Maulwürfe
Die Maulwürfe (Talpidae) sind eine Säugetierfamilie aus der Ordnung der Insektenfresser (Eulipotyphla). Sie umfassen mehr als 60 in Eurasien und Nordamerika verbreitete Arten, wobei der Europäische Maulwurf der einzige in Mitteleuropa lebende Vertreter ist. Die Tiere besitzen vielfältige Anpassungen an unterschiedliche Lebensweisen. Dementsprechend variabel ist auch ihr äußeres Erscheinungsbild. Allgemein kennzeichnend sind ein walzen- bis spindelförmiger Körper mit kurzem Hals, auf dem ein spitz zulaufender Kopf sitzt. Die meisten Arten, so die Eurasischen Maulwürfe und die Neuweltmaulwürfe, leben unterirdisch grabend. Ihre Vordergliedmaßen sind kräftig und schaufelartig gestaltet sowie weit nach vorn verlagert, die Handflächen zeigen beständig nach außen. Daneben gehören auch die wasserbewohnenden Formen wie die Desmane und der Sternmull dazu. Die langen Hinterbeine der Desmane tragen Schwimmhäute. Die oberirdisch vorkommenden Spitzmausmaulwürfe sowie die verschiedenen Spitzmulle hingegen ähneln äußerlich den Spitzmäusen. Allen Maulwürfen ist ein dichtes Fell mit überwiegend dunkler Fellfarbe zu eigen. Bis auf die Spitzmausmaulwürfe haben die meisten Vertreter keine äußerlich sichtbaren Ohren ausgebildet, ebenso wie die Augen häufig verkümmert sind. Eine Besonderheit stellen die Eimerschen Organe als feinfühliges Tastwerkzeug dar.
| Maulwürfe | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Europäischer Maulwurf (Talpa europaea) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Talpidae | ||||||||||||
| Fischer, 1817 |
Überwiegend genutzte Lebensbereiche bestehen aus offenen Landschaften und Flusstälern, aber auch aus Hochgebirgsregionen. Häufig ist das Vorhandensein einer mächtigen Humusschicht Voraussetzung. Das Verhalten ist nur bei wenigen Arten genauer erforscht. Sowohl die grabende als auch die schwimmende Lebensweise bedurfte einzelner spezieller körperlicher Anpassungen. Die unterirdisch grabenden Maulwürfe legen komplexe Tunnel und Gänge an, deren oberflächig kennzeichnendes Merkmal der Maulwurfshügel ist. Die Tunnel können mehrere Ebenen beanspruchen und enthalten Nestkammern. Bodennahe Gänge dienen meist der Nahrungssuche. Bei den wasserbewohnenden Arten erfolgt diese in stehenden und fließenden Gewässern. Zur Ruhe ziehen sich die Tiere in teils selbst gegrabene unterirdische Baue zurück. Dies trifft auch auf die Spitzmulle zu, über die Spitzmausmaulwürfe liegen kaum Informationen vor. Generell sind die Maulwürfe tag- und nachtaktiv. Mit wenigen Ausnahmen leben sie einzelgängerisch, jedes Individuum beansprucht hierbei ein eigenes Territorium. Die Tiere ernähren sich überwiegend von Wirbellosen. Die Paarung findet zumeist einmal jährlich statt. Nach einer gut einmonatigen Tragzeit kommen zwei bis sechs Jungen zur Welt. Der Nachwuchs erlangt dann spätestens im folgenden Jahr die Geschlechtsreife.
Die ersten Arten wurden bereits im Jahr 1758 wissenschaftlich eingeführt. Die Bezeichnung der Familie geht auf das Jahr 1817 zurück. In der Anfangszeit der Erforschung der Maulwürfe bestanden noch unsichere Abgrenzungen zu anderen grabenden Säugetieren. Vor allem im Verlauf des 19. und 20. Jahrhunderts wurde die innere Gliederung der Familie vielfach diskutiert. Eine weitgehende Klärung ließ sich aber erst Anfang des 21. Jahrhunderts mit Hilfe genetischer Untersuchungen bewerkstelligen. Die genetischen Daten setzen den Ursprung der Maulwürfe möglicherweise noch kurz vor der Kreide-Paläogen-Grenze an. Die ältesten Fossilnachweise gehören aber in das ausgehende Eozän. Zu diesem Zeitpunkt traten die Maulwürfe allerdings bereits sowohl in Eurasien als auch in Nordamerika auf. Innerhalb der Stammesgeschichte der Familie können mehrere ausgestorbene Linien unterschieden werden. Von den heute existierenden Arten sind mehrere in ihrem Bestand gefährdet. Hauptursachen hierfür finden sich in der Jagd und in der Lebensraumzerstörung.
Merkmale
Habitus



Die Maulwürfe sind generell eher kleine Tiere. Sie erreichen eine Kopf-Rumpf-Länge von 6,3 bis 24,0 cm und ein Gewicht von 6 bis 440 g. Die kleinsten Vertreter gehören zu den bodenlebenden Spitzmausmaulwürfen (Urotrichinae) sowie zum Amerikanischen Spitzmull (Neurotrichini), während der Russische Desman (Desmana) aus der Gruppe der semi-aquatisch angepassten Desmane (Desmanini) die größte Form repräsentiert. Die grabenden Maulwürfe als Kerngruppe der Familie, hier vor allem die Eigentlichen Maulwürfe (Talpini) und die Neuweltmaulwürfe (Scalopini), werden zwischen 8 und 23 cm lang sowie zwischen 20 und 220 g schwer. Die Schwanzlänge ist bei den Maulwürfen stark variabel: Bei den grabend lebenden Arten ist er vergleichsweise kurz, oft ist er nur mit wenigen Haaren bedeckt, die teils als Tasthaare fungieren und eine Orientierung nach hinten ermöglichen. Bei den spitzmausähnlichen Arten und den Desmanen weist der Schwanz mitunter die gleiche Länge wie der restliche Körper auf. Letztere haben einen partiell oder vollständig abgeplatteten Schwanz, der als Steuerruder im Wasser eingesetzt wird.[1]
Der Rumpf der Maulwürfe ist spindel- bis walzenförmig, der Kopf sitzt auf einem sehr kurzen Hals. Das Fell, das den gesamten Körper bedeckt mit Ausnahme der Füße und der Nasenspitze, besteht bei den unterirdisch lebenden Arten aus zwei Haarschichten, dem Wollhaar und dem Deckhaar. Bei den Desmanen kommen noch Leithaare hinzu. Das Wollhaar ist bei allen Maulwürfen fein und gewellt. Das Deckhaar umfasst zumeist gröbere Haare, die wie bei den Desmanen auch borstenartig sein können. Bei letzteren sind die Deckhaare in der Regel auch abgeplattet und verhindern so das Entweichen der Luft unter Wasser. Insgesamt ist das Fell sehr dicht. Es setzt sich bei einigen Neuweltmaulwürfen aus bis zu 3000 Haaren je Quadratzentimeter zusammen, bei den Desmanen steigt dies auf bis zu 20.300 an. Die Fellfärbung variiert von hellgrau über verschiedene Brauntöne bis schwarz. Häufig tritt ein seidiger Glanz auf. Die Bauchseite ist dabei nur wenig heller als die Oberseite. Alle Arten haben Hautdrüsen, die vorwiegend zur Absonderung von Schweiß dienen. Teilweise bedecken bis zu 600 solcher Drüsen einen Quadratzentimeter Haut. Am auffälligsten sind allerdings die Moschusdrüsen an der Schwanzwurzel der Desmane. Der Kopf wirkt durch die charakteristisch lang ausgezogene Nase zugespitzt. Die Augen der Maulwürfe sind sehr klein und teilweise von Haut überdeckt, Ohrmuscheln fehlen bei allen Arten mit Ausnahme der Spitzmausmaulwürfe.[1]
Die Gliedmaßen der Maulwürfe wirken kurz und enden jeweils in fünf bekrallten Fingern oder Zehen. Die Vordergliedmaßen der unterirdisch lebenden Arten sind zu Grabwerkzeugen umgebildet. Die mit der Handfläche nach außen gedrehten Hände zeigen sich schaufelförmig verbreitert. Die Gliedmaßen der Desmane tragen hingegen Schwimmhäute zwischen den Zehen und seitlich an den Kanten borstenartige Haare. Sie sind ebenso wie die kleinen schlanken Hände und Füße der Spitzmausmaulwürfe nur bedingt zum Graben geeignet, wenngleich auch erstere teilweise ihre eigenen Baue anlegen können.[1]
Schädel- und Gebissmerkmale

Der Schädel der Maulwürfe ist relativ einheitlich gebaut. Er weist eine typisch langgestreckte Form auf mit einem breiten Hirnschädel und einem schmalen Rostrum. Er wirkt dadurch keilartig. Das Längen-Breiten-Verhältnis liegt bei rund 2:1, es treten aber deutliche artliche Variationen in der Breite des Rostrums auf. Eine in Aufsicht auffallende Einschnürung hinter der Orbita tritt außer bei den Desmanen nicht auf. In Längsansicht hebt sich der Hirnschädel nur leicht von der Schnauzenregion ab. Markante Knochenrippeln wie ein Scheitelkamm oder Hinterhauptskamm fehlen, auch hier bilden die Desmane eine Ausnahme. Die einzelnen Schädelknochen verwachsen relativ früh in der Individualentwicklung. Der Jochbogen ist bei allen Maulwürfen entwickelt, jedoch eher schwach ausgeprägt. Zudem kragt er nicht weit aus, vielmehr verläuft er parallel zur Schädelseite. An der Schädelunterseite ist die Paukenblase entweder mit dem Schädel fest verwachsen wie bei den Eurasischen Maulwürfen (Talpa), den Westamerikanischen Maulwürfen (Scapanus), dem Ostamerikanischen Maulwurf (Scalopus) beziehungsweise dem Weißschwanzmaulwurf (Parascaptor) oder über Bindegewebe mit ihm verbunden, wie es beim Amerikanischen Spitzmull oder beim Sternmull der Fall ist. In der Regel sind die Paukenblasen aber klein und rundlich. Die Gehörknöchelchen zeigen bei den wenigsten Arten eine auffallende Vergrößerung, für die beiden letztgenannten Formen sind sie dagegen außerordentlich klein. Lediglich beim Weißschwanzmaulwurf und beim Kurzgesichtmaulwurf (Scaptochirus) weist der Hammer einen deutlich aufgeblähten Kopf auf. Sowohl die Verbindung der Paukenblase mit der Schädelunterseite als auch die Ausprägung der Gehörknöchelchen spiegeln die stärkeren oder schwächeren Anpassungen an ein Leben im Untergrund wider, was wiederum Rückwirkung auf das Hörvermögen hat. Demnach nehmen die Tiere, die weitgehend oberirdisch leben vermehrt höher-, jene, die unterirdisch leben, stärker niederfrequente Geräusche wahr.[2][3] Der Unterkiefer ist bei allen Maulwürfen schlank gebaut. Der Kronenfortsatz zeigt sich häufig gerundet, der Gelenkfortsatz schlank und der Winkelfortsatz kräftig. Die Verbindung mit dem Schädel ist scharnierartig, so dass nur Auf- und Abwärtsbewegungen möglich sind.[4][1]
| Gruppe | Zahnformel | Zahnanzahl | I1 | C1 | Gebissaustausch |
|---|---|---|---|---|---|
| Condylurini | 44 | groß | klein | pränatal | |
| Desmanini | 44 | groß | klein | pränatal | |
| Neurotrichini | 36 | groß | klein | ?? | |
| Scalopini | 36–44 | groß | klein | postnatal | |
| Scaptonychini | 42 | klein | groß | ?? | |
| Talpini | 40–44 | klein | groß | pränatal | |
| Urotrichini | 36–38 | groß | klein | postnatal | |
| Uropsilinae | 34–38 | klein | klein | postnatal |








Das Gebiss der Maulwürfe ist stark variabel, je nach Art sind 34 bis 44 Zähne ausgebildet. Die Zahnformel lautet demnach . Einige Arten hauptsächlich aus der Gruppe der Neuweltmaulwürfe und der Eigentlichen Maulwürfe, darunter auch der Europäische Maulwurf, haben mit 44 Zähnen die ursprüngliche Zahnzahl der Höheren Säugetiere bewahrt. Die größten Unterschiede finden sich im vorderen Gebiss. Dies betrifft allerdings nicht nur die schwankende Zahnanzahl, sondern auch die Ausprägung der Zähne. So ist bei den Eigentlichen Maulwürfen und dem Langschwanzmaulwurf (Scaptonychini) der obere Eckzahn stark vergrößert, alle anderen Vertreter mit Ausnahme der Spitzmausmaulwürfe haben einen hypertrophierten ersten Schneidezahn. Bei letzteren sind alle vorderen Zähne etwa gleich groß. Das hintere Gebiss mit den Prämolaren und den Molaren ähnelt dem anderer Insektenfresser. Die Kauflächen sind hier mit spitzen Höckern und scharfen Schmelzleisten versehen, was als Anpassung an die fleisch- und insektenfressende Lebensweise aufzufassen ist. Auf den Mahlzähnen ordnen sich die Haupthöckerchen in einem dilambdodonten (W-förmigen) Muster an. Bemerkenswert ist eine teils starke Oligodontie bei einigen Formen, insbesondere der Eurasischen Maulwürfe und der Ostasiatischen Maulwürfe (Mogera) innerhalb der Eigentlichen Maulwürfe. Häufig kann dies innerhalb des vorderen Gebisses und der Prämolaren festgestellt werden und betrifft sowohl fehlende als auch überzählige Zähne. Als weitere Auffälligkeit findet bei den Eigentlichen Maulwürfen, den Desmanen, dem Sternmull (Condylurini) der Wechsel vom Milchgebiss zum Dauergebiss schon vor der Geburt statt. Die Spitzmausmaulwürfe, Neuweltmaulwürfe und die Japanischen Spitzmulle (Urotrichini) hingegen zeigen einen nachgeburtlichen Zahnaustausch.[5][1]

Skelettmerkmale

Die Wirbelsäule setzt sich aus 7 Hals-, 13 bis 14 Brust-, 5 bis 7 Lenden-, 5 bis 6 Kreuzbein- und 8 bis 27 Schwanzwirbel zusammen. Die Anzahl der Wirbel vor dem Schwanz variiert somit zwischen 31 und 33, wobei letzteres überwiegt. Die Dornfortsätze der Hals- und Brustwirbel sind in der Regel schwach entwickelt. Die Brustwirbel tragen schlanke Rippen. Sofern das 14. Rippenpaar ausgebildet ist, zeigt es sich deutlich verkleinert und hat keine Funktion.[5][1]
Als teilweise hoch spezialisierte Tiere haben die Maulwürfe einige Besonderheiten im Skelettbau, was vor allem die Arme und den Schultergürtel betrifft. Die Veränderungen sind umso auffälliger, je stärker sich die einzelnen Arten an eine grabende Lebensweise angepasst haben. Generell sind die Knochen der Arme bei den grabenden Maulwürfen massiver und breiter als bei den terrestrisch oder semi-aquatisch lebenden Formen. Der Schultergürtel ist zumeist vorverlagert und liegt bei den grabenden Angehörigen oberhalb des letzten Halswirbels, Ausnahmen bilden hier die Desmane und Spitzmausmaulwürfe. Einzigartig unter den Säugetieren ist die gelenkige Verbindung zwischen dem Schlüsselbein und dem Oberarmknochen, genauer mit dem Großen Knochenvorsprung von letzterem. Die ansonsten übliche Artikulation von Oberarmknochen und Schulterblatt besteht lediglich noch bei den Spitzmausmaulwürfen. Dem Schlüsselbein kommt außerdem eine besondere Bedeutung zu, was die Position der Arme betrifft. Bei den bodenbewohnenden Spitzmausmaulwürfen ist dieses noch lang und schlank, wobei gegenüber den Spitzmäusen bereits einzelne Kürzungen vorliegen. Dennoch sind die Vorderbeine bei diesen unterhalb des Körpers positioniert, was eine terrestrisch laufende Fortbewegung ermöglicht. Von den wasserlebenden Desmanen hin zu den bodengrabenden Formen verkürzt sich das Schlüsselbein immer weiter, bis es bei den Eurasischen Maulwürfen und den Neuweltmaulwürfen nahezu eine Würfelform erreicht. Dies bewirkt durch die direkte Artikulation mit den Oberarmknochen, dass die Arme bei den grabenden Maulwürfen weit vor dem Brustkorb neben dem Kopf liegen und weitgehend in die Körperkontur eingebaut sind, was wiederum die Grabbewegungen im Untergrund erleichtert. Ähnlich stark verändert ist das Schulterblatt. Es wird von den bodenbewohnenden hin zu den grabenden Vertretern immer schlanker und verliert einzelne markante Fortsätze. Unter anderem bildet sich bei den grabenden Maulwürfen das Acromion zurück, ebenso wird das Metacromion, ein Fortsatz des Acromion, reduziert. Einer großen Veränderung unterliegt der Oberarmknochen, der bei den Spitzmausmaulwürfen und den Desmanen relativ schlank und rund im Querschnitt ausgeprägt ist, bei den grabenden Maulwürfen hingegen kurz und robust, er stellt hier den massivsten Knochen des gesamten Skeletts. Erreicht wird dies durch die Verbreiterung des Schaftes, einhergehend mit einer generellen Abplattung des Knochens, sowie einer extremen seitlichen Ausdehnung der Gelenkenden. Das Breiten-Längen-Verhältnis des Oberarmknochens liegt bei den Neuweltmaulwürfen und den Eurasischen Maulwürfen dadurch bei etwa 0,33 bis 0,45, während es bei den Spitzmausmaulwürfen und den Desmanen 0,17 bis 0,22 beträgt. Der kurze und breite Oberarmknochen der grabenden Maulwürfe fungiert als Ansatzstelle für die starke Armmuskulatur, die das Fortbewegen im Untergrund ermöglicht. Allerdings verhindern die breiten Gelenkenden, hauptsächlich jene des Ellenbogengelenks, Drehungen in diesem Bereich, so dass nur Auf- und Abwärtsbewegungen möglich sind. Rotationen des Armes erfolgen aus der Schulter, wodurch sich der Oberarmknochen um seine Längsachse dreht. Trotz seiner Massivität sind am Oberarmknochen der grabenden Maulwürfe keine speziellen Knochenverdichtungen vorhanden.[6] Auffallend am Unterarm ist vor allem der lang ausgezogene obere Gelenkfortsatz der Elle (Olecranon) als Teil des Ellenbogengelenks bei den grabenden Maulwürfen, der wiederum als Anker der Armmuskulatur fungiert. Bei den eher terrestrisch lebenden Formen wird der Fortsatz deutlich kürzer.[7][8][9] Analog zu den Armknochen sind jene der Hand bei den grabenden Maulwürfen ebenfalls stark verdickt im Vergleich zu den anderen Vertretern. Hier kommt zusätzlich ein sichelförmiges Sesambein vor, der sogenannte „Vordaumen“ (Präpollex) oder das Os falciforme. Es setzt seitlich am Kahnbein an. Bei den Eurasischen Maulwürfen und bei den Neuweltmaulwürfen erreicht es durchaus die Länge der Handwurzel zuzüglich des Mittelhandknochens und kommt rudimentär bei den Spitzmullen und den Desmanen vor, fehlt aber vollständig bei den Spitzmausmaulwürfen. Das Os falciforme verbreitert als „sechster Finger“ die Handfläche und unterstützt das Graben im Untergrund. Es wird bereits embryonal angelegt.[10][11][12][1]
Im Gegensatz zu den Armen sind die Beine bei den meisten Maulwürfen weniger spezialisiert. Oberschenkelknochen und Schienbein erreichen zumeist die gleiche Länge. Eine Ausnahme bilden hier die Desmane, bei denen ersterer nur halb so lang wird wie letzterer. Das Schienbein verbindet sich mit dem Wadenbein auf Höhe des Mittelschaftes. Die einzelnen Fußknochen sind lang und schlank. Bei einigen Vertretern wie den Eurasischen Maulwürfen kommt am Fuß ein kleines Sesambein vor, das dem „Vorzeh“ (Prähallux) entspricht. Seine Funktion ist unklar, es könnte aber genetisch an die Bildung des Präpollex gekoppelt sein.[5][10][1]
Weichteilanatomie
Über die inneren Organe der Maulwürfe wurden bisher nur wenige Informationen bereitgestellt. Die Lungen sind durchschnittlich größer als bei den meisten Säugetieren. Für die Eurasischen Maulwürfe beträgt ihr Anteil am Körpergewicht fast ein Fünftel, bei den Desmanen ein Zehntel. Im Vergleich dazu erreichen sie bei den Spitzmäusen weniger als ein Zehntel.[1]
Der Verdauungstrakt zeigt wie bei allen Insektenfressern einen eher simplen Bau. Dies trifft auch auf den Magen zu, der in seiner Verlängerung des Magenpförtners in etwa dem der Spitzmäuse gleicht, jedoch durch eine recht auffällige Einschnürung zwischen diesem und dem Magenkörper Ähnlichkeiten zu dem der Igel aufweist. Im Vergleich zu den Igeln ist die innere Oberfläche vergrößert, was sich in einer höheren Anzahl an Drüsen im Magengrund auszeichnet. Die Verteilung der Drüsen, die die Magenschleimhaut bilden, ist sehr charakteristisch für die Maulwürfe. So entstehen etwa 83 bis 89 % durch die Fundusdrüsen, 4 bis 6 % durch die Pylorusdrüsen und 1 bis 3 % durch die Kardialdrüsen, der Rest in Übergangsbereichen. Bei den Spitzmäusen ist der Anteil der Fundusdrüsen geringer. Insgesamt beansprucht der Magen 15 bis 25 % der Kopf-Rumpf-Länge, er ist am kürzesten beim Amerikanischen Spitzmull, intermediär bei den grabenden Maulwürfen und am längsten beim Sternmull.[13] Der Darm bildet einen einfachen Schlauch. Er ist verglichen mit der Körperlänge sehr kurz, der Blinddarm fehlt.[1]
Im Bau des Fortpflanzungstraktes zeigen Maulwürfe ebenfalls Übereinstimmungen mit anderen Insektenfressern: Generell ist der Geschlechtstrakt der Männchen sehr groß, er kann bis zu 14 % des Gesamtkörpergewichts ausmachen.[14] Die Hoden liegen außerhalb der Bauchhöhle in hodensackähnlichen Hautfalten nahe der Schwanzwurzel, den Cremasterfalten. Im Verhältnis zum Körpergewicht sind die Hoden mitunter sehr groß, beim Sternmull nehmen sie entsprechend fast 4,8 % ein. Der Penis wird von einer weiten Vorhaut umgeben und ist schwanzwärts gerichtet. Es kommt ein Penisknochen vor, der jedoch sehr klein ist und beim Amerikanischen Spitzmull sowie beim Sternmull vollständig fehlt. Die Weibchen zeigen teilweise eine Vermännlichung der äußeren Geschlechtsorgane. So ist analog zum Penis der Männchen die Klitoris tief in die Vorhaut eingebettet. Die Clitoriseichel ragt fast genauso weit auf wie das entsprechende männliche Gegenstück, einige Arten verfügen über einen Klitorisknochen. Zudem kommt eine zweilappige Prostata vor. Die Gebärmutter besitzt typischerweise zwei Hörner (Uterus bicornis).[15][16][1]
Eimersches Organ und weitere Sinnesorgane

Die Schnauzenregion der Maulwürfe ist reich an Tastsinneszellen, den sogenannten Eimerschen Organen. Benannt nach Theodor Eimer handelt es sich hierbei um buckelartige Aufwölbungen mit einem Durchmesser 40 bis 120 μm. Die Aufwölbungen bestehen aus umgestülpter Haut und markieren die Enden von Nervenbahnen, einerseits freie Nervenenden, andererseits solche aus Merkel-Zellen. Damit können die Tiere wahrscheinlich nicht nur Tastreize, sondern auch elektrische Reize wahrnehmen und damit schwache elektrische Felder fühlen, die bei der Muskelbewegung der Beute entstehen. Am ausgeprägtesten findet sich dieses Organsystem beim Sternmull, der 22 radial um die Nase angeordnete fingerförmige Fortsätze besitzt. Auf diesen befinden sich schätzungsweise bis zu 26.000 einzelne Rezeptoren. Bei anderen Maulwürfen sind die Eimerschen Organe weniger komplex ausgebildet beziehungsweise abweichend strukturiert. Ihnen fehlen die fingeartigen Tentakel des Sternmulls, so dass sich die Buckel direkt auf dem Nasenspiegel anordnen, der in mehrere Parzellen aufgeteilt ist. Hierbei können zwischen 1300 und über 2400 einzelne Rezeptoren vorkommen. Bei einigen Formen wie dem Kurzgesichtmaulwurf oder dem Ostamerikanischen Maulwurf sind die Rezeptoren möglicherweise zurückgebildet. Bei den Desmanen sprießen zwischen den teils merklich verkleinerten Buckeln feinfühlige Vibrissen. Lediglich die Spitzmausmaulwürfe zeigen keinerlei Hinweise auf die Ausbildung von Eimerschen Organen.[17][18][19]
Genetische Merkmale
| Gattung | Chromosomensatz | Fundamentale Anzahl |
|---|---|---|
| Condylurini | 2n = 34 | FNa = 64 |
| Desmanini | 2n = 32–42 | FNa = 64–68 |
| Neurotrichini | 2n = 38 | FNa = 72 |
| Scalopini | 2n = 34 | FNa = 56–64 |
| Scaptonychini | 2n = 34 | FNa = 64 |
| Talpini | 2n = 30–48 | FNa = 52–64 |
| Urotrichini | 2n = 34 | FNa = 62–64 |
| Uropsilinae | 2n = 34–36 | FNa = 46–54 |
Der diploide Chromosomensatz der Maulwürfe besteht aus 30 bis 48 Chromosomenpaaren. Bei einem größeren Teil der Arten lautet er 2n = 34, was als ursprüngliche Form angesehen wird. Er findet sich in dieser Version bei den als eher ursprünglich geltenden Spitzmausmaulwürfen, aber auch bei einigen Spitzmullen, dem Sternmull und den an das Graben angepassten Neuweltmaulwürfen wieder. Zudem tritt er bei den gleichfalls unterirdisch lebenden Eigentlichen Maulwürfen auf, unterlag hier aber stärkeren Abwandlungen. In dieser Gruppe ließen sich auch die bisher ermittelten Extremwerte feststellen. Demnach kommt der einfachste Chromosomensatz bei den Ostasiatischen Maulwürfen vor, der umfangreichste beim Kurzgesichtmaulwurf. Stärkere Abweichungen vom Ursprungstypus finden sich bei den Desmanen und dem Amerikanischen Spitzmull. Die ursprüngliche Annahme einer weitgehend konservativen Entwicklung der Maulwürfe bezüglich ihrer cytogenetischen Eigenschaften lässt sich durch die zunehmenden Analysen seit den 2000er Jahren nicht bestätigen. Allerdings ist bisher ungeklärt, warum innerhalb der grabenden Formen die Neuweltmaulwürfe eine ausgesprochene Stabilität bezüglich des Chromosomensatzes aufweisen, die Eigentlichen Maulwürfe hingegen nicht. Ähnlich variantenreich erweist sich die fundamentale Anzahl, also die Anzahl der Arme der Autosomenpaare, die zwischen 46 und 72 schwankt. Hierbei ist eine bimodale Verteilung auffällig, bei der ein Schwerpunkt um 54, der andere bei Werten um 62 bis 64 schwankt. Als Ursache kommen Neuarrangements in Form von Mutationen in Frage, bei denen einarmige Autosomen zunehmend in zweiarmige umgewandelt werden. Nahezu alle Vertreter der Familie besitzen ein großes metazentrisches X-Chromosom und ein kleines fleckenartiges Y-Chromosom.[20][21][22][23][24] Das Mitogenom der Maulwürfe ist bei den bisher untersuchten Arten relativ einheitlich aufgebaut, die Anzahl der Basenpaare reicht von 16.530 bis 16.960.[25]
Verbreitung und Lebensraum
Die Maulwürfe sind ausschließlich auf der Nordhalbkugel beheimatet, namentlich in Eurasien und Nordamerika. In Eurasien besiedeln sie von der Iberischen Halbinsel ausgehend weite Teile des westlichen, mittleren, östlichen und südlichen Europas, des Weiteren treten sie von Westasien über Zentral- und Ostasien bis zur Japanischen Inselwelt auf. Nach Süden erstreckt sich das Vorkommen dort über das festländische Südostasien bis zur Malaiischen Halbinsel. Der nordamerikanische Verbreitungsschwerpunkt reicht vom südlichen Kanada bis in das nördliche Mexiko. Die Tiere bewohnen damit sowohl die gemäßigten Zonen, als auch subtropische und tropische Bereiche. Einige Formen sind auch an boreale Verhältnisse angepasst. Prinzipiell fehlen die Maulwürfe jedoch in polaren und subpolaren Regionen. Insgesamt bewohnen die Vertreter der Familie unterschiedliche Lebensräume. Sie reichen von offenen Wiesen- und Steppenarealen über Flusstäler bis hin zu Küstendünen und semiariden Landschaften. Mitunter werden auch Kultur- und Gartenflächen besiedelt. Einige Arten wie die Spitzmausmaulwürfe kommen in extremen Hochgebirgslagen bis zu 4600 m über dem Meeresspiegel vor. Zumeist bevorzugen die Maulwürfe Gegenden mit feuchten Böden und einer dicken Humusschicht. In sehr trockenen Gebieten oder Lebensräumen mit trockenen Jahresabschnitten beschränken sich die Tiere auf Bereiche mit feuchterem Untergrund oder Flussauen. In den mittleren Breiten neigen einige Arten wie der Europäische Maulwurf, der Ostamerikanische Maulwurf oder der Sternmull zu ausgedehnten Verbreitungsgebieten. Vor allem in den eher semiariden Habitaten in Westasien und dem südlichen Schwarzmeergebiet, aber auch in den tropischen Gebieten Südostasiens sind die Vorkommen der einzelnen Arten sehr begrenzt. Diese Regionen gelten als bedeutende Diversifikationszentren.[26][27][28] In einigen Bereichen des Verbreitungsgebietes bewohnen mehrere Maulwurfarten eine Region, etwa im Mittelmeergebiet oder auf den Japanischen Inseln sowie in Südostasien. Häufig unterscheiden sich die Arten dann in ihrer Größe und der Nutzung spezieller ökologischer Nischen. Ihr gemeinsames Auftreten ist daher nur bedingt syntopisch.[1]
Lebensweise
Ökomorphologische Typen


Die teils morphologische Vielgestaltigkeit der Maulwürfe reflektiert ihre unterschiedlichen Anpassungen an verschiedene Lebensräume. Prinzipiell können vier Ökomorphotypen unterschieden werden:[29]
- oberirdisch (terrestrisch): Spitzmausmaulwürfe
- teils wasserbewohnend (semi-aquatisch): Desmane, Sternmull
- teils unterirdisch (semi-fossorial): Japanische Spitzmulle, Amerikanischer Spitzmull, Langschwanzmaulwurf
- unterirdisch (fossorial): Eigentliche Maulwürfe, Neuweltmaulwürfe
Die unterschiedlichen ökologischen Anpassungen führen zu variierenden Lebensweisen. Die meisten Vertreter der Maulwürfe leben unterirdisch. Sie sind an diese Lebensweise mit ihrem walzenförmigen Körper, den vorverlagerten Vordergliedmaßen und den nach außen gedrehten sowie breiten und damit zu Grabwerkzeugen umgestalteten Händen angepasst. Da ausschließlich die Vorderbeine zum Graben eingesetzt werden, sind diese am stärksten entwickelt. Die Tiere verbringen den Großteil ihres Lebens in einem selbst gegrabenen, unterirdischen Gangsystem. Das Aushubmaterial wird dabei häufig in Form von Maulwurfshügeln zur Oberfläche gebracht. Je nach Art und Lebensraum können sich diese Gänge knapp unter der Oberfläche oder auch in größere Tiefen erstrecken beziehungsweise mehrere Ebenen umfassen. In kühleren Regionen halten Maulwürfe sich im Winter oft in größerer Tiefe auf als während der wärmeren Monate. In den Gangsystemen befinden sich mehrere Nestkammern, die mit Pflanzenmaterial gepolstert und häufig tiefer im Erdreich positioniert sind. Teilweise werden auch sehr große, „burg“ähnliche Hügel aufgehäuft. Bekannt ist dies vom Europäischen Maulwurf, aber auch vom Kaukasischen Maulwurf und vom Townsend-Maulwurf.[1]
Die meisten Arten dürften im Bedarfsfall schwimmen können. Dennoch haben sich mit den Desmanen und dem Sternmull teils wasserlebende Formen herausgebildet. Die Tiere verbringen einen Großteil ihrer aktiven Zeit im Wasser und suchen auch einen Teil ihrer Nahrung am Grund von Gewässern. Der Körper der Tiere ist stromlinienförmig gestaltet, die vorderen Gliedmaßen sind nicht so charakteristisch vorverlagert wie bei den grabenden Maulwürfen. Der Sternmull paddelt mit allen vier Gliedmaßen im Wasser. Bei den Desmanen sorgen ausschließlich die Hinterbeine für den Vortrieb, die abweichend von den grabenden Maulwürfen daher auch deutlich kräftiger entwickelt sind. An Land nutzen die wasserlebenden Maulwürfe teils eigens gegrabene Gangsysteme, mitunter auch Felsspalten als Rückzugsorte. Die Eingänge liegen ausschließlich unterhalb der Wasserlinie, die Ruhekammern jedoch oberhalb.[1]
Die Spitzmausmaulwürfe sind vermutlich reine Bewohner der Erdoberfläche und suchen wohl dort auch ihre Nahrung. Ihr Körperbau entspricht dadurch weitgehend dem der Spitzmäuse mit unter dem Körper liegenden Gliedmaßen. Der Bau der Hände und Füße erlaubt einen Zehengang. Über ihre Lebensweise ist allerdings so gut wie nichts bekannt. Ähnlich verhält es sich auch mit den verschiedenen Spitzmullen. Diese können ebenfalls vergleichbar den Spitzmausmaulwürfen vierfüßig laufen, legen jedoch auch teils einfache Gänge und Tunnel an, die in der Regel nur flach in den Untergrund reichen oder Laubfallschichten queren. Die Gänge dienen weitgehend der Nahrungssuche, können zudem auch einzelne Nestkammern enthalten.[1]
Spezielle Anpassungen und Sinneswahrnehmungen
Das Leben im Untergrund bei den grabenden Arten erforderte spezielle Modifikationen. Sowohl beim Ostamerikanischen Maulwurf als auch beim Europäischen Maulwurf finden sich als Anpassung an die sauerstoffarme Luft in den Gangsystemen Veränderungen in den biochemischen Eigenschaften des Hämoglobins. Infolgedessen ist bei beiden Arten die Aufnahmefähigkeit von CO2 erhöht, wodurch wiederum mehr Sauerstoff zur Verfügung gestellt werden kann. So besitzt das Hämoglobin des Europäischen Maulwurfs eine im Vergleich zu ausschließlich oberirdisch lebenden Kleinsäugern höhere Sauerstoffaffinität (nativer P50-Wert = 2,9 kPa gegenüber 6,5 kPa bei Mäusen), während die Sauerstoffaffinität von CO2-beladenem Carbaminohämoglobin relativ hoch ist.[30][31] Die Konzentration von 2,3-BPG in den Erythrozyten des Europäischen Maulwurfs liegt unter Normalbedingungen bei rund 5,3 mmol/l und ist damit gegenüber anderen Säugetieren nicht auffällig hoch.[31] Außerdem variiert sie zwischen verschiedenen Maulwurf-Arten. Die Bindungsaffinität des Hämoglobins gegenüber 2,3-BPG ist hingegen bei einigen Arten deutlich herabgesetzt und wird daher als weitere Ursache für die hohe Sauerstoffaffinität des Hämoglobins, sowie dessen CO2-Beladungskapazität diskutiert.[32][1]
Bei einigen Arten der Eurasischen Maulwürfe, der Ostasiatischen Maulwürfe, der Desmane und zusätzlich beim Sternmull, beim Haarschwanzmaulwurf und beim Amerikanischen Spitzmull treten die Weibchen als Hermaphroditen auf, was einmalig unter Säugetieren ist.[33] Sie stellen damit phänotypisch Weibchen dar, besitzen jedoch Zwitterdrüsen. Diese produzieren zwar keine Spermien, können aber, durch genetische Schalter gesteuert, sowohl weibliche als auch männliche Geschlechtshormone ausstoßen. Diese Tiere sind uneingeschränkt reproduktionsfähig, teilweise kommt es aber zum Geschlechtertausch, wodurch solche Individuen dann in der Regel unfruchtbar sind und zumeist kleinere Hoden als typische Männchen haben.[34][35][36] Bei genetischen Untersuchen wurden mehrere Faktoren für das Phänomen ausfindig gemacht, sie sind bisher ausschließlich bei Maulwürfen nachgewiesen. Dazu gehört einerseits eine Verdreifachung des Gens CYP17A1, wodurch zwei zusätzliche Kopien des entsprechenden Abschnittes vorhanden sind. Das Gen ist für ein Enzym verantwortlich, welches die Bildung von Androgenen, also männliche Sexualhormonen, kontrolliert. Außerdem fehlt in den Hodenabschnitten der Zwitterdrüsen das Gen CYP19A1, es kommt ausschließlich im Eierstockabschnitt vor. Dieses wiederum steuert die Synthese von Aromatase, ein Enzym, das Androgene in Östrogene umwandelt. Durch die genannten Veränderungen können größere Mengen an männlichen Sexualhormonen ausgeschüttet werden, das Vorkommen von CYP19A1 im Eierstockabschnitt der Zwitterdrüse hält wohl die Fortpflanzungsfähigkeit der Weibchen aufrecht. Andererseits liegt am Gen FGF9, das die Hodenbildung beeinflusst und in Verbindung mit anderen Genloci verschiedene Signaltransduktionswege zur Bildung der Eierstöcke hemmt, eine Drehung (Inversion) vor. Durch diese Neuarrangierung wird in einem frühen Stadium der Gonadenentwicklung die Meiose im Eierstockabschnitt der Zwitterdrüse verhindert und die stärkere Ausbildung des Hodenabschnittes befördert. Die „Vermännlichung“ der weiblichen Individuen bringt wahrscheinlich bei den Arten Vorteile, die eine unterirdische Lebensweise verfolgen. Da das Graben der Gänge sehr kraft- und energieaufwendig ist, kann durch die verstärkte Ausschüttung an männlichen Sexualhormonen die Muskelmasse gesteigert werden. Außerdem befördert es die Aggressivität, was wiederum bei der häufig einzelgängerischen Lebensweise behilflich ist.[33][1]

Aufgrund der Rückbildung der Augen ist der Sehsinn stark reduziert und beschränkt sich zumeist auf eine Unterscheidung von hell und dunkel. Dagegen sind andere Sinnesorgane teils außerordentlich entwickelt. Hierzu gehört der Tastsinn, der in der gesamten Sinneswahrnehmung eine wichtige Rolle spielt. Ein Teil wird dabei durch die Eimerschen Organe der Nasenregion übernommen, zusätzlich sind über die Schnauzen und an andere Körperpartien verteilt zahlreiche Vibrissen ausgebildet. Weite Bereiche der Neocortex der Großhirnrinde dienen der Erfassung und Verarbeitung taktiler Reize. Als Bote fungiert der Nervus trigeminus, welcher komplex aufgebaut ist. Beim Ostamerikanischen Maulwurf weist dieser bis zu 30.000 Afferenten auf, also Abzweigungen für den Signaltransport, beim Sternmull sind es bis 84.000. Der extrem hohe Wert bei letzterer Art ist auf die fingerförmigen Hautanhängsel der Nase mit dem komplexen Eimerschen Organsystem zurückzuführen.[37][38][39] Das dadurch entstehende hochsensible Tastorgan ermöglicht es dem Sternmull, hocheffizient seine zumeist kleinen Beutetiere zu erkennen, zu orten und einzufangen. Die sich daraus ergebenden Reaktionszeiten gehören zu den schnellsten im Tierreich.[19] Daneben nimmt auch der Geruchssinn eine wichtige Funktion ein. Für die grabenden Maulwürfe ist er von Bedeutung, um sich in den dunklen Gängen und Tunneln zu orientieren sowie die Beute zu orten. Beim Ostamerikanische Maulwurf konnte mit Hilfe von Experimenten festgestellt werden, dass dieser mit beiden Nasenlöchern getrennt Gerüche wahrzunehmen vermag. Der als Stereoriechen bekannte Effekt erhöht die Genauigkeit von Lokalisierungen.[40][41][39] Die semi-aquatisch lebenden Desemane und der Sternmull haben wiederum Techniken zur Aufnahme von Duftspuren unter Wasser entwickelt. Dies erfolgt über das Ausstoßen von Luftblasen, an denen die Geruchspartikel anhaften und welche dann über das Einatmen der Blasen aufgenommen werden.[42][43][1]
Sozialverhalten und Aktivitätszeiten

Die Sozialstruktur der Maulwürfe ist nur bei den wenigsten Arten untersucht. Wie die meisten Insektenfresser sind die Tiere in der Regel territoriale Einzelgänger und reagieren aggressiv auf Artgenossen. Ein gemeinsames Auftreten männlicher und weiblicher Individuen findet nur während der Fortpflanzungsphase statt. Die einzelnen Territorien können sich zwischen den Geschlechtern überschneiden, nicht aber bei Geschlechtsgenossen. Grenzmarkierungen erfolgen mit Sekreten aus den Analdrüsen. Nur wenige Arten teilen sich Gangsysteme mit Artgenossen, die sie nur in ihrer Nähe dulden, wenn reichlich Nahrung vorhanden ist. Hierzu zählen beispielsweise die Desmane und der Sternmull, eventuell auch der Amerikanische Spitzmull. In der Regel neigen Maulwürfe nicht zu ausgedehnten Wanderungen, sondern verbleiben innerhalb ihres etablierten Territoriums. Über die Kommunikation liegen nur wenige Informationen vor. Generell gelten Maulwürfe als nicht sehr lautgeberisch. Bekannt sind einige Quieklaute, die von Jungtieren oder bei Irritationen ausgestoßen werden.[1]
Die meisten unterirdisch lebenden Maulwürfe haben wie andere grabende Tiere keinen ausgeprägten Tag-Nacht-Rhythmus. So ist etwa beim Europäischen Maulwurf und beim Ostamerikanischen Maulwurf die Aktivitätszeit in mehrere Wach- und Schlafphasen aufgeteilt, wobei die Wachphasen jeweils bei einer Dauer von etwa 4 bis 5 Stunden liegen. Ähnliches konnte beim Sternmull und bei den Japanischen Spitzmullen beobachtet werden. Die weitgehend oberirdisch lebenden Maulwürfe können sowohl tag- als auch nachtaktiv sein, wenngleich öfter die Nachtaktivität überwiegt als dies bei den Desmanen der Fall ist. Die Arten der kühleren Regionen halten keinen Winterschlaf, in den Wintermonaten ziehen sich die Tiere in tiefere Erdschichten zurück.[1]
Ernährung und Energiehaushalt

Die Maulwürfe sind fleischfressend und ernähren sich generalistisch von einer weiten Bandbreite an Beutetieren. Der Hauptbestandteil der Nahrung setzt sich vornehmlich aus Wirbellosen zusammen, in seltenen Fällen kommen auch Wirbeltiere hinzu. Genauere Untersuchungen liegen aber nur von wenigen Arten vor. Die grabenden Maulwürfe suchen ihre Nahrung in oberflächennahen Gängen und Tunneln. Sie setzt sich überwiegend aus Regenwürmern zusammen und wird zusätzlich durch Insekten und deren Larven sowie Hundert- und Tausendfüßer oder Schnecken bereichert. In geringem Ausmaß kommen Pflanzen (vorwiegend deren unterirdische Teile) hinzu. Bei den bodenbewohnenden Arten erfolgt die Nahrungssuche oberflächig, teilweise in Laubfallschichten. Allerdings können wie bei den verschiedenen Spitzmullen hierbei auch flach im Erdboden verlaufende Gänge angelegt werden. Das Nahrungsspektrum ähnelt dem der grabenden Vertreter. Die Desmane und der Sternmull sind auf Gewässer angewiesen, wo sie schwimmend und tauchend auf Beutezug gehen. Hierbei bilden Insekten wie Eintags- und Köcherfliegen, aber auch Flohkrebse die primäre Bezugsquelle.[1]
Allgemein verdauen Maulwürfe ihrer Nahrung relativ schnell, beim Amerikanischen Spitzmull dauert der gesamte Vorgang von der Nahrungsaufnahme bis zum Ausscheiden insgesamt 45 Minuten. Zudem ist auch der Energiebedarf relativ hoch, wodurch die Tiere täglich sehr viel Nahrung zu sich nehmen müssen. Dies trifft insbesondere auf die Arten zu, welche im Untergrund graben. Beim Zurücklegen einer Entfernung von 1 m auf dem Erdboden verbraucht ein 120 g schweres Tier rund 0,002 kcal, für die gleiche Entfernung grabend im Untergrund steigt der Bedarf auf fast 1,2 kcal. Die täglich aufgenommene Nahrungsmenge für den Europäischen Maulwurf und den Ostamerikanischen Maulwurf entspricht so nahezu der des eigenen Körpergewichts. Gleiches trifft möglicherweise auf die Spitzmulle zu, bei den Desmanen liegt er etwa bei der Hälfte des eigenen Körpergewichts. Über körpereigene Energiespeicher verfügen nur die wenigsten Arten, so etwa der Sternmull und der Japanische Spitzmull, die beide Fett in ihrem Schwanz ablagern können. Aus diesem Grund und wegen des hohen Nahrungsbedarfs überleben die Tiere längere Nahrungspausen über 12 bis 24 Stunden nicht. Einige Arten umgehen dies, indem sie Nahrungsvorräte anhäufen, so etwa der Europäische Maulwurf.[1]
Die Körpertemperatur der Maulwürfe reicht von 34,5 bis 38,4 °C. Die Stoffwechselrate ist vergleichbar der anderer Kleinsäuger, sie tendiert aber bei den grabenden Vertretern hin zu einem niedrigeren Bereich. Dies begründet sich in den hohen Energiekosten des Grabens und schützt die Tiere zudem vor Überhitzung. Aufgrund des dichten Fells können die Maulwürfe nur schlecht überschüssige Wärme ableiten. Bei warmer Umgebung erhöht sich daher die Blutzirkulation vor allem an der Schnauze, am Schwanz und an den Füßen. Grabende Tiere ziehen sich dann auch in größere Tiefen zurück, semi-aquatische Formen nutzen zur Kühlung das Wasser.[1]
Fortpflanzung und Lebenserwartung
Über die Fortpflanzung ist wenig bekannt, die meisten Maulwürfe vermehren sich nicht in menschlicher Gefangenschaft. Die Paarungsphase ist jahreszeitlich begrenzt. Bei paarungsbereiten Männchen schwellen die Hoden stark an. Die Tragzeit beträgt zwischen 26 und 45 Tage. Ein Großteil des Nachwuchses kommt in den Frühlingsmonaten zur Welt. Muttertiere tragen bis zu zehn Embryonen aus, durchschnittlich sind es allerdings 4,5. Teilweise besteht eine hohe vorgeburtliche Sterblichkeit von fast einem Viertel. Würfe umfassen demnach zwischen zwei und sechs Neugeborene. Nur bei wenigen Arten ist ein zweiter Fortpflanzungszyklus im Jahr vermerkt, dazu zählen etwa der Japanische Maulwurf und der Kleine Japanische Maulwurf. Neugeborene sind Nesthocker, sie haben nackte Haut und geschlossene Augen und Ohren. Zumeist übernimmt nur das Muttertier die Aufzucht, eine Ausnahme bilden die Desmane. Die Saugphase währt rund einen Monat. Für die Jungen besteht die größte Sterblichkeitsrate im ersten Lebensjahr, wenn sie das mütterliche Nest verlassen. Die Geschlechtsreife setzt mit sechs bis zwölf Monaten ein, so dass der Nachwuchs sich in der Regel erst im Jahr nach der Geburt verpaart. Die Lebenserwartung ist relativ gering und dürfte bei den wenigsten Tieren drei bis sechs Jahre übersteigen.[1]
Ökologische Bedeutung
Durch ihre Grabtätigkeit haben die unterirdisch lebenden Maulwürfe als Vertreter des wühlenden (fodenten) Edaphons einen gewissen Anteil an der Bodenbildung (Pedogenese). Sie tragen mitunter zur Durchmischung der humosen oberen Bodenabschnittes (Ah-Horizont) im Rahmen der Bioturbation bei. Außerdem verfrachten sie humusfreies Unterbodenmaterial nach oben. Durch ihren lockernden Einfluss auf das Bodengefüge begünstigen sie ferner die Durchlüftung beziehungsweise bedingt auch die Drainage. Die Grabetätigkeit befördert zudem den Pflanzenwuchs und verhilft durch das Öffnen der Vegetationsdecke konkurrenzschwächere Pflanzen zur Entfaltung. Dies hat wiederum Auswirkungen auf die unmittelbare Tierwelt.[44][45] Des Weiteren profitieren zahlreiche andere Tiere von den durch die Maulwürfe angelegten Gänge. Der semi-aquatischen Maulwürfe können als Anzeiger für die Wasserqualität und die Intaktheit des Gewässernetzes gewertet werden.[1]
Systematik
Äußere Systematik
Innere Systematik der Insektenfresser nach Brace et al. 2016[46]
|
Die Maulwürfe sind eine Familie aus der Ordnung der Insektenfresser (Eulipotyphla). Diese Ordnung hat eine taxonomisch wechselhafte Geschichte, bei der immer wieder verschiedene Gruppen ein- oder ausgegliedert wurden. Aus heutiger Sicht gehören neben den Maulwürfen noch die Spitzmäuse (Soricidae), Igel (Erinaceidae) und die Schlitzrüssler (Solenodontidae) dazu, ebenso wie die in jüngerer Zeit ausgestorbenen Karibischen Spitzmäuse (Nesophontidae). Molekulargenetische Untersuchungen sehen die Maulwürfe hierbei in einem engeren Verhältnis zu den Igeln und Spitzmäusen, wobei erstere die Außengruppe von letzteren beiden bilden.[47][46][48] Die Insektenfresser stellen eine alte Gruppe der Höheren Säugetiere (Eutheria) dar. Sie lassen sich genetisch bis in die Oberkreide vor rund 72 Millionen Jahren zurückverfolgen. Die Maulwürfe trennten sich von ihren verwandten Linien bereits vor rund 68 Millionen Jahren ab, noch kurz bevor der Kreide-Paläogen-Grenze.[48] Teilweise werden auch jüngere Daten angegeben,[49] doch beruhen diese vermutlich auf ungenauen Kalibrierungen.[48]
Innerhalb der Insektenfresser formen die Maulwürfe gemeinsam mit einigen ausgestorbenen Gruppen einen engeren Verwandtschaftskreis, der als Überfamilie der Talpoidea bezeichnet wird. Diese schließen als eine weitere Einheit die Proscalopidae ein, die ursprünglich als Unterfamilie der Maulwürfe geführt wurde, seit Beginn der 1980er Jahre jedoch als eigenständige Familie gilt. Deren Mitglieder waren ebenfalls stark an das unterirdische Leben angepasst, zeigten aber Abweichungen im Bau der Gliedmaßen und Schädelgestaltung, wobei letzterer im Unterschied zu den Maulwürfen ebenfalls zum Graben eingesetzt wurde. Die Tiere lebten vom Paläozän bis zum Miozän in Nordamerika.[50][51] Etwa zur gleichen Zeit traten in Europa die Dimylidae auf, deren markantes Kennzeichen neben einem generell niederkronigeren Gebiss eine Reduktion des letzten oberen Mahlzahns ist.[52]
Innere Systematik
Innere Systematik der Maulwürfe nach He et al. 2016[53]
|
Die Maulwürfe werden heute in zwei Unterfamilien untergliedert. Die eine umfasst die weitgehend auf Ostasien beschränkten, oberirdisch lebenden Spitzmausmaulwürfe (Uropsilinae). Aufgrund ihres generalisierten Körperbaus, der stark an die Spitzmäuse erinnert, können diese als relativ ursprünglich aufgefasst werden. Die zweite Unterfamilie besteht aus einer Gruppe spezialisierterer Formen, welche einer grabenden Lebensweise nachgehen, sich teilweise oberirdisch aufhalten oder aber semi-aquatisch auftreten. Aus forschungsgeschichtlichen Gründen trägt sie die Bezeichnung Altweltmaulwürfe (Talpinae), kommt aber sowohl in Eurasien als auch in Nordamerika vor. Den Altweltmaulwürfen werden verschiedene Triben zugeordnet. Die strikt unterirdisch lebenden Maulwürfe teilen sich hierbei einerseits auf die Eigentlichen Maulwürfe (Talpini), andererseits auf die Neuweltmaulwürfe (Scalopini) auf. Beide Gruppen sind innerhalb der Altweltmaulwürfe eher entfernter verwandt. Erstere schließt mit den Eurasischen Maulwürfen (Talpa) und somit mit dem in Mitteleuropa heimischen Europäischen Maulwurf (Talpa europaea) das bekannteste Mitglied ein, umfasst aber auch mehrere weitere Gattungen mit ähnlichem Körperbau aus dem ost- und südostasiatischen Raum. Die Neuweltmaulwürfe (Scalopini) sind abweichend von ihrem Trivialnamen nicht nur in Nordamerika, sondern auch in Ostasien heimisch. Als näher mit den Eigentlichen Maulwürfen verbunden gelten die an das Wasserleben angepassten Desmane (Desmanini) und der hoch spezialisierte Sternmull (Condylurini). Eine weitere, engere Verwandtschaftsgruppe bilden die verschiedenen „Spitzmulle“. Hierzu zählen der Langschwanzmaulwurf (Scaptonychini), die Japanischen Spitzmulle (Urotrichini) und der Amerikanische Spitzmull (Neurotrichini), die jeweils einer eigenen Tribus angehören. Einige Systematiken vereinen alle drei Gruppen auch innerhalb der Urotrichini, wobei der Langschwanzmaulwurf dann als eigenständiger Untertribus der Scaptonychina von den Japanischen Spitzmullen und dem Amerikanischen Spitzmull separiert wird. Letztere beiden finden sich in dieser Version innerhalb der Urotrichina.[53][54][1]
Diese innere Gliederung der Maulwürfe basiert auf molekulargenetischen Analysen. Bei diesen erwiesen sich alle Maulwürfe abseits der Spitzmausmaulwürfe als monophyletische Einheit. Zuvor wurden mehrere eigenständige Unterfamilien ausgewiesen. Die Neuweltmaulwürfe standen demgemäß als Scalopinae den Talpinae beziehungsweise den Altweltmaulwürfen gegenüber. Aufgrund der in beiden Gruppen ausgeprägten grabenden Lebensweise stufte man sie als eng miteinander verwandt ein. Ebenso wurden die Desmane als separate Unterfamilie geführt, während der Sternmull den Neuweltmaulwürfen näher stand. Über die Beziehungen der „Spitzmulle“ zueinander herrschten indes unterschiedliche Auffassungen vor.[55][56][53][54] Die genetischen Untersuchungen, die verstärkt seit den 2010er Jahren an den Maulwürfen durchgeführt wurden, gaben somit Klärung über die innere Gliederung der Familie. Sie zeigten aber auch auf, dass die anhand morphologischer und verhaltensbiologischer Kriterien vermuteten Verwandtschaftsbeziehungen nicht zutreffen. Demnach entwickelte sich die grabende Lebensweise der Maulwürfe, die besonders extrem bei den Eigentlichen Maulwürfen und den Neuweltmaulwürfen ausgeprägt ist, mehrfach innerhalb der Familie. Gleiches wird für die semi-aquatische Lebensweise der Desmane und des Sternmulls angenommen, die jeweils auf unterschiedlichen anatomischen Anpassungen beruhen.[57][58][53][29]
Neben den heute noch bestehenden Gruppen sind zusätzlich einige ausgestorbene Gattungsgemeinschaften bekannt. Eine davon bildet die Unterfamilie der Suleimaninae, die im Unteren Miozän in Westasien und Südosteuropa auftrat. Deren vergleichsweise großen Vertreter zeichneten sich unter anderem durch eine Reduktion der hintersten Zähne aus.[59] Eine weitere erloschene Linie wird durch die Unterfamilie der Gaillardinae repräsentiert. Bei diesen handelt es sich um desmanartige Maulwürfe, die wahrscheinlich ebenfalls an eine semi-aquatische Lebensweise angepasst waren. Die Tiere lebten im Übergang vom Miozän zum Pliozän in Nordamerika.[60]
Überblick über die Unterfamilien und Gattungen der Maulwürfe
Die rezenten Maulwürfe verteilen sich auf gut 20 Gattungen mit über 60 Arten, wobei sich innerhalb einiger Gruppen auch kryptische Formen verbergen können. Hinzu kommen zahlreiche ausgestorbene Vertreter. Die Übersicht zur inneren Systematik bis zur Gattungsebene beruht auf den Arbeiten von Rainer Hutterer 2005, Kai He und Kollegen 2016 sowie Boris Kryštufek und Masaharu Motokawa 2018,[61][53][1] berücksichtigt aber auch einzelne neuere Entwicklungen:[62][28]
- Familie: Talpidae Fischer, 1817
- Achlyoscapter Hutchison, 1968
- Eotalpa Sigé, Crochet & Insole, 1977
- Mongolopala Ziegler, Dahlmann & Storch, 2007
- Nuragha de Brujin & Rümke, 1974
- Oreotalpa Lloyd & Eberle, 2008
- Tegulariscaptor Sansalone, Kotsakis, Schwermann, Van den Hoek Ostende & Piras, 2017
- Unterfamilie: Gaillardinae Hutchison, 1968
- Gaillardia Matthew, 1932
- Unterfamilie: Suleimaninae Van den Hoek Ostende, 2001
- Suleimania Van den Hoek Ostende, 2001
- Unterfamilie: Talpinae Fischer, 1817 (Altweltmaulwürfe)
- Tribus: Condylurini Gill, 1875
- Condylura Illiger, 1811 (Sternmull, einschließlich 1 rezenten Art)
- Tribus: Desmanini Thomas, 1912 (Desmane)
- Archaeodesmana Topachevski & Pashkov, 1983
- Asthenoscapter Hutchison, 1974
- Desmana Güldenstädt, 1777 (Russischer Desman, einschließlich 1 rezenten Art)
- Galemys Kaup, 1829 (Pyrenäen-Desman, einschließlich 1 rezenten Art)
- Gerhardstorchia Dahlmann & Doğan, 2011
- Lemoynea Bown, 1980
- Magnatalpa Oberg & Samuels, 2022
- Mygalea Schreuder, 1940
- Mygalinia Schreuder, 1940
- Mygatalpa Schreuder, 1940
- Tribus: Neurotrichini Hutterer, 2005
- Neurotrichus Günther, 1880 (Amerikanischer Spitzmull, einschließlich 1 rezenten Art)
- Quyania Storch & Qiu, 1983
- Rzebikia Sansalone, Kotsakis & Piras, 2016
- Tribus: Scalopini Gill, 1875 (Neuweltmaulwürfe)
- Alpiscaptulus Jiang & Chen, 2021 (einschließlich 1 rezenten Art)
- Domninoides Green, 1956
- Hugueneya van den Hoek Ostende, 1989
- Leptoscaptor Ziegler, 2003
- Mioscalops Ostrander, Mebrate & Wilson, 1986
- Parascalops True, 1894 (Haarschwanzmaulwurf, einschließlich 1 rezenten Art)
- Proscapanus Gaillard, 1899
- Scalopus É. Geoffroy Saint-Hilaire, 1803 (Ostamerikanischer Maulwurf, einschließlich 1 rezenten Art)
- Scapanoscapter Hutchison, 1968
- Scapanulus Thomas, 1912 (Gansu-Maulwurf, einschließlich 1 rezenten Art)
- Scapanus Pomel, 1848 (Westamerikanische Maulwürfe, einschließlich 5 rezente Arten)
- Yanshuella Storch & Qiu, 1983
- Yunoscaptor Storch & Qiu, 1990
- Tribus: Scaptonychini Van Valen, 1967
- Mongoloscapter Lopatin, 2002
- Scaptonyx Milne-Edwards, 1872 (Langschwanzmaulwurf, einschließlich 1 rezenten Art)
- Tribus: Talpini Fischer, 1817 (Eigentliche Maulwürfe)
- Euroscaptor Miller, 1940 (Südostasiatische Maulwürfe, einschließlich 10 rezente Arten)
- Geotrypus Pomel, 1848
- Mogera Pomel, 1848 (Ostasiatische Maulwürfe, einschließlich 9 rezente Arten)
- Oreoscaptor Kawada, 2016 (Japanischer Bergmaulwurf, einschließlich 1 rezenten Art)
- Parascaptor Gill, 1875 (Weißschwanzmaulwurf, einschließlich 1 rezenten Art)
- Scaptochirus Milne-Edwards, 1867 (Kurzgesichtmaulwurf, einschließlich 1 rezenten Art)
- Skoczenia Rzebik-Kowalska, 2014
- Talpa Linnaeus, 1758 (Eurasische Maulwürfe, einschließlich 16 rezenten Arten)
- Tribus: Urotrichini Dobson, 1883 (Japanische Spitzmulle)
- Desmanodon Engesser, 1980
- Dymecodon True, 1886 (True-Spitzmull, einschließlich 1 rezenten Art)
- Myxomygale Filhol, 1890
- Paratalpa Lavocat, 1951
- Percymygale Hugueney & Maridet, 2017
- Pseudoparatalpa Lopatin, 1999
- Tenuibrachiatum Ziegler, 2003
- Urotrichus Temminck, 1841 (Japanischer Spitzmull, einschließlich 1 rezenten Art)
- Unterfamilie: Uropsilinae Dobson, 1883
- Desmanella Engesser, 1972
- Mystipterus Hall, 1930
- Theratiskos van den Hoek Ostende, 2001
- Uropsilus Milne-Edwards, 1871 (Spitzmausmaulwürfe, einschließlich 10 rezente Arten)
Stammesgeschichte
Ursprünge und Eozän
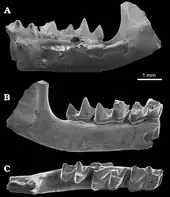
Der Ursprung der Maulwürfe ist nicht geklärt. Eine Zeitlang wurde eine Herkunft aus dem Eozän in Europa angenommen, das damals eine isolierte Insel formte. Von dort sind auch die ältesten Fossilien bekannt. Sie werden der Gattung Eotalpa zugesprochen, die mit mehreren Zahn- und Gebissfunden, aber auch Teilen des Bewegungsapparates von der Isle of Wight sowie dem Hordle Cliff in der englischen Grafschaft Hampshire und aus Creechbarrow in der Grafschaft Dorset überliefert ist. Die Funde entstammen der Headon-Hill-Formation und der Bembridge-Limestone-Formation, welche beide im Oberen Eozän entstanden.[63][64][65] Eotalpa als der bisherige älteste Vertreter der Maulwürfe zeigt einzelne Besonderheiten, die die Form nicht an die Basis der Entwicklung der Familie stellen, so etwa in einzelnen Zahnmerkmalen. Auch die aufgefundenen Fußknochen lassen bestimmte Spezialisierungen erkennen, beispielsweise sind die Metapodien und Phalangen im Vergleich zu den Spitzmausmaulwürfen als ursprünglichste rezente Gruppe kürzer und flacher. Sie verweisen auf eine bereits einfache Anpassung an grabende Eigenschaften, was auch durch die Bildung einer schwachen Leiste am oberen Gelenkende der Elle gestützt wird, welche bei den Spitzmausmaulwürfen fehlt. Dahingegen spricht die Struktur des Sprung- und Fersenbeins für eher oberirdische Aktivitäten. Damit zeigten die Maulwürfe bereits im Eozän gewisse Anzeichen einer grabenden Lebensweise, was einer teils vertretenen Meinung einer semi-aquatischen Herkunft widerspricht. Ein markanter Unterschied zu den heutigen Maulwürfen besteht bei Eotalpa darin, dass das Schien- und Wadenbein noch nicht miteinander verwachsen waren. Die Mischung aus ursprünglichen und entwickelten Merkmalen verschieben den Ursprung der Maulwürfe bis möglicherweise in das Paläozän und machen eine Herkunft aus dem heutigen Asien wahrscheinlich.[65]
Noch in das Obere Eozän fällt der Erstnachweis der Maulwürfe in Nordamerika. Es handelt sich hierbei um einen partiellen Unterkiefer mit hinterer Bezahnung. Er wurde in der Florissant-Formation im Florissant Fossil Beds National Monument im US-Bundesstaat Colorado entdeckt und der Gattung Oreotalpa zugewiesen. Der Gattungsname referenziert den heutigen gebirgigen Charakter der Fundregion. Das Alter der Ablagerungen mit dem Unterkiefer wird auf rund 34 Millionen Jahren datiert.[66]
Oligozän
Eotalpa ist im Unteren Oligozän weiterhin präsent, isolierte Zähne wurden in der Borgloon-Formation bei Boutersem in Belgien gefunden.[67] Im gleichen Zeitraum lassen sich Maulwürfe in Asien belegen. Die Hsanda-Gol-Formation im Tal der Gobiseen in der zentralen Mongolei erbrachte Ober- und Unterkieferbruchstücke von Mongolopala. Die frühe asiatische Form ist in Größe und Zahncharakteristika relativ ähnlich zum nordamerikanischen Oreotalpa. Die Funde zeigen auf, dass die Maulwürfe bereits im Übergang vom Eozän zum Oligozän kontinentalweite Wanderungen unternommen und somit ihre heutigen Verbreitungsgebiete erreicht hatten.[68][66]
Während sich die frühesten Maulwürfe keiner bestimmten Linie zuweisen lassen, repräsentiert Percymygale einen Angehörigen der Japanischen Spitzmulle. Die Gattung trat im Unteren Oligozän in Erscheinung und ist aus mehreren Karstspalten im südwestlichen Deutschland dokumentiert, so etwa in Möhren nordöstlich von Ulm, wobei einige Zahn- und Kieferfunde aus der Spalte Möhren 20 für eine Anwesenheit bereits im Obersten Eozän sprechen. Die Form war geographisch recht weit verbreitet und ist während des gesamten Oligozäns im mittleren und westlichen Europa nachweisbar. Ihr zur Seite steht das nahe verwandte Myxomygale, das zeitlich etwas später in Erscheinung tritt, aber ein räumlich ähnliches Vorkommen aufweist. Beide Formen zeichnen sich durch einen schlanken Oberarmknochen aus. Bei Percymygale war das Rostrum aber deutlich länger ausgebildet, was häufig mit einer nur geringen Anpassung an bodengrabende Aktivitäten verbunden wird.[69][70] Allgemein bilden die zahlreichen Karstspalten Südwestdeutschlands eine reichhaltige Quelle zur Entwicklung der frühen Maulwürfe. Jene von Ehrenstein ebenfalls bei Ulm erbrachten unter anderem Gebiss- und Beinreste von Tegulariscaptor, ein Vertreter, der ebenfalls in die nähere Verwandtschaft zu den Japanischen Spitzmullen eingeordnet wird. Der wiederum recht schlanke Oberarmknochen weist aber schon gewisse Eigenschaften auf, etwa das breite Ellenbogengelenk, die eine teils grabende Lebensweise andeuten.[71] Die Karstspalten von Ehrenstein lieferten zusätzlich auch Geotrypus, mit dem erstmals die Eigentlichen Maulwürfe im ausgehenden Unteren und im Oberen Oligozän dokumentiert sind. Die Tiere waren offensichtlich bereits vollständig an das Graben im Untergrund angepasst. Hinweise hierfür geben mehrere Teilskelette, von denen eines aus Enspel in Rheinland-Pfalz stammt. Das dort aufgefundene Individuum verfügte über den charakteristischen „Vordaumen“ zur Verbreiterung der Handfläche, zudem waren die Vorderbeine relativ massiv ausgebildet.[72][73] Im Oberen Oligozän sind des Weiteren die Spitzmausmaulwürfe fassbar, so mit der Gattung Demanella aus einer Karstspalte bei Eggingen nahe Ulm. Gleiches gilt für die Neuweltmaulwürfe mit Hugueneya aus derselben Lokalität. Beide Gattungen sind hier aber weitgehend nur über Zähne bekannt.[74][75] Abseits des südwestdeutschen Gebietes haben die Desmane mit Mygatalpa ihren frühesten Nachweis, hier liegen einzelne Unterkieferfragmente aus dem Oberen Oligozän des nördlichen Frankreichs vor.[76]
Außerhalb des europäischen Raumes sind im Oberen Oligozän weitere Vertreter der Maulwürfe fassbar. Im östlichen Asien ist aus oberen Abschnitten des Hsanda-Gol-Formation Mongoloscapter über einzelne Unterkieferfragmente dokumentiert, das wohl in einer näheren Beziehung zum Langschwanzmaulwurf steht.[77][78] In Nordamerika wiederum stellt ein Unterkieferbruchstück aus der Brule-Formation im südwestlichen North Dakota den ersten Hinweis auf die Spitzmausmaulwürfe dar. Der Fund wird der dortigen Gattung Mystipterus zugerechnet.[79]
Miozän
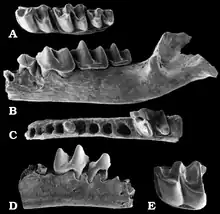
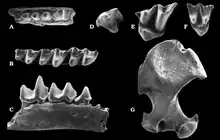
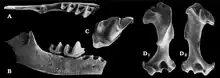
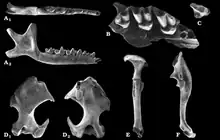
Im Unteren Miozän erfolgte eine weitere Diversifizierung der Maulwürfe. Neben den heute noch bestehenden Linien brachte die Familie zusätzlich noch die Suleimaninae hervor. Deren Charakterform Suleimania ist erstmals in Vorderasien nachweisbar. Abweichend von allen anderen Maulwürfen war der hinterste obere Molar zurückgebildet und der entsprechende untere verkleinert. Rein oberflächlich erinnern die Tiere dadurch an die Dimylidae. Zahlreiche Zahnfunde wurden an Fundstellen wie Harami und Gökler in Anatolien entdeckt,[59][80] einzelne Fossilien liegen auch von der Balkanhalbinsel vor.[81] Eine bisher rätselhafte Form kam mit Nuragha im Unteren Miozän auf dem heutigen Sardinien vor. Benannt wurde sie nach den konischen Höckern auf den unteren Mahlzähnen, die ein wenig an die als Nuraghen bezeichneten Festungstürme der sardischen Bronzezeit erinnern.[82] Desmanella bleibt eine dominante Form der Spitzmausmaulwürfe, die bis in das Obere Miozän Bestand hatte und weite Teile Europas und Vorderasiens bewohnte.[83] Im Vorderasien teilte sich Desmanella den Lebensraum mit Theratiskos, das wie sein Verwandter über recht schlanke Gliedmaßen verfügte. Belegt ist die Gattung bis nach Zentralasien.[59][84][85] Innerhalb der Neuweltmaulwürfe löst Proscapanus das ältere Hugueneya ab und avanciert zu einem wichtigen Vertreter. Nachweisbar ist er bis zum ausgehenden Mittleren Miozän in West- und Mitteleuropa. Funde liegen aus mehreren Karstspalten Südwestdeutschlands vor. Zu nennen wäre hier etwa Petersbuch bei Eichstätt.[86] Von großer Bedeutung erweist sich auch ein Teilskelett aus den untermiozänen Ablagerungen von Viehausen bei Regensburg. Dessen massiver Oberarmknochen ebenso wie der „Vordaumen“ der Hand zeigen die Grabbefähigung der Tiere an.[73] Petersbuch barg zudem noch Reste von Leptoscaptor, das hier lokal auftritt und mitunter eine große Fundhäufigkeit erreicht. Parallel zu Myxomygale ist im Unteren Miozän noch Paratalpa als Angehöriger der Japanischen Spitzmulle belegt. Die Gattung verschwindet aber alsbald wieder und wird durch Desmanodon abgelöst. Die Form findet sich in zahlreichen europäischen und westasiatischen Fundstellen wieder. Ihr im Vergleich zu Paratalpa etwas kräftigerer Oberarmknochen lässt gewisse Grabbefähigungen vermuten.[59][80] Dies trifft wiederum nicht auf das verwandte Tenuibrachiatum zu, das im Mittleren Miozän lokal in Petersbuch erscheint.[86] Eine ebenfalls regionale Variante der Japanischen Spitzmulle bildet Pseudoparatalpa, welches unter anderem aus der Aral-Formation in Kasachstan berichtet wurde.[84][85] Die Desmane werden im Unteren Miozän durch Mygalea vertreten. Auch hier erbrachten die Ablagerungen von Viehhausen eines der wenigen bekannten Teilskelette,[73] während aus den Karstspalten von Petersbuch Einzelzähne und Kieferfragmente vorliegen.[86] Im Mittleren Miozän treten noch Asthenoscaptor und Gerhardstorchia hinzu, letztere Gattung besteht noch bis zum Oberen Miozän.[87][88] Beide Formen sind aber eher selten. In das Obere Miozän fällt der Erstnachweis von Archaeodesmana, das im Folgenden fast europaweit dokumentiert werden konnte, einen Verbreitungsschwerpunkt aber auf der Iberischen Halbinsel findet.[89] Geotrypus als Repräsentant der Eigentlichen Maulwürfe ist nach dem Unteren Miozän weitgehend nicht mehr nachweisbar. Abgelöst wird die Form durch die frühesten Vertreter der Eurasischen Maulwürfe (Talpa). Diese werden häufig als Seitenzweig interpretiert, der im Oberen Miozän vorläufig wieder aus dem Fossilbericht verschwand.[90]
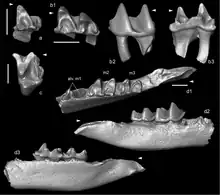
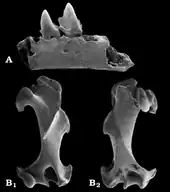
Nachdem bereits im Oligozän nahezu alle heutigen Linien der Maulwürfe etabliert sind, treten im östlichen Asien im Unteren Miozän erstmals die Vorfahren des Amerikanischen Spitzmulls (Neurotrichus) hervor. Es handelt sich hierbei um die Gattung Quyania, die mit wenigen isolierten Zähnen aus dem Gebiet des Aoerban in der zentralen Inneren Mongolei erscheint und in der Region noch bis zum Oberen Miozän verbleibt. Das Fundgebiet lieferte zudem Reste von Desmanella, Proscapanus und zusätzlich von Yanshuella.[91] Quyania und Yanshunella, letzterer repräsentiert die Neuweltmaulwürfe, sind mit zusammen mehreren hundert Forssilresten in Form von Zähnen, Unterkiefern und Beinresten aus Ertemte und Harr Obo im Süden der Autonomen Region Chinas dokumentiert. Beide Fundstellen gehören dem Oberen Miozän an.[92] Weitere gemeinsame Fundpunkte wurden am Baikalsee aufgeschlossen, wo zusätzlich noch Desmanodon auftrat.[93] Von Ostasien aus fand Quyania seinen Weg nach Europa. Hier sind obermiozäne Fossilfunde unter anderem aus einer Spaltenfüllung bei Petersbuch beschrieben worden.[86] Yanshuella hingegen besiedelte im gleichen Zeitraum Nordamerika, Unterkiefer und Zähne der Gattung kamen in Oregon zu Tage.[60][92] In Ostasien tritt mit Yunoscaptor im Oberen Miozän ein weiterer Angehöriger der Neuweltmaulwürfe hinzu. Fundpunkte dieser sehr kleinen Form liegen im nördlichen und südlichen Teil des heutigen Chinas, wobei für erstere Region, speziell dem Yushe-Becken, ein gemeinsames Vorkommen mit Yanshuella verzeichnet ist.[94][95]
Neben den eingewanderten Maulwurfsvertretern bildeten sich in Nordamerika im Miozän eigenständige Linien aus. Das bereits im Oligozän nachgewiesene Mystipterus aus der Gruppe der Spitzmausmaulwürfe hielt sich noch bis in das Obere Miozän. Sehr umfangreich fällt der Fossilbericht der Neuweltmaulwürfe aus. Eine frühe Form bildet Mioscalops, deren älteste Funde aus der Harrison-Formation in Nebraska noch in das Untere Miozän datieren, aber nur einzelne Unterkiefer beinhalten.[96] Weitere Belege sind aus Colorado bekannt. Ein sehr umfangreiches Fossilienmaterial mit Schädel- und Beinresten erbrachte das Quartz Basin in Oregon mit einer Altersstellung im Mittleren Miozän. Von hier wurde auch Domninoides berichtet. Aus relativer regionaler Nähe liegen Reste von Scapanoscapter vor. Alle drei Vertreter waren mit ihren massiven Beinknochen gut an die grabenden Lebensbedingungen angepasst. Die beiden ersteren verfügten über ein reduziertes Gebiss, letzterer besaß eine vollständige Bezahnung und stand dadurch eventuell den Westamerikanischen Maulwürfen (Scapanus) näher. Parallel zu Scapanoscapter sind die Westamerikanischen Maulwürfe im Mittleren Miozän ebenfalls fassbar.[97][60] Die Vorfahren des Ostamerikanischen Maulwurfs (Scalopus) zeigen sich erstmals im Oberen Miozän.[98] Im Gegensatz dazu sind die Vorfahren des Sternmulls (Condylura) weitgehend unbekannt. Einige Autoren ordnen Achlyoscapter in dessen nähere Verwandtschaft ein. Von ihm wurden mehrere Kieferfragmente im Quartz Basin von Oregon entdeckt, die wie die Funde der Neuweltmaulwürfe dort dem Mittleren Miozän angehören. Eine heute nicht mehr bestehende Linie der Maulwürfe repräsentiert Gaillardia, die Typusform der Gaillardinae. Reste in Form von Kieferfragmenten und postcranialen Skelettelementen liegen auf dem nördlichen Oregon, zusätzlich auch aus Nebraska vor. Ähnlich den Desmanen lebten die Tiere semi-aquatisch und bewegten sich im Wasser mit Hilfe der Beine und des Schwanzes fort. Es bestehen aber deutliche anatomische Unterschiede zu den Desmanen, etwa am Oberarmknochen und am Schlüsselbein, die in einzelnen Aspekten eher an die Spitzmausmaulwürfe erinnern. Auch der Unterkiefer weist markante Abweichungen zu den Desmanen auf.[60][99] Parallel zu den Gailardinae bestätigt Lemoynea mit einzelnen Zahn- und Kieferresten aus Nebraska auch die Anwesenheit der Desmane im Oberen Miozän in Nordamerika.[100][51]
Pliozän und Pleistozän
Das Pliozän und das Pleistozän zeichnen sich durch das Verschwinden zahlreicher älterer Formen der Maulwürfe und das Aufkommen der moderneren Linien aus. In Eurasien sind vor allem für das Pliozän noch einzelne stammesgeschichtlich ältere Vertreter nachgewiesen. Bedeutung erhalten hier mehrere Fundstellen im südlichen Polen wie Podlesice, Węże und Rębielice Królewskie. Es handelt sich hierbei um Quyania als Nahverwandter des Amerikanischen Spitzmulls, während etwa gleichzeitig noch das mit ihm eng verbundene Rzebikia hinzukommt. Für beide Formen sind zahlreiche Kieferreste und Gliedmaßenfragmente dokumentiert. Die südpolnischen Lokalitäten bargen darüber hinaus auch Reste des Haarschwanzmaulwurfs (Parascalops) aus der Gruppe der Neuweltmaulwürfe, der hier einen seinen frühesten Beleg aufweist. Gleiches gilt für die Vorfahren des heutigen Sternmulls, der abseits eines etwas fraglichen Fundes aus dem Mittleren Miozän Zentralasiens hier seinen ersten Auftritt hat, was für einen eurasischen Ursprung der Linie von Condylura spricht.[101][102][103][104] Parallel zu dieser wasserlebenden Form erreichen die Desmane mit Archaeodesmana im Pliozän eine weite Verbreitung im westlichen Eurasien. Die Form gilt als Ausgangspunkt der Entwicklung der heutigen Vertreter. Deren Gattungen Galemys und Desmana kommen ebenfalls im Pliozän erstmals auf. Abweichend von den heutigen Verhältnissen mit einem eher eingeschränkten Lebensraum waren sie aber vor allem im Pleistozän vielfach in Europa präsent.[89] Die Eigentlichen Maulwürfe werden kurzfristig im Pliozän durch Skoczenia vertreten, das weitgehend nur über Gebissreste aus dem südlichen Polen bekannt ist.[103] Talpa als Gattung der Eurasischen Maulwürfe hingegen hält nach seinem Verschwinden aus dem Fossilbericht im Oberen Miozän im Pliozän wieder Einzug in Europa und tritt mit mehreren, morphometrisch voneinander abweichenden Formen auf. Teilweise sind diese sympatrisch für einzelne Fundstellen dokumentiert, exemplarisch dafür ist eine der jüngsten Spaltenfüllungen aus Petersbuch erwähnt, die in das Mittelpleistozän datiert und rund 500 Reste mehrerer Angehöriger der Eurasischen Maulwürfe enthielt.[105][106] Andere Mitglieder der Eigentlichen Maulwürfe wie die artenreichen Ostasiatischen Maulwürfe (Mogera) oder aber der Japanische Bergmaulwurf (Oreoscaptor) erscheinen dagegen relativ spät und sind erstmals für das Mittlere Pleistozän auf dem Japanischen Archipel verzeichnet. Dies trifft auch auf die beiden heutigen Angehörigen der Japanischen Spitzmulle zu.[107][108] Auf dem ostasiatischen Festland lassen sich im Pliozän mit Yunoscaptor noch die Neuweltmaulwürfe im Yushe-Becken verzeichnen. Gleichzeitig sind aber in der Region auch erstmals die Vorfahren des Kurzgesichtmaulwurf (Scaptochirus) als weitere grabende Maulwürfe präsent.[95]

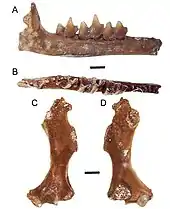
Auch in Nordamerika setzen sich im Pliozän und Pleistozän weitgehend die heutigen Linien durch. Auf der bedeutenden Gray Fossil site des Unteren Pliozän in Tennessee kam mit Magnatalpa noch ein Vertreter der Desmane vor, der in etwa die Ausmaße des Russischen Desman (Desmana) aufwies. Die dortige Landschaft wurde zusätzlich von den Vorfahren des Amerikanischen Spitzmulls bewohnt, ebenso wie jenen des Haarschwanzmaulwurfs. Als weiteres Mitglied der Neuweltmaulwürfe war auch Mioscalops präsent.[109] Abseits davon erreichen die Vorfahren des Ostamerikanischen Maulwurfs im östlichen, südlichen und zentralen Nordamerika im Pliozän und Pleistozän ein weites Verbreitungsgebiet.[110][111] In Teilen entsprach es in etwa dem, das auch die frühen Angehörigen des Sternmulls nutzten.[104] Im westlichen Teil des Kontinentes breiteten sich in diesem Zeitraum hingegen frühe Angehörige der Westamerikanischen Maulwürfe aus. Zu nennen ist die bedeutende Hagerman Fauna als artenreiche Säugetiergemeinschaft, hinzu kommen mehrere pleistozäne Fundstellen in Kalifornien.[112][113][51]
Forschungsgeschichte
Etymologie
Die Maulwürfe waren den Menschen in vielen Regionen seit langem bekannt. Der im Deutschen übliche Trivialname „Maulwurf“ (zumeist für den Europäischen Maulwurf verwendet), bezieht sich, anders als es sich phonetisch vermuten ließe, nicht auf das „Maul“ und der möglicherweise damit bewegten Erde. Letzteres ist von Maulwürfen nicht bekannt und wird überwiegend von bodenwühlenden Nagetieren getätigt. Die Bezeichnung „Maulwurf“ hat ihren Ursprung im mittelhochdeutschen Wort moltwërf. Dieses leitet sich vermutlich vom althochdeutschen mū-wërf ab, wobei mū soviel wie „Haufen“ und wërf soviel wie „werfen“ bedeuten. Da für das Wort mū im Deutschen keine Entsprechung vorliegt, wurde es teilweise zu „mul“ im Sinne von „Mull“, „Staub“ umgedeutet, woraus im Hochdeutschen schließlich volksetymologisch „Maul“ entstand.[114] Der Wortteil „Mull“ findet sich noch bei einigen Maulwurfsarten wie dem Sternmull und den verschiedenen Spitzmullen, ebenso wie bei nicht verwandten, ökologisch aber ähnlich lebenden Tieren, etwa den Beutelmullen, den Blindmullen, den Graumullen, den Nacktmullen, den Goldmullen oder den Gürtelmullen. Auf eine ähnliche Wurzel geht das englische Wort mole für „Maulwurf“ zurück. Es leitet sich vom altenglischen moldwarp her, das auf mold für „Erde“ und weorpan für „durch“ basiert.[115] In zahlreichen romanischen Sprachen hingegen werden Variationen des lateinischen Wortes talpa für die Maulwürfe verwendet, so etwa das französische taupe oder das spanische topo. Als weitere Bedeutung für talpa wird für mehrere Sprachen im romanischen Raum zudem „Sohle“ angegeben. Einige Sprachwissenschaftler nehmen an, dass beides auf eine ältere indogermanische Wurzel zurückgeht, in der die ursprüngliche Bedeutung in etwa „Erde“ lautete.[116] Zahlreiche slawische Bezeichnungen wie krtek im Tschechischen oder крот (krot) im Russischen basieren auf dem altkirchenslawischen Wort крътъ, dass sich wiederum von krŭtŭ herleitet. Einige Linguisten übersetzen dies mit „sich schnell bewegen“.[117][118]
Erstbeschreibungen

Aufgrund der langen Bekanntheit der Maulwürfe wurden die Tiere schon früh in naturforschenden Berichten erwähnt. Bereits Albert Seba führte in seinem im Jahr 1734 erschienenen Thesaurus mehrere Formen auf, die sowohl in der Alten wie auch in der Neuen Welt beheimatet waren.[119] Sebas teils pragmatisches Vorgehen unterschied die Maulwürfe unter anderem nach ihrer Färbung, wobei nicht jedes Exemplar einem heutigen Vertreter exakt zugeordnet werden kann.[120] Der Thesaurus diente jedoch als Basis für die Arbeiten nachfolgender Naturforscher. Hierzu gehört unter anderem auch Carl von Linné, der im Jahr 1758 in der zehnten Ausgabe seines Werkes Systema naturae mehreren Arten der Maulwürfe erstmals einen heute wissenschaftlich gültigen Artnamen gab. Linnaeus erkannte zu diesem Zeitpunkt aber noch nicht die nähere Verwandtschaft der einzelnen Formen, da er lediglich den Europäischen Maulwurf der Gattung Talpa zuwies (zusätzlich noch den Kap-Goldmull, der jedoch mit den Maulwürfen nicht näher verwandt ist). Den Ostamerikanischen Maulwurf und den Sternmull sah er hingegen in einer näheren Beziehung zu den Rotzahnspitzmäusen, den Russischen Desman ordnete er zu den Bibern.[121] Die Gattungsbezeichnung Talpa war allerdings schon vor Linnaeus in Gebrauch und wurde unter anderem von Seba genutzt.[122]

Die Bezeichnung Talpa als Basis für die weitere wissenschaftliche Benennung der Maulwürfe war damit bereits in der zweiten Hälfte des 18. Jahrhunderts etabliert. Weitere Gattungen wurden in der Folgezeit eingeführt, etwa Desmana im Jahr 1777, Scalopus im Jahr 1803 und Condylura im Jahr 1812. Rund 60 Jahre nach Linnaeus’ wegweisendem Werk zur systematischen Gliederung der Tierwelt fasste Johann Fischer von Waldheim die Maulwürfe erstmals stärker zusammen. Er etablierte hierfür im Jahr 1817 die Bezeichnung Talpini, die er allerdings abweichend von der heutigen Auffassung als ein Taxon oberhalb der Tribusebene sah. Innerhalb der Talpini führte er die damals bekannten Formen wie die Eurasischen Maulwürfe und den Ostamerikanischen Maulwurf, analog zu Linnaeus aber auch die Goldmulle. In einem ähnlichen Zusammenhang hierzu hatte Fischer von Waldheim drei Jahre zuvor die Maulwürfe in einer übergeordneten Gruppe namens Talpinorum eingeordnet. Fischer von Waldheim gilt heute als Erstautor der Familie der Maulwürfe, wobei sowohl 1814 als auch 1817 als Ursprungsjahr angegeben werden.[123][124] Die heute korrekte Form des Familiennamens als Talpidae stammt allerdings von John Edward Gray aus dem Jahr 1825. Gray vereinte innerhalb der Talpidae aber nicht nur die Maulwürfe, sondern auch die Goldmulle, Rotzahnspitzmäuse, Igel, Tenreks und Spitzhörnchen, die er jeweils auf tribunaler Ebene voneinander abtrennte. Innerhalb der von ihm als eigentliche Maulwürfe gesehenen Gruppe gliederte er lediglich die Eurasischen Maulwürfe ein, den Sternmull und den Ostamerikanischen Maulwurf wies er den Goldmullen zu, die Desmane wiederum den Rotzahnspitzmäusen.[125]
Gliederungsversuche
In den folgenden Dekaden beschäftigten sich zahlreiche Naturforscher mit den Maulwürfen und versuchten sich an einer sinnvollen Gliederung dieser im Verhältnis zu anderen Insektenfresser-Gruppen. Hierzu gehören unter anderem Auguste Pomel im Jahr 1848 oder Paul Gervais im Jahr 1854.[126] Aus heutiger Sicht handelte es sich dabei zumeist um sehr weitläufige Auffassungen der Maulwürfe, die wie zuvor auch andere Gruppen wie die Goldmulle oder die Schlitzrüssler mit einschlossen. Auch bei Wilhelm Peters Arbeit aus dem Jahr 1864 war dies der Fall, in der er aber erstmals die Insektenfresser in zwei große Formengruppen aufteilte und diese nach der Ausprägung des Blinddarms unterschied (und für die Ernst Haeckel drei Jahre später die Bezeichnungen Menotyphla für jene mit und Lipotyphla für jene ohne Blinddarm einführte).[127] Erst St. George Mivart reduzierte im Jahr 1867 die Maulwürfe weitgehend auf die Familie im heutigen Sinne. Hierbei unterschied er zwei Unterfamilien, die er einerseits mit „Talpina“ für alle damals bekannten grabenden Maulwürfe und andererseits mit „Myogalina“ benannte, letztere Bezeichnung bezieht sich auf die Desmane, schloss aber auch die Spitzmulle mit ein. Die Unterscheidung erfolgte unter anderem anhand des Bewegungsapparates, der sich bei den „Talpina“ laut Mivart durch einen massiven Oberarmknochen und ein kurzes, kompaktes Schlüsselbein auszeichnete. Bei den „Myogalina“ waren beide Knochen eher schlank gebaut.[128]
Mivarts Schema wurde nur wenig später, im Jahr 1875, von Theodore Gill übernommen, der aber beide Unterfamilien noch einmal deutlicher differenzierte. Gill sah innerhalb seiner von ihm als Talpinae bezeichneten Unterfamilie zwei größere Einheiten. Er grenzte dadurch eine eurasische Gruppe grabender Maulwürfe („Talpae“) von einer nordamerikanischen („Condylurae“) ab. Die Unterfamilie der Myogalinae mit ihren weitgehend oberirdisch lebenden Vertretern teilte er in die eigentlichen Desmane („Mygalae“) und die Spitzmulle und Spitzmausmaulwürfe („Urotrichini“) auf.[129] Im Jahr 1883 veröffentlichte George Edward Dobson seine umfassende Abhandlung zu den Insektenfressern. In dieser widmete er sich umfänglich auch den Maulwürfen. Seine Gliederung der Familie bestand aus überarbeiteten Versionen von Mivart und Gill. Er führte im Unterschied zu Gill allerdings die Spitzmausmaulwürfe als eigenständige höhere Gruppe („Uropsili“). Ihnen gegenüber stand eine Einheit aus allen anderen Maulwürfen. Für diese behielt er die bestehende Aufteilung weitgehend bei, sah aber die Desmane („Myogalae“ bei Dobson) und die verschiedenen Spitzmulle („Urotrichini“) als eigenständige Einheiten an. Die grabenden Maulwürfe wies Dobson als eigene Gruppe aus, unterschieden in die eurasischen „Talpae“ und die nordamerikanischen „Condylurae“ und „Scalopes“, letzteren beiden gestand er eine engere Bindung zu.[5]
Anfang des 20. Jahrhunderts beschäftigte sich Oldfield Thomas mehrfach mit den Maulwürfen. Auf ihn basiert ein neues Gliederungsschema, in dem er insgesamt fünf Unterfamilien ausdifferenzierte. In einer kurzen Abhandlung aus dem Jahr 1912 benannte er mit den Talpinae und Scalopinae die alt- beziehungsweise neuweltlichen grabenden Maulwürfen, zusätzlich unterschied er die Desmaninae mit den wasserlebenden Formen Eurasiens, die Condylurinae mit dem ebenfalls semi-aquatischen Sternmull Nordamerikas und die Uropsilinae mit den bodenlebenden Spitzmausmaulwürfen. Die Spitzmulle zeigten nach Thomas Beziehungen zu den Scalopinae.[130] Für die nächsten 50 Jahre sollte dieses Gliederungskonstrukt Bestand haben und wurde so auch von George Gaylord Simpson in seiner generellen Taxonomie der Säugetiere aus dem Jahr 1945 übernommen.[131] Erst Leigh Van Valen sah sich 1967 veranlasst, innerhalb einer umfassenden Studie zu den rezenten und fossilen Insektenfressern eine Neubewertung vorzunehmen. In seinem Ansatz bestanden nur drei Unterfamilien, neben den Talpinae zusätzlich noch die Desmaninae und die Uropsilinae. Alle anderen Gruppen verschob er auf die Ebene der Tribus, wodurch die grabenden Maulwürfe Eurasiens und Nordamerikas (Talpini und Scalopini), der Sternmull (Condylurini) sowie die Spitzmulle (Urotrichini) innerhalb der Talpinae positioniert waren. Allerdings trennte Van Valen mit den Scaptonychini den Langschwanzmaulwurf von den übrigen Spitzmullen als eigenständige Tribus ab.[132] Diese Gliederung adoptierten dann auch spätere Autoren weitgehend, sie findet sich demnach unter anderem bei J. Howard Hutchison im Jahr 1968,[60] aber auch bei Malcolm C. McKenna und Susan K. Bell in ihrer Klassifikation der Säugetiere aus dem Jahr 1997.[133] Rainer Hutterer hob im Jahr 2005 innerhalb des Werkes Mammal Species of the World die neuweltlichen grabenden Maulwürfe wieder auf das Niveau der Unterfamilie und stellte sie den altweltlichen gegenüber, die Spitzmausmaulwürfe formten die dritte Unterfamilie der Maulwürfe. Innerhalb der Scalopinae schloss er auch den Sternmull ein, während er die Desmane den Talpinae zuschlug. Als weitere Neuerung löste Hutterer den Amerikanischen Spitzmull als eigene Tribus der Neurotrichini aus den restlichen Spitzmullen heraus. In seiner Auffassung verblieben alle Spitzmulle bei den Talpinae.[134] Tatsächliche Klarheit über die Gliederung der Familie lieferten ab den 2010er Jahren verstärkt vorgenommene molekulargenetische Studien, die zu der heutigen, allgemein akzeptierten Aufteilung der Maulwürfe führten.[53]
Maulwürfe und Menschen
Bedrohung und Schutzstatus

In früheren Zeiten wurden einige Maulwurfsarten wegen ihres Felles bejagt, wozu unter anderem der Russische Desman, der Sibirische Maulwurf und der Kaukasische Maulwurf gehören. Diese Praxis spielt jedoch heute keine Rolle mehr. Heutige Konflikte zwischen Menschen und Maulwürfen basieren vorwiegend auf der Grabetätigkeit dieser Tiere. Obwohl die meisten Arten sich von Wirbellosen ernähren und nur sporadisch pflanzliche Nahrung verzehren, kann ihre grabende Lebensweise Schäden an Pflanzenwurzeln anrichten. Nicht selten nutzen Ratten und Mäuse die durch Maulwürfe gegrabenen Gänge, um an die Wurzeln und Knollen zu gelangen. Die Hügel und Tunnel können unter Umständen zur Beschädigung von Mäh- und Erntegeräten führen. Im gärtnerischen und landwirtschaftlichen Bereich werden zudem die Maulwurfshügel häufig missbilligt, was zur Verfolgung der Tiere führt. Auch die Lebensraumzerstörung stellt für einige Arten eine Bedrohung dar. Verursacht wird diese nicht nur durch die Ausdehnung menschlicher Siedlungs- und Wirtschaftsflächen, sondern auch durch die zunehmende Austrocknung der Böden oder die Zerstückelung und Verunreinigung der Gewässernetze. Die IUCN listet rund drei Fünftel aller Arten als nicht bedroht, je ein weiteres Fünftel sind in ihrem Bestand im unterschiedlichen Maße bedroht oder aber die Informationslage ist zu dürftig, als dass sie in eine spezielle Gefährdungskategorie eingestuft werden können. Allerdings berücksichtigt die Naturschutzorganisation nicht alle bekannten Arten.[135] Für die meisten Vertreter der Maulwürfe kommt nur ein Vor-Ort-Schutz in Frage, da Aufzuchtprogramme in menschlicher Obhut bisher kaum Erfolg zeigten. Zahlreiche Arten sind in geschützten Gebieten präsent.[135][1]
Kulturelle Bezüge
_047.jpg.webp)
Die charakteristische und mitunter den Menschen störende Lebensweise der grabenden Maulwürfe fand recht früh Einzug in das Kunstschaffen. Bereits im Jahr 1874 ließ Wilhelm Busch in seinem Bildergedicht Der Maulwurf aus der Sammlung Dideldum! den Gärtner Knoll einen Kampf mit einem Maulwurf austragen.[136] Anfang des 20. Jahrhunderts erschien der Kinderbuchklassiker Der Wind in den Weiden von Kenneth Grahame, in dem ein vermenschlichter und manierlicher Maulwurf als Protagonist auftritt. Von herausragender Bedeutung ist auch die Figur Der kleine Maulwurf. Im Jahr 1956 von Zdeněk Miler erschaffen, erlebte sie zahlreiche Trickfilm-Abenteuer in über fünf Dekaden Fernsehgeschichte.[137][1] Weiter erwähnenswert sind Der Maulwurf Grabowski aus dem gleichnamigen Kinderbuch von Luis Murschetz oder Werner Holzwarths kleiner Maulwurf, der wissen wollte, wer ihm auf den Kopf gemacht hat. Im Kinderfernsehen des Deutschen Fernsehfunks kam neben Figuren wie zum Beispiel Herrn Fuchs und Frau Elster und dem Raben Meister Schwarzrock im Märchenwald auch die des Maulwurfs Buddelflink vor. Im allgemeinen Sprachgebrauch stand die Grabetätigkeit der Maulwürfe Pate für die Bezeichnung „Maulwurf“ für einen Informanten, der sich getarnt in eine Organisation eingeschlichen hat und dort (verdeckt) Informationen für seine Auftraggeber beschafft.
Literatur
- Tom S. Kemp: The Origin and Evolution of Mammals. Reprinted edition. Oxford University Press, Oxford u. a. 2005, ISBN 0-19-850761-5
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 ISBN 978-84-16728-08-4
- Ronald M. Nowak: Walker's Mammals of the World. 2 Bände. 6. Auflage. Johns Hopkins University Press, Baltimore MD u. a. 1999, ISBN 0-8018-5789-9
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. A taxonomic and geographic Reference. 2 Bände. 3. Auflage. The Johns Hopkins University Press, Baltimore MD 2005, ISBN 0-8018-8221-4
Einzelnachweise
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 ISBN 978-84-16728-08-4
- Matthew J. Mason: Evolution of the Middle Ear Apparatus in Talpid Moles. Journal of Morphology 267, 2006, S. 678–695
- Daisuke Koyabu, Misato Hosojima und Hideki Endo: Into the dark: patterns of middle ear adaptations in subterranean eulipotyphlan mammals. Royal Society Open Science 4, 2017, S. 170608, doi:10.1098/rsos.170608
- Eugene Barrow und Norman Macleod: Shape variation in the mole dentary (Talpidae: Mammalia). Zoological Journal of the Linnean Society 153, 2008, S. 187–211
- George Edward Dobson: A Monograph of the Insectivora, systematic and anatomical. Part II. London, 1883, S. 126–172 ()
- Patricia S. Meier, Constanze Bickelmann, Torsten M. Scheyer, Daisuke Koyabu und Marcelo R. Sánchez-Villagra: Evolution of bone compactness in extant and extinct moles (Talpidae): Exploring humeral microstructure in small fossorial mammals. BMC Evolutionary Biology 13, 2013, S. 55, doi:10.1186/1471-2148-13-55
- Berry Campbell: The shoulder anatomy of the moles. A study in phylogeny and adaptation. The American Journal of Anatomy 64, 1939, S. 1–39
- Charles A. Reed: Locomotion and Appendicular Anatomy in Three Soricoid Insectivores. The American Midland Naturalist 45, 1951, S. 513–671 ()
- Marcelo R Sánchez-Villagra, Peter R. Menke und Jonathan H. Geisler: Patterns of evolutionary transformation in the humerus of moles (Talpidae, Mammalia): A character analysis. Mammal Study 29 (2), 2004, S. 163–170, doi:10.3106/mammalstudy.29.163
- Marcelo R. Sánchez-Villagra und Peter R. Menke: The mole’s thumb – evolution of the hand skeleton in talpids (Mammalia). Zoology 108, 2005, S. 3–12
- Christian Mitgutsch, Michael K. Richardson, Rafael Jiménez, José E. Martin, Peter Kondrashov, Merijn A. G. de Bakker und Marcelo R. Sánchez-Villagra: Circumventing the polydactyly ‚constraint‘: The mole’s ‚thumb‘. Biology Letters 8, 2011, S. 74–77, doi:10.1098/rsbl.2011.0494
- Constanze Bickelmann, Christian Mitgutsch, Michael K. Richardson, Rafael Jiménez, Merijn A. G. de Bakker und Marcelo R. Sánchez-Villagra: Transcriptional heterochrony in talpid mole autopods. EvoDevo 3, 2012, S. 16, doi:10.1186/2041-9139-3-16
- Andrzej Myrcha: Comparative studies on the morphology of the stomach in the Insectivora. Acta Theriologica 12 (14), 1967, S. 223–244
- W. Robert Eadie: The male accessory reproductive glands of Condylura with notes on a unique prostatic secretion. The Anatomical Record 101, 1948, S. 59–78
- Adriana Watkins Sinclair, Stephen E. Glickman, Laurence Baskin und Gerald R. Cunha: natomy of Mole External Genitalia: Setting the Record Straight. The Anatomical Record 299, 2016, S. 385–399, doi:10.1002/ar.23309
- Adriana Watkins Sinclair, Stephen E. Glickman, Keneth C. Catania, Akio Shinohara, Laurence Baskin und Gerald R. Cunha: Comparative morphology of the penis and clitoris in four species of moles (Talpidae). Journal of Experimental Zoology B (Molcular and Developmental Evolution) 328B, 2017, S. 275–294, doi:10.1002/jez.b.22732
- Kenneth C. Catania: A Comparison of the Eimer’s Organs of Three North American Moles: The Hairy-Tailed Mole (Parascalops breweri), the Star-Nosed Mole (Condylura cristata), and the Eastern Mole (Scalopus aquaticus). The Journal of Comparative Neurology 354, 1995, S. 150–160
- Kenneth C. Catania: Epidermal Sensory Organs of Moles, Shrew-Moles, and Desmans: A Study of the Family Talpidae with Comments on the Function and Evolution of Eimer’s Organ. Brain Behavior and Evolution 56, 2000, S. 146–174
- Kenneth C. Catania und Fiona E. Remple: Asymptotic prey profitability drives star-nosed moles to the foraging speed limit. Nature 433, 2005, S. 519–522
- Shin-ichiro Kawada, Masashi Harada, Kazuhiru Koyasu und Sen-ichi Oda: Karyological on the short-faced mole, Scaptochirus moschatus (Insectivore, Talpidae). Mammal Study 27, 2002, S. 91–94, ()
- Shin-ichiro Kawada, Song Li, Ying-Xiang Wang, Orin B. Mock, Sen-ichi Oda und Kevin M. Campbell: Karyotype evolution of shrew moles (Soricomorpha: Talpidae). Journal of Mammalogy 89 (6), 2008, S. 1428–1434
- Shin-ichiro Kawada, Nguyen Truong Son und Dang Ngoc Can: Karyological diversity of talpids from Vietnam (Insectivora, Talpidae). In: Dang Ngoc Can, Hideki Endo, Nguyen Truong Son, Tatsuo Oshida, Le Xuan Canh, Dang Huy Phuong, Darrin Peter Lunde, Shin-ichiro Kawada, Akiko Hayashida und Montoki Sasaki (Hrsg.): Checklist of wild mammals of Vietnam. Hanoi, 2008, S. 384–390
- Kai He, Jin-Huan Wang, Wei-Ting Su, Quan Li, Wen-Hui Nie und Xue-Long Jiang: Karyotype of the Gansu mole (Scapanulus oweni): further evidence for karyotypic stability in talpid. Mammal Study 37, 2012, S. 341–348, doi:10.3106/041.037.0408
- Svetlana V. Pavlova, Vladimir S. Lebedev, Vasily D. Yakushov, Yongke Zhu, Yun Fang, Yue‑Hua Sun und Boris I. Sheftel: High diversity of small insectivorous mammals on Qinghai–Tibet Plateau and first description of karyotype for four endemics of China. Scientific Reports 11, 2021, S. 24496, doi:10.1038/s41598-021-03809-4
- Di Xu, Mengyao Sun, Zenghao Gao, Yiping Zhou, Qingqian Wang und Lei Chen: Comparison and Phylogenetic Analysis of Mitochondrial Genomes of Talpidae Animals. Animals 13 (2), 2023, S. 186, doi:10.3390/ani13020186
- Anna A. Bannikova, Elena D. Zemlemerova, Paolo Colangelo, Mustafa Sözen, M. Sevindik, Artem A. Kidov, Ruslan I. Dzuev, Boris Kryštufek und Vladimir S. Lebedev: An underground burst of diversity – a new look at the phylogeny and taxonomy of the genus Talpa Linnaeus, 1758 (Mammalia: Talpidae) as revealed by nuclear and mitochondrial genes. Zoological Journal of the Linnean Society 175, 2015, S. 930–948
- E. D. Zemlemerova, A. A. Bannikova, V. S. Lebedev, V. V. Rozhnov und A. V. Abramov: Secrets of the underground Vietnam: an underestimated species diversity of Asian moles (Lipotyphla: Talpidae: Euroscaptor). Proceedings of the Zoological Institute RAS 320 (2), 2016, S. 193–220
- İslam Gündüz, Sadık Demirtaş, Metin Silsüpür, Medine Özmen, P. David Polly und David T. Bilton: Notes from the Anatolian underground: two new mole taxa from Eastern Turkey, together with a revised phylogeny of the genus Talpa (Mammalia: Eulipotyphla: Talpidae). Zoological Journal of the Linnean Society, 2023, S. zlad049, doi:10.1093/zoolinnean/zlad049
- Kai He, Triston G Eastman, Hannah Czolacz, Shuhao Li, Akio Shinohara, Shin-ichiro Kawada, Mark S Springer, Michael Berenbrink und Kevin L Campbell: Myoglobin primary structure reveals multiple convergent transitions to semiaquatic life in the world’s smallest mammalian divers. eLife 10, 2021, S. e66797, doi:10.7554/eLife.66797
- Gerhard Heldmaier und Gerhard Neuweiler: Vergleichende Tierphysiologie. Springer Verlag Berlin Heidelberg, 2004, ISBN 978-3-642-62374-5, S. 241
- Wolfgang Jelkmann, Walter Oberthür, Traute Kleinschmidt und Gerhard Braunitzer: Adaptation of hemoglobin function to subterranean life In the mole, Talpa europaea. Respiration Physiology 46 (1), 1981, S. 7–16, doi:10.1016/0034-5687(81)90064-5
- Kevin L. Campbell, Jay F. Storz, Anthony V. Signore, Hideaki Moriyama, Kenneth C. Catania, Alexander P. Payson, Joseph Bonaventura, Jörg Stetefeld und Roy E. Weber: Molecular basis of a novel adaptation to hypoxic-hypercapnia in a strictly fossorial mole. BMC Evolutionary Biology 10, 2010, S. 214, doi:10.1186/1471-2148-10-214
- Francisca M. Real, Stefan A. Haas, Paolo Franchini, Peiwen Xiong, Oleg Simakov, Heiner Kuhl, Robert Schöpflin, David Heller, M.-Hossein Moeinzadeh, Verena Heinrich, Thomas Krannich, Annkatrin Bressin, Michaela F. Hartmann, Stefan A. Wudy, Dina K. N. Dechmann, Alicia Hurtado, Francisco J. Barrionuevo, Magdalena Schindler, Izabela Harabula, Marco Osterwalder, Michael Hiller, Lars Wittler, Axel Visel, Bernd Timmermann, Axel Meyer, Martin Vingron, Rafael Jiménez, Stefan Mundlos, Darío G. Lupiáñez: The mole genome reveals regulatory rearrangements associated with adaptive intersexuality. Science 370 (6513), 2020, S. 208–214, doi:10.1126/science.aaz2582
- Rafael Jiménez, Miguel Burgos, L. Caballero und Rafael Díaz de la Guardia: Sex reversal in a wild population of Talpa occidentalis (Insectivora, Mammalia). Genetic Research 52, 1988, S. 135–140
- Rafael Jiménez, Miguel Burgos, Antonio Sánchez, Andrew H. Sinclair, Francisco J. Alarcón, Juan J. Marín, Esperanza Ortega, Rafael Díaz de la Guardia: Fertile females of the mole Talpa occidentalis are phenotypic intersexes with ovotestes. Development 118, 1993, S. 1303–1311
- F. David Carmona, Masaharu Motokawa, Masayoshi Tokita, Kimiyuki Tsuchiya, Rafael Jiménez und Marcelo R. Sánchez-Villagra: The Evolution of Female Mole Ovotestes Evidences High Plasticity of Mammalian Gonad Development. Journal of Experimental Zoology 310B (3), 2008, S. 259–266
- Duncan B. Leitch, Diana K. Sarko und Kenneth. C. Catania: Brain mass and cranial nerve size in shrews and moles. Scientific Reports 4, 2014, S. 6241, doi:10.1038/srep06241
- Kenneth C. Catania: Mole senses. Current Biology 29, 2019, S. R819–R828
- Kenneth C. Catania: All in the Family – Touch Versus Olfaction in Moles. The Anatomical Record 303, 2020, S. 65–76, doi:10.1002/ar.24057
- Kenneth C. Catania: Evolution of brains and behavior for optimal foraging: A tale of two predators. PNAS 109 (suppl. 1), 2012, S. 10701–10708, doi:10.1073/pnas.1201885109
- Kenneth C. Catania: Stereo and serial sniffing guide navigation to an odour source in a mammal. Nature Communications 4, 2012, S. 1441, doi:10.1038/ncomms2444
- Kenneth C. Catania: Underwater 'sniffing' by semi-aquatic mammals. Nature 444, 2006, S. 1024–1025, doi:10.1038/4441024a
- Yu. F. Ivlev, M. V. Rutovskaya und O. S. Luchkina: The Use of Olfaction by the Russian Desman (Desmana moschata L.) during Underwater Swimming. Doklady Biological Sciences 452, 2013, S. 280–283
- Naohiko Sagara und Yu Fukasawa: Inhabitant changes in long-term mole nesting at the same site, revealed by observing mushroom fruiting at the site. Mammalia 78 (3), 2014, S. 383–391, doi:10.1515/mammalia-2013-0073
- Merle Streitberger und Thomas Fartmann: Bodenstörende Ökosystem-Ingenieure im mitteleuropäischen Grasland und ihre Bedeutung für die Biodiversität. Eine Analyse am Beispiel der Gelben Wiesenameise und des Europäischen Maulwurfs. Naturschutz und Landschaftsplanung 49 (8), 2017, S. 252–259
- Selina Brace, Jessica A. Thomas, Love Dalén, Joachim Burger, Ross D. E. MacPhee, Ian Barnes und Samuel T. Turvey: Evolutionary History of the Nesophontidae, the Last Unplaced Recent Mammal Family. Molecular Biology and Evolution 33 (12), 2016, S. 3095–3103, doi:10.1093/molbev/msw186
- Christophe J. Douady, Pascale I. Chatelier, Ole Madsen, Wilfried W. de Jong, Francois Catzeflis, Mark S. Springer und Michael J. Stanhope: Molecular phylogenetic evidence confirming the Eulipotyphla concept and in support of hedgehogs as the sister group to shrews. Molecular Phylogenetics and Evolution 25 (1), 2002, S. 200–209
- Mark S. Springer, William J. Murphy und Alfred L. Roca: Appropriate fossil calibrations and tree constraints uphold the Mesozoicdivergence of solenodons from other extant mammals. Molecular Phylogenetics and Evolution 121, 2018, S. 158–165, doi:10.1016/j.ympev.2018.01.007
- Jun J. Sato, Satoshi D. Ohdachi, Lazaro M. Echenique-Diaz, Rafael Borroto-Páez, Gerardo Begué-Quiala, Jorge L. Delgado-Labañino, Jorgelino Gámez-Díez, José Alvarez-Lemus, Son Truong Nguyen, Nobuyuki Yamaguchi und Masaki Kita: Molecular phylogenetic analysis of nuclear genes suggests a Cenozoic over-water dispersal origin for the Cuban solenodon. Scientific Reports 6, 2016, S. 31173, doi:10.1038/srep31173
- Anthony D. Barnosky: A Skeleton of Mesoscalops (Mammalia, Insectivora) from the Miocene Deep River Formation, Montana, and a Review of the Proscalopid Moles: Evolutionary, Functional, and Stratigraphic Relationships. Journal of Vertebrate Paleontology 1 (3/4), 1981, S. 285–339
- Gregg F. Gunnell, Thomas M. Bown, J. Howard Hutchison und Jonathan I. Bloch: Lipotyphla. In: C. M. Janis, G. F. Gunnell, und M. D. Uhen (Hrsg.): Evolution of Tertiary Mammals of North America, Vol. 2. Cambridge University Press, 2007, S. 89–125
- Kenneth D. Rose: The beginning of the age of mammals. Johns Hopkins University Press, 2006, S. 1–428 (S. 154–155)
- Kai He, Akio Shinohara, Kristofer M. Helgen, Mark S. Springer, Xue-Long Jiang, Kevin L. Campbell: Talpid Mole Phylogeny Unites Shrew Moles and Illuminates Overlooked Cryptic Species Diversity. Molecular Biology and Evolution 34 (1), 2016, S. 78–87
- A. A. Bannikova, E. D. Zemlemerova, V. S. Lebedev, D. Yu. Aleksandrov, Yun Fang und B. I. Sheftel: Phylogenetic Position of the Gansu Mole Scapanulus oweni Thomas, 1912 and the Relationships Between Strictly Fossorial Tribes of the Family Talpidae. Doklady Biological Sciences 464, 2015, S. 230–234
- Akio Shinohara, Kevin L. Campbell und Hitoshi Suzuki: Molecular phylogenetic relationships of moles, shrew moles, and desmans from the new and old worlds. Molecular Phylogenetics and Evolution 27, 2003, S. 247–258, doi:10.1016/S1055-7903(02)00416-5
- Marcelo R. Sánchez-Villagra, Inés Horovitz und Masaharu Motokawa: A comprehensive morphological analysis of talpid moles (Mammalia) phylogenetic relationships. Cladistics 22, 2006, S. 59–88
- Akio Shinohara, Hitoshi Suzuki, Kimiyuki Tsuchiya, Ya-Ping Zhang, Jing Luo, Xue-Long Jiang, Ying-Xiang Wang und Kevin L. Campbell: Evolution and Biogeography of Talpid Moles from Continental East Asia and the Japanese Islands Inferred from Mitochondrial and Nuclear Gene Sequences. Zoological Science 21, 2004, S. 1177–1185
- Masaharu Motokawa: Phylogenetic relationships within the family talpidae. Journal of Zoology 263, 2004, S. 147–157
- Lars W. van den Hoek Ostende: Insectivore faunas from the Lower Miocene of Anatolia – Part 5: Talpidae. Scripta Geologica 122, 2001, S. 1–35
- J. Howard Hutchison: Fossil Talpidae (Insectivora, Mammalia) from the Later Tertiary of Oregon. Bulletin of the Museum of Natural History University of Oregon 11, 1968, S. 1–117 (S. 58–96)
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. 3. Auflage. 2005
- Zhong-Zheng Chen, Shui-Wang He, Wen-Hao Hu, Wen-Yu Song, Kenneth O Onditi, Xue-You Li und Xue-Long Jiang: Morphology and phylogeny of scalopine moles (Eulipotyphla: Talpidae: Scalopini) from the eastern Himalayas, with descriptions of a new genus and species. Zoological Journal of the Linnean Society 193 (2), 2021, S. 432–444, doi:10.1093/zoolinnean/zlaa172
- Bernard Sigé, Jean-Yves Crochet und Allen Insole: Les plus vieilles taupes. Geobios, Memoire Special, 1, 1977, S. 141–157
- David L. Harrison, Paul J. J. Bates, Malcolm Pearch, Chris Michaels und David J. Ward: New additions to the late middle Eocene mammal fauna ofCreechbarrow, Dorset, southern England. Cainozoic Research 9 (1), 2012, S. 65–85
- Jerry J. Hooker: Skeletal adaptations and phylogeny of the oldest mole Eotalpa (Talpidae, Lipotyphla, Mammalia) from the UK Eocene: The beginning of the fossoriality in moles. Palaeontology 59 (2), 2016, S. 195–216
- Karen J. Lloyd und Jaelyn J. Eberle: A new talpid from the late Eocene of North America. Acta Palaeontologica Polonica 53 (3), 2008, S. 539–543
- Richard Smith: Présence du genre Eotalpa (Mammalia, Talpidae) dans l'Oligocène inférieur de Belgique (Formation de Borgloon, MP 21). Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre 77, 2007, S. 159–165
- Reinhard Ziegler, Thomas Dahlmann und Gerhard Storch: Oligocene–Miocene vertebrates from the Valley of Lakes (central Mongolia): Morphology, phylogenetic and stratigraphic implications: 4. Marsupialia, Erinaceomorpha and Soricomorpha (Mammalia). Annalen des Naturhistorischen Museums Wien 108A, 2007, S. 53–164
- Reinhard Ziegler: Moles (Talpidae, Mammalia) from Early Oligocene karstic fissure fillings in South Germany. Geobios 45, 2012, S. 501–513, doi:10.1016/j.geobios.2011.11.017
- Marguerite Hugueney und Olivier Maridet: Evolution of Oligo-Miocene talpids (Mammalia, Talpidae) in Europe: focus on the genera Myxomygale and Percymygale n. gen. Historical Biology 30 (1/2), 2018, S. 267–275, doi:10.1080/08912963.2017.1282477
- G. Sansalone, T. Kotsakis, A. H. Schwermann, L. W. Van den Hoek Ostende und P. Piras: When moles became diggers:Tegulariscaptorgen. nov., from the early Oligocene of south Germany, and the evolution of talpid fossoriality. Journal of Systematic Palaeontology 16 (8), 2018, S. 645–657, doi:10.1080/14772019.2017.1329235
- Achim H. Schwermann und Thomas Martin: A partial skeleton of Geotrypus antiquus (Talpidae, Mammalia) from the Late Oligocene of the Enspel fossillagerstätte in Germany. Paläontologische Zeitschrift 86, 2012, S. 409–439, doi:10.1007/s12542-012-0129-1
- Achim H. Schwermann und Richard S. Thompson: Extraordinarily preserved talpids (Mammalia, Lipotyphla) and the evolution of fossoriality. Journal of Vertebrate Paleontology 35 (4), 2015, S. e934828, doi:10.1080/02724634.2014.934828
- Lars W. van den Hoek Ostende: The Talpidae (Insectivora, Mammalia) of Eggingen-Mittelhart (Baden-Württemberg, F.R.G.) with special reference to the Paratalpa-Desmanodon lineage. Stuttgarter Beiträge zur Naturkunde Serie B 152, 1989, S. 1–29
- Reinhard Ziegler: Talpidae (Insectivora, Mammalia) aus dem Oberoligozän und Untermiozän Süddeutschlands. Stuttgarter Beiträge zur Paläontologie Serie B 167, 1990, S. 1–80
- Antje Schreuder: A revision of the fossil water-moles (Desmaninae). Archives Néerlandais de Zoologie 4, 1940, S. 201–233
- A. V. Lopatin: An Oligocene Mole (Talpidae, Insectivora, Mammalia) from Mongolia. Paleontological Journal 36 (5), 2002, S. 531–534
- A. V. Lopatin: A New Record of Mongoloscapter (Talpidae, Lipotyphla, Mammalia) from the Oligocene of Mongolia. Paleontological Journal 52 (6), 2018, S. 677–681
- William W. Korth, Clint A. Boyd, Robert J. Emry und Jeff J. Person: Additional small mammals from the Oligocene Brule Formation (Orellan-Whitneyan) of Southwestern North Dakota. Paludicola 14 (2), 2023, S. 57–74 ()
- Lars W. van den Hoek Ostende, Peter Joniak, Bora Rojay, Cathelijn Aten, Melike Bilgin und Pablo Pelaez-Campomanes: Early Miocene insectivores of Gökler (Kazan Basin, Central Anatolia, Turkey). Palaeobiodiversity and Palaeoenvironments 99, 2019, S. 701–722, doi:10.1007/s12549-019-00396-1
- Froukje N. van der Sar, Roos van Glabbeek, Wilma Wessels, Zoran Marković und Hans de Bruijn: Insectivores and marsupials from the upper Oligocene of Banovići (Bosnia and Herzegovina). Journal of Vertebrate Paleontology 37 (6), 2017, S. e1368529, doi:10.1080/02724634.2017.1368529
- H. De Bruijn und C. G. Rümke: On a peculiar mammalian association from the Miocene of Oschiri (Sardinia). II. Koninklijke Nederlandse Akademie Van Wetenschappen B 77 (1), 1974, S. 59–79
- Gerhard Storch und Thomas Dahlmann: Desmanelia rietscheli, ein neuer Talpide aus dem Obermiozän von Dorn-Dürkheim 1, Rheinhessen (Mammalia, Lipotyphla). Carolinea – Beiträge zur naturkundlichen Forschung in Südwestdeutschland 58, 2000, S. 65–69 ()
- A. V. Lopatin: Early Miocene small mammals from the North Aral Region (Kazakhstan) with special reference to their biostratigraphic significance. Paleontological Journal 38 (suppl 3), 2004, S. S217–S323
- Oleg G. Bendukidze, Hans de Bruijn und Lars W. van der Hoek Ostende: A revision of Late Oligocene associations of small mammals from the Aral Formation (Kazakhstan) in the National Museum of Georgia, Tbilissi. Palaeodiversity 2, 2009, S. 343–377
- Reinhard Ziegler: Moles (Talpidae) from the late Middle Miocene of South Germany. Acta Palaeontologica Polonica 48 (4), 2003, S. 617–648
- J. Howard Hutchison: Notes on type specimens of European Miocene Talpidae and a tentative Classification of Old World Tertiary Talpidae (Insectivora: Mammalia). Geobios 7 (3), 1974, S. 211–256
- Thomas Dahlmann und Salih Doğan: Gerhardstorchia nomen novum: a new name for Storchia Dahlmann 2001 (Mammalia: Lipotyphla: Talpidae: Desmaninae). Paläontologische Zeitschrift 85, 2011, S. 91
- Raef Minwer-Barakat, Antonio García-Alix, Elvira Martín-Suárez und Matthijs Freudenthal: Early Pliocene Desmaninae (Mammalia, Talpidae) from Southern Spain and the Origin of the Genus Desmana. Journal of Vertebrate Paleontology 40 (5), 2020, S. e1835936, doi:10.1080/02724634.2020.1835936
- P. Colangelo, A. A. Bannikova, B. Kryštufek, V. S. Lebedev, F. Annesi, E. Capanna und A. Loy: Molecular systematics and evolutionary biogeography of the genus Talpa (Soricomorpha: Talpidae). Molecular Phylogenetics and Evolution 55, 2010, S. 372–380
- Wang Xiao-Ming, Qiu Zhu-Ding, Li Qiang, Yukimitsu Tomida, Yuri Kimura, Zhijie Jack Tseng und Wang Hong-Jiang: A New Early to Late Miocene Fossiliferous Region In Central Nei Mongol: Lithostratigraphy and Biostratigraphy In Aoerban Strata. Vertebrata Palasiatica 47 (2), 2009, S. 111–134
- Gerhard Storch und Shuding Qiu: The Neogene mammalian faunas of Ertemte and Harr Obo in Inner Mongolia (Nei Mongol), China. 2. Moles – Insectivora: Talpidae. Senckenbergiana lethaea 64, 1983, S. 89–127
- Leonid L. Voyta, Thomas Mörs und Margarita A. Erbajeva: Erinaceomorpha and Soricomorpha (Mammalia) of the Miocene Tagay fauna (Olkhon Island, Lake Baikal, Eastern Siberia): A preliminary report. Palaeobiodiversity and Palaeoenvironments 102, 2022, S. 897–914, doi:10.1007/s12549-022-00557-9
- Gerhard Storch und Shuding Qiu: Insectivores (Mammalia: Erinaceidae, Soricidae, Talpidae) from the Lufeng hominoid locality, late Miocene of China. Geobios 24 (5), 1991, S. 601–621
- Lawrence J. Flynn und Wen-Yu Wu: The Lipotyphla of Yushe Basin. In: Lawrence J. Flynn und Wen-Yu Wu (Hrsg.): Late Cenozoic Yushe Basin, Shanxi Province, China: Geology and Fossil Mammals, Volume II: Small Mammal Fossils of Yushe Basin, Vertebrate Paleobiology and Paleoanthropology. Springer Science+Business Media B.V., 2017, S. 11–26
- William W. Korth: Fossil small mammals from the Harrison Formation (Late Arikareean: Earliest Miocene), Cherry County, Nebraska. Annals of the Carnegy Museum 61 (2), 1992, S. 69–131 ()
- Richard H. Tedford: Clarendonian Insectivora from the Ricardo Formation, Kern County, California. Bulletin, Southern California Academy of Sciences 60, 1961, S. 57–76
- Walter W. Dalquest: Mammals of the Coffee Ranch local fauna, Hemphilian of Texas. The Pearce-Sellards Series 38, 1983, S. 1–41
- James E. Martin: A rare occurence of the fossil water mole Gallardia (Desmanini, Talpidae) from the Neogene in North America. Proceedings of the South Dakota Academy of Science 96, 2017, S. 94–98
- Thomas M. Bown: The Fossil Insectivora of Lemoyne Quarry (Ash Hollow Formation, Hemphillian), Keith County, Nebraska. Transactions of the Nebraska Academy of Sciences and Affiliated Societies 284, 1980, S. 99–122
- Stanisław Skoczeń: Condylurini Dobson, 1883 (Insectivora, Mammalia) in the Pliocene of Poland. Acta Zoologica Cracoviensia 21, 1976, S. 291–314
- Stanisław Skoczeń: New records of Parascalops, Neurotrichus and Condylura (Talpinae, Insectívora) from the Pliocene of Poland. Acta Theriologica 38 (2), 1993, S. 125–137
- Barbara Rzebik-Kowalska: Review of the Pliocene and Pleistocene Talpidae (Soricomorpha, Mammalia) of Poland. Palaeontologia Electronica 17 (2), 2014, S. 17.2.26A, doi:10.26879/457
- Gabriele Sansalone, Tassos Kotsakis und Paolo Piras: Condylura (Mammalia, Talpidae) reloaded: New insights about the fossil representatives of the genus. Palaeontologia Electronica 19 (3), 2016, S. 54A ()
- Wighart von Koenigswald: Mittelpleistozäne Kleinsäugerfauna aus der Spaltenfüllung Petersbuch bei Eichstätt. Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie 10, 1970, S. 407–432
- Gabriele Sansalone, Tassos Kotsakis und Paolo Piras: Talpa fossilis or Talpa europaea? Using geometric morphometrics and allometric trajectories of humeral moles remains from Hungary to answer a taxonomic debate. Palaeontologia Electronica 18 (2), 2015, S. 42A ()
- Yoshinari Kawamura: Quaternary Rodent Faunas in the Japanese Islands (Part 1). Memoirs of the Faculty of Science, Kyoto University, Series of Geology and Mineralogy 53 (1/2), 1988, S. 31–348 ()
- Yoshinari Kawamura: Quaternary Mammalian Faunas in the Japanese Islands. The Quaternary Research 30 (2), 1991, S. 213–220 ()
- Danielle E. Oberg und Joshua X. Samuels: Fossil moles from the Gray fossil site (Tennessee): Implications for diversification and evolution of North American Talpidae. Palaeontologia Electronica 25 (3), 2022, S. a33, doi:10.26879/1150
- Walter W. Dalquest: Vertebrate fossils from the Blanco local fauna of Texas. Occasional Papers from the Museum Texas Tech University 30, 1975, S. 1–52
- Gary S. Morgan, Paul L. Sealey und Spencer George Lucas: Pliocene and Early Pleistocene (Blancan) vertebrates from the Palomas Formation in the vicinity of Elephant Butte Lake and Caballo, Sierry County, Southwestern New Mexico. New Mexico Museum of Natural History and Science, Bulletin 53, 2011, S. 664–736
- J. Howard Hutchison: Late Pliocene (Blancan) Scapanus (Scapanus) (Talpida: Mammalia) from the Glenns Ferry Formation of Idaho. PaleoBios 12 (45), 1987, S. 1–7
- J. Howard Hutchison: Moles of the Scapanus latimanus group (Talpidae, Insectivora) from the Pliocene and Pleistocene of California. Contributions in Science, Natural History Museum of Los Angeles County 386, 1987, S. 1–15
- Friedrich Kluge und Elmar Seebold: Etymologisches Wörterbuch der deutschen Sprache. Verlag Walter de Gruyter, Berlin 1989, 1–822 (S. 468)
- John Considine: Stephen Skinner’s Etymologiconand other English etymological dictionaries 1650-1700. Studia Etymologica Cracoviensia 14, 2009, S. 123–151
- Simona Georgescu und Oana-Alexandra Buzea: Sur l’origin du roum. Talpă, ou la recontre entre ‘plante du pied’ et ‘taupe’. Philologica Jassyensia 15 (2), 2019, S. 189–202
- Robert Orr: Again the *ŭ-stems in Common Slavic. Journal of Slavic Linguistics 4 (2), 1996, S. 312–343
- Ecaterina Pleșca: Denumirile cîrtiței: Între motivație și etimologie. Paralele lexico-semantice de areal. Revistă de lingvistică și știință literară 1–2, 2008, S. 92–100
- Albert Seba: Locupletissimi rerum naturalium thesauri accurata descriptio, et iconibus artificiosissimis expressio, per universam physices historiam. Amsterdam, 1734, Taf. 32 sowie S. 51 ()
- Neal Woodman: American Recent Eulipotyphla. Nesophontids, Solenodons, Moles, and Shrews in the New World. Smithsonian Institution Scholary Press, 2018, S. 1–107
- Carl von Linné: Systema naturae. 10. Auflage, 1758, Band 1, S. 52, 53, 59 ()
- T. S. Palmer: Index Generum Mammalium: A List of the Genera and Families of Mammals. North American Fauna 23, 1904, S. 1–984 (S. 659) ()
- Johann Fischer von Waldheim: Zoognosia tabulis synopticis illustrata. Moskau, 1814, S. X ()
- Johann Fischer von Waldheim: Adversaria zoologica. Mémoires de la Société impériale des naturalistes de Moscou 5, 1817, S. 357–428 ()
- John Edward Gray: An outline of an attempt at the disposition of Mammalia into tribes and families, with a list of the genera apparently appertaining to each tribe. Annals of Philosophy N. S. 10, 1825, S. 337–344 ()
- Paul Gervais: Histoire naturelle des mammifères, avec l’indication de leurs moeurs, et de leurs rapports avec les arts, le commerce et l’agriculture. Band 1, Paris, 1854, S. 1–420 (S. 239–259) ()
- Wilhelm Peters: Über die Säugethiergattung Selenodon. Abhandlungen der Königlichen Akademie der Wissenschaften zu Berlin 1863, S. 1–22 ()
- St. George Jackson Mivart: Notes on the Osteology of the Insectivora. Journal of Anatomy and Physiology 2 (1), 1868, S. 117–154 (S. 150) ()
- Theodore Gill: Synopsis of the insectivorous mammals. Bulletin of the United States Geological and Geographical Survey of the Territories 1, 1874–1875, S. 91–140 (S. 102) ()
- Oldfield Thomas: On the collection of mammals from the Tsin-ling mountains, Central China, presented by Mr. G. Fenwick Owen to the National Museum. Annals and Magazine of Natural History 8 (10), 1912, S. 395–403 ()
- George Gaylord Simpson: The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History 85, 1945, S. 1–350 (S. 52–53)
- Leigh van Valen: New Paleocene Insectivores and Insectivore classification. Bulletin of the American Museum of Natural History 135, 1967, S. 217–284
- Malcolm C. McKenna und Susan K. Bell: Classification of mammals above the species level. Columbia University Press, New York, 1997, S. 1–631 (S. 279–283)
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. A taxonomic and geographic Reference. 2 Bände. 3. Auflage. Johns Hopkins University Press, Baltimore MD 2005, ISBN 0-8018-8221-4 ()
- Gefährdungsgrad der einzelnen Arten laut IUCN Suche nach „talpidae“ in der Roten Liste gefährdeter Arten der IUCN., abgerufen am 18. August 2023
- Wilhelm Busch: Dideldum! Heidelberg, 1874 ()
- Nadine Kulbe: Maulwurfmonitoring. Über das Vorkommen der Familie der Talpidae in wissenschaftlichen Sammlungen. Dresden, 2020
Weblinks
- Uni Zürich: Wie der Maulwurf zu seinen zwölf Fingern kommt