Süßgräser
Die Süßgräser (Poaceae = Gramineae) sind eine Pflanzenfamilie in der Ordnung der Grasartigen (Poales). Mit etwa 12.000 Arten in rund 780 Gattungen[1] sind sie eine der größten Familien innerhalb der Blütenpflanzen. Sie sind weltweit in allen Klimazonen verbreitet und durch eine typische grasartige Gestalt gekennzeichnet.
| Süßgräser | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Grannen-Ruchgras (Anthoxanthum aristatum) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Poaceae | ||||||||||||
| (R.Br.) Barnhart |



Viele Arten der Süßgräser gehören zu den ältesten Nutzpflanzen und sind seit alters für den Menschen von lebenswichtiger Bedeutung. Alle Getreide wie Weizen, Roggen, Gerste, Hafer, Hirse, Mais und Reis zählen zu dieser Pflanzengruppe. Sie stellen in Form von Marktfrüchten oder als Viehfutter in der Veredelung heute die Basis für die Ernährung der Weltbevölkerung dar. Als Vorkommen in Gras- oder Grünland – wie Wiesen und Weiden, aber auch Steppen und Savannen – prägen sie in weiten Teilen der Erde das Landschaftsbild.
Lebenszyklus und Morphologie
Süßgräser umfassen sowohl kurzlebige als auch langlebige Arten. Sie weisen eine charakteristische Morphologie sowohl der vegetativen als auch der generativen Organe auf, mit einem gemeinsamen „grasförmigen“ Grundbauplan der verschiedenen Arten. Gräser sind meist schlankwüchsig und verfügen über lange, dünne, durch Knoten gegliederte Halme, parallelnervige, lange Blätter und oft unauffällige, einfache Blütenstände. Innerhalb der Unterfamilien, Tribus und Gattungen sind dagegen deutliche taxonspezifische Abwandlungen der Merkmale vorhanden.
Lebensformen und Ausdauer
Viele Arten sind einjährig und schließen ihren gesamten Lebenszyklus in einer Vegetationsperiode ab. Sie leben meist nur wenige Monate und überdauern die ungünstige Jahreszeit als Samen im Boden. Diese Formen, zu denen auch viele Getreidesorten gehören, werden Therophyten genannt.
Andere Arten sind mehrjährig und damit Hemikryptophyten. Sie verfügen über bodennahe Erneuerungsknospen und überdauern ungünstige Zeiten geschützt durch den Boden, Laubstreu oder Schnee. Dazu gehören zweijährige Arten, die im Laufe des Sommers oder Herbstes keimen und erst im folgenden Jahr Früchte und Samen bilden, ebenso wie ausdauernde und mehrjährige Arten, die wenige oder viele Jahre leben. Diese besitzen überwinterungsfähige Horste oder Rosetten. Die Individuen einer Generation ausdauernder Arten können bis zu 400 Jahre alt werden, so zum Beispiel der Rot-Schwingel (Festuca rubra).[2] Die Gemeine Quecke (Elymus repens) ist ein Beispiel dafür, dass sich Gräser aus Ausläuferfragmenten erneuern können (Rhizom-Geophyten).[3] Die meisten Hemikryptophyt-Arten sind krautig; deren Halme nach etwa einem Jahr Lebensdauer oberirdisch absterben. Ausnahmen bilden holzige Bambus-Arten (Bambuseae), deren Triebe dickwandig und fest sind und mehrere Jahrzehnte ausdauern können.
Wuchsformen und Wurzeln
Etliche Süßgräser sind zart gebaut und werden nur wenige Zentimeter groß (z. B. Einjähriges Rispengras). Andere Arten haben verholzte Halme und erreichen Wuchshöhen bis zu 40 Metern und mehr, wie beispielsweise die Bambus-Art Dendrocalamus giganteus. Ein- und zweijährige Arten haben gewöhnlich einzelne oder wenige Triebe in lockeren Büscheln mit weicheren Blättern. Bei diesen Süßgräsern tragen alle oder die meisten der Sprossachsen Blütenstände. Die ausdauernden Arten bilden in den meisten Fällen festere Halme und Blattspreiten und neben blühenden Trieben eine größere oder kleinere Anzahl an nicht blühenden Trieben. Sie wachsen in lockeren oder dichten Horsten oder rasenförmig. Letztere Wuchsform ergibt sich, indem sich die Pflanzen entweder über mehr oder weniger lange, oberirdisch kriechende, grünliche oder rötliche Sprossachsen, namentlich Stolonen (z. B. das Weiße Straußgras) oder über unterirdische, weiße oder braune Rhizome ausbreiten (z. B. die Kriech-Quecke). Außer an der Farbe lassen sich die beiden Typen von sich an den Knoten bewurzelnden Ausläufern auch daran unterscheiden, dass Stolonen an jedem Knoten (Nodus) über vollständige Blätter mit Blattscheide und Blattspreite verfügen, Rhizome dagegen an diesen Punkten lediglich kleine, dünne, schuppenförmige Niederblätter entwickeln. Bei horstbildenden Arten bilden sich nur kurze Ausläufer, oder die jungen Seitentriebe entwickeln sich innerhalb der Blattscheiden des Muttertriebes (intravaginal), so beim Schaf-Schwingel (Festuca ovina). Auf diese Weise entsteht durch die gedrängt stehenden Triebe die typische büschelige, dicht horstige Wuchsform vieler Gräser. Wachsen die Triebe die untere Blattscheide durchstoßend (extravaginal), ist der Aufwuchs meist locker-horstig oder rasenförmig (z. B. Rot-Schwingel). Die meisten Süßgräser sind Flachwurzler; sie bilden keine Haupt- und Pfahlwurzeln. Am Stängelgrund und an den Knoten der Ausläufer werden zahlreiche sprossbürtige Wurzeln gebildet, die ihrerseits Seitenwurzeln 1. und 2. Ordnung entwickeln können. Auf diese Weise können Wurzelsysteme von beachtlicher Länge entstehen. So kann sich eine einzige Pflanze des Rot-Schwingels etwa 250 Meter im Durchmesser ausbreiten.[2]
Halme und Blätter
.jpg.webp)
Die Stängel der Süßgräser werden als Halme (über althochdeutsch halm ableitbar von germanisch halma, „Stroh, Getreidestängel, Grasstängel“[4]) bezeichnet. Sie sind meist hohl und rund. Nur wenige Grasarten besitzen markige Stängel. Sie sind durch feste, mit Gewebe gefüllte Knoten (Nodien) gegliedert. Die Abschnitte zwischen den Knoten werden als Internodien bezeichnet. Unmittelbar oberhalb der Knoten liegen die Wachstumszonen, die Halme wachsen also mit eingelagerten Meristemen. An diesen Stellen setzen die faserigen Verstärkungselemente, die den Halmen zusätzliche Stabilität und Zugfestigkeit verleihen, aus. Die Halme bleiben auf diese Weise beweglich und biegsam. Sie sind so in der Lage, sich nach Wind- und Regeneinwirkung wieder aufzurichten. Sie können entweder senkrecht hochwachsen, von einem gebogenen Grund aufsteigen oder gänzlich am Boden niederliegend wachsen. Grashalme variieren in Größe, Festigkeit und Zahl der Knoten. Sie sind im Querschnitt meist rund, selten etwas zusammengedrückt wie beim Zusammengedrückten Rispengras (Poa compressa). Bei einigen Süßgrasarten sind die untersten Internodien mehr oder weniger angeschwollen und verdickt. Die Halme etlicher Gräser sind unverzweigt, bei einigen Arten bilden sich von den Knospen in den Blattachseln ausgehende Seitenzweige. Die Beblätterung der Halme ist bei Süßgräsern immer wechselständig und fast ausnahmslos zweizeilig (distich) – im Gegensatz zur dreizeiligen Beblätterung der Sauergräser (Cyperaceae).
Die Blätter der Süßgräser bestehen immer aus zwei verschiedenen Abschnitten: der Blattscheide und der Blattspreite. Die Blattscheide entspricht dem Blattgrund, setzt am Knoten an und umschließt das Internodium bis fast zum nächsten Halmknoten. Die Scheiden sind bei der Mehrzahl der Gräser an einer Seite offen. Bei wenigen Grasarten sind die Ränder verwachsen und damit die Blattscheiden röhrig geschlossen, wenngleich sie früh im oberen Bereich aufreißen. Während die basalen Blattscheiden die Wachstumspunkte der jungen Triebe schützen, erfüllen diejenigen an den Halmen diese Schutzfunktion für die dortigen Wachstumszonen oberhalb der Knoten und sorgen außerdem für zusätzliche Stabilität. Der obere Teil der Blattscheiden kann bauchig aufgeblasen sein. Die Vorderseite des Blattscheidenendes kann in mehr oder weniger spitze, meist stängelumfassende „Öhrchen“ ausgezogen sein oder Büschel von Haaren tragen. Die Blattscheide geht am oberen Ende in die vom Halm abstehende Blattspreite über. Diese ist flach, gerollt oder gefaltet; stets länglich und mehr oder weniger spitz zulaufend. Sie zeigt die kennzeichnende Paralleladerung einkeimblättriger Pflanzen. Jede Blattader entspricht einem Leitbündel, der dem Stofftransport und der Aussteifung der Blattfläche dient.
Am plötzlichen Übergang von der Blattscheide zur Blattspreite sitzt bei den meisten Arten ein häutiges Anhängsel, das Blatthäutchen (Ligula). Es erscheint meistens als farbloser, durchscheinender Fortsatz der Oberhaut auf der Innenseite der Blattscheide und stellt eine Verlängerung der inneren Epidermis der Blattscheide dar. Es schützt vor Verletzungen durch Reibung des sich beim Wind hin und her bewegenden Halmgliedes sowie vor dem Eindringen von Schmutz und Parasiten in den Raum zwischen Halm und Scheide. Wegen seiner Gestaltungsvielfalt ist das Blatthäutchen für die Artbestimmung hilfreich. Es ist behaart oder unbehaart, kragenförmig, zugespitzt, langgezogen, sehr kurz oder sehr lang. Teilweise ist das Blatthäutchen durch eine Reihe von Haaren ersetzt, selten fehlt es ganz.
Blütenstände und Blüten


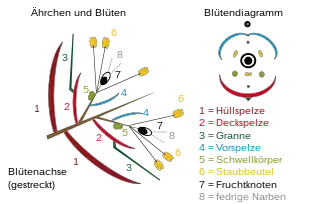

Die Blütenstände (Infloreszenzen) der Süßgräser bestehen aus einer Vielzahl von Teilblütenständen, seltener Einzelblüten, die in Ähren, Rispen und Trauben an einer Blütenstandsachse (Rhachis spicae) angeordnet sind. Die Teilblütenstände werden als Ährchen bezeichnet. Sie bestehen ihrerseits aus ein- bis mehreren, überwiegend zweigeschlechtigen Blüten. Sitzen die Ährchen ungestielt direkt an der Blütenstandsachse, handelt es sich um eine Ähre. Bei Fingergräsern befinden sich mehrere Ähren am Halmende in fingerartiger Anordnung. Sogenannte Kolben entstehen durch Abwandlungen von Ähren durch Vergrößerung des Achsengewebes. In Trauben befinden sich die Ährchen an unverzweigten Stielen. Die Ährchen können alle in die gleiche Richtung weisen (einseitswendig) oder sich in zwei Reihen an gegenüberliegenden Seiten der Achse befinden. Sind die Seitenäste einseits- oder allseitswendig verzweigt, handelt es sich um Rispen. In Ährenrispen oder Scheinähren sind die Seitenäste so kurz, dass die Blütenstände äußerlich wie Ähren erscheinen. Erst beim Umbiegen einer solchen Ährenrispe werden die tatsächlichen Verzweigungsmuster erkennbar.
Süßgräser zeichnen sich durch eine charakteristische Reduzierung der Blüten aus. Die Ährchen werden am Grunde von einer inneren und einer äußeren Hüllspelze (Gluma), die miteinander verwachsen sein können, eingefasst. Oberhalb davon stehen ein oder mehrere Blüten, jede mit einer Deck- sowie Vorspelze. Die Deckspelzen können als Tragblätter der Einzelblüten aufgefasst werden. Die Spelzen variieren in ihrer Form und Größe sehr. Die beiden Hüllspelzen können gleich oder verschieden gestaltet sein. Die Deckspelzen sind vielförmiger gestaltet. Sie können an den Enden spitz, stumpf oder verschiedenartig gezähnt sein. Auf dem Rücken sind sie gerundet, zusammengedrückt oder gekielt. Die Mittelrippe kann in einen Stachel oder eine Granne verlängert sein.
Die Blüten bestehen aus einer Vorspelze und zwei, selten drei, zuweilen an den Rändern verwachsenen Schwellkörperchen (Lodiculae), durch deren Anschwellen die Spelzen geöffnet werden. Es sind ferner meist drei Staubblätter (Stamina) vorhanden (selten sechs, zwei oder nur eines), von denen jedes einen Stiel (Filament) und einen den Pollen tragenden, zweiteiligen Staubbeutel (Anthere) aufweist. In jeder Blüte gibt es schließlich einen runden, aus zwei oder drei Fruchtblättern verwachsenen, oberständigen Fruchtknoten (Ovarium). Dieser verfügt an seiner Spitze über einen Stempel (Pistillum), der seinerseits auf kurzen Stielen ein, zwei oder selten drei fedrige Narbenäste (Stigmae) trägt. Der Fruchtknoten enthält die Samenanlage, die mit Fruchtknotenwänden zu einer Einheit, der Karyopse, verwächst.
Bei manchen Arten enthalten einige Blüten nur männliche Organe oder sind steril. Ferner sind etliche Arten verschiedenährig, das heißt, die Blüten mit nur weiblichen und nur männlichen Organen befinden sich getrennt in verschiedenen Blütenständen desselben Individuums (einhäusig), so beim Mais. Bei anderen Arten wie dem Pampasgras befinden sich die Geschlechter getrennt in den Blütenständen verschiedener Individuen einer Grasart. Sie sind zweihäusig.

Früchte und Samen
Die Frucht ist bei den meisten Grasarten eine trockene Karyopse, eine Sonderform der Nussfrucht. Seltener sind die Früchte Beeren oder Steinfrüchte mit saftigen oder fleischigen Fruchtwänden, so wie bei einigen Bambus-Arten. Während der Reifezeit verwächst die Fruchtwand (Perikarp) mit der Samenschale (Testa) zu einer einsamigen, trockenen Schließfrucht. Die „Samenkörner“ stellen also keine Samen, sondern vielmehr Früchte dar. Unterhalb der Fruchtwand und der Samenschale liegt die eiweißreiche Aleuronschicht. Darunter folgt das den restlichen Samen ausfüllende stärkereiche Nährgewebe, das Endosperm. Gräser sind einkeimblättrig (monokotyl); bei ihnen ist das eine Keimblatt (Kotyledon) zu einem Scutellum (Schildchen) und zu einer Keimscheide (Koleoptile) umgestaltet. Das Scutellum liegt zwischen dem Endosperm und dem Embryo und spielt eine wichtige Rolle für den Stofftransport und die Hormonsynthese. Der Embryo verfügt bereits über deutlich erkennbare Wurzel- und Sprossanlagen. Die Koleoptile ist ein zylinderförmiges Schutzorgan, welches das Primärblatt des auskeimenden Embryos umgibt. Da die Koleoptile ein umgewandeltes Keimblatt darstellt, ist es als Organ ein Blatt. Wie alle Blätter besitzt es zwei Epidermen (außen und innen), Stomata und Leitbündel. Die Stärke und die Proteine dienen dem Embryo als Starthilfe für die Keimung, bevor es sich durch Photosynthese selbst versorgen kann. Die ausgereiften Früchte der Gräser sind in ihrer Gestalt und ihrem Aufbau charakteristisch. Die ehemalige Bauchnaht des Fruchtknotens erscheint auf einer Flanke des Korns als tiefe Furche.
An verschiedenen Stellen des Fruchtstandes bilden sich Zonen eines speziellen Gewebes, entlang dessen ein glatter Bruch entsteht, sobald der Samen reif ist. Bei den meisten Gräsern erfolgt dieser Bruch in der Ährchenachse unterhalb der Deckspelze. Die Karyopse ist in diesen Fällen meistens in Deckspelzen und Vorspelzen fest eingeschlossen und stellt als Gesamtheit die Ausbreitungseinheit (Diaspore) dar. Bei einigen Arten erfolgt der Bruch unterhalb der untersten Deckspelze des Ährchens (z. B. Perlgräser), unter dem einzelnen Ährchen oder in einem Büschel von Ährchen (Gerste), selten in der Hauptachse des Fruchtstandes (Dünnschwanz). Gräser mit nackten Früchten sind in den tropischen Gattungen Sporobolus und Eragrostis häufig. Bei diesen steht das Korn frei und wird ausgestreut, nachdem sich ein Bruch am Grunde der sie haltenden Deckspelze entwickelt hat.
Chemische Merkmale
Die Samen sind reich an Stärke. Diese kann aus einzelnen Stärkekörnern (Roggen, Weizen, Gerste) bestehen oder aus zu mehreren zusammengesetzten (Hafer). Auch in den Rhizomen und anderen vegetativen Organen speichern die Gräser Stärke, Saccharose und/oder Fructane. Bei den Fructanen kommt neben dem unverzweigten „Inulin-Typ“ der verzweigte „Phlein-Typ“ vor. Das Fructanmuster ist wie der Polymerisationsgrad oft kennzeichnend für die Art. Die äußere Endospermschicht (Aleuronschicht) der Karyopsen ist reich an Reserveproteinen. Sie enthält vor allem Albumine, Globuline, Gluteline (nur in verdünnten Säuren und Laugen löslich) und Prolamine (in 70–80%igem Ethanol löslich). Letztere sind beim Roggen- oder Weizenmehl Voraussetzung für die Backfähigkeit.
Einige Triben der Panicoideae bilden ätherische Öle in schlauchförmigen, verkorkten Zellen. Cymbopogon nardus liefert das Aetheroleum Citronella, das hauptsächlich aus Citronellal und Geraniol besteht und bei der Herstellung von Melissengeist oft das echte Melissenöl ersetzt. Weitere Cymbopogon-Arten werden angebaut, da sie Parfümöle wie Palmarosaöl und Lemongrasöl liefern. Diese Öle bestehen überwiegend aus Mono- und Sesqui-Terpenen, während Phenylpropanoide selten sind. Alkaloide sind selten. Es gibt Protoalkaloide und vereinzelt Pyrrolizidin- und β-Carbolintyp-Alkaloide. Cyanogene Glykoside (blausäure-produzierende Verbindungen) sind weit verbreitet, kommen aber immer nur in geringen Mengen vor. Cumarine kommen wahrscheinlich bei allen Vertretern vor, aber nur beim Gewöhnlichen Ruchgras (Anthoxanthum odoratum) und beim Duftenden Mariengras (Hierochloe odorata) in größeren Mengen. Polyphenole sind in geringeren Mengen enthalten.
In den Blattepidermen wird wie bei den Sauergräsern (Cyperaceae) häufig Kieselsäure in Form von Kieselsäurekörpern eingelagert. Oxalatkristalle scheinen vollkommen zu fehlen.
Etliche dieser Inhaltsstoffe zeigen als Bitterstoffe eine fraßhemmende Wirkung oder wirken toxisch auf Bakterien oder Pilze.
Ökologie

.jpg.webp)

Vegetatives Wachstum, Ausbreitung und Regeneration
Bei ausdauernden Arten erfolgt die vegetative Ausbreitung überwiegend über Stolonen und Rhizome, die sich an den Knoten bewurzeln. Etliche Arten bedienen sich zusätzlich der unechten Viviparie, bei der keine Samen gebildet, sondern Brutknospen (Bulbillen), die erbgleiche Tochterpflanzen hervorbringen. Ein bekanntes Beispiel ist das Alpen-Rispengras (Poa alpina). Bei diesem Gras entwickeln sich im Blütenstand anstelle von Blüten grüne Pflänzchen, die an der Mutterpflanze verbleiben oder zu Boden fallen und als Diasporen dienen. Beim Zwiebel-Rispengras (Poa bulbosa) bilden sich basale, zwiebelartige Brutknospen, in denen Reservestoffe eingelagert sind. Jede Brutknospe bildet die Grundlage für eine neue Pflanze.
Gräser sind zur raschen Regeneration nach Verbiss oder Mahd befähigt. Dies liegt in der geschützten Lage ihrer Blattwachstumszonen (Meristeme) und Nebentriebknospen begründet. Die Wachstumszonen befinden sich an der Basis der Graspflanzen nahe der Erdoberfläche. Die Triebe bestehen aus unterschiedlich alten und gegenständig angeordneten Blättern. Junge Blätter wachsen an der Basis der Blattscheide (Interkalarmeristem). Ein erneutes Wachstum der Blätter nach Verlust durch Mahd oder Beweidung wird dadurch ermöglicht. Auch die einzelnen Blätter verfügen wie die Halme am oberen Ende der Blattscheiden im Übergang zu den Blattspreiten über teilungsfähiges Gewebe, das Nebentriebe bilden kann. Ferner sind die Halme durch das unterschiedliche Wachstum der teilungsfähigen Zonen oberhalb der Knoten zu einem Wiederaufrichten des Stängels nach Regen oder Tritt befähigt.
Die beschriebenen Wachstumsbereiche sind in verschiedene Zonen unterteilt: An der Basis findet die Zellteilung und damit eine Zellproduktion statt. Darauf folgt ein Bereich der Zellstreckung. In der folgenden Zone der Zelldifferenzierung erfolgt die Ausbildung der Blattzellen. Die Zellproduktion und Zellstreckung verschieben das ausdifferenzierte Blatt nach oben. Sobald das Blatt aus der Blattscheide ans Licht tritt, ist es photosynthetisch aktiv.[5]
Generative Vermehrung und Ausbreitung
Alle Süßgräser sind windblütig (anemogam). Die Blüten öffnen sich nur wenige Stunden am Tag, um Staubblätter und Narben dem Wind auszusetzen. Eine Selbstbestäubung wird durch die meist frühere Reife der Staubblätter verhindert (Proterandrie). Die starke Reduzierung der Blüten ist eine Anpassung an diese Form der Bestäubung. Gräser können auf auffällige Formen und Farben der Blüten und auf ein Nektarangebot zur Anlockung von Tieren verzichten. Die passive Pollenübertragung über den Wind und Luftströmungen erfolgt dabei weit weniger gezielt als bei der Tierbestäubung. Diesen Mangel gleichen die Windblüher mit der Massenproduktion von Blütenstaub aus. Dies führt während der Blütezeit zu regelrechten Staubwolken, die garantieren, dass zumindest ein kleiner Teil des weniger als einen Tag lebensfähigen Pollens seinen Bestimmungsort, die weiblichen Narben, erreicht. Beispielsweise bildet der Roggen (Secale cereale) pro Ähre etwa vier Millionen Pollenkörner; eine einzelne Blüte bis zu 57.000.[6] Eine große Blütenhülle wäre bei der Pollenverbreitung nur hinderlich. Die Lodiculae schwellen durch Wasseraufnahme an und drängen die Spelzen auseinander – die „Grasblüte“ öffnet sich. Die Filamente sind lang und dünn und lassen die Staubbeutel frei aus der Blüte heraushängen. So kann der Wind ungehindert den trockenen, nicht verklebten und leichten Pollen heraustragen. Die Fruchtknoten haben gefiederte und dadurch mit großer Oberfläche versehene Narben, die den Pollen gewissermaßen wieder aus Luft herauskämmen können. Die Effizienz dieser Form der Pollenverbreitung wird durch das Herausheben der Blütenstände über die Ebene des Blattwerkes sowie durch eine hohe Individuendichte der Graspflanzen verstärkt. Eine Sonderform der geschlechtlichen Ausbreitung ist die echte Viviparie, bei der die Samen schon auf der Mutterpflanze auskeimen. Die Samenausbreitung erfolgt auf vielfältige Weise; überwiegend durch den Wind (Anemochorie), über das Wasser (Hydrochorie) oder durch Tiere (Zoochorie).
Mykorrhiza
Das Wurzelsystem der Wiesengräser bildet arbuskuläre Mykorrhiza (AM), eine Symbiose mit einem Pilz. Diese erleichtert der Graspflanze die Erschließung und Aufnahme von Nährstoffen aus dem Boden. Ein Pilzmycel verbindet mehrere Pflanzen derselben Art und andere Pflanzenarten, wodurch nicht nur das Gras selbst und der Pilz, sondern schließlich Wiese und Pilz eine Lebensgemeinschaft bilden.[7]
Photosynthese
Unter den Gräsern gibt es sowohl C3- (die meisten heimischen Gräser wie Deutsches Weidelgras) als auch C4-Pflanzen (z. B. Mais, Hirse und Zuckerrohr), letztere mit effizienterer Photosynthese bei hohem Wärme- und Lichtangebot. C3-Pflanzen weisen dagegen bei kühleren Temperaturen und weniger Licht eine effizientere Photosynthese auf. Die Forschung hat gezeigt, dass der C4-Mechanismus zuerst bei den Gramineen, wahrscheinlich im Oligozän vor etwa 23 bis 34 Millionen Jahren entwickelt wurde, wobei es Hinweise auf über zehn unabhängige Entwicklungen gibt. Bei der geographischen Verbreitung ergibt sich eine auffällige klimatische Abhängigkeit der Photosynthesetypen. So ist der Anteil der C4-Pflanzen unter den Gräsern in kühlen und humiden Klimaten deutlich niedriger als in trockenen bis extrem ariden Regionen der Erde.[7]










Synökologie, Biotope und Bedeutung
Natürliche und anthropogene Grasländer
Etwa ein Fünftel der Pflanzendecke der Erde wird von Gräsern eingenommen.[8] Savannen und Steppen bilden die großen, natürlichen Grasländer der Erde in Klimazonen, die für Wald nicht geeignet sind. Dem gegenüber stehen die durch menschliche Tätigkeit entstandenen Kulturgrasländer vor allem Mitteleuropas, die in einem langen nacheiszeitlichen Prozess vom Wald zur offenen, durch Wiesen und Weiden geprägten Landschaft entstanden.
Die dauerhaften, mehr oder weniger geschlossenen Grasbestände erfüllen vielfältige ökologische und biologische Aufgaben. Sie verhindern vor allem durch ihr dichtes und eng vernetztes Wurzelsystem die Abtragung der Bodenschicht durch Wind und Wasser (Erosion). Ferner erzeugen sie durch ihr Wurzelwerk einen hohen Gehalt an organischer Substanz im Boden. Etwa zwei Drittel der pflanzlichen Primärproduktion bleiben in Grasländern unterirdisch zurück und führen so zur Humusbildung. Dazu trägt das jährlich absterbende oberirdische Pflanzenmaterial zusätzlich bei, das als Mulch zurückbleibt und nur langsam zersetzt wird. In vielen Grasländern spielen natürliche Feuer eine Rolle. Blitze entzünden am Ende der Vegetationszeit die abgestorbene Pflanzenmasse. Die in der Asche enthaltenen anorganischen Nährstoffe fördern als Dünger den Neuaustrieb der Pflanzen. Darüber hinaus wird der Gehölzaufwuchs zerstört, die Brände tragen so zur Offenhaltung der Graslandschaft bei. Grasländer beherbergen und ernähren eine artenreiche und vielfältige Tierwelt: eine Vielzahl von Insekten (Termiten und Ameisen), Spinnen, Vögel, Kleinsäuger und zahlreiche im Boden lebende Tiere, nicht zuletzt Großsäuger wie jene der großen Tierherden in den afrikanischen Savannen. Letztere tragen wie die regelmäßigen Feuer dazu bei, die Verjüngung der Gehölze zu hemmen. Ihre Exkremente düngen den Boden. Durch den Fraß wird die Regeneration der Gräser so angeregt, dass die Primärproduktion um mehr als zwei Drittel zunimmt. Ferner sind sie für die Verbreitung der Früchte und Samen durch Epi-, Endo- oder Dysochorie von Bedeutung. Naturnahe Grasländer gehen heute weltweit zurück. Die Ursachen liegen in der Umwandlung in Acker- und Siedlungsland, der Aufgabe traditioneller Wiesen- oder Weidenutzungen sowie in der Intensivierung (Düngung) und Degradierung (Überweidung).[9]
Steppen und Prärien
Die baumfreien Steppen finden sich in den semiariden, gemäßigten Zonen vorwiegend auf der Nordhalbkugel. Auf der Südhalbkugel ist die argentinische Pampa eine zu den eurasischen Steppen und den nordamerikanischen Prärien analoge Vegetationsform. Es wird kontrovers diskutiert, ob sie auf natürliche Weise entstanden ist. Steppen unterliegen durch strenge Kälte im Winter und anhaltende Trockenheit im Sommer im Jahresverlauf zwei Perioden der Vegetationsruhe. Die im Frühjahr, Frühsommer und Spätherbst anfallenden Niederschläge reichen für das Wachstum der Steppenvegetation aus. Kennzeichnend für Steppen ist ihre hohe bodenbiologische Aktivität bei einem hohen Humusanteil (bis zu 10 %). Es können sich fruchtbare Schwarzerdeböden mit Humushorizonten bis zu einem Meter Mächtigkeit bilden. Die osteuropäischen Steppen lassen sich grob in etwa vier Vegetationstypen gliedern, die der zunehmenden Kontinentalität in Richtung Südost folgen. In Russland und der Ukraine finden sich Wiesensteppen mit Aufrechter Trespe (Bromus erectus), Flaumigem Wiesenhafer (Avenula pubescens), Schillergräsern (Koeleria) und vielen anderen Grasarten. Sie sind reich an einjährigen, und nicht grasartigen ausdauernde krautige Pflanzen. Darauf folgt die durch Pfriemengräser der Gattung Stipa dominierte Federgrassteppe mit schmalblättrigen „Horstgräsern“ und weniger Stauden, und schließlich die Kurzgrassteppe mit xerophytischen Festuca-Arten. Westliche Vorposten der osteuropäischen Steppen finden sich beispielsweise in der Pannonischen Tiefebene Ungarns. Im gemäßigten Nordamerika entspricht den Steppen die flächenmäßig deutlich kleinere Prärie zwischen Mississippi und den Rocky Mountains. Sie ist im Gegensatz zu den osteuropäischen Steppen weniger kontinental geprägt. Sie erreicht in West-Ost-Ausdehnung 1000 Kilometer, in Nord-Süd-Ausdehnung 2750 Kilometer. Im Osten findet sich die Hochgras-Prärie mit Wiesen-Rispengras (Poa pratensis), dem Pyramiden-Schillergras (Koeleria pyramidata), Prärie-Bartgras (Andropogon scoparius), Rutenhirse (Panicum virgatum) und vielen krautigen Pflanzen. Im Südteil folgt Richtung Westen die Mischgras-Prärie im Übergang zur am Fuße der Rocky-Mountains befindlichen Kurzgras-Prärie mit dem Moskitogras (Bouteloua gracilis) und Buchloe dactyloides. Die Bedeutung geschlossener Grasdecken als Schutz vor Erosion zeigen die verheerenden Sandstürme der 1930er Jahre in der danach benannten „Dust Bowl“ Nordamerikas durch großflächige Bodenzerstörungen im Zuge der Umwandlung in Ackerland. Noch im 17. und 18. Jahrhundert zogen Büffelherden mit einer geschätzten Bestandsgröße von 50 bis 70 Millionen Tieren über die Prärien. Heute sind es über 100 Millionen Hausrinder.[9][10]
Savannen
Die Savannen umfassen etwa 15 Millionen Quadratkilometer. In den wechselfeuchten Sommerregengebieten der Tropen der Südhalbkugel erreichen sie ihre größte Ausdehnung in Afrika. Analoge Formationen der Savannen sind die Llanos Venezuelas und Kolumbiens, die Cerrados Brasiliens sowie die Eukalyptus-Steppen Nordaustraliens. Savannen sind im Gegensatz zu den Steppen und Prärien mit Bäumen und Sträuchern durchsetzt. Die Savannen Afrikas würden ohne Feuer und den Einfluss der großen Elefanten- und Huftierherden in kurzer Zeit mit Gehölzen zuwachsen. In den Savannen findet sich häufig ein kleinräumiges Oberflächenrelief aus flachen Hügeln und Senken mit Höhenunterschieden unter einem Meter. Dadurch unterscheiden sich die Standorte vor allem hinsichtlich der Wasserverfügbarkeit. Die unterschiedliche Wasserverfügbarkeit bestimmt schließlich die Nährstoffverfügbarkeit und die Vegetation. In Savannen spielen neben den Großtieren Termiten, Ameisen und Heuschrecken eine maßgebliche Rolle als Regulative im Ökosystem. Der Artenreichtum der Pflanzen der Savannen ist vergleichsweise gering. Die Hauptkomponenten sind C4-Gräser wie die Lampenputzergräser (Pennisetum) und Andropogon-Arten in Afrika. In Australien sind die Savannen, das sogenannte Spinifex- oder Hummock-Grasland, durch Igelkopfgräser der Gattungen Triodia und Plectrachne gekennzeichnet. Dagegen ist der Artenreichtum der Tiere ausgesprochen groß. So leben etwa 1,5 Millionen Großtiere in den Savannen der Erde, allein in der Serengeti Ostafrikas sind es 98 große Weidetiere pro Quadratkilometer. Die Zoomasse wird auf 150 bis 250 Kilogramm Trockenmasse pro Hektar geschätzt – jene der Wälder der gemäßigten Zone wird mit nur 10 Kilogramm Trockenmasse pro Hektar angegeben.[9][10]
Kulturgrasland
In der Naturlandschaft Europas ist natürliches Grasland auf wenige Bereiche beschränkt. Nur in hohen Berglagen oberhalb der Waldgrenze, in Seemarschen, in oft überschwemmten Auenbereichen und im Randbereich von Hochmooren konnten sich kleinräumig natürliche, weitgehend baumfreie Grasländer, sogenannte Urwiesen, entwickeln. Sie sind heute stark vom Menschen überprägt. Kulturgrasland dient dem Menschen wirtschaftlich als Grundlage der Viehzucht. Die durch Gräser dominierten Wiesen- und Weidelandschaften des gemäßigten Europa sind im Wesentlichen das Ergebnis jahrhundertelangen menschlichen Wirkens. Noch vor etwa 10.000 Jahren war Mitteleuropa nahezu reines Waldland. Die Entwicklung bäuerlicher Kulturen, die sich vom Nahen Osten ausgehend vor etwa 6700 bis 6400 Jahren (Neolithikum) nach Mitteleuropa ausbreiteten, ermöglichte das Sesshaftwerden der Menschen und führte zu immer stärkeren Eingriffen in die natürliche Pflanzendecke. Es gab Siedlungen, erste Äcker und Nutztiere, die ihre erste Nahrung im Wald suchten. Der Fraß der Tiere, Brand und Holzeinschlag führten im Laufe längerer Zeit zu Auflichtungen in den Wäldern. Mit Beginn der Eisenzeit wurde die Landnutzung verstärkt, und mit Erfindung der Sense wurde die Gewinnung von Heu und Streu möglich. Auf diese Weise entstanden erste größere Wiesenareale. Im Mittelalter vollendete sich die Landschaftsentwicklung in einer offenen und differenzierten Kulturlandschaft aus Siedlungen, Waldresten, Feldgehölzen, Gebüschen, Äckern sowie artenreichen Wiesen und Weiden. Die typische Landschaft wird in der Literatur vielfach als „Parklandschaft“ oder „europäische Savanne“ beschrieben. Im Zuge der Agrarentwicklung in der Neuzeit wird die Landwirtschaft durch die Technisierung, Flurbereinigungen, Melioration sowie gezielte Ansaat ausgewählter Grasarten immer unabhängiger von natürlichen Gegebenheiten. Die artenreichen, extensiven Wiesen und Weiden wurden weitgehend von artenarmen, monotonen Wirtschaftswiesen und -weiden abgelöst. Zu den wichtigsten angebauten Futtergräsern des Dauergrünlandes gehören heute das Deutsche Weidelgras (Lolium perenne), das Wiesen-Lieschgras (Phleum pratense), das Gewöhnliche Knäuelgras (Dactylis glomerata) sowie Wiesen- und Rohr-Schwingel (Festuca pratensis, Festuca arundinacea).[9][11]
Küsten- und Hochwasserschutz
Etliche Gräser verhindern durch ihr dünnes Netzwerk aus Wurzeln und die Bedeckung des Bodens mit ihren oberirdischen Pflanzenteilen nicht nur dessen Abtragung durch Wind und Wasser, sondern sorgen zusätzlich für dessen Aufhöhung. Dafür sind besonders solche Arten geeignet, die unter vergleichsweise ungünstigen Standortbedingungen weitreichende Rhizome und Stolonen bilden können. So ist beispielsweise der Gewöhnliche Strandhafer (Ammophila arenaria) maßgeblich an der Festlegung der Treibsande sowie am Aufbau der Weißdünen auf den Inseln und an den Festlandsküsten beteiligt und erfüllt so eine wichtige Funktion im Küstenschutz. Auf regelmäßig überfluteten Schlickflächen der Küsten ist es der Strand-Salzschwaden (Puccinellia maritima), der mit seinen sich bewurzelnden Stolonen allmählich die kurzen dichten Rasen der Salzwiesen bildet und mit seinen kurzen steifen Halmen und Blättern den Schlamm gewissermaßen einfängt und die Oberfläche der Marsch langsam aufhöht. Dort, wo andere Süßgräser nicht mehr gedeihen können, übernimmt das Salz-Schlickgras (Spartina anglica) an ähnlichen Standorten die Funktion der Festlegung und Aufhöhung von Schlick der seewärtigen Seiten im Wattenmeer und entlang der Priele. Die Deiche der Küsten und Ströme werden schließlich mit einer Pflanzendecke ausgestattet, die von bodenhaltenden Süßgräsern dominiert wird.
In den Ebenen entlang der Flussufer des Binnenlands schützen Gräser den Boden vor Erosion und erfüllen eine ebenso wichtige Aufgabe im Hochwasserschutz. Beispielsweise bilden das Rohr-Glanzgras (Phalaris arundinacea) und der Wasser-Schwaden (Glyceria maxima) auf Schlammflächen und an Ufern dichte und hohe Aufwüchse mit kräftigen Rhizomen. Flussauen stellen nicht nur Retentionsflächen für Hochwässer dar, sondern sind aufgrund der Großgräser eine Senke („Falle“) für Sedimente, Nähr- und Schadstoffe.
Nutzung und Bedeutung für den Menschen
Die Familie der Süßgräser bietet ein breites Spektrum an Nutzungsmöglichkeiten. Demgegenüber stehen jedoch nur relativ wenige Gattungen, die schließlich als Nutzpflanzen für den Menschen von Bedeutung sind. So werden lediglich 6 bis 7 Prozent der 600 bis 700 Gattungen als Nahrung oder als Werk- und Baustoffe verwendet. Nur etwa 15 Gattungen, das sind knapp 2 Prozent (ohne Berücksichtigung der Bambus-Gattungen), spielen dabei eine größere Rolle.[12]
Getreide



Von großer weltwirtschaftlicher Bedeutung sind die Getreide. Grasfrüchte, beziehungsweise Getreidekörner, dienen dem Menschen als Grundnahrungsmittel. Sie liefern über 50 % der Welternährungsenergie. Weizen (Triticum-Arten), Mais (Zea mays) und Reis (Oryza sativa) nehmen dabei eine führende Rolle ein. Gerste, Roggen, Hirsen und Hafer decken etwa ein Zehntel ab. Weizen, Gerste und Roggen haben ihren Ursprung im sogenannten Fruchtbaren Halbmond, der sich von Ägypten über Palästina bis zum Persischen Golf erstreckt. Hier wurden die Wildformen in Kultur genommen, die über verschiedene Auslese- und Kreuzungsprozesse zu den heutigen Kulturformen entwickelt wurden. Reis hat seinen Ursprung in China oder Indien; Mais stammt aus Mexiko. Unter Hirsen werden Gräser verschiedener Gattungen mit kleinfrüchtigen Körnern zusammengefasst, wie Digitaria, Echinochloa, Eragrostis, Panicum, Setaria, Sorghum.[12]
Im Jahr 2019 wurden weltweit folgende Anbauergebnisse erzielt (Tabelle nach Erntemengen weltweit sortiert):
| 2019 | Anbaufläche in Hektar | Ertrag in dt pro ha | Getreidemenge in Tonnen | ||||
|---|---|---|---|---|---|---|---|
| Getreideart | Welt | D | Welt | D | Welt | D | |
| 1 | Weizen | 215.901.958 | 3.118.100 | 35,5 | 74,0 | 765.769.635 | 3.344.000 |
| 2 | Mais | 197.204.250 | 416.000 | 58,2 | 88,1 | 1.148.487.291 | – |
| 3 | Reis | 162.055.938 | – | 46,6 | – | 755.473.800 | - |
| 4 | Gerste | 51.149.869 | 1.708.800 | 31,1 | 67,8 | 158.979.610 | 9.583.600 |
| 5 | Hirse* | 71.728.545 | – | 11,7 | – | 86.265.170 | – |
| 6 | Hafer | 9.418.493 | 126.300 | 24,5 | 41,1 | 23.104.147 | 577.600 |
| 7 | Roggen | 4.213.392 | 636.300 | 30,4 | 50,9 | 12.801.441 | 2.201.400 |
| Summe | 711.672.445 | 6.005.500 | 2.950.881.094 | 42.075.800 | |||
| * Hirse umfasst hier Sorghum und Millet („Echte Hirse“) Quelle: FAO: Faostat 2019[13] | |||||||
Sonstige Nutzungen
Neben den Grasfrüchten werden die Stängel, die Blätter und Wurzeln genutzt. Süßgräser sind eine wichtige Rohstoffquelle zur Gewinnung von Stärke, Zellulose, Zucker sowie Fetten und ätherischen Ölen. Sie können als Werk-, Bau- und Füllstoffe verwendet werden. Vor allem werden die verholzten Halme verschiedener Bambus-Arten in tropischen und subtropischen Gebieten Asiens zur Herstellung von Möbeln, Ess- und Trinkgefäßen oder Zäunen verwendet und nicht zuletzt im Haus- und Gerüstbau eingesetzt. In Nordwesteuropa wird das hier im Überfluss wachsende Schilf zu Eindeckung von Häusern verwendet.
Die Eismumie Ötzi soll einen Mantel aus Süßgräsern getragen haben.

Bambussprosse werden als Gemüse gegessen.

Zitronengräser (Cympopogon) werden als Gewürz- und Heilpflanzen verwendet. Ferner dienen Gräser zur Herstellung von alkoholischen Getränken wie Bier, Rum oder Korn. Als nachwachsende Rohstoffe gewinnen Süßgräser, vor allem Bambus und Zuckerrohr, zunehmende Bedeutung zur Herstellung von Bioalkohol als Treibstoff.
Schließlich sei noch die Verwendung zahlreicher Süßgrasarten mit auffälligen Blütenständen, wie beispielsweise das Pampasgras, als Ziergräser im Garten- und Landschaftsbau genannt.[12]
Die landwirtschaftliche Nutzung umfasst neben dem Getreideanbau die Nutzung zahlreicher Grasarten als Futterpflanzen für Rinder, Schafe oder Pferde in Form von Kulturgrasland wie Wiesen (Mahd zur Heugewinnung, Streunutzung, Silage) oder Weiden. Darüber hinaus werden geeignete Gräser für Rasen in privaten Gärten, in Parks, auf Golf- oder Sportplätzen eingesetzt, mit der Nutzungsart und -intensität angepassten Sortenmischungen.
Gesundheit
Bei empfindlichen Menschen können Pollen von Süßgräsern die Bildung von Antikörpern Immunglobulin E (IgE) auslösen, was als Heuschnupfen bekannt ist. Sogenannte wasserlösliche I-Glykoproteine haften an der Pollenoberfläche, werden leicht an die Schleimhäute abgegeben und können allergische Reaktionen erzeugen. Ferner können in der Aleuronschicht der Getreidekörner enthaltene Prolamine durch eine immunologische Überempfindlichkeitsreaktion die als Zöliakie bezeichnete Krankheit auslösen.
Vorkommen
Süßgräser sind weltweit verbreitet. Sie kommen von den Meeresküsten bis ins Hochgebirge, vom Äquator bis jenseits der Polarkreise in nahezu allen terrestrischen Ökosystemen vor und besiedeln dabei Standorte von großer ökologischer Bandbreite. Sie wachsen sowohl auf dauernassen bis extrem trockenen Böden als auch in sehr heißen bis arktisch kalten Klimaten.
Man findet Süßgräser flutend in Gewässern, bestandsbildend als Röhrichte, als Unterwuchs in Wäldern, auf wechselfeuchten wie auch trockenen Böden, an Straßenrändern, an Böschungen, auf Felsen – selbst Schotterflächen und Mauerkronen werden besiedelt. Die Familie der Süßgräser deckt nahezu alle denkbaren Standorttypen ab, wobei die einzelnen Arten und Populationen im Rahmen des Wettbewerbs um die Ressourcen (Konkurrenz) ihre jeweils eigenen Vorzugs- oder Existenzbereiche besiedeln. Etliche Pflanzenformationen außerhalb der Wälder werden im Wesentlichen durch Gräser aufgebaut. Die nordamerikanische Prärie, die Steppen Osteuropas, die Savannen Afrikas und die Pampa Südamerikas, aber auch die Wirtschaftswiesen und -weiden Europas sind die landschaftsprägenden natürlichen sowie unter menschlichen Einfluss entstandenen Grasländer der Erde, in denen Bäume und Sträucher zurücktreten oder ganz fehlen.
Stammesgeschichte
In der Erdneuzeit (Känozoikum) entstanden die modernen Familien der Blütenpflanzen, so auch die Gräser. Sie waren zunächst auf bewaldete und sumpfige Gebiete beschränkt. Mit der Entwicklung des kontinuierlichen Wachstumsprozesses und der Windbestäubung wurden ab dem Oligozän die offenen Länder erobert. Steppen und Grasländer breiteten sich vor allem im Miozän aus. Man nimmt an, dass die Evolution der Süßgräser mit jener der großen Weidetiere (Wiederkäuer, Pferde, Kamele etc.) parallel ging.[10]
Erkenntnisse indischer Wissenschaftler aus dem Jahr 2005 gehen einem Bericht der Zeitschrift Science zufolge davon aus, dass sich Gräser bereits in der Kreidezeit, dem letzten Abschnitt des Erdmittelalters (Mesozoikum), entwickelt haben. Diese Annahme geht auf Funde von Pflanzenbestandteilen zurück, die im fossilen Dung (Koprolith) von Dinosauriern gefunden wurden und auf reis- und bambusähnliche Gräser deuten.[14]
Süßgräser gehören zu den im Verlauf der Evolution sekundär entstandenen windblütigen Angiospermen. Spuren von Pollenkitt in Gräsern weisen darauf hin, dass die Vorgänger biotisch durch Vögel und Insekten bestäubt wurden. Pollenkitt verklebt die Pollenkörner zu größeren Übertragungseinheiten, was bei der Windbestäubung, die schwebfähige und leichte Pollen verlangt, störend wäre.[15]
Im Zuge des Übergangs zur Windblütigkeit wurden die Blüten reduziert. Entwicklungsgenetische Befunde deuten darauf hin, dass die Vorspelzen ein Verwachsungsprodukt zweier Blütenhüllblätter von ursprünglich drei und die Schwellkörperchen aus inneren Tepalen hervorgegangen sind. Die Gräserblüte lässt sich somit vom Grundtypus der dreizähligen Blüten einkeimblättriger Pflanzen ableiten mit zwei Kreisen à drei Blütenhüllblättern, zwei Kreisen à drei Staubblättern sowie drei Fruchtblättern. Der dreifächrige Fruchtknoten der Süßgräser wurde einfächrig und enthält nur noch eine Samenanlage. Vom äußeren und inneren Staubblattkreis blieb nur der äußere Ring erhalten. Vom inneren Hüllblattkreis blieben nur die zwei als Schwellkörperchen dienenden Schuppen, die dritte Tepale fiel aus. Nur bei einigen tropischen Arten sind noch drei Lodiculae vorhanden. Der äußere Hüllblattkreis besteht nur noch aus der Vorspelze, die aus zwei getrennten Blütenhüllblättern entstanden ist. Bei wenigen tropischen Gräsern sind zwei getrennte Vorspelzen erhalten. Die dritte Tepale fiel wiederum aus.
Etwa 80 % der Grasarten haben mehr als einen Chromosomensatz im Zellkern. Hybride, zum Teil auch fruchtbare, sind bei Süßgräsern selbst zwischen Gattungen nicht selten.[7] Viele der heutigen Gräser waren in der Naturlandschaft vermutlich nicht in der jetzigen Form vorhanden. Es wird angenommen, dass viele Graslandarten ihren Ursprung in diploiden Sippen haben, die während der Eiszeiten in südlichen Rückzugsgebieten überdauerten. In einem langen Prozess der Rückwanderung, der Anpassung an veränderte Standortbedingungen und verschärfter Konkurrenz sowie durch die vom Menschen seit dem Neolithikum neu geschaffenen Lebensräume konnten durch Kreuzungen diploider Elternarten tetra-, bis polyploide Sippen (Allopolyploidie) entstehen. So sind zum Beispiel Anthoxanthum odoratum, Agrostis stolonifera, Dactylis glomerata und Poa pratensis Hybride alter diploider Sippen.[11]
Systematik
Die Typusgattung der Familie der Poaceae ist Poa. Der Gattungsname Poa ist vom griechischen Wort póa abgeleitet und bedeutet Kraut, Gras, Pflanze. Die veraltete Bezeichnung für die Familie lautet Gramineae. Nach dem Internationalen Code der Botanischen Nomenklatur Artikel 14 ist die weitere Verwendung des alten Begriffes als Ausnahme von den strengen Regeln erlaubt und damit legitim: Poaceae = Gramineae Jussieu nom. cons. (nomina conservanda) et nom. alt. (nomen alternativum).[16][17][18]
Süßgräser sind Bedecktsamer (Magnoliopsida). Im Gegensatz zu den Nacktsamern (Gymnospermae) ist bei ihnen die Samenanlage im Fruchtknoten eingeschlossen. Die Familie der Süßgräser gehört innerhalb der Einkeimblättrigen Pflanzen (Monokotyledonen) zur Ordnung der Süßgrasartigen (Poales). Die Familie umfasst etwa 10.000 Arten mit je nach verwendeter Systematik 600 bis 700 Gattungen. Die Poaceae sind in 13 Unterfamilien von ungleicher Größe unterteilt, die noch weiter in insgesamt 46 Tribus gegliedert sind. Die Unterfamilien können vom phylogenetischen Standpunkt aus zu zwei Hauptgruppen, „BEP-clade“ und „PACC-clade“ zusammengefasst werden.
.jpg.webp) Unterfamilie Pharoideae: Pharus latifolius
BEP-clade
 Unterfamilie Ehrhartoideae: Reis (Oryza sativa)  Unterfamilie Aristidoideae: Aristida acuta  Unterfamilie Pooideae: Weizen (Triticum aestivum)
PACC-clade
 Unterfamilie Danthonioideae: Habitus, Laubblätter und Blütenstand von Cortaderia jubata
 Unterfamilie Arundinoideae: Pfahlrohr (Arundo donax)  Unterfamilie Chloridoideae: Eragrostis cilianensis  Unterfamilie Panicoideae: Sorghumhirse (Sorghum bicolor)
|
Quellen
Die allgemeinen Informationen dieses Artikels entstammen den unter Literatur und Weblinks aufgeführten Referenzen (Morphologie, Standorte, Verbreitung etc.). Darüber hinaus sind einzelne Aspekte, Spezialthemen, Zahlen usw. den aufgeführten Einzelpublikationen entnommen.
Literatur
- Charles Edward Hubbard: Gräser. Beschreibung, Verbreitung, Verwendung (= UTB. Band 233). 2., überarbeitete und ergänzte Auflage. Eugen Ulmer, Stuttgart (Hohenheim) 1985, ISBN 3-8001-2537-4 (englisch: Grasses. Übersetzt von Peter Boeker).
- Peter Sitte, Elmar Weiler, Joachim W. Kadereit, Andreas Bresinsky, Christian Körner: Lehrbuch der Botanik für Hochschulen. Begründet von Eduard Strasburger. 35. Auflage. Spektrum Akademischer Verlag, Heidelberg 2002, ISBN 3-8274-1010-X.
- Ernst Klapp, Wilhelm Opitz von Boberfeld: Taschenbuch der Gräser. Erkennung und Bestimmung, Standort und Vergesellschaftung, Bewertung und Verwendung. 13. überarbeitete Auflage. Eugen Ulmer, Stuttgart (Hohenheim) 2006, ISBN 3-8001-4775-0.
- Wu Zheng-yi, Peter H. Raven, Deyuan Hong (Hrsg.): Flora of China. Volume 22: Poaceae. Science Press / Missouri Botanical Garden Press, Beijing / St. Louis 2006, ISBN 1-930723-50-4 (englisch).
- Vernon Hilton Heywood (Hrsg.): Blütenpflanzen der Welt. Birkhäuser, Basel / Bonn / Stuttgart 1982, ISBN 3-7643-1305-6 (englisch: Flowering Plants of the World.).
Weblinks
- R. J. Soreng, G. Davidse, P. M. Peterson, F. O. Zuloaga, E. J. Judziewicz, T. S. Filgueiras, O. Morrone, K. Romaschenko: A World-wide Phylogenetic Classification of Poaceae (Gramineae). Die erste Veröffentlichung erfolgte am 13. Januar 2000 als Classification of New World Grasses (Poaceae/Gramineae). Die Daten werden fortlaufend aktualisiert, zuletzt am 5. April 2014.
- Die Familie der Poaceae bei der APWebsite. (Abschnitt Systematik)
- Gramineae Juss. bei L. Watson und M. J. Dallwitz: The Families of Flowering Plants, Beschreibung im DELTA-Format. (Abschnitt Beschreibung)
Einzelnachweise
- M.J.M. Christenhusz, J.W. Byng: The number of known plants species in the world and its annual increase. Phytotaxa. Volume 261, No. 3, 2016, S. 201–217. doi:10.11646/phytotaxa.261.3.1
- Vernon Hilton Heywood (Hrsg.): Blütenpflanzen der Welt. Birkhäuser, Basel / Bonn / Stuttgart 1982, ISBN 3-7643-1305-6 (englisch: Flowering Plants of the World.).
- Gerhard Grümmer: Das Verhalten von Rhizomen der Quecke (Agropyron repens) gegen trockene Luft, Weed Research 3 (1), 1963, S. 44–51.
- Friedrich Kluge, Alfred Götze: Etymologisches Wörterbuch der deutschen Sprache. 20. Auflage, hrsg. von Walther Mitzka, De Gruyter, Berlin/ New York 1967; Neudruck („21. unveränderte Auflage“) ebenda 1975, ISBN 3-11-005709-3, S. 285.
- H. Schnyder: Physiologische und morphogenetische Grundlagen zum Regenerationsvermögen der Gräser In: Bayerische Akademie der Wissenschaften (Hrsg.): Gräser und Grasland: Biologie – Nutzung – Entwicklung, Rundgespräch am 10. Oktober 2005, Verlag Dr. Friedrich Pfeil, München 2006, ISBN 3-89937-070-8, ISSN 0938-5851.
- H. Behrendt, J. Ring: A research strategy for the investigation of the influence of environmental pollutants on the development of allergic sensitization and disease. In: J. Ring, H. Behrendt, D. Vieluf (Hrsg.): New Trends in Allergy IV. Springer, Berlin / Heidelberg 1997, S. 51–60.
- H. Ziegler: Die Familie der Süßgräser. In: Bayerische Akademie der Wissenschaften (Hrsg.): Gräser und Grasland: Biologie – Nutzung – Entwicklung, Rundgespräch am 10. Oktober 2005, Verlag Dr. Friedrich Pfeil, München, ISBN 3-89937-070-8, ISSN 0938-5851.
- H. L. Shantz: The Place of Grasslands in the Earth’s Cover. In: Ecology, Volume 35, No. 2, 1954, S. 143–145.
- W. Haber: Die Grasländer der Erde: Verbreitung und Lebensbedingungen. In: Bayerische Akademie der Wissenschaften (Hrsg.): Gräser und Grasland: Biologie – Nutzung – Entwicklung, Rundgespräch am 10. Oktober 2005, Verlag Dr. Friedrich Pfeil, München, ISBN 3-89937-070-8, ISSN 0938-5851.
- J. H. Reichholf: Gräser, Fleisch und Humus. Tiere in Graslandökosystemen In: Bayerische Akademie der Wissenschaften (Hrsg.): Gräser und Grasland: Biologie – Nutzung – Entwicklung, Rundgespräch am 10. Oktober 2005, Verlag Dr. Friedrich Pfeil, München, ISBN 3-89937-070-8, ISSN 0938-5851.
- H. Dierschke, G. Briemle: Kulturgrasland. Ulmer, Stuttgart 2002, ISBN 3-8001-3816-6.
- E. Bayer: Bedeutende und interessante Nutzpflanzen aus der Familie der Gräser. In: Bayerische Akademie der Wissenschaften (Hrsg.): Gräser und Grasland: Biologie – Nutzung – Entwicklung. Rundgespräch am 10. Oktober 2005, Verlag Dr. Friedrich Pfeil, München 2006, ISSN 0938-5851, ISBN 3-89937-070-8.
- Ernährungs- und Landwirtschaftsorganisation der Vereinten Nationen (FAO): Produktionsstatistik 2019, abgerufen am 7. Juni 2021 (englisch).
- Vandana Prasad, Caroline A. E. Strömberg, Habib Alimohammadian, Ashok Sahni: Dinosaur Coprolites and the Early Evolution of Grasses and Grazers. In: Science, Volume 310 (5751), 2005, S. 1177.
- Michael Hesse: Entwicklungsgeschichte und Ultrastruktur von Pollenkitt und Exine bei nahe verwandten entomophilen und anemophilen Angiospermensippen der Alismataceae, Liliaceae, Juncaceae, Cyperaceae, Poaceae und Araceae, Plant Systematics and Evolution, Springer Wien 1980.
- Internationaler Code der Botanischen Nomenklatur (PDF; 1,5 MB) abgerufen 7. September 2007.
- Poaceae bei Tropicos.org. Missouri Botanical Garden, St. Louis, abgerufen am 9. Dezember 2014.
- Poaceae im Germplasm Resources Information Network (GRIN), USDA, ARS, National Genetic Resources Program. National Germplasm Resources Laboratory, Beltsville, Maryland. Abgerufen am 9. Dezember 2014.
- Grass Phylogeny Working Group, Nigel P. Barker, Lynn G. Clark, Jerrold I. Davis, Melvin R. Duvall, Gerald F. Guala, Catherine Hsiao, Elizabeth A. Kellogg, H. Peter Linder: Phylogeny and Subfamilial Classification of the Grasses (Poaceae). Annals of the Missouri Botanical Garden, 2001, Volume 88, No. 3, 2001, S. 373–457 (Online. (Memento des vom 25. November 2015 im Internet Archive) Info: Der Archivlink wurde automatisch eingesetzt und noch nicht geprüft. Bitte prüfe Original- und Archivlink gemäß Anleitung und entferne dann diesen Hinweis.)
- Rafaël Govaerts (Hrsg.): Poaceae – Datenblatt bei World Checklist of Selected Plant Families des Board of Trustees of the Royal Botanic Gardens, Kew. Abgerufen am 9. Dezember 2014.
- R. J. Soreng, G. Davidse, P. M. Peterson, F. O. Zuloaga, E. J. Judziewicz, T. S. Filgueiras, O. Morrone, K. Romaschenko: A World-wide Phylogenetic Classification of Poaceae (Gramineae). Die erste Veröffentlichung erfolgte am 13. Januar 2000 als Classification of New World Grasses (Poaceae/Gramineae). Die Daten werden fortlaufend aktualisiert, zuletzt am 5. April 2014.