Zilie
Als Zilie oder Cilium (lat. cilium ‚Wimper‘) bezeichnet man eine besondere Form des Zellfortsatzes bei Zellen von eukaryoten Organismen. Die zylindrische, 5–10 µm lange und etwa 250 nm schlanke Ausstülpung der Zellmembran enthält neben Zytoplasma ein röhrenförmiges Skelett aus Bündeln von Mikrotubuli, das sogenannte Axonem. Dieses Gerüst besteht aus neun kreisförmig angeordneten Mikrotubuli-Dubletts (9×2), in deren Mitte zwei zentrale Mikrotubuli liegen können (9×2+2) oder nicht (9×2+0). Man unterscheidet danach
- Sekundäre Zilien mit zentralen Mikrotubuli, Bauplan 9×2+2, die zumeist aktiv bewegbar, motil sind.

- Solche werden auch Kinozilien, Flimmerhärchen oder Wimpern genannt und mit den ähnlich aufgebauten, etwas längeren Geißeln der Eukaryonten als Undulipodien („wellenschlagende Füßchen“) zusammengefasst. Während einzelne Zellen durch Bewegungen ihrer Geißeln sich selber fortbewegen, bewegen miteinander verbundene Epithelzellen mittels ihrer Flimmerhärchen in abgestimmter Aktion beispielsweise Flüssigkeiten in den Bronchien oder ein Ei im Eileiter über die Epithelfläche.
- • Möglich wird die Eigenbeweglichkeit sekundärer Zilien erst durch eine Ausrüstung mit zusätzlichen Proteinen, welche die Mikrotubulipaare untereinander verbinden (Nexin-Band, Radialspeichen) und gegeneinander verschieben (Dynein-Arme). Fehlen diese, sind auch Zilien mit 9×2+2 Muster nicht motil, so etwa die Geruchsrezeptoren tragenden Zilien[1] der Riechzellen.
- Primäre Zilien ohne zentrale Mikrotubuli, Bauplan 9×2+0, die in der Regel nur passiv bewegbar sind.

- Womöglich tragen alle Zellen des menschlichen Körpers je eine solche primäre Zilie, die während der embryonalen Entwicklung und bei der Differenzierung von Geweben eine wichtige Rolle spielt und als Mechano- oder Chemosensor dienen kann.
- • Doch schließt ein (primärer) Aufbau ohne zentral gelegene Mikrotubuli eine aktive Beweglichkeit nicht prinzipiell aus. Die nodalen Zilien – benannt nach dem embryonalen Primitivknoten (lat. nodus ‚Knoten‘), einer Bildung während der Gastrulaphase der Embryonalentwicklung von Säugern, wo dieser Zilientyp entdeckt wurde – beispielsweise sind 9×2+0 gebaut und dennoch motil. Ihre Aktivität erzeugt einen asymmetrischen periembryonalen Fluss, der zur seitendifferenten Expression von Signalproteinen (wie Nodal) führt und darüber die Lateralisierung innerer Organe bewirkt.
Das Forschungsinteresse galt lange Zeit fast ausschließlich den durch ihre Beweglichkeit auffälligen Kinozilien, während man die unscheinbaren primären Zilien zwar sah, aber wenig beachtete. Seitdem sich jedoch ab etwa 2000 zeigte, dass die Entwicklung des tierischen Organismus und die Funktion vieler seiner Organe entscheidend von der Anwesenheit gesunder primärer Zilien abhängt, werden diese intensiv untersucht, beziehungsweise die als Ziliopathien damit verknüpften Erkrankungen, etwa ein Kartagener-Syndrom beim Menschen.
Zilien kommen bei vielen Eukaryonten vor. Im Tierreich sind sie auf fast allen Zelltypen zu finden, bei Pflanzen allerdings selten, insbesondere nicht in Blütenpflanzen, beispielsweise aber in Palmfarnen vorhanden. Bei Fadenwürmern und Gliederfüßern werden nur primäre Zilien in einigen Nervenzellen gefunden. Einzeller wie Wimpertierchen besitzen nur sekundäre Kinozilien.
Sekundäre Zilien
Aufbau und Vorkommen
Jede Zilie ist außen von einer Plasmamembran umgeben. Innen liegt im Zytoplasma als besondere Bildung des Zytoskeletts das sogenannte Axonem aus Bündeln feiner Tubulin-Hohlfasern, der Mikrotubuli. Bei sekundären Zilien sind diese nach dem 9×2+2-Muster angeordnet: In der Mitte der Zilie liegen zwei einzelne Zentraltubuli (Singletts) in einer scheidenartigen Umhüllung (Zentralscheide). Diese beiden zentralen Einzeltubuli sind kreisförmig umgeben von neun Doppeltubuli (Dubletts), die je aus einem A- und einem damit verschmolzenen B-Tubulus bestehen.
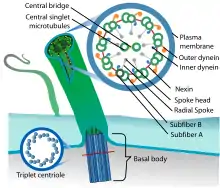
An jedem A-Tubulus befinden sich Paare von armartigen Strukturen (Dyneinarme), die zum B-Tubulus des benachbarten Dubletts gerichtet sind. Die im Kreis angeordneten Dubletts werden sowohl durch Nexinbindeglieder ringförmig untereinander verbunden als auch durch sogenannte Radialspeichen je mit den umhüllten Zentraltubuli.
Alle zwanzig Mikrotubuli gehen von einem Basalkörperchen aus, dem Bildungszentrum der Zilie an der Zilienbasis. Fasern der Übergangszone (Transitionszone) verankern dieses Kinetosom in der Zellmembran und setzen die membranumhüllte Zilie mit dem Axonem vom übrigen Zellkörper so ab, dass von einer Kompartimentierung gesprochen werden kann.
Der Schlag einer sekundären Zilie als aktive Bewegung wird durch energiebedürftige Verspannungen der Mikrotubuli im Inneren der Zilie hervorgebracht. Der Bewegungsprozess kann als ein Gleitmechanismus („sliding filament mechanism“) beschrieben werden. Dabei stellen Arme aus Dynein, stets am A-Tubulus verankert, mit ihren Spitzen Kontakte jeweils zum B-Tubulus des nächsten Nachbardubletts her und können mit dieser Verbindung eine ATP-abhängige Verschiebung der Dubletts gegeneinander bewirken, sodass eine Biegung erzeugt wird. Nexin, ein stark dehnbares Protein, hält benachbarte Dubletts während dieses Gleitvorgangs beieinander.
Derartige Kinozilien sind öfters in großer Anzahl an der Zelloberfläche von Einzellern oder Vielzellern ausgebildet. Geißeln und Zilien werden wissenschaftlich auch unter dem Oberbegriff Undulipodien (Einzahl Undulipodium) zusammengefasst, aufgrund des gleichen Bauprinzips.

Einige Wimpertierchen besitzen Gruppen von Zilien, die miteinander in Verbindung stehen. Solche Gruppierungen von Fortsätzen werden auch Cirrus (lat. für Locke) genannt.
Gelegentlich werden Zilien nicht deutlich genug von Mikrovilli unterschieden. Diese jedoch tragen kein Gerüst aus Mikrotubuli, sondern sind mit Aktinfilamenten ausgerüstet, darüber hinaus anderen Ursprungs und in der Regel nur passiv beweglich. Mikrovilli, nicht Zilien, befinden sich beispielsweise im Darm von Säugetieren, wo sie der Oberflächenvergrößerung dienen. Bewegt wird der Nahrungsbrei hier über die Peristaltik von Muskelzellen. Ein anderes Beispiel für fälschlich als Zilien bezeichnete Zellfortsätze sind die Bildungen der Haarzellen im Innenohr. Die früher als Stereozilien bezeichneten, reizaufnehmenden Fortsätze dieser Sinneszellen sind ebenfalls Mikrovilli und werden heute daher als Stereovilli bezeichnet. Eine Zilie existiert bei der menschlichen Haarzelle nur in der embryonalen Anlage und degeneriert während der Entwicklung.
Nicht zu verwechseln ist die Zilie mit den Flagellen der Bakterien. Diese sind wesentlich schlanker, bestehen vollständig aus Protein (Flagellin) und werden nicht von einer Membran umgeben. Auch liegt ihrer Arbeitsweise ein völlig anderes Prinzip zugrunde (nämlich eine Rotation ähnlich einer Schiffs-Schraube).
Bewegung und Funktion
Kinozilien werden nur selten allein gefunden, sondern meist in größerer Anzahl und oft in Reihen oder Feldern auf einer Zelle. Das koordinierte, ruderartige Schlagen der beweglichen Zilien dient folgenden Funktionen:
- Fortbewegung der einzelnen Zelle, so bei Wimpertierchen, zahlreichen Larvenstadien kleinerer, wasserbewohnender Tiere oder auch dem Spermium höherer Tiere
- Herbeistrudeln von Nahrungsteilchen
- Transport von Partikeln und Flüssigkeiten innerhalb eines Organismus, beispielsweise durch das Flimmerepithel in den Luftwegen ein Transport von Schleim und Fremdstoffen aus den Bronchien oder durch zilientragende Epithelzellen im Eileiter der Transport einer Eizelle.

Kinozilien oder Wimpern sind sozusagen flexible Miniatur-Ruder, die im Gegensatz zu Geißeln uniplanar (in einer Ebene) schlagen. Ebene und Schlagrichtung liegen für jede Zilie fest. Während des kraftvollen Vorschlages ist die Zilie nahezu gestreckt. Der langsamere Rückschlag erfolgt gekrümmt, wobei eine Biegungswelle von der Zilienbasis bis zur Zilienspitze läuft und die Zilie bei geringem Widerstand des umgebenden Mediums wieder in ihre Ausgangsposition zurückgeführt wird. Dabei kann gleichzeitig eine Kurve im Raum durchlaufen werden.
Jede Zilie einer Zilienreihe schlägt gegenüber der vorhergehenden um einen Bruchteil später. Man nennt dies eine metachrone Bewegung. Der kollektive Bewegungsablauf ist dabei wellenförmig, vergleichbar einem im Wind wogenden Kornfeld.
Die Schlagfrequenz einer Zilie kann je nach Umweltbedingungen zwischen 5 und 20 Hz betragen. Dabei gibt es Faktoren, die die Frequenz beschleunigen können, wie Wärme oder auch einige Medikamente. Andere Faktoren hemmen dagegen die Frequenz oder führen gar zum Stillstand, wie beispielsweise Nikotin oder ein bakterieller Infekt.
Primäre Zilien

Im Unterschied zu den meist aktiv bewegbaren, „motilen“ sekundären Zilien sind primäre Zilien zumeist nur passiv beweglich. In der Regel existiert pro Zelle nur eine solche „nicht-motile“ Zilie, die nach dem 9×2+0 Schema gebildet ist – das zentrale Paar fehlt also.
Fast alle Zellen von Wirbeltieren besitzen eine einzelne nichtmotile Zilie, auch „primäres Cilium“ genannt, welches lange in der Forschung vernachlässigt wurde. Diese primären Zilien stellen oft einen sensitiven Fortsatz der Zelle dar. Aus diesen nichtmotilen Zilien haben sich auch spezialisierte Strukturen herausgebildet; so ist beispielsweise das Außensegment von Photorezeptorzellen im Auge über ein spezialisiertes Cilium, das sogenannte Verbindungscilium, mit dem Innensegment verbunden.
Dagegen haben die Riechzellen, als Sinneszellen des Geruchs bei Wirbeltieren zugleich olfaktorische Nervenzellen, je bis zu zwanzig nichtmotile Zilien mit besonderen Geruchsrezeptoren; diese speziellen Nervenzellfortsätze sind allerdings nach dem 9×2+2 Schema sekundärer Zilien aufgebaut.[1][2]
Einzelnachweise
Literatur
- James R. Davenport, Bradley K. Yoder: An incredible decade for the primary cilium: a look at a once-forgotten organelle. In: Am J Physiol Renal Physiol. 289. Jahrgang, Nr. 6, 2005, S. F1159–1169, doi:10.1152/ajprenal.00118.2005, PMID 16275743 (physiology.org).
Siehe auch
Weblinks
- Denis Wheatley: Primary Cilia