Rotalgen
Die Rotalgen (Rhodophyta,[1] Synonym: Rhodophyceae) sind eine Abteilung von Algen, von denen viele durch die an der Photosynthese beteiligten Phycobiline rot gefärbt sind. Sie bilden eine der drei Gruppen der Archaeplastida. Ihre Fortpflanzung ist durch einen dreigliedrigen Generationswechsel gekennzeichnet. Alle Formen und Stadien sind unbegeißelt. Rotalgen kommen in der Mehrzahl in der Litoralzone des Meeres vor, einige Arten auch im Süßwasser und in feuchtem Erdreich. Fossil sind sie seit dem Erdzeitalter des Ectasiums (vor etwa 1400 bis 1200 Millionen Jahren) bekannt.[2]
| Rotalgen | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|

Asparagopsis taxiformis | ||||||||||
| Systematik | ||||||||||
| ||||||||||
| Wissenschaftlicher Name | ||||||||||
| Rhodophyta | ||||||||||
| Wettstein |
.jpg.webp)

Merkmale
Rotalgen sind zumeist mehrzellig und bestehen aus Zellfäden, die zu einem Plektenchym (Scheingewebe) aneinandergelagert sein und bandförmige oder lappige Formen ausbilden können.[3] Die Zellwand besteht aus Cellulose und einem Schleim aus Galactanen, zumeist Agar oder Carrageen. Einzellige Arten sind nur von einer Schleimschicht ohne Cellulose umgeben. Begeißelte Stadien fehlen im Unterschied zu anderen Algen vollständig.[4]
Bei vielzelligen Rotalgen befindet sich in der Mitte jeder Trennwand eine Struktur, die traditionell als „Tüpfel“ (engl. pit connection) bezeichnet wird, aber nicht wie die Tüpfel bei höheren Pflanzen eine Verbindung zwischen den Zellen herstellt, sondern im Gegenteil eine Pore, die bei der zentripetalen Bildung der Zellwand zunächst offen bleibt, verschließt.[5]
Die Farbe der Rotalgen variiert je nach den vorhandenen Photosynthese-Pigmenten. Arten der Litoral-Zone sind olivgrün bis rotbraun, weil sie neben Chlorophyll viel blaues Phycocyanin sowie Carotinoide enthalten. Arten tieferer Zonen enthalten viel rotes Phycoerythrin und sind rot bis violett gefärbt.[6]
Photosynthese und Plastiden
Die Chloroplasten, hier auch Rhodoplasten genannt, enthalten Chlorophyll a und seine Begleitcarotinoide, aber keine anderen Chlorophylle. In den Chloroplasten liegen die Thylakoide nicht wie bei den Chloroplastida in Stapeln vor, sondern sind in gleichen Abständen nebeneinander angeordnet.
Auf den Thylakoiden sitzen 30 bis 40 nm große, mehr oder weniger kugelige Strukturen: die Phycobilisomen. Sie enthalten wasserlösliche Photosynthese-Hilfspigmente, die Phycobiliproteine. Die Hauptmasse bilden dabei mehrere Varianten von rotem Phycoerythrin; Allophycocyanine und Phycocyanine kommen ebenfalls vor. Bau und Pigmentausstattung der Rhodoplasten deuten auf ihre Verwandtschaft mit den Cyanobakterien hin (siehe auch Endosymbiontentheorie).
Das Pigment Phycoerythrin ist photodestruktiv. Das bedeutet, dass dieses Pigment mit zunehmender Lichtintensität zunehmend zerstört wird. Dies ist der Grund dafür, dass viele oberflächennah lebende Rotalgen nicht rot aussehen. Mit Hilfe der Phycobiliproteine minimieren die Rotalgen die Grünlücke der grünen Pflanzen und können auch „mittelwellige VIS“ (Bereich der visuellen und gleichzeitig größtenteils auch photosynthetisch aktiven Strahlung) absorbieren und auf die photosynthetisch aktiven Chlorophyll a-Moleküle übertragen. Da Licht mit einer Wellenlänge von ca. 380 nm bis 550 nm (blau bis grün) schlecht von Wassermolekülen und ihren Soluten absorbiert wird, gelangt es in große Tiefen (über 250 m). Phycoerythrin verstärkt die Absorption vor allem von 490 nm bis 580 nm (vom hellblauen bis zum Anfang des gelben Bereiches).
Das primäre Produkt der Photosynthese ist hauptsächlich der Zuckeralkohol Floridosid. Der Gehalt an gelöstem Floridosid korreliert mit dem Salzgehalt des Wassers und kann bis zu 10 % der Trockenmasse der Alge ausmachen. Dies wirkt einem Wasserverlust durch Osmose entgegen.[7] Das Reservepolysaccharid ist die Florideenstärke, ein Polymer des Zuckeranteils des Floridosids. Sie wird nicht wie die Stärke der höheren Pflanzen in den Plastiden, sondern an der Oberfläche der Chloroplasten oder im Cytosol gespeichert. Chemisch ist sie näher mit dem tierischen Glykogen als mit pflanzlicher Stärke verwandt.
Fortpflanzung
Neben der ungeschlechtlichen Vermehrung durch Sporen weisen Rotalgen typischerweise einen dreigliedrigen Generationswechsel auf, bei dem nacheinander Gametophyten, Karposporophyten (oder Gonimoblasten) und Tetrasporophyten auftreten. Die Bildung eines Karposporophyten wird als Apomorphie der Rotalgen betrachtet.
Das weibliche Geschlechtsorgan, das einzellige Karpogon, bildet einen dünnen fädigen Fortsatz aus, die Trichogyne, die dem Kontakt mit den männlichen Gameten (Spermatien) dient. Das männliche Geschlechtsorgan, das Spermatangium, bringt nur ein kugelförmiges oder längliches Spermatium hervor, das von einer klebrigen Masse umgeben ist. Bei manchen Rotalgen hat diese Umhüllung faserige Fortsätze, welche die Chance erhöhen, Kontakt zu einer Trichogyne zu finden. Die Trichogyne wächst gewöhnlich so lange weiter, bis sich ein Spermatium an sie bindet. Die Spermatien sind – wie alle Zellen der Rotalgen – stets unbegeißelt und werden passiv durch die Wasserströmung bewegt.
Das Karpogon enthält zwei Zellkerne, von denen einer in der Trichogyne sitzt und später vergeht. Der andere, basale Zellkern verschmilzt bei der Befruchtung mit dem Kern eines Spermatiums, der in die Trichogyne entlassen wurde und durch diese hinabwandert. Aus der nun diploiden Zygote wächst der wenigzellige fädige Gonimoblast (auch Karposporophyt genannt) heraus. Dieser stellt wegen des erfolgten Wechsels der Kernphase eine neue Generation dar, bleibt aber mit der Mutterpflanze verbunden. Er setzt schließlich diploide Karposporen frei, aus denen die diploiden Tetrasporophyten als dritte Generation hervorgehen, welche äußerlich meist den Gametophyten der ersten Generation gleichen. In ihren Tetrasporangien erfolgt die Meiose, und es entstehen zwei weibliche und zwei männliche Tetrasporen. Mit den aus diesen hervorgehenden Gametophyten schließt sich der Lebenszyklus.[8][9]
Von diesem typischen Lebenszyklus gibt es viele Ausnahmen. So sind bei einzelligen Rotalgen wie den Cyanidiales und den Porphyridiales keine sexuellen Vorgänge bekannt, und die Bangiales zeigen einen zweigliedrigen Generationswechsel, bei dem die Zygote unmittelbar die Karposporen bildet, also kein Gonimoblast ausgebildet wird.
Vorkommen
Die Rotalgen leben ganz überwiegend im Meer, und sie sind unter den Meeresalgen mit mehr Arten repräsentiert als alle sonstigen Algengruppen zusammen. Allerdings sind sie weitgehend auf gemäßigte bis tropische Breiten beschränkt, während in kalten Meeresbereichen Braunalgen und Grünalgen überwiegen. Dank der Phycobiline können sie in größeren Wassertiefen Photosynthese betreiben als andere Algen.[10]
Rotalgen im Süßwasser
Im Süßwasser leben etwa 178 Arten von Rotalgen, das sind etwas mehr als drei Prozent der gesamten Artenzahl. Soweit bekannt, bevorzugen alle Arten Fließgewässer (oder zumindest bewegtes Wasser) und kommen am Boden (benthisch), meist aufwachsend auf Hartsubstraten, vor. Wie typisch für Rotalgen, werden relativ gering belichtete Habitate, zum Beispiel durch Huminstoffe getrübtes Wasser, bevorzugt. Einige Arten bevorzugen Sonderhabitate, so zum Beispiel Cyanidium und Galdieria in Thermalquellen in Italien.[11] Die meisten süßwasserlebenden Arten sind nicht rot gefärbt, stattdessen kommen gelbgrüne, blaugrüne, olivgrüne, violette, bräunliche bis zu grauschwarze Arten vor; meist ist das Phycoerythrin bei ihnen durch Phycocyanin überdeckt. Viele Süßwasser-Rotalgen sind empfindlich gegenüber Gewässerverschmutzung, einige gelten als bedroht. In Europa sind Süßwasser-Rotalgen in den nördlichen Breiten artenreicher als im Süden; die meisten Arten (28) sind in Finnland nachgewiesen, in Deutschland sind es 21, in Österreich 22 und in der Schweiz 12 Arten. Die häufigsten und am weitesten verbreiteten Gattungen sind Audouinella, Bangia, Batrachospermum, Hildenbrandia und Lemanea.[12]
Epiphyten und Parasiten

Manche Rotalgen wachsen als Epiphyten oder als Parasiten auf anderen Algen. Bei ersteren unterscheidet man fakultative, die auch auf toten Substraten wachsen können, und obligate Epiphyten, die nur auf einem passenden Wirt gedeihen können. Ein obligater Epiphyt ist die Pinselbüschelalge. Deren Sporen keimen auf der Oberfläche des Knotentangs (einer Braunalge). Sie bilden ein Rhizoid aus, das mittels Verdauungsenzymen in das Wirtsgewebe eindringt und den Epiphyten verankert. Parasitische Rotalgen nehmen im Unterschied zu bloßen Epiphyten Nährstoffe aus dem Wirt auf. Dabei können sie auch selbst noch Photosynthese betreiben oder als Vollparasiten keine eigenen Photosynthese-Pigmente mehr ausbilden und ganz auf die Ernährung durch den Wirt angewiesen sein. Ein interessantes Beispiel für einen Vollparasiten ist Choreocolax polysiphoniae, der die Pinselbüschelalge parasitiert, die wiederum als Epiphyt auf dem Knotentang wächst. Auch Choreocolax dringt in den Wirt ein, bildet aber Tüpfelverbindungen zu dessen Zellen aus, um Nährstoffe aufzunehmen. Deshalb spricht man hier nicht von einem Rhizoid, sondern von einem Haustorium.[13]
Nutzung
Menschlicher Verzehr

- In Ländern wie Kanada, Island, Irland und Frankreich ist der leicht nussig schmeckende Lappentang (Palmaria palmata, franz. dulce) als Nahrungsmittel weit verbreitet. In Irland wird er darüber hinaus als Bestandteil von Gin verwendet.[14]
- In Wales wird laverbread, eine Algenpaste aus gekochten und zerkleinerten Purpurtangen, frittiert verzehrt oder auch mit Orangensaft zu Hammel gereicht.
- In Ostasien werden Purpurtange der Gattungen Porphyra und Pyropia (japanisch 海苔 nori, chinesisch 海苔 haitai oder 紫菜 zicai, koreanisch 김 gim) in Suppen mitgekocht, in Gewürzmischungen (beispielsweise für Ramen) verwendet oder als ganze gepresste Blätter um Sushi oder Gimbap gewickelt.
- Knorpeltang (Chondrus crispus oder „Irisch Moos“) wird unter anderem für die Herstellung von Pudding, Milchgetränken, Hustentee und Husten-Lutschpastillen verwendet. Durch den hohen Anteil an Carrageen findet der Tang darüber hinaus bei der Herstellung von Kosmetika, Schuhcreme und Medizinprodukten Verwendung, wo er zur Stabilisierung von Suspensionen eingesetzt wird.[15]
- Aus verschiedenen Rotalgen lassen sich Agar (Agar-Agar) und Carrageen gewinnen. Diese werden nicht nur als Gelatineersatz für Nahrungsmittel verwendet, sondern auch für Kosmetik sowie für die experimentelle Biologie, z. B. als Nährmedium zur Kultivation von Bakterien.[16]
- Darüber hinaus werden Rotalgen als Nahrungsergänzungsmittel verwendet, da sie reich an Vitaminen und Calcium sowie Iod und weiteren Spurenelementen sind.[17][18] Die in Pflanzen eher selten vorkommende „physiologisch bedeutende“ (aber nicht generell essenzielle) Aminosäure Taurin findet sich bei verschiedenen essbaren und in der japanischen und koreanischen Küche verwendeten Rotalgen-Arten wie Gelidium subcostatum (Hirakusa), Grateloupia elliptica (Tanbanori), Gloiopeltis tenax (Manufori), Gloiopeltis furcata (Fukurofunori), Gracilaria textorii (Kabanori) und Gracilaria vermiculophylla (Ogonori).[19][20]
Landwirtschaftliche Nutzung
- Als Naturkalk wird Algenkalk als Hilfs- und Düngestoff für Landwirtschaft und Gartenbau, zur Kalkung von Böden und als Gegenmaßnahme bei Bodenversauerung verwendet. Algenkalk ist auch im Ökolandbau zugelassen, wo er unter anderem zur Steigerung der Erträge eingesetzt wird.[21]
- Rotalgen der Art Asparagopsis taxiformis als Futtermittelzusatz haben unter verschiedenen untersuchten Algen das größte Potential zur Verringerung des Methangas-Ausstoßes bei Rindern und Schafen.[22]
Damit der hohe Methan-Ausstoß bei der Produktion von Rindfleisch oder Milch in Zukunft gezielt gesenkt werden kann, werden Rotalgen durch Anbau in künstlich angelegten Tangwäldern gewonnen, z. B. vor der Küste Australiens.[23] oder durch Ernte natürlicher Vorkommen beispielsweise im Atlantik um die Azoren.[24] Auf der Nordhemisphäre ist die Zucht von Asparagopsis taxiformis problematisch, da sie dort als invasive Art gilt.[25] Geforscht wird unter anderem an der Universität Kiel, wo die Agrar- und Ernährungswissenschaftliche Fakultät in Zusammenarbeit mit Versuchsbetrieben, zukunftstaugliche Verbesserungen für die Rinderzucht auch hinsichtlich der Emissionen eruiert.[26] Im Jahr 2022 wurde in Kalifornien die Nutzung als Futtermittel-Zusatz genehmigt.[27]
Systematik









Die Rotalgen werden seit der Mitte des 19. Jahrhunderts als eigenes Taxon anerkannt. 1851 stellte Franz Joseph Ruprecht die Klasse Rhodophyceae auf. 1901 stufte Richard Wettstein die Rotalgen als Rhodophyta im höheren Rang einer Abteilung ein. Im 20. Jahrhundert wurden die Rhodophyta bzw. Rhodophyceae meist in zwei Hauptgruppen eingeteilt, die Florideophyceae (Florideen) und die Bangiophyceae. Neuere molekulargenetische Untersuchungen führten jedoch zu einem stärker unterteilten System.[28][29]
Adl et al. schlugen 2012 eine strikt phylogenetische Einteilung vor, die auf jegliche Rangstufen verzichtet:[30]
- Rhodophyceae Turet (emend. Rabenhorst) (Syn. Rhodophyta Wettstein, Rhodoplantae Saunders & Hommersand)
- Cyanidiales T.Christensen (Syn. Cyanidiophyceae Merola et al., Cyanidiophyta Moehn ex Doweld)
- Rhodellophyceae Cavallier-Smith (Syn. Rhodellophytina Cavallier-Smith)
- Stylonematales K. Drew (Syn. Stylonematophyceae H. S. Yoon et al.)
- Porphyridiophyceae H. S. Yoon et al.
- Compsopogonales Skuja (Syn. Compsopogonophyceae G. W. Saunders & Hommersand)
- Bangiales Nägeli (Syn. Bangiophyceae A. Wettstein)
- Florideophycidae Cronquist
- Nemaliophycidae Christensen
- Corallinophycidae L. Le Gall & G. W. Saunders
- Ahnfeltiophycidae G. W. Saunders & Hommersand
- Rhodymeniophycidae G. W. Saunders & Hommersand
Diese ranglose Klassifikation wurde jedoch von den meisten Phykologen nicht aufgegriffen. Im Syllabus of Plant Families von 2017 gliedern Kamiya et al. die Rotalgen wie folgt (Artenzahlen ergänzt nach AlgaeBase, Stand Juli 2018):[1][31]
Abteilung Rhodophyta, mit etwa 7130 Arten
- Unterabteilung Cyanidiophytina, mit etwa 7 Arten
- Klasse Cyanidiophyceae
- Ordnung Cyanidiales, mit etwa 7 Arten
- Klasse Cyanidiophyceae
- Unterabteilung Rhodophytina / Proteorhodophytina (die ersten vier Klassen wurden von Muñoz-Gómez 2017 als eigene Unterabteilung eingestuft),[32] mit etwa 140 Arten
- Klasse Porphyridiophyceae
- Ordnung Porphyridiales, mit etwa 15 Arten
- Klasse Stylonematophyceae, mit etwa 43 Arten
- Ordnung Rufusiales, mit nur einer Art
- Ordnung Stylonematales, mit etwa 42 Arten
- Klasse Compsopogonophyceae, mit etwa 76 Arten
- Ordnung Compsopogonales, mit etwa 11 Arten
- Ordnung Erythropeltidales, mit etwa 64 Arten
- Ordnung Rhodochaetales, mit nur einer Art
- Klasse Rhodellophyceae, mit etwa 6 Arten
- Ordnung Dixonelliales, mit 2 Arten
- Ordnung Glaucosphaerales, mit 3 Arten
- Ordnung Rhodellales, mit nur einer Art
- Klasse Porphyridiophyceae
- Unterabteilung Rhodophytina / Eurhodophytina, mit etwa 6950 Arten
- Klasse Bangiophyceae
- Ordnung Bangiales, mit etwa 186 Arten
- Klasse Florideophyceae, mit etwa 6771 Arten
- Unterklasse Hildenbrandiophycidae
- Ordnung Hildenbrandiales, mit etwa 19 Arten
- Unterklasse Nemaliophycidae, mit etwa 903 Arten
- Ordnung Acrochaetiales, mit etwa 235 Arten
- Ordnung Balbianiales, mit 3 Arten
- Ordnung Balliales, mit etwa 7 Arten
- Ordnung Batrachospermales, mit etwa 264 Arten (darunter die Froschlaichalgen)
- Ordnung Colaconematales, mit etwa 50 Arten
- Ordnung Entwisleiales, mit nur einer Art
- Ordnung Nemaliales, mit etwa 269 Arten
- Ordnung Palmariales, mit etwa 52 Arten
- Ordnung Rhodachlyales, mit etwa 2 Arten
- Ordnung Thoreales, mit etwa 20 Arten
- Unterklasse Corallinophycidae, mit etwa 798 Arten
- Ordnung Corallinales, mit etwa 611 Arten
- Ordnung Hapalidiales, mit etwa 137 Arten
- Ordnung Rhodogorgonales, mit 3 Arten
- Ordnung Sporolithales, mit etwa 42 Arten
- sowie einige Arten mit unsicherer Zuordnung
- Unterklasse Ahnfeltiophycidae, mit etwa 11 Arten
- Ordnung Ahnfeltiales, mit etwa 10 Arten
- Ordnung Pihiellales, mit nur einer Art
- Unterklasse Rhodymeniophycidae, mit etwa 5192 Arten
- Ordnung Acrosymphytales, mit etwa 16 Arten
- Ordnung Bonnemaisoniales, mit etwa 33 Arten
- Ordnung Ceramiales, mit etwa 2695 Arten
- Ordnung Gelidiales, mit etwa 234 Arten
- Ordnung Gigartinales, mit etwa 962 Arten
- Ordnung Gracilariales, mit etwa 238 Arten
- Ordnung Halymeniales, mit etwa 330 Arten
- Ordnung Nemastomatales, mit etwa 63 Arten
- Ordnung Peyssonneliales, mit etwa 121 Arten
- Ordnung Plocamiales, mit etwa 75 Arten
- Ordnung Rhodymeniales, mit etwa 404 Arten
- Ordnung Sebdeniales, mit etwa 18 Arten
- Unterklasse Hildenbrandiophycidae
- Klasse Bangiophyceae
Die Erforschung der Systematik der Rotalgen ist noch nicht abgeschlossen. Durch molekulargenetische Untersuchungen werden jährlich neue Arten entdeckt und weitere Änderungen der Einteilung notwendig werden.
Arten (Auswahlen)
In Europa heimische Arten (Auswahl)
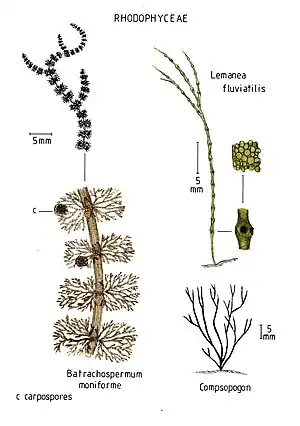
- Borsten-Rotalge (Lemanea fluviatilis) im Süßwasser
- Froschlaichalge (Batrachospermum gelatinosum) im Süßwasser
- Roter Horntang (Ceramium virgatum)
- Knorpeltang (Chondrus crispus)
- Korallenmoos (Corallina officinalis)
- Blutroter Meerampfer (Delesseria sanguinea)
- Schnurtang (Dumontia contorta)
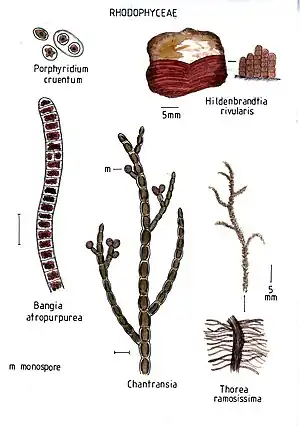
- Speckkrustenrotalge (Hildenbrandia rubra)
- Krustensteinblatt (Lithophyllum incrustans)
- Kraussterntang (Mastocarpus stellatus)
- Roter Hautflügeltang, Flügel-Seeampfer (Membranoptera alata)
- Lappentang (Palmaria palmata)
- Roter Eichentang (Phycodrys rubens)
- Kammtang (Plocamium cartilagineum)
- Zierliche Seefeder (Plumaria plumosa)
- Zarter Fadentang (Polysiphonia stricta)
- Porphyra purpurea, „Purpurblatt“
- Nabel-Purpurtang, Hauttang (Porphyra umbilicalis)
- Purpur-Rotalge (Porphyridium cruentum)
- Kalkkrustenrotalge (Phymatolithon polymorphum)
- Braunroter Zweigtang (Rhodomela confervoides)
- Pinselbüschelalge (Vertebrata lanosa)
Für die in der Deutschen Bucht vorkommenden Rotalgen siehe auch die Liste der Meeresalgen von Helgoland.
Außereuropäische Arten
- Vanvoorstia bennettiana, eine in Australien endemische Art, ist die einzige Rotalge, die bislang von der IUCN als ausgestorben gelistet wird.
Literatur
- Harold C. Bold, Michael J. Wynne: Introduction to the Algae – Structure and Reproduction. (= Prentice-Hall Biological Sciences Series). Prentice-Hall, 1978, ISBN 0-13-477786-7.
- Heinz Streble, Dieter Krauter: Das Leben im Wassertropfen. Mikroflora und Mikrofauna des Süßwassers. Ein Bestimmungsbuch. 10. Auflage. Kosmos, Stuttgart 2006, ISBN 3-440-10807-4.
- Klaus Lüning: Meeresbotanik – Verbreitung, Ökophysiologie und Nutzung der marinen Makroalgen. Georg Thieme Verlag, Stuttgart 1985, ISBN 3-13-667501-0.
- Reece Campbell: Biologie. Pearson Studium, 2006, ISBN 3-8273-7180-5.
- P. H. Raven, R. F. Evert, S. E. Eichhorn: Biologie der Pflanzen. Walter de Gruyter, 2006, ISBN 3-11-018531-8.
- Strasburger: Lehrbuch der Botanik. 36. Auflage. Akademie Verlag, 2008, ISBN 978-3-8274-1455-7.
Weblinks
Einzelnachweise
- Mitsunobu Kamiya, Sandra C. Lindstrom, Takeshi Nakayama, Akiko Yokoyama, Showe-Mei Lin, Michael D. Guiry, Carlos Frederico D. Gurgel, John Marinus Huisman, Taiju Kitayama, Masahiro Suzuki, Tae Oh Cho, Wolfgang Frey: Rhodophyta. In: Syllabus of Plant Families, 13. Auflage. Teil 2/2: Photoautotrophic eukaryotic Algae. (Frey, W. Hrsg.), S. [i]–xii, [1]–171. Stuttgart: Borntraeger Science Publishers. ISBN 978-3-443-01094-2. 2017.
- Nicholas J. Butterfield: Bangiomorpha pubescens n. gen., n. sp.: implications for the evolution of sex, multicellularity, and the Mesoproterozoic/Neoproterozoic radiation of eukaryotes. In: Paleobiology. 26(3), Jacksonville, NY, 2000, S. 386–404. doi:10.1666/0094-8373(2000)026<0386:BPNGNS>2.0.CO;2
- Focko Weberling, Hans Otto Schwantes: Pflanzensystematik. 2. Aufl., Ulmer, Stuttgart 1975, S. 242.
- Robert Edward Lee: Phycology. 5. Auflage, Cambridge University Press, Cambridge 2018, S. 84 f.
- Robert Edward Lee: Phycology. 5. Auflage, Cambridge University Press, Cambridge 2018, S. 86 f.
- Focko Weberling, Hans Otto Schwantes: Pflanzensystematik. 2. Aufl., Ulmer, Stuttgart 1975, S. 241.
- Robert Edward Lee: Phycology. 5. Auflage, Cambridge University Press, Cambridge 2018, S. 85.
- Robert Edward Lee: Phycology. 5. Auflage, Cambridge University Press, Cambridge 2018, S. 97–101.
- Focko Weberling, Hans Otto Schwantes: Pflanzensystematik. 2. Aufl., Ulmer, Stuttgart 1975, S. 244 f.
- Robert Edward Lee: Phycology. 5. Auflage, Cambridge University Press, Cambridge 2018, S. 84.
- Ein Beispiel ist Galdieria sulphuraria, siehe Julia Thurau: Rotalge: Galdi ist einfach anders. In: Der Tagesspiegel. 3. April 2001, abgerufen am 16. Oktober 2021.
- Janina Kwandrans, Pertti Eloranta (2010): Diversity of freshwater red algae in Europe. In: Oceanological and Hydrobiological Studies 39 (1): 161-169. doi:10.2478/v10009-010-0015-7
- Robert Edward Lee: Phycology. 5. Auflage, Cambridge University Press, Cambridge 2018, S. 92 f.
- This gin distilled in Donegal is flavored with seaweed. Algaeworld, abgerufen am 12. September 2021 (englisch).
- Irish moss, red algae Britannica, abgerufen am 12. September 2021 (englisch).
- Rotalgen (Agar-Agar). Apotheken.de, abgerufen am 12. September 2021.
- Rotalge: Diese Vitamine stecken drin. GEO, abgerufen am 12. September 2021.
- Nahrungsergänzung im Blick: Algen. Apotheken.de, abgerufen am 12. September 2021.
- Hiroyuki Kataoka, Naomi Ohnishi: Occurrence of Taurine in Plants. In: Agricultural and Biological Chemistry. Band 50, Nr. 7, Juli 1986, S. 1887–1888, doi:10.1080/00021369.1986.10867664 (PDF).
- Azusa Kawasaki, Ayuko Ono, Shoshi Mizuta, Mitsunobu Kamiya, Takaaki Takenaga: The Taurine Content of Japanese Seaweed. In: Advances in Experimental Medicine and Biology. 975, Teil 2, 2017, S. 1105–1112, doi:10.1007/978-94-024-1079-2_88, PMID 28849526.
- Versorgung mit Kalk, Magnesium, Schwefel und Natrium. Kalk (Ca). Ökolandbau.de, abgerufen am 12. September 2021.
- Seaweed and Seaweed Bioactives for Mitigation of Enteric Methane: Challenges and Opportunities. Abgerufen am 29. August 2022.
- Cultivating a natural solution to climate change. Sea Forest, abgerufen am 12. September 2021 (englisch).
- . Abgerufen am 29. August 2022.
- Seaweed and Seaweed Bioactives for Mitigation of Enteric Methane: Challenges and Opportunities. Abgerufen am 29. August 2022.
- Wie sieht die Rinderhaltung der Zukunft aus? Verbundvorhaben InnoRind erarbeitet innovative Ansätze für die Rinderhaltung von morgen. Uni Kiel, abgerufen am 12. September 2021.
- Tara Duggan: To fight climate change, California approves seaweed that cuts methane emissions in cow burps. In: San Francisco Chronicle. 6. Mai 2022, abgerufen am 9. Mai 2022 (amerikanisches Englisch).
- Gary W. Saunders, Max Hoyt Hommersand: Assessing red algal supraordinal diversity and taxonomy in the context of contemporary systematic data. In: American Journal of Botany. 91, 2004, S. 1494–1507. doi:10.3732/ajb.91.10.1494
- Hwan Su Yoon, Kirsten M. Müller, Robert G. Sheath, Franklyn D. Ott, Debashish Bhattacharya: Defining the major lineages of red algae (Rhodophyta). In: Journal of Phycology. Band 42, 2006, S. 482–492. (PDF) doi:10.1111/j.1529-8817.2006.00210.x
- S. M. Adl, A. G. B. Simpson, C. E. Lane, J. Lukeš, D. Bass, S. S. Bowser, M. W. Brown, F. Burki, M. Dunthorn, V. Hampl, A. Heiss, M. Hoppenrath, E. Lara, L. le Gall, D. H. Lynn, H. McManus, E. A. D. Mitchell, S. E. Mozley-Stanridge, L. W. Parfrey, J. Pawlowski, S. Rueckert, L. Shadwick, C. L. Schoch, A. Smirnov, F. W. Spiegel: The Revised Classification of Eukaryotes. In: Journal of Eukaryotic Microbiology. 59, 2012, S. 429–514. (PDF; online), S. 475–476.
- Michael D. Guiry, G. M. Guiry: Rhodophyta. In: Algaebase. - World-wide electronic publication, National University of Ireland, Galway, abgerufen am 23. Juli 2018
- Sergio A. Muñoz-Gómez, Fabián G. Mejía-Franco, Keira Durnin, Morgan Colp, Cameron J. Grisdale, John M. Archibald, Claudio H. Slamovits: The new red algal subphylum Proteorhodophytina comprises the largest and most divergent plastid genomes known. In: Current Biology, Heft 27, 2017, Seiten 1–8. doi:10.1016/j.cub.2017.04.054