oldfield Thomas: On mammals collected in the provinces of Sze-chwan and Yunnan, W. China, by Mr. Malcolm Anderson. Proceedings of the Zoological Society 1911, S. 48–50 ()
- Oldfield Thomas: The Duke of Bedford's zoological exploration of Eastern Asia. – XV. On the mammals from the provinces of Szechwan and Yunnan, Western China. Proceedings of the Zoological Society 1912, S. 127–141 ()
- Oldfield Thomas: On mammals from the Yunnan Highlands collected by Mr. George Forrest and presented to the British Museum by Col. Stephenson R. Clarke D.S.O. Annals and Magazine of Natural History 9 (10), 1922, S. 391–406 ()
- Glover M. Allen: New Chinese insectivores. American Museum Novitates 100, 1923, S. 1–11
- Glover M. Allen: The mammals of China and Mongolia. Natural History of Central Asia Volume XI, Part I, American Museum of Natural History, 1938, S. 1–620 (S. 63–64) ()
- Wilfred Hudson Osgood: Variable dentition in a Chinese insectivore. Field Museum of Natural History Zoological Series 20, 1937, S. 365–368 ()
- J. R. Ellerman und T. C. S. Morrison-Scott: Checklist of Palaearctic and Indian Mammals 1758 to 1946. Zweite Ausgabe, London, 1966, S. 1–809 (S. 32) ()
- Robert S. Hoffmann: A review of the shrew moles (Genus Uropsilus) of China and Burma. Journal of the Mammalogical Society of Japan 10 (2), 1984, S. 69–80
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. A taxonomic and geographic Reference. 2 Bände. 3. Auflage. Johns Hopkins University Press, Baltimore MD 2005, ISBN 0-8018-8221-4 ()
- Gerhard Storch und Thomas Dahlmann: Desmanelia rietscheli, ein neuer Talpide aus dem Obermiozän von Dorn-Dürkheim 1, Rheinhessen (Mammalia, Lipotyphla). Carolinea – Beiträge zur naturkundlichen Forschung in Südwestdeutschland 58, 2000, S. 65–69 ()
- Barbara Rzebik-Kowalska: The Pliocene and Early Pleistocene Lipotyphla (Insectivora, Mammalia) from Romania. Acta zoologica cracoviensia 45 (2), 2002, S. 251–281
- Lars W. van den Hoek Ostende und Oldrich Fejfar: Erinaceidae and Talpidae (Erinaceomorpha, Soricomor-pha, Mammalia) from the Lower Miocene of Merkur-Nord (Czech Republic, MN 3). Beiträge zur Paläontologie 30, 2006, S. 175–203
- Reinhard Ziegler: Talpidae (Insectivora, Mammalia) aus dem Oberoligozän und Untermiozän Süddeutschlands. Stuttgarter Beiträge zur Paläontologie Serie B 167, 1990, S. 1–80
- Thomas Mörs: Die Säugetiere oberoligozänen Fossillagerstätte Rott bei Bonn (Rheinland). Decheniania 149, 1996, S. 205–232
- Johannes Klietmann, Brian D. Metscher, Lars W. van den Hoek Ostende, Doris Nagel und Michael Rummel: First record of an upper deciduous molar in Desmanella (Uropsilinae, Talpidae, Mammalia). Geobios 46, 2013, S. 503–510, doi:10.1016/j.geobios.2013.07.005
- Reinhard Ziegler: Talpiden (Mammalia, Insectivora) aus dem Orleanium und Astaracium Bayerns. Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie 25, 1985, S. 131–175
- Reinhard Ziegler: Moles (Talpidae) from the late Middle Miocene of South Germany. Acta Palaeontologica Polonica 48 (4), 2003, S. 617–648
- Burkart Engesser: Die obermiozäne Säugetierfauna von Anwil (Baselland). Tätigkeitsbericht der naturforschenden Gesellschaft Baselland 28, 1972, S. 37–363 (S. 115–120) ()
- Reinhard Ziegler: Miocene Insectivores from Austria and Germany - An Overview. Beiträge zur Paläontologie 30, 2006, S. 481–494
- Reinhard Ziegler: Insectivores (Lipotyphla) and bats (Chiroptera) from the Late Miocene of Austria. Annalen des Naturhistorischen Museums Wien 107 A, 2006, S. 93–196
- Florentin Cailleux, Lars W. van den Hoek Ostende und Peter Joniak: The Late Miocene Talpidae (Eulipotyphla, Mammalia) from the Pannonian Region, Slovakia. Journal of Paleontology 98 (1), 2024, S. 128–151, doi:10.1017/jpa.2023.95
- David L. Harrison und Barbara Rzebik-Kowalska: A note on the occurrence of Desmanella cf. dubia Rümke, 1976 (Insectivora: Talpidae: Uropsilinae) in the Lower Pliocene of Podlesice, Poland. Cranium 11 (1), 1994, S. 3–6
- Vicente D. Crespo, Rafael Marquina-Blasco, Francisco Javier Ruiz-Sánchez und Plini Montoya: An unusual insectivore assemblage from the early Miocene of southwestern Europe: The talpids and dimylids from the Ribesalbes–Alcora Basin (Spain). Comptes Rendus Palevol 18, 2019, S. 407–416, doi:10.1016/j.crpv.2019.03.003
- C. G. Rümke: A new Desmanella species (Talpidae, Insectivora) from the Turolian of Concud and Los Mansuetos (Prov. of Teruel, Spain). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen (physical sciences) 77 (4), 1974, S. 359–374
- Antonio García-Alix, Marc Furió, Raef Minwer-Barakat, Elvira Martínsuárez und Matthijs Freudenthal: Environmental control on the biogeographical distribution of Desmanella (Soricomorpha, Mammalia) in the Miocene of the Iberian Peninsula. Palaeontology 54 (4), 2011, S. 753–762, doi: 10.1111/j.1475-4983.2011.01062.x
- Jean-Yves Crochet: Insectivores Pliocènes du Sud de la France (Languedoc-Roussillion) et du Nord-Est de l'Espagne. Palaeovertebrata 16 (3), 1986, S. 145–171
- Wang Xiao-Ming, Qiu Zhu-Ding, Li Qiang, Yukimitsu Tomida, Yuri Kimura, Zhijie Jack Tseng und Wang Hong-Jiang: A New Early to Late Miocene Fossiliferous Region In Central Nei Mongol: Lithostratigraphy and Biostratigraphy In Aoerban Strata. Vertebrata Palasiatica 47 (2), 2009, S. 111–134
- Lars W. van den Hoek Ostende, Peter Joniak, Bora Rojay, Cathelijn Aten, Melike Bilgin und Pablo Pelaez-Campomanes: Early Miocene insectivores of Gökler (Kazan Basin, Central Anatolia, Turkey). Palaeobiodiversity and Palaeoenvironments 99, 2019, S. 701–722, doi:10.1007/s12549-019-00396-1
- A. V. Lopatin: Early Miocene small mammals from the North Aral Region (Kazakhstan) with special reference to their biostratigraphic significance. Paleontological Journal 38 (suppl 3), 2004, S. S217–S323
- Oleg G. Bendukidze, Hans de Bruijn und Lars W. van der Hoek Ostende: A revision of Late Oligocene associations of small mammals from the Aral Formation (Kazakhstan) in the National Museum of Georgia, Tbilissi. Palaeodiversity 2, 2009, S. 343–377
- Varun Parmar, Rigzin Norboo und Rahul Magotra: First record of Erinaceidae and Talpidae from the Miocene Siwalik deposits of India. Historical Biology 35 (2),2023, S. 276–283, doi:10.1080/08912963.2022.2034806
- William W. Korth, Clint A. Boyd, Robert J. Emry und Jeff J. Person: Additional small mammals from the Oligocene Brule Formation (Orellan-Whitneyan) of Southwestern North Dakota. Paludicola 14 (2), 2023, S. 57–74 ()
- Robert W. Wilson: Early Miocene rodents and insectivores from northeastern Colorado. University of Kansas, Paleontological Contributions Vertebrata 7, 1960, S. 1–92 (S. 38–42)
- William W. Korth: Fossil small mammals from the Harrison Formation (Late Arikareean: Earliest Miocene), Cherry County, Nebraska. Annals of the Carnegy Museum 61 (2), 1992, S. 69–131 ()
- IUCN: The IUCN Red List of Threatened Species. ()
Weblinks
- Gefährdungsgrad der einzelnen Arten in der Roten Liste gefährdeter Arten der IUCN.