River Continuum Concept
Das River Continuum Concept ist ein Modell zur Einteilung und Beschreibung von Fließgewässern, das neben der Klassifizierung einzelner Gewässerabschnitte nach dem Vorkommen von Leitorganismen auch die Geomorphologie nutzt. In der Limnologie werden Modellbeschreibungen und Klassifikationsmodelle wie dieses benutzt, um den Zustand eines Gewässers zu beschreiben und zu beurteilen. Es bietet über die bis zu seiner Einführung übliche reine Einordnung von Lebensgemeinschaften hinaus auch eine Erklärung für deren Abfolge im Verlauf des Gewässers sowie über weitere voraussagbare und tatsächlich beobachtbare biologische Eigenschaften des Gewässers. Entwickelt wurde das Konzept 1980 durch eine amerikanische Forschungsgruppe um Robin L. Vannote.

Hintergrund
Das River Continuum Concept basiert auf der Idee, nach der ein Fließgewässer als offenes Ökosystem betrachtet wird, das in dauernder Interaktion mit dem Ufer steht und sich im Verlauf von der Quelle bis zur Mündung stetig verändert. Grundlage für diese Veränderung des Gesamtsystems ist eine graduelle Änderung physikalischer Umweltbedingungen wie beispielsweise der Breite, der Tiefe, der Wassermenge, der Strömungseigenschaften, der Temperatur sowie der Komplexität des Gewässers. Nach der Hypothese sind die Lebewesen einem solchen kontinuierlichen System angepasst und bilden ihrerseits ein Kontinuum, bei denen die Lebensgemeinschaften über längere Flussbereiche mit den physikalischen Gegebenheiten in Einklang stehen und sich ein Gleichgewicht zwischen Produzenten organischen Materials (primär Pflanzen) und Konsumenten desselben einstellt. Entlang des Flussverlaufs kommt es zu einer Veränderung im Verhältnis zwischen der Produktion und dem Verbrauch (Respiration) des Materials.
Lebensgemeinschaften und Ernährungstypen
Die kontinuierlichen Veränderungen der Eigenschaften innerhalb des Fließgewässers bedingen vor allem eine spezifische Zusammensetzung der Organismen in den unterschiedlichen Abschnitten des Gewässers. Wichtig ist dabei der Anteil der vier wichtigen Ernährungstypen, die als Zerkleinerer, Sammler, Weidegänger (bzw. engl. "Grazer") und Räuber bezeichnet werden. Mit Ausnahme der Räuber ernähren sich alle diese Organismen direkt von pflanzlichem Material und zersetzen dieses dadurch. Sie werden Saprobier genannt.

Die Zerkleinerer sind Organismen, die grobes organisches Material wie Laub in Teile zerkleinern. Sie nutzen Material mit Größen über einem Millimeter (Grobpartikuläres Material, CPOM = coarse particulate organic matter) sowie dessen Aufwuchs (Pilze, Mikroorganismen) als Nahrung, lassen zugleich aber auch viel Material zurück. Typische Zerkleinerer mitteleuropäischer Gewässer sind Flohkrebse, Wasserasseln, verschiedene Eintagsfliegen- und Steinfliegenlarven.
Als Sammler werden Organismen bezeichnet, die durch Reusen- oder andere Sammelstrukturen in der Lage sind, Schwebstoffe aus dem Wasserstrom zu fischen oder vom Sediment aufzunehmen, von denen sie sich ernähren. Die Partikelgrößen liegen dabei zwischen 0,5 und 50 Mikrometer (Ultrafeinpartikuläres organisches Material, UPOM = ultrafine particulate organic matter, bis Feinpartikuläres Material, FPOM = fine particulate organic matter). Zu dieser Gruppe gehören die Larven der Kriebelmücken, viele Zuckmückenlarven, Fadenwürmer und viele weitere Tiergruppen.
Die Weidegänger grasen den Aufwuchs größerer Strukturen im Wasser ab, etwa von Steinen, Holzstücken oder größeren Wasserpflanzen. Hierzu gehören etwa Schnecken, Lidmückenlarven und ebenfalls viele Zuckmückenlarven.
.jpg.webp)
Räuber sind schließlich Organismen, die sich von anderen Tieren ernähren, darunter etwa die Libellenlarven sowie verschiedene Käfer.
Aufgrund der Veränderung des vorhandenen organischen Materials ändern sich im Flussverlauf die Anteile dieser Gruppen an den Lebensgemeinschaften. Im Oberlauf und der Bachregion gibt es aufgrund der groben Pflanzenstoffe ein Übergewicht an Zerkleinerern und auch an Sammlern, Weidegänger machen einen kleinen Anteil aus. Im Mittellauf erhöht sich der Anteil der Weidegänger und vor allem der Sammler, während die Zerkleinerer deutlich zurückgehen und schließlich vollständig verschwinden. Im Unterlauf gibt es fast nur noch partikuläres Material, wodurch die Sammler den größten Teil ausmachen. Der Anteil der Räuber bleibt in allen Abschnitten weitgehend konstant und ändert sich nur in der Artenzusammensetzung, da diese nicht von der Größe des organischen Materials abhängig sind, sondern von der Verfügbarkeit von Beutetieren. Atypische Änderungen in der Zusammensetzung dieser Organismengruppen innerhalb des Verlaufs eines Fließgewässers, etwa eine erhöhte Anzahl von Zerkleinerern in größeren Flussbereichen oder ein Fehlen dieser Tiere im Oberlauf, lassen auf eine Störung schließen.
Die Einteilung des Fließgewässers

Die Einteilung des Fließgewässers nach dem River Continuum Concept lässt eine grobe Zuordnung in drei Abschnitte zu, die auf alle Fließgewässer anwendbar ist. Demnach lassen sich Kleingewässer als Bäche gegenüber mittleren Flüssen und großen Flüssen abgrenzen.
Der Bachbereich ist im Oberlauf sehr schmal und meistens von einer starken Ufervegetation gesäumt. Diese verhindert das Durchdringen von Sonnenlicht und damit die Produktion von organischem Material mittels Photosynthese in dem Gewässer, zugleich liefert sie jedoch große Mengen an pflanzlichem Material, das in den Fluss fällt (allochthones organisches Material). In diesem Abschnitt überwiegt also die Respiration gegenüber der Produktion und das Verhältnis beträgt P:R<1. Hier spielen Lebewesen eine große Rolle, die das meist grobe pflanzliche Material zerkleinern, sowie Organismen, die zerkleinertes Material sammeln und nutzen. Hinzu kommen Weidegänger und Räuber. In diesem Bereich ist außerdem die größte Diversität an organischem Material zu erwarten. Vollständig abgebaut werden hier allerdings nur Kleinstpartikel, größere und schwerer abbaubare Bestandteile driften weiter flussabwärts.

Im weiteren Verlauf des Gewässers nimmt die Bedeutung des Ufers als Lieferant organischen Materials zunehmend ab, und die Produktion innerhalb des Gewässers wird wichtiger (autochthones organisches Material). Das Verhältnis der Produktion zur Respiration wird größer und beträgt P:R>1. Der Anteil an Zerkleinerern nimmt entsprechend ab, da das pflanzliche Material hier in Form von Algen vorliegt. Größer wird der Anteil der Sammler und auch der Weidegänger, der Räuberanteil bleibt unverändert.
Im letzten Abschnitt liegt im Fluss sehr viel partikuläres Material vor, außerdem findet weitere Produktion durch Photosynthese statt, die jedoch durch trübende Schwebstoffe auf den obersten Bereich des Gewässers beschränkt wird. Hier überwiegt wieder die Respiration, und das Verhältnis beträgt wiederum P:R<1. Die Lebensgemeinschaft besteht in diesen Flussbereichen fast ausschließlich aus Sammlern sowie einem weiterhin unveränderten Anteil an Räubern.
Das Kontinuum
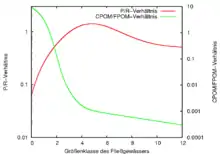
Die kontinuierlichen Veränderungen über die Strecke des Gewässers lassen sich an verschiedenen Faktoren belegen. Wie oben beschrieben beginnt der Fluss mit einem anfangs sehr stark von außen beeinflussten System, in dem vor allem organisches Material verbraucht wird. Danach geht er über in ein System mit starker Eigenproduktion organischen Materials, das tagesperiodisch abhängig vom Sonnenlicht variiert. Der letzte Bereich ist zwar wenig abhängig von außen, aber trotzdem sehr stark von Abbauprozessen geprägt. In einem durchgehenden System ohne Störungen, etwa durch Zuflüsse, kann diese Entwicklung bei allen Flusssystemen betrachtet werden, wobei Variationen durch jahreszeitliche Rhythmik der Umweltfaktoren (vor allem der Temperatur) möglich sind. Die Partikelgröße des organischen Materials sowie seine Diversität nehmen zudem im Laufe des Flusses immer weiter ab.
Ressourcen und Stabilität des Systems
Einen wesentlichen Punkt des Konzepts macht die Betrachtung der Ressourcennutzung des organischen Materials und der darin enthaltenen Energie im Fließgewässer aus. An jeder Stelle im Fluss wird Energie eingebracht in Form von organischem Material, sie wird genutzt, gespeichert und teilweise an flussabwärts gelegene Stellen weitergegeben. Die vorhandene Energie stellt zugleich den limitierenden Faktor des Systems dar, und das System ist bestrebt, diesen möglichst effizient und verlustfrei zu nutzen. Freie Ressourcen ermöglichen es neuen Arten, sich in der Lebensgemeinschaft zu etablieren, so dass diese im Modell auch schnell wieder genutzt werden. Dieses Prinzip ist nicht nur für das Ökosystem „Fluss“ wichtig, sondern gilt für jedes andere System gleichermaßen. Hier spielt es jedoch eine größere Rolle, da nicht verbrauchte Ressourcen an einer Stelle durch den ständigen Weitertransport verlorengehen. Nach dem River Continuum Concept wird entsprechend postuliert, dass in einem Fluss ein konstanter starker Zwang zur optimalen Ressourcennutzung und zu einer zeitlich gleichmäßigen kontinuierlichen Nutzung besteht.
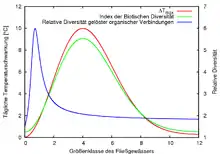
Der zeitliche Aspekt dieser Kontinuität lässt sich vor allem an der Tagesperiodik und der jahreszeitlichen Periodik erkennen. Im Verlauf des Tages wechseln die Lebensgemeinschaften vor allem aufgrund des erhöhten Fraßdrucks am Tag (Fische jagen vor allem optisch) und der Änderungen abiotischer Faktoren wie der Temperatur und dem Licht. Dabei ist die größte tagesperiodische Veränderung der Faktoren im Mittellauf festzustellen, hier existiert auch das größte Spektrum verschiedener Tierarten (Artendiversität), die die unterschiedlichen Bedingungen optimal nutzen können.
Aufgrund der, durch diese gleichmäßige Nutzung der Ressourcen, vorhandenen Konstanz ist eine sehr hohe Stabilität des Systems bei Störungen und Schwankungen bedingt. Ungleichmäßigkeiten in der Nutzung werden entsprechend sehr schnell ausgeglichen, und es stellt sich relativ bald danach ein neues Gleichgewicht ein. Zudem gibt es keine ökologische Entwicklung des Systems (Sukzession), und Veränderungen des Systems sind nur von außen durch geologische Veränderungen wie eine Veränderung des Einzugsgebietes, Veränderung des organischen Inputs oder geologischer Erdbewegung möglich. Auch nach diesen Veränderungen kommt es jedoch wieder zu einem Fließgleichgewicht und einem veränderten, aber optimal funktionierenden Flusssystem.
Entwicklung und Anwendung des Konzepts
Die erste Vorstellung des umfassenden Konzeptes fand 1980 im Rahmen eines zweitägigen Kongresses des Stroud Water Research Center statt, dessen Leiter Robin Vannote war. Es war das Ergebnis einer mehrjährigen Studie, die von der Rockefeller-Stiftung bezahlt wurde. Die Veröffentlichung der Hypothese erfolgte im selben Jahr unter dem Titel „The River Continuum Concept“ in der Zeitschrift Canadian Journal of Fisheries and Aquatic Sciences. Das Konzept baute auf Arbeiten weiterer amerikanischer Limnologen auf, wie Ruth Patrick, die den ökosystemaren Aspekt des Fließgewässers herausgestellt hat, sowie Luna Leopold, der sich mit den physikalischen Veränderungen des Gewässers auseinandersetzte. Die wesentliche Ergänzung der Arbeitsgruppe von Vannote war die Ergänzung und Erklärung der Lebensgemeinschaften in diesem System. Vannote selbst schilderte die Ausgangslage wie folgt:
- „In those days, most people studied a square meter of water to death.“[1]
(Deutsch: „Damals analysierten die meisten Leute einen Quadratmeter Wasser zu Tode.“)
Die Forschung erfolgte also immer nur an kleinen Teilstücken der Gewässer, und diese wurden nur ausgesprochen selten in ihrer Gesamtheit als Fließgewässer betrachtet.
Nach ihrer Veröffentlichung fand das River Continuum Concept sehr schnell die Akzeptanz der Fachwelt und wurde zum favorisierten Modell zur Beschreibung der Lebensgemeinschaften in Fließgewässern. Hier löste es die klassische Gliederung der Gewässer in Fischregionen, die von Robert Lauterborn zwischen 1916 und 1918 anhand der Lebensgemeinschaften im Rhein entwickelt wurde, sowie die Einteilung in die Lebensräume Krenal, Rhithral und Potamal von Joachim Illies in seiner Veröffentlichung „Versuch einer allgemeinen biozönotischen Gliederung der Fließgewässer“ (1961) ab. Beide Konzepte hatten den Nachteil, dass sie immer nur Zonen des Gewässers beschrieben und keine Betrachtung des Systems in seiner Gesamtheit ermöglichten, wie dies durch das River Continuum Concept ermöglicht wird.
In der Praxis wird das River Continuum Concept heute vor allem zur ökologischen Beurteilung von Fließgewässern und deren Störungen eingesetzt. So kann nach einer Untersuchung der Biozönosen in einem Flusslauf die Artenzusammensetzung bestimmt und mit dem Idealfall nach den River Continuum Concept verglichen werden. Vor allem ein Übergewicht oder ein Fehlen von Ernährungstypen kann Aufschluss über eine eventuell vorhandene Störung geben.
Probleme, Grenzen und Modifikationen

Obwohl das River Continuum Concept auf eine breite Akzeptanz gestoßen ist, ist es in seiner Anwendbarkeit begrenzt. Es beschreibt als Modell ein optimales und sich gleichmäßig veränderndes Fließgewässer ohne Störungen und Unregelmäßigkeiten. Nicht enthalten sind dabei etwa Zuströme, Stauungen durch Dämme oder in Seen oder auch unregelmäßige Ereignisse wie Überflutungen des Ufers.
Um auch diese Unregelmäßigkeiten im Modell zu erfassen wurde das River Continuum Concept von verschiedenen Autoren ausgebaut und variiert. So entwickelten beispielsweise J. V. Ward und J. A. Stanford 1983 das Serial Discontinuity Concept, in dem sie die Auswirkungen von geomorphologischen Störungen wie die angesprochenen Stauungen und Zuflüsse integrierten. Dieselben Autoren stellten 1993 das Hyporheic Corridor Concept vor, in dem auch die vertikale (in die Tiefe) und laterale (von Ufer zu Ufer) Strukturkomplexität mit dem Kontinuum des Flusses verbunden wurde. Das Flood Pulse Concept, entwickelt von W. J. Junk et al. 1989 und weiter modifiziert von P. B. Bayley 1990 und K. Tockner et al. 2000, brachte schließlich auch noch ein, dass ein großer Teil der Nährstoffe und des organischen Materials aus dem Sediment und überfluteten Landschaftsteilen in den Fluss kommt. Vor allem die Produktivität sehr großer Flüsse mit großen Überflutungsflächen schildert das 1994 von J. H. Thorp und M. D. Delong entwickelte Riverine Productivity Model (RPM).
Quellenangaben
- River Continuum (Memento vom 28. Oktober 2008 im Internet Archive)
Literatur
River Continuum Concept
- R. L. Vannote, G. W. Minshall, K. W. Cummins, J. R. Sedell und C. E. Cushing: The River Continuum Concept. (PDF; 956 kB) In: Canadian Journal of Fisheries and Aquatic Sciences. Band 37, Ottawa 1980, Nr. 1, S. 130–137. ISSN 0706-652X
- Winfried Lampert und Ulrich Sommer: Limnoökologie. Georg Thieme, Stuttgart 1993, 1999. ISBN 3-13-786402-X
- D. N. Gordon, T. A. McMahon, B. L. Finlayson, C. J. Gippel und R. J. Nathan: Stream Hydrology - An Introduction for Ecologists. John Wiley & Sons, Chichester W Suss 2004. ISBN 0-470-84357-8
Weiterführende Konzepte
- J. W. Junk, P. B. Bayley und R. E. Sparks: The flood pulse concept in river floodplain systems. In: Canadian Special Publications of Fisheries and Aquatic Sciences. Band 106, 1989, S. 110–127. ISSN 0706-652X
- J. V. Ward und J. A. Stanford: The serial discontinuity concept of river ecosystems. In: T. D. Fontaine und S. M. Bartell (Hrsg.): Dynamics of lotic ecosystems. Science Publications, Ann Arbor Mich 1983, S. 29–42. ISBN 0-250-40612-8
- J. A. Stanford und J. V. Ward: An ecosystem perspective of alluvial rivers, connectivity and the hyporheic corridor. In: Journal of the North American Benthological Society. Band 12, Allen, Lawrence Kan 1993, S. 48–60. ISSN 0887-3593
- P. B. Bayley: The flood pulse advantage and the restoration of river -floodplain-systems. In: Regulated Rivers. Research & Management. Band 6, 1990, S. 75–86. ISSN 0886-9375
- K. Tockner, F. Malard und J. V. Ward: An extension of the flood pulse concept. In: Hydrological Processes. Band 14, Whiley, Chichester 2000, S. 2861–2883. ISSN 0885-6087
- J. H. Thorp und M. D. Delong: The riverine productivity model: an heuristic view of carbon sources and organic processing in large river ecosystems. In: Oikos. Band 70, Blackwell, Oxford 1994, Nr. 2, S. 305–308. ISSN 0030-1299