Sumpfkrüge
Die Sumpfkrüge (Heliamphora) sind eine Pflanzengattung in der Familie der Schlauchpflanzengewächse (Sarraceniaceae). Es sind präkarnivore Pflanzen.
| Sumpfkrüge | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Heliamphora chimantensis (in Kultur) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Heliamphora | ||||||||||||
| Benth. |
Beschreibung




Vegetativer Habitus
Die Arten der Gattung Heliamphora sind ausdauernde krautige Pflanzen. Bis auf die bis zu 4 Meter hoch wachsende Heliamphora tatei bilden alle Arten bodenständige Rosetten. Das einfache oder verzweigte, robuste Rhizom bildet horizontale Ausläufer und wächst dann aufwärts, so entstehen weitere Wuchspunkte. Ausgehend von einer einzelnen Pflanze können so (insbesondere bei Heliamphora chimantensis) über Jahrzehnte umfangreiche Horste aus tausenden von Individuen entstehen, die mehrere Quadratmeter groß sind. Die faserigen, braunen Wurzeln erreichen eine Länge von bis zu 25 Zentimetern.
Blätter
Alle Heliamphora-Arten weisen einen altersabhängigen Blattdimorphismus auf, was bedeutet, dass die Blätter einer jungen Pflanze sich in der Form deutlich von jenen einer älteren unterscheiden. Die jungen Blätter sind bei allen Arten annähernd gleich geformt; die arttypischen Merkmale werden erst nach bis zu 4 Jahren ausgeformt.
Die drei bis zehn gänzlich aufrechten bis schwach zum Zentrum der Rosette gebogenen, trichterförmigen bis zylindrischen Blätter sind Fallen vom Fallgrubentyp und erreichen eine Höhe von 8 bis 10 Zentimetern bei Heliamphora minor und Heliamphora pulchella über 20 Zentimetern bei Heliamphora nutans bis hin zu etwa einem halben Meter bei Heliamphora ionasii.
Die Fallen besitzen am äußeren Rand des Peristoms (der Umrandung der Schlauchöffnung) ein sehr kleines, oft kontrastreich gefärbtes, zipfel- oder helmförmig über die Schlauchöffnung gebogenes aufrechtes Deckelrudiment. Eine Ausnahme bildet hier die Art Heliamphora sarracenioides, deren Deckel ähnlich einem Dach die gesamte Öffnung seitlich und senkrecht zum Falleninneren überdeckt und nur eine kleine Öffnung zur Rosettenmitte hin übrig lässt. Die Außenseite der Fallen ist haarlos, die Innenseite des Trichters ist abschnittweise mit abwärtsgerichteten Haaren bewachsen, die die Flucht von hineingefallener Beute erschwert. Die Form der Fallen verhindert ein Hinausfliegen hineingefallener Insekten, da durch deren Flügelbewegung in den Fallen ein Abwärtssog entsteht. An der Vorderseite des Schlauches haben die Schläuche eine sogenannte Ala, einen Flügel, der dicht mit Nektarien besetzt ist. Auf halber Höhe der Ala befindet sich eine kleine Öffnung, durch die überschüssiges Regenwasser ablaufen kann, eine leichte Behaarung darin fungiert als Sieb und verhindert das Hinausgeschwemmtwerden von Beute.
Blüten, Früchte und Samen
An den langen, aufrechten Blütenstandsschäften stehen die wenigen Blüten in einem traubigen Blütenstand. Die zwittrigen Blüten sind radiärsymmetrisch. Die zumeist vier, selten fünf oder sechs, zugespitzt-eiförmigen Kronblätter sind weiß (manchmal grün überhaucht) bis rot, bei einigen Arten verfärben sie sich im Lauf der Anthese von weiß zu rot. Es sind zehn bis zwanzig Staubblätter vorhanden. Der Fruchtknoten ist oberständig und dicht behaart. Es ist nur ein Griffel vorhanden.


Ökologie
Karnivorie
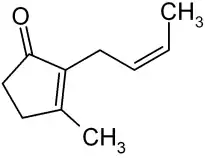
Wege der Anlockung von Insekten sind zum einen die auffällige Ausfärbung der Blätter, insbesondere des Deckelrudiments und zum anderen die Reflexion des UV-Lichtes allein durch die innere Blattoberfläche[1], die für Insektenaugen die Fallenöffnung deutlich gegen die Umgebung und die Blattaußenseite abhebt. Unterstützt wird dies durch die Absonderung von Nektar durch Nektardrüsen an der Ala sowie besonders stark am Deckelrudiment, an dem auch weitere chemische Lockstoffe ausgeschieden werden. Die Schläuche lassen sich in vier Zonen unterteilen: Zone 1 ist das Deckelrudiment, Zone 2 der oberste Schlauchabschnitt und die Zonen 3 und 4 die jeweils tieferliegenden Abschnitte des Schlauches. Jede dieser Zonen hat dabei beim Beutefang eine spezielle Funktion, für die sie entsprechend unterschiedlich ausgestattet ist. Die erste Zone, das dicht mit Nektarien besetzte Deckelrudiment, scheidet größere Mengen an stark duftendem Nektar aus sowie Sarracenin (ein antimikrobiell wirkendes und Fressfeinde abhaltendes Iridoid), Cineron (eine Insektizidvorstufe), mehrere als Insektenpheromone wirksame Ester, Phenylacetaldehyd (eine Chemikalie zur Kommunikation unter Insekten), Erucamid (ein Gleitmittel, das zum einen das Sekret flüssig hält und zugleich den Fang der Insekten begünstigt), zahlreiche Alkane sowie Xylol[2]. Diese Stoffe dienen vielfach der Anziehung bzw. dem Fang von Beuteinsekten, haben aber möglicherweise auch weitere, z. B. betäubende oder verklebende Wirkungen.
Insekten, die die Pflanzen besuchen, streben zum Deckelrudiment (Bodeninsekten entlang der Ala, Fluginsekten unmittelbar) und stürzen entweder von dort in den Schlauch oder werden von weiteren Nektarien zum Schlauchinneren geführt. So gelangen sie in die dicht mit abwärtsweisenden Haaren besetzte Zone 2, die nur mehr inselartig mit Nektardrüsen besetzt ist. Da die abwärtsweisenden Haare den Weg zurück zum Rand versperren, kann sich das Insekt nur weiter abwärts bewegen, bis es an den abrupt beginnenden Ansatz der Zone 3 gelangt, dessen Oberfläche unbehaart und glatt ist, so dass das Tier den Schlauch hinabrutscht und in die Flüssigkeit von Zone 4 stürzt, wo die Verdauung stattfindet.
Nur von einer Sumpfkrugart, Heliamphora tatei, konnte bisher die Produktion von Enzymen durch die Pflanzen selbst nachgewiesen werden, auch hier jedoch nur bei geschlossenen und jungen Schläuchen. Tests bei einigen anderen Arten blieben erfolglos. Damit ist (mit eben der Ausnahme von Heliamphora tatei) die Gattung nicht als karnivor im strengen Sinne, sondern als „präkarnivor“ einzustufen. Auf welchem Weg genau die Verdauung der Beute geschieht, ist bisher nicht gesichert, zwei Hypothesen gelten dabei als wahrscheinlich: Zum einen könnte eine in der Flüssigkeit ansässige Bakterienfauna die Beutetiere zersetzen. Zum anderen wäre eine sogenannte Autolyse, also eine Selbstauflösung der Beute möglich, denn der Hauptanteil der Beute der Sumpfkrüge besteht zu 80 bis 95 % aus Ameisen mit einer häufig räuberischen Lebensweise, die Verdauungsenzyme in ihrem Inneren mitbringen. Da Enzyme katalytisch wirken, sich also nicht verbrauchen und sich auch nicht zersetzen, könnten sie sich akkumulieren und so zur Verdauung weiterer Beutetiere dienen, die wiederum erneut Enzyme einbringen. Möglich ist auch, dass mehrere dieser Faktoren zusammenspielen.
Neben Ameisen (vorzugsweise der Gattungen Linepithema, Camponotus und Solenopsis) erbeuten Sumpfkrüge zu nennenswerten Anteilen auch Käfer, Mücken, Trauermücken, Zweiflügler, Hautflügler, vereinzelt aber auch größere Beutetiere wie Schmetterlinge, Skorpione oder Steinfliegen.
Sumpfkrüge bilden viele der Merkmale ihrer Karnivorie ausschließlich an äußerst lichtintensiven Standorten aus, die Überschattung durch Vegetation oder Landschaftsmerkmale führt neben einer relativ langsamen Wuchsgeschwindigkeit auch zum Ausbleiben der Nektarsekretion, der starken Ausfärbung, der Behaarung und zu nur schwach ausgeprägten Deckelrudimenten. Das hat zu der Vermutung geführt, dass Sumpfkrüge die Karnivorie nur in Situationen gesteigerten Nährstoffbedarfs (z. B. im Falle schnellen Wuchses durch optimale Standortbedingungen) als zusätzliche Nährstoffquelle benötigen.
Verbreitung
_edit.jpg.webp)
Die Gattung Heliamphora ist fast ausschließlich auf den sogenannten Tepuis, den schwer zugänglichen Tafelbergen auf der Hochebene im Grenzgebiet zwischen Brasilien, Venezuela und Guyana, beheimatet. Nur Heliamphora heterodoxa und Heliamphora sarracenioides sind auch in den tieferen Lagen dieser Region zu finden.
Evolution
Lange Zeit wurde Heliamphora wegen ihres einfachen Fallenbaus für eine der ursprünglichsten fleischfressenden Pflanzen gehalten. Neuere genetische und blütenbiologische Untersuchungen legen aber nahe, dass Heliamphora wahrscheinlich eine lange Entwicklungsgeschichte hat und ihre einfache Form erst in jüngerer Vergangenheit angenommen hat.
Systematik

Botanische Geschichte
Die Gattung Heliamphora wurde 1840 durch George Bentham in Proceedings of the Linnean Society of London, 1, S. 53 aufgestellt.[3]
Die Typusart ist Heliamphora nutans, die 1840 durch George Bentham anhand eines von Robert Hermann Schomburgk gesammelten Exemplars erstbeschrieben wurde. Der Gattungsname Heliamphora ist aus den altgriechischen Bezeichnungen für „Sumpf“ (ἕλος helos) und „Krug“ (άμφορεύς amphoreus) zusammengesetzt. Im englischen Sprachraum wurde der Gattungsname gelegentlich fälschlicherweise mit „Sun Pitcher“ (vom griechischen helios) übersetzt.
Über neunzig Jahre hinweg war die Gattung Heliamphora monotypisch, bevor Henry Allan Gleason 1931 Heliamphora tatei, Heliamphora tyleri und 1939 Heliamphora minor beschrieb. Zwischen 1978 und 1984 bearbeiteten Julian Alfred Steyermark (der bereits 1951 die Gattung um Heliamphora heterodoxa ergänzt hatte) und Bassett Maguire die Gattung weiter und beschrieben neben zahlreichen infraspezifischen Taxa auch zwei neue Arten, Heliamphora ionasii und Heliamphora neblinae. Seit 1999 hat – resultierend aus mehrfachen Expeditionen sowie der Sichtung vorhandenen Herbarmaterials – sich vor allem eine Gruppe deutsche Forscher, bestehend aus Thomas Carow, Peter Harbarth, Joachim Nerz, Andreas Wistuba und Andreas Fleischmann, um die Gattung verdient gemacht und kontinuierlich neue Arten beschrieben, die Beschreibung weiterer Taxa steht noch aus.
Arten
Bis 2010 wurden 19 Arten beschrieben, damit ist Heliamphora die artenreichste Gattung der Sarraceniaceae. Die genaue Taxonomie dieser Gattung ist noch nicht zweifelsfrei geklärt; vor allem der Status von Heliamphora tyleri und Heliamphora macdonaldae ist umstritten, wobei sich bezüglich des Status von Heliamphora tyleri die Ansicht einer Synonymie zu Heliamphora tatei durchsetzt.
- Heliamphora chimantensis Wistuba, Carow & Harbarth
- Heliamphora ciliata Wistuba, Nerz & A.Fleischm.
- Heliamphora elongata Nerz
- Heliamphora exappendiculata (Maguire & Steyerm.) Nerz & Wistuba
- Heliamphora folliculata Wistuba, Harbarth & Carow
- Heliamphora glabra (Maguire) Nerz, Wistuba & Hoogenstr.
- Heliamphora heterodoxa Steyerm.
- Heliamphora hispida Wistuba & Nerz
- Heliamphora huberi A.Fleischm., Wistuba & Nerz
- Heliamphora ionasii Maguire
- Heliamphora macdonaldae Gleason
- Heliamphora minor Gleason
- Heliamphora neblinae Maguire
- Heliamphora nutans Benth.
- Heliamphora pulchella Wistuba, Carow, Harbarth & Nerz
- Heliamphora sarracenioides Carow, Wistuba & Harbarth
- Heliamphora tatei Gleason
- Heliamphora tyleri Gleason
- Heliamphora uncinata Nerz, Wistuba & A.Fleischm.
Literatur
- Wilhelm Barthlott, Stefan Porembski, Rüdiger Seine, Inge Theisen: Karnivoren. Biologie und Kultur fleischfressender Pflanzen. Ulmer, Stuttgart 2004, ISBN 3-8001-4144-2.
- Klaus Jaffe, Fabian Michelangeli, Jorge M. Gonzalez, Beatriz Miras, Marie Christine Ruiz: Carnivory in Pitcher Plants of the Genus Heliamphora (Sarraceniaceae). In: The New Phytologist. Band 122, Nr. 4, 1992, ISSN 0028-646X, S. 733–744, JSTOR:2557442.
- Francis Ernest Lloyd: The Carnivorous Plants (= A New Series of Plant Science Books. 9, ZDB-ID 415601-8). Chronica Botanica Company, Waltham MA 1942.
- Stewart McPherson: Pitcher Plants of the Americas. The McDonald & Woodward Publishing Company, Blacksburg VA 2007, ISBN 978-0-939923-74-8.
Einzelnachweise
- Daniel M. Joel, Barrie E. Juniper, Amots Dafni: Ultraviolet patterns in the traps of carnivorous plants. In: The New Phytologist. Bd. 101, Nr. 4, 1985, S. 585–593, JSTOR:2432892.
- Klaus Jaffé, Murray S. Blum, Henry M. Fales, Robert T. Mason, Aivlé Cabrera: On Insect Attractants from Pitcher Plants of the Genus Heliamphora (Sarraceniaceae). In: Journal of Chemical Ecology. Band 21, Nr. 3, 1995, ISSN 0098-0331, S. 379–384, doi:10.1007/BF02036725.
- Heliamphora bei Tropicos.org. Missouri Botanical Garden, St. Louis, abgerufen am 29. August 2017.