Russischer Desman
Der Russische Desman (Desmana moschata) ist eine Säugetierart aus der Familie der Maulwürfe (Talpidae). Gemeinsam mit dem Pyrenäen-Desman bildet er die Gruppe der Desmane (Desmanini), die von allen Maulwürfen am ausgeprägtesten an eine wasserbewohnende Lebensweise angepasst sind. Er stellt den größten Vertreter der Maulwürfe dar. Äußerlich kennzeichnend sind der gedrungene Körperbau, die lange und sehr bewegliche Schnauze sowie der lange, abgeplattete Schwanz. Die großen Hinterfüße dienen der Fortbewegung im Wasser. Hingegen sind die Vorderfüße im Unterschied zu anderen, grabenden Maulwürfen eher klein. Die Zehen werden durch Schwimmhäute miteinander verbunden. Das Fell ist dicht und zumeist dunkel gefärbt. Das Vorkommen des Russischen Desmans ist auf Osteuropa beschränkt. Dort bewohnt er breite Flussniederungen im westlichen Teil Russlands und im Nordosten der Ukraine, mit dem Nordwesten Kasachstans wird auch ein Teil von Zentralasien erreicht. Die bewohnten Lebensräume zeichnen sich durch eine reiche Vegetation und zahlreiche Versteckmöglichkeiten an den Uferrändern aus.
| Russischer Desman | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Russischer Desman (Desmana moschata) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name der Gattung | ||||||||||||
| Desmana | ||||||||||||
| Güldenstädt, 1777 | ||||||||||||
| Wissenschaftlicher Name der Art | ||||||||||||
| Desmana moschata | ||||||||||||
| (Linnaeus, 1758) |
Die Tiere leben weitgehend sozial und sind nicht territorial. Ausnahmen treten während der Paarungszeit auf. Die hauptsächlichen Aktivitäten finden in der Dämmerungszeit oder nachts statt. Der Russische Desman hält sich zum Großteil im Wasser auf und ist ein guter Schwimmer. An Land gräbt er einen Unterschlupf, der aus einem komplexen Gangsystem mit Nestkammern besteht. Infolge häufiger auftretender Überschwemmungen wandern die Tiere entlang der Uferlinien. Die Nahrung setzt sich aus Wirbellosen, Wirbeltieren und Pflanzen zusammen. Die bevorzugte tierische Beute besteht aus Würmern, Insekten und Weichtieren. Die Fortpflanzung kann ganzjährig erfolgen, eine Hauptphase beschränkt sich auf das Frühjahr. Ein Wurf umfasst bis zu fünf Neugeborene. Diese werden in einer speziellen Nestkammer aufgezogen. Zur Paarung und während der Aufzucht der Jungen schließen sich einzelne Familienverbände zusammen, bestehend aus den Elterntieren und dem Nachwuchs. Die Tragzeit währt etwa 40 bis 45 Tage, die Jungen verlassen nach rund drei Wochen das Nest.
Die wissenschaftliche Erstbeschreibung des Russischen Desmans erfolgte im Jahr 1758. Zunächst den Bibern zugewiesen, wurde die eigenständige Gattung Desmana im Jahr 1777 aufgestellt. Im 19. Jahrhundert waren die Tiere teilweise unter der Gattungsbezeichnung Mygale bekannt. Fossil traten Vorgänger des Russischen Desmans bereits im Unteren Pliozän auf. In der stammesgeschichtlichen Vergangenheit kamen diese in weiten Teilen Europas vor und sind zudem aus Ostasien dokumentiert. Der heutige Bestand wird als bedroht eingestuft. Historisch war vor allem die Jagd auf das Desmanfell von Bedeutung, die die lokalen Populationen teils stark dezimierte. Heute sind neben der Jagd auch Gewässerverschmutzung und Landschaftszerstörung für den Rückgang des Russischen Desmans verantwortlich.
Merkmale
Habitus

Der Russische Desman ist der größte Vertreter der Maulwürfe. Er erreicht eine Kopf-Rumpf-Länge von 18,5 bis 24,0 cm, der Schwanz wird 18,0 bis 21,0 cm lang. Er besitzt dadurch gut 90 bis 95 % der Länge des restlichen Körpers. Das Gewicht beträgt 370 bis 440 g. Ein ausgeprägter Geschlechtsdimorphismus ist nicht vorhanden. Der Körperbau wirkt kompakt und gedrungen. Einzelne Besonderheiten ergeben sich aus der semi-aquatischen Lebensweise. Der Kopf ist insgesamt konisch geformt und endet in einer langgestreckten und sehr beweglichen Schnauze. Sie erinnert dadurch an einen Rüssel. Nach vorn ist sie ober- und unterseits abgeplattet, auf der Unterseite verläuft eine längsgerichtete Rille. Die Nasenlöcher sind nach oben gerichtet und können unter Wasser geschlossen werden. Die Schnauze ist mit zahlreichen Vibrissen in mehreren Reihen ausgestattet. Die vorderen Tasthaare werden nur 1 bis 2 mm lang und bilden teilweise eine Art Teppich. Nach hinten nehmen sie kontinuierlich an Länge zu und erreichen am Ende, kurz vor dem Auge, gut 39 mm. Ebenso sind am Kinn kurze Tasthaare in mehreren Reihen ausgebildet. Allgemein haben die Vibrissen einen schmalen Schaft und eine etwas verbreiterte kegelförmige Spitze.[1] Der Nasenspiegel ist scheinbar nackt, wird aber von zahlreichen Mikrovibrissen von rund 200 μm Länge bedeckt, die zwischen kleinen buckelförmigen Erhebungen in rosettenartiger Anordnung sprießen. Sie stellen wohl ein Äquivalent zum Eimerschen Organ der grabenden Maulwürfe dar.[2] Die Augen sind klein, äußere Ohren fehlen.[3][4][5]
Das Fell ist auf der Rückseite dunkel rötlichbraun bis nahezu schwarz gefärbt. Jungtiere sind generell blasser. Die Unterseite hebt sich aschgrau hervor. Albinismus wurde bisher nur selten beobachtet und tritt bei einem von rund 5000 bis 6000 Individuen auf. Es lässt sich eine dichte Unterwolle und ein langes, gröberes Deckhaar unterscheiden. Das Deckhaar umfasst einerseits Leithaare, die mit 14,1 bis 21,5 mm am längsten sind und einen zylindrischen Schaft sowie eine etwas verbreiterte spindelförmige Spitze besitzen. Andererseits werden die Grannenhaare mit 10 bis 20 mm etwas kürzer, haben eine gewellten Schaft mit bis zu zehn Krümmungen und einen lanzettförmigen Abschluss. Beiden Haartypen fehlt die Markröhre in den oberen Abschnitten. Die Wollhaare der Unterwolle erreichen nahezu die Länge der Grannenhaare, sind aber mit bis zu 16 Krümmungen stärker gewellt und nur einfach aufgebaut. Sie stehen in Büscheln von bis zu sieben Haaren. Generell ist das Fell sehr dicht, wobei dieses bei den Weibchen deutlicher zum Tragen kommt als bei den Männchen. Am Bauch besteht es aus bis zu 20.750 Haaren je Quadratzentimeter und ist damit dichter als am Rücken mit 15.990 Haaren. Sehr dicht behaarte Bereiche können bis zu 25.000 Haare je Quadratzentimeter aufweisen.[6] Davon entfallen am Rücken 86 bis 89 % auf die Unterwolle, am Bauch sind es 90 bis 93 %. Den Rest macht das Deckhaar aus. Die Haut hat einen silbrigen Farbton und ist mit 0,43 mm an der Brust dicker als am Rücken, wo sie etwa 0,28 mm misst. Der Schwanz weist nur eine spärliche Behaarung auf. Er ist vollständig seitlich abgeplattet, im Gegensatz zum Pyrenäen-Desman (Galemys pyrenaicus) mit seinem nur Ende abgeflachten Schwanz. An der Wurzel weist er die größte Breite auf. Hier sind Drüsen ausgebildet, die einen moschusartig duftendes Sekret absondern. Bedeckt wird der Schwanz von Schuppenringen.[3][4][5]
Die Hinterfüße sind fast doppelt so groß wie die Vorderfüße, was einen auffallenden Unterschied zu den grabenden Maulwürfen bildet. Sie weisen jeweils fünf Strahlen auf, die kräftige Krallen tragen. Zwischen den Fingern und Zehen sind Schwimmhäute ausgebildet. Des Weiteren werden die Fußkanten von steifen und platten Borstenhaaren gesäumt, was die Gesamtfläche der Fußsohle verbreitert und so hauptsächlich den Hinterfüßen beim Paddeln zugutekommt. Die Hinterfußlänge beträgt 5,1 bis 5,8 cm. Weibchen verfügen über vier Zitzenpaare. Beim Männchen ist der Kopf des Penis mit Stacheln besetzt.[3][4][5]
Schädel- und Gebissmerkmale
Der Schädel wird 55 bis 58 mm lang und am Hirnschädel 28,9 bis 30,8 mm breit. Die Breite an den Jochbögen beträgt 21,5 bis 22,9 mm. Er ist groß und massiv mit einem breiten, aber kaum aufgewölbten Hirnschädel. Die Einziehung im Bereich der Orbita zeigt sich nur schwach. Das Rostrum zieht lang aus, die Seitenkanten verlaufen parallel zueinander. Am Stirn- und Scheitelbein bestehen markante Rippeln als Ansatzstellen einer massiven Kaumuskulatur. Der Jochbogen ist relativ schmal. Am Hinterhauptsbein wölben sich die Gelenke zur Artikulation mit der Halswirbelsäule nur schwach hervor, ihre Gelenkflächen weisen nach unten. Der Unterkiefer wiederum besitzt einen kräftigen Bau mit hoch aufsteigendem und fast im rechten Winkel zum horizontalenm Knochenkörper stehenden Kronenfortsatz sowie einem deutlichen Winkelfortsatz. Das Gebiss umfasst mit 44 Zähnen die ursprüngliche Anzahl, die bei den Höheren Säugetieren üblich ist. Häufig wird folgende Zahnformel angegeben: .[7] Die Knochennaht zwischen dem Mittelkieferknochen und dem Oberkiefer verläuft entlang des zweiten vorderen oberen Zahnes. Da bei den Säugetieren der Mittelkieferknochen alleinig die Schneidezähne trägt, besteht die Möglichkeit, dass nur zwei Incisiven ausgebildet sind. Alternativ wird daher auch als Zahnformel angegeben. Sie setzt aber voraus, dass beim Russischen Desman evolutiv ein Schneidezahn verloren ging und ein zusätzlicher Prämolar ausgebildet wurde. Der vorderste obere Schneidezahn ist sehr groß und dolchartig gestaltet. Dahinter liegt ein Diastema, auf das sechs kleine einspitzige Zähne folgen, von denen der dritte etwas größer ist. Dieser stellt je nach Zahnformel den Eckzahn oder ersten Prämolaren dar. Die Molaren sind insgesamt sehr groß und voluminös, haben jedoch niedrige (brachyodonte) Zahnkronen. Die obere Zahnreihe misst 28,8 bis 31,0, die untere 24,2 bis 26,7 mm in der Länge.[3][8][4][5]


Genetische Merkmale
Der diploide Chromosomensatz lautet 2n = 32. Alle Autosomen sind zweiarmig mit sechs metazentrischen, acht submetazentrischen und einem subtelozentrischen Paar. Das X-Chromosom ist groß und submeta- bis subtelozentrisch, das Y-Chromosom klein und metazentrisch. Die Anzahl an Armen der Autosomen (fundamentale Anzahl) beträgt 64.[9][4][5]
Verbreitung und Lebensraum
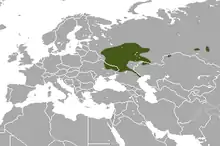
Das Verbreitungsgebiet des Russischen Desmans liegt in Osteuropa und erreicht auch Zentralasien. Es erstreckt sich über den südlichen europäischen Teil Russlands, der östlichen Ukraine und den Nordwesten Kasachstans. Die Hauptvorkommen finden sich entlang der Flussläufe des Dneprs, des Dons, der Wolga einschließlich der Kama und des Urals. In der Ukraine ist die Art lediglich noch am Seim zu finden.[10] Einzelne neuangesiedelte Populationen sind am Oberlauf des Ob im westlichen Sibirien zu finden, so beispielsweise bei Tscheljabinsk, Tomsk und Nowosibirsk.[11] In anderen Regionen wie etwa am Kerschenez bei Nischni Nowgorod, wo der Russische Desman bis in die 1990er Jahre vorkam, wurde er wieder eingeführt.[12][4][5]
Als Lebensräume bevorzugen die Tiere permanente Gewässer in Überflutungsebenen. Häufig handelt es sich um kleinere Seen und langsam fließende Flüsse mit einem schlammigen Boden und einem hohen Reichtum an Wirbellosen, als Anzeiger dienen unter anderem Schnecken wie Lymnaea und Viviparus. Die Mindestgröße der Gewässer sollte hierbei 60 bis 100 m² nicht unterschreiten, allerdings werden große Wasserflächen wie ausgedehnte Seen oder Wasserreservoire eher gemieden, ebenso wie Flüsse mit großer Wasserkraft. Zudem muss die Wassertiefe wenigstens 2 bis 3 m betragen, so dass die Gewässer im Winter nicht zufrieren. Besonders zuträglich sind steilere Hänge mit Versteck- und Grabmöglichkeiten. Eine weitere Voraussetzung ist eine dichte holzige Vegetation, welche vor zu strengen Wetterverhältnissen schützt.[13] Nach Untersuchungen an der Kljasma und der Oka reichen die Sommertemperaturen dadurch von 8 bis 23 °C, die Wintertemperaturen von 0 bis 7 °C. Resultierend daraus fallen während der kalten Jahreszeit die Temperaturen in den Unterschlüpfen nicht in den Minusbereich.[14] Die Populationsdichte wird überwiegend anhand der Anzahl der Unterschlüpfe entlang der Fluss- und Seeuferlinie bestimmt. In den meisten untersuchten Habitaten beträgt sie heute 0,5 bis 15,2 Unterschlüpfe auf einem Kilometer Uferlänge. An verschiedenen Gewässern in Mordwinien an der mittleren Wolga wurden in einem Untersuchungszeitraum von 2009 bis 2018 maximal 8,2 bis 10,5 Baue auf entsprechender Uferlänge registriert.[15] Mit zehn bis elf Unterschlüpfen vergleichbare Werte stammen von der Sura aus Beobachtungen im Jahr 2018.[13] In historischer Zeit lag die Anzahl beobachteter Baue teilweise zehn- bis zwanzigmal höher.[5]
Lebensweise
Territorialverhalten
Die Lebensweise des Russischen Desmans ist in Teilen gut erforscht. Die Tiere sind gut an eine semi-aquatische Lebensweise angepasst. Beim Schwimmen nutzen sie die breiten Hinterfüße zum alternierenden Paddeln und den Schwanz zum Steuern. Die Vorderbeine sind meist nach vorn gerichtet. In dieser Haltung schwimmt ein Tier sowohl an der Wasseroberfläche als auch bei Tauchgängen und kann dadurch auch abrupte Wendungen von 90° durchführen. In einzelnen Fällen werden beide Hinterbeine gleichzeitig eingesetzt.[16] Beim Tauchen liegt der Körper üblicherweise schräg mit dem Kopf nach unten in einem 45°-Winkel zum Untergrund.[17] An der Wasseroberfläche hält ein Tier seine lange und bewegliche Schnauze aufrecht, so dass er wie ein Schnorchel wirkt. Gegen die Strömung bei mäßigen Fließgeschwindigkeiten kann ein Tier in rund sechs Stunden 100 m zurücklegen, flussab ist es rund anderthalb mal so schnell. In der Regel hält sich der Russische Desman relativ ufernah auf, selten dringt er mehr als 4 m weit in ein Gewässer vor. Landgänge absolviert er nur selten, verursacht wird dies meist durch einen starken Anstieg des Wasserspiegels. Dort ist er eher unbeholfen, die breit stehenden Beine verhindern ein schnelles Laufen über weite Strecken. Die niedrige Körperhaltung lässt den Bauch und Schwanz jeweils den Boden berühren. Höchstgeschwindigkeiten wurden mit etwa 1 m/s gemessen, die sich aber nach kurzen Distanzen halbiert haben. Die teils steilen Uferböschungen werden mit Hilfe der Krallen der Vorderbeine erklommen, allerdings vermag der Russische Desman nicht hangab zu klettern. Die Orientierung erfolgt über das Gehör, den Geruch und durch Tasten mittels der Vibrissen. Zum Riechen unter Wasser stößt ein Tier Luftblasen aus der Nase, an die Geruchsstoffe bestimmter Objekte anhaften. Durch das erneute Einatmen der Luftblasen können diese wahrgenommen werden.[18] Zur Orientierung dient höchstwahrscheinlich auch das Sekret der Moschusdrüse, mit dem die Tiere ihre Wege und Pfade einreiben.[3][19][20][5]
Der Russische Desman ist im Unterschied zu zahlreichen anderen Maulwurfsarten mit ihrem einzelgängerischen und territorialen Verhalten ein eher soziales Tier. In menschlicher Obhut leben männliche und weibliche Individuen oft friedlich zusammen. Allerdings findet man in freier Wildbahn selten mehr als zwei fortpflanzungsfähige Weibchen in einem Gewässer. Männchen bilden hingegen Gruppen und sind weniger aggressiv untereinander als die Weibchen. In der Fortpflanzungszeit bilden sich Familiengruppen heraus, die aus den Elterntieren und dem Nachwuchs bestehen und bis zu sieben Individuen umfassen können. Hier ist dann vor allem in der Anfangsphase eine stärkere Territorialität zu beobachten, da die einzelnen Familiengruppen ihr Revier verteidigen. Intraspezifisch beginnt dies mit einem Betasten mit der langen Schnauze, was nach mehrmaliger Wiederholung in Kämpfe münden kann. Dabei richtet sich ein Tier auf den Hinterbeinen auf und lässt sich dann auf den Rücken fallen, um sich mit den Krallen zu verteidigen. Zudem ist der Russische Desman stärker lautgeberisch. Bekannt ist ein lautes Kreischen als Droh- oder Abwehrmaßnahme, das bereits in einer gegenseitigen Distanz von 1,5 m ausgestoßen wird. Ein lautes Knattern oder Trillern, das je nach Erregungszustand bis zu 40 m weit hörbar ist und weitgehend in der Dämmerungszeit ertönt, wird als Kontaktruf eingestuft. Dagegen dient ein Piepsen möglicherweise eher der Beschwichtigung, da es zum Teil auch beim Nachwuchs eingesetzt wird. Die Laute nehmen einen Frequenzbereich on 300 bis 1700 hz ein.[21][17] Zum Komfortverhalten zählt die Fellpflege, die sehr intensiv sein kann und überwiegend mit den Hinter- seltener mit den Vorderfüßen durchgeführt wird. In Ruheposition ist ein Tier eingerollt mit der Schnauze unter dem Bauch, der Schwanz steht jedoch ab.[19][3][20][5]
Abweichend vom Pyrenäen-Desman zeigt der Russische Desman stärkere Grabungsaktivitäten. Jede Familiengruppe nutzt ein komplexes Behausungssystem, das in der Regel in steile Uferböschungen gegraben ist. Die Eingänge, die rund 14 cm messen, liegen immer unter Wasser, zumeist so tief, dass sie im Winter bei gefrorenem Wasser zugänglich bleiben. Häufig hat ein System mehrere Eingänge, Ring- und Verbindungswege sowie Nestkammern. Die Gänge verlaufen landein und können sich bis zu 10 m vom Ufer entfernen. In Steilhängen sind sie in der Regel kürzer und gut 5 m lang, in flacheren Uferböschungen dehnen sie sich auf bis zu 22 m aus. Die Nestkamern befinden sich oberflächennah, etwa 20 bis 30 cm unter dem Erdboden, aber hoch genug, dass sie von Wasserhochstand verschont bleiben. Umgeben sind sie in vielen Fällen vom Wurzelwerk der Bäume, das eine natürliche Barriere für die Grabetätigkeit des Russischen Desmans darstellen. Eine einzelne Nestkammer hat einen Durchmesser von 17 bis 35 cm und ist mit einem Nest aus Pflanzenmaterial ausgestattet. Etwas geräumiger mit bis zu 40 cm Durchmesser sind jene zur Aufzucht des Nachwuchses. Von den Nestkammern gehen bis zu drei Sackgassen ab, die 20 bis 30 cm lang sind. Das Behausungssystem schließt auch verschiedene Vorratslager oder Fresskammern ein. Eine Familiengruppe legt zusätzlich drei bis fünf weitere Nebenbehausungen an. Sie dienen als Ruhe- und Fressplatz und sind im letzteren Fall mit zahlreichen Nahrungsresten gefüllt. Zeitweise genutzte Unterschlüpfe können auch während Hochwasserereignissen aufgesucht werden, in manchen Fällen ziehen sich die Tiere dann in Baumstümpfe oder -höhlen zurück. Zudem ist der Russische Desman migratorisch, was sich aus den instabilen lokalen Verhältnissen mit auftretenden Hochwassern ergibt. Wanderungen bis zu 7 km flussab oder 3,5 km flussauf wurden beobachtet. Die längste ist mit 16 km in 69 Tagen verzeichnet. Verschiebungen der Aktionsräume infolge von Dürre sind weniger umfangreich und reichen oft nur wenige hundert Meter weit.[3][20][5]
Die überwiegenden Aktivitäten des Russischen Desmans finden nachts statt mit den Hauptphasen während der Abend- und Morgendämmerung. Dies kann im Winter, wenn die Gewässer gefroren sind, in eine dreiphasige Aktivität überleiten. Die aktiven Phasen verteilen sich dann auf die Zeiträume von 05.30 bis 07.30 Uhr, 12.30 bis 14.30 Uhr und 19.30 bis 21.30 Uhr. Tagsüber zieht sich ein Tier in seinem Bau zurück. Nach Untersuchungen in Mordwinien mit Stimmenrekordern zeigt der Russische Desman hierbei eine größere Aktivität am frühen Nachmittag und in den frühen Morgenstunden kurz vor seinem Aufbruch. Insgesamt verbringt er 17 Stunden in seinen Tunneln und Gängen, weitere sieben Stunden nutzt er diese nicht, wobei er dann entweder ruht oder außerhalb der Tunnel im freien Wasser aktiv ist. Tiere am Fluss Oka hingegen sind weiteren Analysen zufolge täglich zwischen 11,4 und 15,5 Stunden aktiv, was 48 bis 65 % der gesamten Tageszeit entspricht. Im Wasser halten sie sich, individuell abweichend, durchschnittlich 5,3 bis 9,0 Stunden auf.[22][3][20][5]
Ernährungsweise
Die Nahrung des Russischen Desmans weist ein vielfältiges Spektrum auf und besteht aus Wirbellosen, Wirbeltieren und Pflanzen. Unter den Wirbellosen sind verschiedene Würmer von Bedeutung, wie etwa Gordius aus der Gruppe der Saitenwürmer und Lumbricus aus der Gruppe der Regenwürmer. Hinzu kommen Egel und Spinnen sowie Krebse. Bei letzteren haben Gattungen wie Asellus und Potamobius größere Bedeutung. Weitere Beutetiere sind Insekten und deren Larven. Hier dominieren Köcherfliegen, Libellen, Zweiflügler, Käfer und Eintagsfliegen. Eine umfangreiche Gruppe bilden die Weichtiere mit unter anderem Viviparus, Lymnaea, Planorbis, Bithynia, Valvata oder Anodonta. Neben Fröschen ergänzen unter den Wirbeltieren vor allem Fische, darunter Hecht, Rotauge, Europäischer Schlammpeitzger und Flussbarsch, den Speiseplan. Tiere in Gefangenschaft fraßen aber auch Mäuse und anderes Fleisch. Zu den bevorzugten Pflanzen zählen Schachtelhalmartige, wie Schachtelhalme selbst, Froschlöffelartige wie Froschlöffel, Pfeilkraut, Drachenwurz und Laichkräuter, daneben auch Süßgrasartige wie Schilfrohr, Rohrkolben, Igelkolben, Seggen und Simsen oder Seerosenartige wie Seerosen und Teichrosen sowie Nelkenartige wie Ampfer.[3][20][5]
Der Anteil der einzelnen Komponenten der Nahrung kann über das Jahr variieren und wird vom Angebot bestimmt. Pflanzen werden überwiegend in den Frühjahr- und Herbstmonaten konsumiert, können aber das gesamte Jahr über Bestandteil bleiben. Fische nehmen im Winter einen größeren Anteil ein. Zudem scheinen regionale Unterschiede zu bestehen, da Untersuchungen zufolge Tiere an der Wolga im Sommer häufig Köcherfliegen als Nahrungsressource nutzen, solche am Don hingegen Zweiflügler, Egel und Weichtiere. Generell spielen bei ersteren Zweiflügler und Weichtiere nur eine untergeordnete Rolle, während sie bei letzteren einen bedeutenden Prozentsatz in der Nahrung ausmachen. Der Magen eines Individuums fasst im Durchschnitt 1,7 g, im Maximum bis zu 7 g. Der Flüssigkeitsbedarf ist mit durchschnittlich 120 cm³ am Tag vergleichsweise hoch, die gebrauchte Menge hängt aber vom Feuchtigkeitsgehalt der Nahrung ab.[3][20][5]
Seine Nahrung sucht der Russische Desman auf dem Grund der Gewässer. Auf seinen Tauchgängen inspiziert er zumeist verschiedene Furchen, die er in den Boden gegraben hat. Diese bilden natürliche Fallen für die häufig wenig bewegliche Nahrung wie Weichtiere. Das Aufspüren erfolgt mit der rüsselartigen Nase oder mit den Tasthaaren. Erbeutete Nahrung wird in den Bau oder an Land verbracht, nur selten frisst ein Tier sie direkt vor Ort.[3][17]
Fortpflanzung
Die Fortpflanzung des Russischen Desmans ist nur partiell gut untersucht. Er kann sich prinzipiell ganzjährig paaren, ein Höhepunkt wird im Frühjahr erreicht. Zu diesem Zeitpunkt ist die höchste Aktivität der Tiere zu verzeichnen. Die Tiere sind monogam und formen kleine Familiengruppen aus drei bis sieben Individuen. Inwiefern Weibchen zwei Geburten im Jahr absolvieren, ist bisher nicht vollständig geklärt. Die mit 80 % meisten Jungtiere werden in den Monaten Juli und August geboren, säugende oder milchproduzierende Weibchen können aber von März bis Oktober beobachtet werden. Ein Muttertier trägt zwischen eins und fünf Embryonen aus. Der Durchschnitt variiert regional von 2,5 bis 3,7. Die Anzahl der Neugeborenen ist etwas geringer, allerdings besteht die Tendenz, das Würfe im Frühjahr und Sommer größer sind. Die Tragzeit beträgt rund 40 bis 45 Tage. Neugeborene sind zwischen 75 und 85 mm lang und wiegen etwa 16 g. Sie haben nackte Haut, geschlossene Ohren und sind blind sowie zahnlos. In den ersten zehn Tagen verbleibt das Muttertier im Nest des Nachwuchses, wechselt dann aber in ein nahegelegenes. Von da an besucht sie die Jungen regelmäßig. Das Vatertier ist an der Aufzucht beteiligt. Nach fünf bis sieben Tagen wachsen bei den Jungtieren silbergraue Haare auf dem Rücken, der Bauch bleibt vorerst nackt. Das Fell ist mit einem Alter von zehn Tagen ausgeprägt, zu diesem Zeitpunkt sind die Jungen rund 110 mm lang. Die Ohren öffnen sich nach 18 Tagen. Das Nest verlassen die Jungen erstmals nach rund 23 Tagen, die erste feste Nahrung nehmen sie nach fast vier Wochen zu sich. Die Gewichtszunahme verläuft regelmäßig. Bei einem Alter von anderthalb bis zwei Monaten wiegt ein Junges etwa 196 g, nach zehn Monaten sind es etwa 333 g. Männchen erreichen mit vier bis sechs Monaten die Geschlechtsreife, bei Weibchen dauert der Prozess fast doppelt so lang. Die Familiengruppen bleiben den gesamten Sommer bis zum Herbst zusammen. Danach wandert der Nachwuchs ab und die Elterntiere trennen sich.[3][20][5]
Die Sterblichkeitsrate der Neugeborenen liegt bei 40 %. Für alle Altersklassen gilt, dass das Überleben maßgeblich von äußeren Faktoren wie den Umweltbedingungen beeinflusst wird. Eine große Rolle spielt der Wasserstand. Dieser kann durch Regen, frühjährliche Schneeschmelze oder Überflutungen ansteigen, wodurch die Tiere gezwungen sind, ihre Baue zu verlassen und somit Fressfeinden potentiell ausgeliefert sind. Trockenheiten senken den Wasserspiegel und mindern das Nahrungsangebot. Lokal ist die Bestandsgröße somit größeren Fluktuationen unterworfen. Nach Untersuchungen im Oka-Naturschutzgebiet schwankte sie im Zeitraum von 1938 bis 1957 zwischen 69 und 207 Individuen. Höchststände wurden hier jeweils in Zeiten mit mäßigem Niederschlag erreicht. Das Höchstalter unter günstigen Bedingungen liegt bei rund vier Jahren.[3][20][5]
Fressfeinde, Kommensalen und Parasiten
Durch Nachweise in Kothaufen sind der Rotfuchs, Marderhund, Hermelin und Iltis, aber auch der Europäische Nerz und Baummarder als Fressfeinde belegt. Jedoch hat der Russische Desman jeweils nur einen geringen Anteil im Nahrungsspektrum. Unter den Vögeln sind der Schelladler und Uhu zu nennen.[23] Die Hauptgefahr für den Russischen Desman erbeutet zu werden, besteht bei seinen gelegentlichen Landgängen. Im Wasser fallen manchmal Jungtiere Hechten oder Europäischen Welsen zum Opfer.[3]
Darüber hinaus tritt die Wasserspitzmaus als Konkurrent auf, die eine ähnliche ökologische Nische wie der Russische Desman besetzt. Ähnliches gilt für die Schleie und die Karausche. Andere Tiere nutzen die durch den Russischen Desman gegrabenen Baue nach. Bekannt ist dies etwa von der Ostschermaus, der Feldmaus und der Brandmaus. Andere werden durch die Ringelnatter, die Rotbauchunke, die Erdkröte und den Seefrosch repräsentiert. Hinzu kommt der Schlammpeitzger. Gegenüber der Ostschermaus sind alle anderen genannten nur untergeordnet vertreten.[3]
Die Parasiten sind recht gut untersucht. Äußerliche Schmarotzer treten mit Milben wie Labidophorus, Eadiea, Haemogamasus, Macrolaelaps und Echinolaelaps in Erscheinung. Sie befallen sowohl die Nester als auch den Körper und werden wohl durch Nagetiere eingeschleppt. Von Bedeutung ist der „Desmanfloh“ Silphopsyllus, ein Käfer, der in der Unterwolle sitzt und sich wahrscheinlich von Haut ernährt. Innere Parasiten umfassen neben Kratzwürmern wie Skrjabinonerus, Cyathocotyle und Alaria sowie Bandwürmern wie Dilepis auch mehrere Formen der Fadenwürmer. Dazu gehören Porrocaecum, Agamospirura, Metastrongyloides und Thominx. Ein bekanntes Krankheitsbild ist das Erysipeloid.[24][25][3]
Systematik
Innere Systematik der Maulwürfe nach He et al. 2016[26]
|
Der Russische Desman ist eine Art aus der Gattung Desmana. Er repräsentiert deren einzigen rezenten Vertreter, wodurch die Gattung monotypisch ist. Art und Gattung gehören wiederum der Familie der Maulwürfe (Talpidae) an. Gemeinsam mit dem nahe verwandten Pyrenäen-Desman (Galemys) wird der Russische Desman innerhalb der Familie der Tribus der Desmane (Desmanini) zugewiesen. Die Desmane unterscheiden sich von anderen Maulwürfen in ihrer semi-aquatische Lebensweise. Sie weisen dadurch einige anatomische Besonderheiten auf. Dies betrifft ihren Körperbau, der weniger an eine grabende Aktivität angepasst ist. Erkennbar wird dies unter anderem an dem schlanken Oberarmknochen und den gegenüber den vorderen Füßen größeren hinteren. Des Weiteren sind die unteren Beinabschnitte länger und die Zehen durch Schwimmhäute miteinander verbunden, gleichfalls ist der Schultergürtel anders konfiguriert.[5] Ursprünglich wurden die Desmane als eigenständige Unterfamilie neben den grabenden Maulwürfen geführt.[27][28] Molekulargenetische Untersuchungen ergaben jedoch eine nähere Beziehung mit den Eigentlichen Maulwürfe (Talpini) und dem Sternmull (Condylurini). Als enger verwandte Gruppe zu den genannten Triben werden die verschiedenen Spitzmulle Ostasiens und Nordamerikas betrachtet. Die Trennung von den anderen Linien der Maulwürfe fand bereits im Oberen Eozän vor rund 37 bis 34 Millionen Jahren statt, die Aufspaltung der Desmane in die heutigen Linien setzte aber erst deutlich später, im Verlauf des Mittleren Miozäns vor 13,9 Millionen Jahren ein. Übergeordnet lassen sich alle genannten Maulwurfsgruppen der Unterfamilie der Altweltmaulwürfe (Talpinae) zuweisen. Innerhalb letzterer stehen zusätzlich noch die Neuweltmaulwürfe (Scalopini). Die Altweltmaulwürfe umfassen sowohl unterirdisch grabende als auch semi-aquatische lebende Tiere aus Eurasien und aus Nordamerika.[29][30][26]
Wie die Gattung so wird auch die Art als monotypisch eingestuft, Unterarten sind dadurch nicht bekannt.[31][4][5] Allerdings wurden mehrere fossile Unterarten benannt, dazu gehören unter anderem D. m. magna, D. m. mosbachensis, D. m. lebenstedtensis oder D. m. moravica. Der größte Teil davon stammt aus dem Pleistozän. Die Unterscheidung basiert weitgehend auf metrischen Unterschieden, doch halten einige Forscher das bisherige Fundmaterial für zu gering, um derartige Abtrennungen sicher vornehmen zu können.[32][33]
Neben dem heutigen Russischen Desman enthält die Gattung Desmana noch mehrere fossile Formen:[34][35]
- Desmana inflata Rümke, 1985
- Desmana kovalskae Rümke, 1985
- Desmana marci Minwer-Barakat, García-Alix, Martín-Suárez & Freudenthal, 2020
- Desmana nehringi Kormos, 1913
- Desmana themalis Kormos, 1930
- Desmana verestchagini Topachevski, 1961
Weitere Formen sind aus dem östlichen Europa benannt worden. Das betrifft unter anderm Desmana jalpugensis, Desmana kujalnikensis, Desmana meridionalis oder Desmana moldavica. Ihre Beschreibungen basieren häufig auf isolierten Zähnen, ein Vergleich mit den anerkanntzen Fossilvertretern fand nur selten statt. Aufgrund dessen ist ihr Status unklar.[36][37] Desmana pontica wiederum, 1940 durch Antje Schreuder basierend auf Funden aus Polgardi in Ungarn etabliert,[32] wurde später durch Cornelia G. Rümke zu Dibolia[34] und anschließend zu Archaeodesmana überführt.[35]
Forschungsgeschichte

Einen der frühesten Hinweise auf den Russischen Desman gab Charles de l’Écluse im Jahr 1605. Er stellte hierbei ein Tier vor, dass er Mus aquaticus nannte.[38] Auf ihn berief sich dann gut 150 Jahre später Carl von Linné, als er im Jahr 1758 in seinem für die biologische Nomenklatur wegweisenden Werk Systema Naturae die wissenschaftliche Erstbeschreibung des Russischen Desmans veröffentlichte. Linné ordnete hierin die Art mit der Bezeichnung Castor moschatus der Gattung der Biber zu.[39] Als Besonderheit hob er den langen, lanzettförmigen und abgeplatteten Schwanz hervor (cauda longa lancelota plana), ein Charakteristikum, das er schon zwölf Jahre zuvor in seinem Werk zur Fauna Schwedens erwähnt hatte.[40] in beiden Veröffentlichungen verwies Linné auf die umgangssprachliche Bezeichnung „Desman“. Als Typusregion gab er Russland an. Nur wenige Jahre nach Linné, 1771, stellte Peter Simon Pallas im ersten Band seiner Reisebeschreibungen durch Russland den Russischen Desman genauer vor. Er tätigte dies unter dem Trivialnamen „Bisamratte“ und widersprach zudem Linné, indem er die Form als Sorex moschatus zu den Rotzahnspitzmäusen verwies.[41] Die Einschätzung übernahm im Jahr 1778 auch Johann Christian Schreber.[42] Bereits ein Jahr zuvor hatte Johann Anton Güldenstädt jedoch die Gattung Desmana aufgestellt. Er tat dies mit der Begründung, dass die Trivialbezeichnung „Desman“ sehr geläufig sei, zumal sie auch von Georges-Louis Leclerc de Buffon in seiner mehrbändigen Histoire Naturelle im Französischen verwendet worden war.[43] Ähnlich wie zuvor Pallas vermutete Güldenstädt eine engere Verwandtschaft des Russischen Desmans mit den Spitzmäusen.[44]

Sowohl die Gattungsbezeichnung Desmana als auch der Trivialname „Desman“ wurden von einigen Naturforschern nachfolgend als unpassend gesehen. Ursprünglich ein schwedisches Wort verweist dieses einerseits als desmansråtta auf den Russischen Desman, andererseits steht desman auch für den moschusartigen Duft und ist mit dem mittellateinischen bisamum für den Geruchsstoff Bisam verwandt. Aufgrund seiner osteuropäischen Verbreitung plädierte im Jahr 1836 Johann Friedrich von Brandt in einem Aufsatz für den umgangssprachlichen Namen „Wychochol“, wie er vor allem in Russland gebraucht wurde. Als wissenschasftliche Gattungsbezeichnung schlug er Myogale vor. In seinem Schreiben behandelte Brandt zudem den inneren Bau des Russischen Desmans und erkannte eine nähere Verwandtschaft zu den Maulwürfen.[45] Myogale wiederum ist eine Entlehnung vom Gattungsnamen Mygale, den Georges Cuvier im Jahr 1800 für den Russischen Desman eingeführt hatte.[46] Cuvier selbst übernahm ihn von Anders Jahan Retzius. der vier Jahre zuvor mit Mygalus auf den Russischen Desman verwiesen hatte.[47] Allerdings erwies sich Mygale als problematisch, da nahezu zeitparallel, im Jahr 1802, mit dem gleichen Gattungsnamen eine Artengruppe der Spinnen belegt worden war.[48] Myogale wurde zwar von diversen Forschern bevorzugt, so etwa von George Edward Dobson im Jahr 1883 in seinem umfassenden Werk über Insektenfresser,[49] im Sinne der Prioritätsregel der biologischen Nomenklatur hat aber Güldenstädts Desmana aufgrund der früheren Benennung Vorrang. Daher sprach sich Antje Schreuder im Jahr 1940 für diese Gattungsbezeichnung aus, was weitgehend akzeptiert ist.[32]
Stammesgeschichte
Die Stammesgeschichte der Gattung Desmana reicht bis in das beginnende Pliozän zurück. Funde verteilen sich im Unterschied zur Verbreitung des heutigen Russischen Desmans auf weite Teile Europas, wurden darüber hinaus auch aus Ostasien beschrieben. Unklar ist die Zuweisung einiger Gebissreste vom Eichkogel südlich von Wien, die noch in das Obere Miozän gehören.[50][51] Die ältesten eindeutigen Hinweise bilden Fossilreste aus Calicasas in der Umgebung von Granada im südlichen Spanien. Das sehr umfangreiche Zahnmaterial lässt auf einen mittelgroßen Angehörigen der Gattung schließen, der Desmana marci zugewiesen wird. Typisch für ältere Vertreter von Desmana war der vorderste obere Schneidezahn mit zwei Spitzen ausgestattet. Außerdem war er bei Desmana marci noch deutlich niederkroniger.[35] im weiteren südlichen Europa kam im Pliozän zusätzlich Desmana verestchagini vor, eine eher kleine Form von Desmana. Ursprünglich 1961 anhand von Funden aus Kosyakin bei Stawropol im Kaukasus benannt, ist die Art darüber hinaus auch von mehreren Lokalitäten in Griechenland dokumentiert worden, so unter anderem von Spilia und Ptolemais. Es liegen hauptsächlich Unterkieferreste vor.[34] Weitere Funde aus dieser Zeit sind unter anderem aus dem Gebiet nördlich der Alpen belegt. Ein bedeutender Fundplatz ist mit Węże im südlichen Polen verzeichnet. Das von hier stammende Material in Form von Unterkieferresten wurde bereits in den 1960er Jahren von Andrzej Sulimski vorgestellt und im Jahr 1985 von Cornelia G. Rümke zu Desmana kovalskae verwiesen.[52][53][34] Möglicherweise taucht die Art auch im entwickelten Pliozän in der Goazhuang-Formation im Yushe-Becken in der heutigen chinesischen Provinz Shanxi auf.[54][55][35]
Zum Ende des Pliozäns tritt mit Desmana nehringi eine relativ große Form auf. Sie wurde erstmals anhand von Unterkieferresten aus Beremend im südlichen Ungarn dokumentiert und von Theodor Kormos im Jahr 1913 aufgestellt.[56] Neben weiteren ungarischen Lokalitäten wie Villany mit einem für die Art recht umfangreichen Material liegen Funde aus Hajnáčka in Tschechien[57] und aus Rebielice Królewskie in Polen vor.[34] Im südwestlichen Europa ist zu diesem Zeitpunkt außerdem Desmana inflata präsent. Die Typuslokalität der Art bildet Valdeganga in der südostspanischen Provinz Albacete. Relativ benachbart kann noch Galera in der Granada genannt werden.[58] Den wissenschaftlichen Namen verdankt Desmana inflata den recht voluminösen Zähnen.[34] Für das Pleistozän bedeutend ist die Art Desmana thermalis, die ebenfalls auf Kormos zurückgeht und von ihm im Jahr 1930 anhand zahlreicher Gebiss- und Skelettreste aus Püspökfürdö im ungarisch-rumänischen Grenzgebiet aufgestellt worden war.[59] Sie ist dann auch aus dem nordalpinen Gebiet wie etwa von den bedeutenden altpleistozänen Fundstellen Hundsheim in Niederösterreich[60] und Tegelen in den Niederlanden belegt.[34] Dem großen Vertreter der Gattung Desmana wird häufig eine Übergangsstellung zwischen den älteren Formen und dem heutigen Russischen Desman bescheinigt. Möglicherweise haben auch die Unterkieferreste aus Schöningen in Niedersachsen eine ähnliche Position inne. Diese datieren in eine Warmzeit des entwickelten Mittelpleistozäns und dürften somit rund 300.000 Jahre alt sein.[61][62]
Wahrscheinlich schon in das ausgehende Altpleistozän datieren die ältesten Fundreste des heutigen Russischen Desmans. Gefunden wurden sie in den Mosbacher Sanden bei Wiesbaden. Die Art kommt dort auch in jüngeren Ablagerungseinheiten vor, die dem beginnenden Mittelpleistozän angehören.[63] Der Russische Desman besaß im Pleistozän ein weit nach Westen ausgedehntes Verbreitungsgebiet. Belegt wurde er im frühen Mittelpleistozän von mehreren Lokalitäten in England. Im späten Mittelpleistozän trat er unter anderem auch im heutigen Baltikum auf, wie Funde aus Nyaravai in Litauen aufzeigen. Etwas jünger, da bereits in einem frühweichsel-kaltzeitlichen Zusammenhang lassen sich Reste aus dem oberen Seeuferhorizont von Neumark-Nord 2 im Geiseltal bei Halle in Sachsen-Anhalt einstufen.[64] Aus der Endphase der letzten Kaltzeit können Funde aus Nørre Lyngby in Dänemark und aus Zachow bei Potsdam in Brandenburg genannt werden, letztere umfassen neben dem Oberarmknochen einzelne weitere Gliedmaßenreste.[65][33] Einige isolierte Zähne wurden aus der Komarowa-Höhle bei Częstochowa im südlichen Polen berichtet.[66] Für ein zu dieser Zeit auch weit nördliches Vordringen der Art im heutigen Russland zeugen unter anderem Fundstellen im mittleren Uralgebiet und am Unterlauf des Irtysch. Noch einmal rund 400 km weiter nördlich liegen zwei Zähne aus der Sedju-Höhle im Becken der Petschora in der russischen Republik Komi vor.[67][68]
Bedrohung und Schutz
_fur_skin.jpg.webp)
Der Russische Desman wird von der IUCN in die Kategorie „stark gefährdet“ (endangered) eingestuft. Die Naturschutzorganisation begründet dies mit einem starken Rückgang der Population. Untersuchungen zufolge verringerte sich der Bestand zwischen 2001 und 2013 um über 50 %. Der Gesamtbestand liegt bei schätzungsweise 13.200 Individuen. Auf die Ukraine entfallen davon nicht mehr als 300 bis 500 Tiere, die allesamt in der Region des Seim im äußersten Nordosten des Landes leben. Andere Populationen wie jene am Dnepr oder im Donezk gelten als erloschen.[10] Größte Bedrohungen für den Bestand stellen die Gewässerverschmutzung verbunden mit Verlust und Zerstückelung des Lebensraumes sowie die Konkurrenz durch eingeschleppte Arten wie die Nutria und die Bisamratte dar.[69] Darüber hinaus hat vor allem die illegale Jagd einen starken Einfluss auf die negative Entwicklung. Diese erfolgt mit fixierten Fischernetzen oder mit Elektrofanggeräten. Die Jagd spielte auch in der historischen Vergangenheit eine große Rolle. Noch bis zum Ende des 19. Jahrhunderts war der Russische Desman relativ häufig. Danach setzte ein massiver Rückgang der Bestände aufgrund der Jagd auf das Desmanfell ein. Schätzungsweise wurden Anfang des 20. Jahrhunderts jährlich zwischen 5500 und 100.000 Tiere getötet, allein für 1913 sind in Nischni Nowgorod rund 60.000 Felle gehandelt worden. Ein Verbot der Jagd auf die Tiere im Jahr 1914 hielt nur kurze Zeit an, bereits zwanzig Jahre später gelangten wieder rund 19.000 Felle auf den Markt. Zwischen 1940 und 1956 durfte der Russische Desman nur mit offizieller Genehmigung getötet werden, innerhalb dieses Zeitraums starben infolgedessen rund 175.000 Tiere. Im Jahr 1957 wurde die Jagd endgültig verboten und der Russische Desman unter Schutz gestellt. Trotzdem schrumpfte die Population weiter, von etwa 70.900 Tieren Anfang der 1970er Jahre auf etwa 40.800 im Jahr 1985 und 25.000 im Jahr 2005.[11] Heutige Schutzmaßnahmen betreffen neben Wiederansiedlungen des Russischen Desmans in Regionen mit erloschenen Beständen auch das Verbot bestimmter Fischernetze oder Elektrofischgeräte. Zudem sind Forschungen zur Populationsgröße und genauen Verbreitung notwendig. Die Art kommt in mehreren Schutzgebieten vor, so unter anderem im Nationalpark Ugra.[70][5]
Literatur
- I. I. Barabasch-Nikiforow: Die Desmane. Neue Brehm Bücherei 474, Wittenberg, 1975, S. 1–100
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 604–605) ISBN 978-84-16728-08-4
- Ronald M. Nowak: Walker's Mammals of the World. 2 Bände. 6. Auflage. The Johns Hopkins University Press, Baltimore MD u. a. 1999, ISBN 0-8018-5789-9.
- М. В. Зайцев, Л. Л. Войта und Б. И. Шефтель: Млекопитающие фауны России и сопредельных территорий. Насекомоядные. Санкт-Петербург, 2014, S. 1–390 (S. 112–117)
Einzelnachweise
- В . Ф. Куликов und М. В. Рутовская: Некоторые особенности строения органов чувств и ориентирования русской выхухоли (Desmana moscha L. 1758). Сенсорные Системы 27 (3), 2013, S. 239–247
- Kenneth C. Catania: Epidermal Sensory Organs of Moles, Shrew-Moles, and Desmans: A Study of the Family Talpidae with Comments on the Function and Evolution of Eimer’s Organ. Brain Behavior and Evolution 56, 2000, S. 146–174
- I. I. Barabasch-Nikiforow: Die Desmane. Neue Brehm Bücherei 474, Wittenberg, 1975, S. 1–100
- М. В. Зайцев, Л. Л. Войта und Б. И. Шефтель: Млекопитающие фауны России и сопредельных территорий. Насекомоядные. Санкт-Петербург, 2014, S. 1–390 (S. 112–117)
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 605–606) ISBN 978-84-16728-08-4
- Yu. F. Ivlev, K. A. Makhotkina und M. V. Rutkovskaya: Fur Structure and Grooming in the Semi-aquatic Mammal Russian Desman (Desmana moschataL.). Doklady Biological Sciences 458, 2014, S. 289–293
- Alan C. Ziegler: Dental homologies and possible relationships of recent Talpidae. Journal of Mammalogy 52 (1), 1971, S. 50–68
- Shin-ichiro Kawada, Kazuhiro Koyasu, Elena I. Zholnerovskaya und Sen-ichi Oda: A discussion of the dental formula of Desmana moschata in relation to the premaxillary suture. Mammal Study 27, 2002, S. 107–111
- В. М. Малыгин und Д. Л. Заславский: Особенности кариотипов Русской (Desmana moschata) и Пиренейской (Galemys pyrenaicus) выхухолей в свази с вопросом о положении етих видов в семействе Talpidae (Mammalia, Insectivora). Зоологический Журнал 65, 1986, S. 1435–1437
- Oleksandr Mikhailovich Yemets, Volodymyr Anatoliyovich Vlasenko, Viktor Mikhailovich Demenko, Valentina Ivanivna Tatarynova, Tetiana Oleksandrivna Rozhkova, Alla Oleksandrivna Burdulaniuk, Olha Mikolaivna Bakumenko, Olena Mikolaivna Osmachko und Yuliia Mikhailivna Shcherbyna: Seymska Population of Russian Desman (Desmana moschata L.) in North-Easten Part of Ukraine: A History of Formation and Current State. Indian Journal of Ecology 47 (4), 2020, S. 1077–1083
- Marina V. Rutovskaya, Maria V. Onufrenya und Alexander S. Onufrenya: Russian desman (Desmana moschata: Talpidae) at the edge of disappearance. Nature Conservation Research 2 (Suppl. 1), 2017, S. 100–112, doi:10.24189/ncr.2017.020
- S. V. Bakka, N. Yu. Kiseleva, I. I. Pankratov, I. A. Tarasov und P. M. Shuko: The story of the creation and monitoring of the Russian Desman (Desmana moschata L.) population reintroduced of in the Kerzhenets river floodplain in the Nizhny Novgorod region. IOP Conference Series: Earth and Environmental Science 115, 2018, S. 012036, doi:10.1088/1755-1315/115/1/012036
- Marina V. Rutovskaya, Alexander N. Aleksandrov, Valentina N. Podshivalina, Alena S. Soboleva und Oleg V. Glushenkov: Habitat conditions of Desmana moschata (Talpidae, Eulipotyphla, Mammalia) in the buffer zone of the Prisurskiy State Nature Reserve (Russia). Nature Conservation Research 5 (2), 2020, S. 36–46, doi:10.24189/ncr.2020.011
- К. А. Еськова, К. И. Беловежец, А. А. Косинский, Ю. О. Морева, И. А. Попов und М. В. Рутовская: Температурный режим мест обитания Русской выхухоли (Desmana moschata, Talpidae, Soricomorpha). Поволжский Экологический Журнал 1, 2018, S. 16–25
- Alexey Andreychev, Vyacheslav Kuznetsov und Alexandr Lapshin: Distribution and population density of the Russian desman (Desmana moschata L., Talpidae, Insectivora) in the Middle Volga of Russia. Forestry Studies 71, 2019, S. 48–68
- Yu. F. Ivlev, A. N. Kuznetsov und M. V. Rutovskaya: Preliminary Data on the Swimming Kinematics of the Russian Desman (Desmana moschata L.). Doklady Biological Sciences 431, 2010, S. 144–148
- A. I. Queiroz und V. Almada: Some behaviour patterns of Russian Desman (Desmana moschata L.), observed in captivity. Council of Europe (Hrsg.): Seminar on the biology and conseration of European desmans and water shrews (Galemys payrenaicus, Desmana moschata, Neomys spp.) Ordesa, Spain, 7-11 June 1995. Strasbourg, 1995, S. 48–54
- Yu. F. Ivlev, M. V. Rutovskaya und O. S. Luchkina: The Use of Olfaction by the Russian Desman (Desmana moschata L.) during Underwater Swimming. Doklady Biological Sciences 452, 2013, S. 280–283
- M. V. Rutovskaya: The behavior of Russian Desman (Desmana moschata). Council of Europe (Hrsg.): Seminar on the biology and conseration of European desmans and water shrews (Galemys payrenaicus, Desmana moschata, Neomys spp.) Ordesa, Spain, 7-11 June 1995. Strasbourg, 1995, S. 55–57
- Martyn L. Gorman und R. David Stone: The Natural History of Moles. London, 1990, S. 1–134 (S. 113–125)
- Peter Romanow, Walter Poduschka und Werner Deutsch: Zur intraspezifischen Lautkommunikation des Russische nDesmans Desmana moschata (Linnaeus, 1758) (Insectivora: Talpidae: Desmaninae) nebst einigen Angaben zu seinem Sozialverhalten. Contributions to Zoology 66 (1), 1996, S. 43–54
- Alexey Andreychev, ycheslav Kuznetsov, Alexandr Lapshin und Maksim Alpeev: Activity of the Russian desman Desmana moschata (Talpidae, Insectivora) in its burrow. Therya 11 (2), 2020, S. 161–167, doi:10.12933/therya-20-801 ISSN 2007-3364
- D. Jánossy und E. Schmidt: Die Nahrung des Uhus (Bubo bubo) Regionale und erdzeitliche Änderungen. Bonner Zoologische Beiträge 21, 1970, S. 25–51
- I. A. Solonitzine: Desmana moschata Brandt, hôte intermédiaire d’un Porrocaecum. Annales de Parasitologie 11 (3), 1933, S. 185–187
- A. Fain und J. O. Whitaker, Jr.: Two new species of Myobiidae from North American mammals (Acarina). Bulletin et Annales de la Société entomologique de Belgique 111, 1975, S. 57–65
- Kai He, Akio Shinohara, Kristofer M. Helgen, Mark S. Springer, Xue-Long Jiang und Kevin L. Campbell: Talpid Mole Phylogeny Unites Shrew Moles and Illuminates Overlooked Cryptic Species Diversity. Molecular Biology and Evolution 34 (1), 2016, S. 78–87
- Leigh van Valen: New Paleocene Insectivores and Insectivore classification. Bulletin of the American Museum of Natural History 135, 1967, S. 217–284
- J. Howard Hutchison: Notes on type specimens of European Miocene Talpidae and a tentative Classification of Old World Tertiary Talpidae (Insectivora: Mammalia). Geobios 7 (3), 1974, S. 211–256
- María Teresa Cabria, Jonathan Rubines, Benjamín Gómez-Moliner und Rafael Zardoya: On the phylogenetic position of a rare Iberian endemic mammal, the Pyrenean desman (Galemys pyrenaicus). Gene 375, 2006, S. 1–13, doi:10.1016/j.gene.2006.01.038
- Javier Igea, Pere Aymerich, Angel Fernández-González, Jorge González-Esteban, Asunción Gómez, Rocío Alonso, Joaquim Gosálbez und Jose Castresana: Phylogeography and postglacial expansion of the endangered semi-aquatic mammal Galemys pyrenaicus. BMC Evolutionary Biology 13, 2013, S. 115, doi:10.1186/1471-2148-13-115
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. A taxonomic and geographic Reference. 2 Bände. 3. Auflage. Johns Hopkins University Press, Baltimore MD 2005, ISBN 0-8018-8221-4 ()
- Antje Schreuder: A revision of the fossil water-moles (Desmaninae). Archives Néerlandais de Zoologie 4, 1940, S. 201–233
- Wolf-Dieter Heinrich und Norbert Hermsdorf: Erster Nachweis eines Desmans, Desmana moschata (Linnaeus, 1758) (Mammalia: Insectivora), im Pleistozän von Brandenburg. Brandenburger geowissenschaftliche Beiträge 12 (1/2), 2005, S. 55–60
- Cornelia G. Rümke: A review of fossil and recent Desmaninae (Talpidae, Insectivora). Utrecht Micropaleontological Bulletins, Special Publication 4, 1985, S. 1–241
- Raef Minwer-Barakat, Antonio García-Alix, Elvira Martín-Suárez und Matthijs Freudenthal: Early Pliocene Desmaninae (Mammalia, Talpidae) from Southern Spain and the Origin of the Genus Desmana. Journal of Vertebrate Paleontology 40 (5), 2020, S. e1835936, doi:10.1080/02724634.2020.1835936
- Barbara Rzebik-Kowalska und Leonid I. Rekovets: Recapitulation of data on Ukrainian fossil insectivore mammals (Eulipotyphla, Insectivora, Mammalia). Acta zoologica cracoviensia 58 (2), 2015, S. 137–171, doi:10.3409/azc.58_2.137
- Barbara Rzebik-Kowalska und Leonid I. Rekovets: New data on Eulipotyphla (Insectivora, Mammalia) from the Late Miocene to the Middle Pleistocene of Ukraine. Palaeontologia Electronica 19 (1), 2016, S. 9A ()
- Charles de l’Écluse: Exoticorum libri decem: quibus animalium, plantarum, aromatum, aliorumque peregrin. fructuum historiae describuntur / item Pt. Bellonii Observationes, eodem C.C. interprete. Leiden 1605, S. 1–378 (S. 375) ()
- Carl von Linné: Systema naturae. 10. Auflage, 1758, Band 1, S. 59 ()
- Carl von Linné: Fauna Svecica, sistens animalia Sveciae regni: quadrupedia, aves, amphibia, pisces, insecta, vermes, distributa per classes & ordines, genera & species, cum differentiis specierum, synonymis autorum, nominibus incolarum, locis habitationum, descriptionibus insectorum. Leiden, 1746, S. 1–411 (S. 9) ()
- Peter Simon Pallas: Reise durch verschiedene Provinzen des Rußischen Reichs. Erster Theil. St. Petersburg, 1771, S. 156 ()
- Johann Christian Daniel Schreber: Die Säugthiere in Abbildungen nach der Natur, mit Beschreibungen. Dritter Theil. Erlangen, 1778, S. 1–590 (S. 567–571) ()
- Georges-Louis Leclerc de Buffon: Histoire naturelle, générale et particulière. Tome Dixième. Paris, 1763, S. 1–20 () und Tafel 2 ()
- Johann Anton Güldenstädt: Beschreibung des Desmans. Beschäftigungen der Berlinischen Gesellschaft Naturforschender Freunde 3, 1777, S. 107–137 ()
- Johann Friedrich von Brandt: Bemerkungen über den inneren Bau des Wuychuchol (Myogale moschaqta), im Vergleich zu dem des Maulwurfs und der Spitzmaus (Sorex araneus). Archiv für Naturgeschichte 2 (1), 1836, S. 176–182 ()
- Georges Cuvier: Leçons d’anatomie comparée. Tome 1. Paris, 1800, S. 1–521 ()
- Anders Jahan Retzius: Animadversiones in classem mammalium Linneanam. Lund, 1796, S. 1–22 (S. 22) ()
- Pierre André Latreille: Histoire naturelle, générale et particulière des Crustacés et des Insectes. Tome Troisième. Paris, 1802, S. 1–467 (S. 48–59) ()
- George Edward Dobson: A Monograph of the Insectivora, systematic and anatomical. Part II. London, 1883, S. 126–172 ()
- Gernot Rabeder: Die Wirbeltierfauna aus dem Alt-Pliozän (O-Pannon) vom Eichkogel bei Mödling (NÖ.) I. Allgemeines — II. Insectivora. Annalen des Naturhistorischen Museums in Wien 74, 1970, S. 589–595
- Reinhard Ziegler: Insectivores (Lipotyphla) and bats (Chiroptera) from the Late Miocene of Austria. Annalen des Naturhistorischen Museums Wien 107A, 2006, S. 93–196
- Andrzej Sulimski: Pliocene insectivores from Węże. Study on the Tertiary bone breccia fauna from Węże near Dzialoszyn in Poland. Part XI. Acta Palaeontologica Polonica 4 (2), 1959, S. 119–173
- Andrzej Sulimski: Supplementary studies on the insectivores from Węże 1 (Poland). Study on the Tertiary bone breccia fauna from Węże near Dzialoszyn in Poland. Part xVII. Acta Palaeontologica Polonica 7 (3/4), 1962, S. 441–498
- Lawrence J. Flynn und Wenyu Wu: Two new shrews from the Pliocene of Yushe Basin, Shanxi Province, China. Vertebrata Palasiatica 32 (2), 1994, S. 73–86
- Lawrence J. Flynn und Wen-Yu Wu: The Lipotyphla of Yushe Basin. In: Lawrence J. Flynn und Wen-Yu Wu (Hrsg.): Late Cenozoic Yushe Basin, Shanxi Province, China: Geology and Fossil Mammals, Volume II: Small Mammal Fossils of Yushe Basin, Vertebrate Paleobiology and Paleoanthropology. Springer Science+Business Media B. V., 2017, S. 11–26
- Theodor Kormos: Trois nouvelles espèces fossiles des Desmans en Hongrie. Annales Musei Nationalis Hungarici 11, 1913, S. 135–145
- Martin Sabol, Vlastimil Konečný, Dionýz Vass, Marianna Kováčová, Anna Ďurišová und Igor Túnyi: Early Late Pliocene site of Hajnáčka I (Southern Slovakia) – geology, palaeovolcanic evolution, fossil assemblages and palaeoenvironment. Courier Forschungsinstitut Senckenberg 256, 2006, S. 261–274
- Jordi Agustí, Hugues-Alexandre Blain, Marc Furió, Roger De Marfá und Andrés Santos-Cubedo: The early Pleistocene small vertebrate succession from the Orce region (Guadix-Baza Basin, SE Spain) and its bearing on the first human occupationof Europe. Quaternary International 223-224, 2010, S. 162–169, doi:10.1016/j.quaint.2009.12.011
- Theodor Kormos: Desmana thermalis, n. sp., eine neue präglaziale Bisamspitzmaus aus Ungarn. Annales Musei Nationalis Hungarici 27, 1930, S. 1–19
- Erich Thenius: Fischotter und Bisamspitzmaus aus dem Altquartär von Hundsheim in Niederösterreich. Sitzungsberichte der Akademie der Wissenschaften mathematisch-naturwissenschaftliche Klasse 157, 1948, S. 187–145
- Hartmut Thieme, Dietrich Mania, Brigitte Urban und Thijs van Kolfschoten: Schöningen (Nordharzvorland) – Eine altpaläolithische Fundstelle aus dem mittleren Eiszeitalter. Archäologisches Korrespondenzblatt 23, 1993, S. 147–163
- Thijs van Kolfschoten: Die Kleinsäugerreste aus dem Reinsdorf-Interglazial von Schöningen. In: Hartmut Thieme (Hrsg.): Die Schöninger Speere. Mensch und Jagd vor 400.000 Jahren. Stuttgart 2007, S. 112–117
- Wighart von Koenigswald und Heinz Tobien: Bemerkungen zur Altersstellung der plaistozänene MosbachSande bei Wiesbaden. Geologisches Jahrbuch 115, 1987, S. 227–237
- Wim J Kuijper: Investigation of inorganic, botanical, and zoological remains of exposure of Last Interglacial (Eemian) sediments at Neumark-Nord 2 (Germany). In: Sabine Gaudzinski-Windheuser und Wil Roebroeks (Hrsg.): Multidisciplinary studies of the Middle Palaeolithic record from Neumark-Nord (Germany). Veröffentlichungen des Landesmuseums für Vorgeschichte Halle 69. Halle/Saale, 2014, S. 79–95
- Kim Aaris-Sørensen: Palaeoecoloy of a Late Weichselian vertebrate fauna from Nørre Lyngby, Denmark. Boreas 24, 1995, S. 355–365
- Barbara Rzebik-Kowalska: Erinaceomorpha and Soricomorpha (Mammalia) from the Late Pleistocene and Holocene of Krucza Ska³a Rock Shelter and Komarowa Cave (Poland). Acta zoologica cracoviensia 49A (1-2), 2006, S. 83–118
- N. G. Smirnov und D. V. Ponomarev: News about the padt distribution of the desman (Desmana moschata L.). Doklady Biological Sciences 414, 2007, S. 219–220, doi:10.1134/S0012496607030143
- Dmitry Ponomarev, Thijs van Kolfschoten, Johannes van der Plicht und Pavel Kosintsev: Lateglacial desman discovered in Sed’yu-1 (Komi Republic, Russia), a site in the far northeast of Europe. Quaternary International 378, 2015, S. 88–98, doi:10.1016/j.quaint.2014.03.001
- V. G. Skoptsov: The Desman and the Musk-rat. Oryx 9 (1), 1967, S. 54–56
- R. Kennerley und S. T. Turvey: Desmana moschata. The IUCN Red List of Threatened Species 2016. e.T6506A22321477 (); zuletzt aufgerufen am 30. November 2022
{kind=link}
Weblinks
- Desmana moschata in der Roten Liste gefährdeter Arten der IUCN 2016. Eingestellt von: R. Kennerley & S. T. Turvey, 2016. Abgerufen am 30. November 2022.