Response Priming
Mit dem Begriff Response Priming bzw. Reaktionsbahnung bezeichnet man eine besondere Form des Priming in der Wahrnehmungspsychologie. Allgemein bestehen Priming-Effekte darin, dass die Reaktion (engl. response) auf einen Zielreiz (engl. target) von der vorausgehenden Präsentation eines Bahnungsreizes (engl. prime) beeinflusst wird. Die Besonderheit des Response Priming ist, dass beide Reize sehr schnell aufeinander folgen und mit motorischen Antwortalternativen verknüpft sind. Wenn eine Versuchsperson eine schnelle Reaktion ausführt, um den Zielreiz zu klassifizieren, kann ein kurz zuvor erscheinender Prime Antwortkonflikte auslösen, wenn er einer anderen Antwortalternative zugeordnet ist als der Zielreiz. Diese Antwortkonflikte schlagen sich in den Verhaltensdaten in Form von so genannten Priming-Effekten nieder, etwa in Reaktionszeiten und Fehlerraten. Eine besondere Eigenschaft von Response-Priming-Effekten ist ihre Unabhängigkeit von der bewussten Wahrnehmung (Sichtbarkeit) des Primes.
Response Priming als visomotorischer Effekt
Bereits 1962 berichteten Fehrer und Raab[1] Experimente, in denen Versuchspersonen durch das Drücken einer einzelnen Taste so schnell wie möglich auf das Erscheinen eines Reizes reagieren sollten, dessen Sichtbarkeit durch so genannte Metakontrast-Maskierung (s. u.) stark reduziert wurde. Sie fanden heraus, dass die Reaktionszeit unabhängig von der bewussten Sichtbarkeit des Reizes war, d. h. auf gut sichtbare Reize wurde ebenso schnell reagiert wie auf kaum sichtbare Reize (Fehrer-Raab-Effekt). Den Begriff Response Priming verwendeten 1982 zum ersten Mal Rosenbaum und Kornblum[2] im Zusammenhang mit einem experimentellen Paradigma, in dem Teile von motorischen Reaktionen durch Bahnungsreize geprimt wurden. Das moderne Verfahren des Response Priming wurde in den 80er und 90er Jahren von Peter Wolff, Werner Klotz, Ulrich Ansorge und Odmar Neumann an der Universität Bielefeld entwickelt.[3][4][5][6] Eine weitere wichtige Weiterentwicklung erfolgte Mitte der 90er Jahre durch die Arbeitsgruppe um Dirk Vorberg an der Technischen Universität Braunschweig.[7]
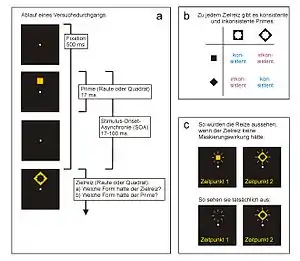
In experimentellen Paradigmen, die die Methode des Response Priming verwenden, müssen Probanden auf einen spezifischen Zielreiz reagieren. In einem einfachen Experiment könnte es sich dabei um einen von zwei geometrischen Reizen handeln, die zwei entsprechenden Reaktionstasten zugeordnet sind (z. B. Raute – linke Taste, Quadrat – rechte Taste). Das Experiment besteht dann aus vielen aufeinander folgenden Durchgängen, in denen die Versuchsperson immer links bei Erscheinen einer Raute und rechts bei Erscheinen eines Quadrats drücken muss. In jedem Durchgang erscheint dabei kurz vor der Präsentation des jeweiligen Zielreizes ein Prime, der ebenfalls die antwortauslösenden Eigenschaften des Zielreizes hat, also selbst eine Raute oder ein Quadrat ist (Abb. 1a). Sind Prime und Zielreiz mit gleichen Reaktionen verknüpft (Raute folgt auf Raute, Quadrat auf Quadrat), gelten sie als konsistent (auch „kongruent“, „kompatibel“); sind sie mit unterschiedlichen Reaktionen verknüpft (Raute folgt auf Quadrat, Quadrat auf Raute), als inkonsistent (auch „inkongruent“, „inkompatibel“; Abb. 1b). Das Zeitintervall zwischen Einsetzen des Primes und Einsetzen des Zielreizes wird Stimulus-Onset-Asynchronie (SOA) genannt. Typischerweise werden SOAs von ca. 100 ms (Millisekunden) oder weniger verwendet.
Priming-Effekte treten dann auf, wenn der Prime die motorische Reaktion auf den Zielreiz beeinflusst: konsistente Primes beschleunigen die Reaktionszeiten, inkonsistente Primes verlangsamen sie (Abb. 2). Priming-Effekte in Reaktionszeiten werden üblicherweise als Differenz der mittleren Reaktionszeiten in den konsistenten und inkonsistenten Durchgängen bestimmt. Außerdem führen konsistente Primes nur sehr selten zu Antwortfehlern (also falschen Reaktionen auf den Zielreiz), während die Fehlerhäufigkeit bei inkonsistenten Primes sehr hoch sein kann. Sowohl in den Reaktionszeiten als auch in den Fehlerraten steigen die Priming-Effekte typischerweise mit der SOA an, wodurch sich in Diagrammen der typische scherenartige Verlauf des Effekts ergibt.[7] Das bedeutet: je später der Zielreiz auf den Prime folgt, desto mehr Einfluss hat der Prime auf die Reaktionszeiten. Bei einer durchschnittlichen Reaktionszeit von 350 bis 450 ms kann der Response-Priming-Effekt auf über 100 ms ansteigen; er zählt damit zu den numerisch größten Effekten der Reaktionszeitforschung.
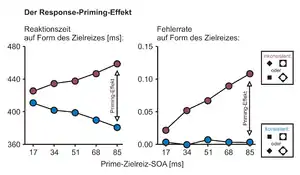
Heute gilt als gesichert, dass der Anstieg des Effekts mit der SOA darauf zurückzuführen ist, dass der Prime immer mehr Zeit hat, die Reaktion zu beeinflussen, bevor der eigentliche Zielreiz motorisch wirksam werden kann. Dies folgt klar aus der Analyse des Zeitverlaufs der motorischen Aktivität im EEG,[8][9][10][11][12] in geprimten Zeigebewegungen[13][14][15] in Kraftmessungen[16] und in Simulationsstudien.[7] Wie groß der Priming-Effekt wird, hängt daher sowohl von den Reizeigenschaften als auch von Eigenschaften der Aufgabe ab. Primes mit höherer Reizenergie (d. h. höherem Kontrast, längerer Dauer etc.) sowie Aufgaben mit einfachen Reizdiskriminationen führen zu großen Priming-Effekten, während Primes mit geringer Reizenergie und schwierige Unterscheidungsaufgaben zu geringeren Effekten führen.[14][15] Priming-Effekte können durch visuelle Aufmerksamkeit verstärkt werden, die rechtzeitig vor Präsentation des Primes auf seine Position oder seine relevanten Merkmale gelenkt wird.[17][18][19]
Der beschriebene Zeitverlauf gilt für SOAs von bis zu ca. 100 ms. Für längere SOAs kann der Effekt noch weiter ansteigen. Unter bestimmten Bedingungen kann man aber auch eine Umkehrung des Effekts beobachten, bei der inkonsistente Primes zu schnelleren Reaktionen auf den Zielreiz führen als konsistente Primes. Dieser Effekt wird häufig als „negativer Kompatibilitätseffekt“ bezeichnet.[9][10][20][21][22][23][24]
Maskiertes Priming
Response Priming kann genutzt werden, um Phänomene der unbewussten Wahrnehmung zu untersuchen. Dabei kann die bewusste Sichtbarkeit des Primes mit Hilfe eines Maskierreizes systematisch verändert werden, bis zur völligen Unsichtbarkeit des Primes. Dies geschieht durch die Präsentation des Maskierreizes kurz vor oder nach der des Primes.[25] Die Sichtbarkeit des Primes kann durch verschiedene Maße erhoben werden, z. B. Forced-Choice-Diskrimination, Entdeckungsurteile, Helligkeitsurteile und andere Maße.[26][27] In vielen Response-Priming-Experimenten dient der Zielreiz selbst zur Maskierung des Primes (Abb. 1). Die sogenannte Metakontrast-Maskierung wird dadurch erreicht, dass auf den Prime eine Maske folgt, die diesen umschließt, so dass beide Reize angrenzende Konturen haben. Ein Kreis kann zum Beispiel durch einen größeren Ring maskiert werden, dessen Innenmaße den Außenmaßen des Kreises entsprechen. Metakontrast ist eine Form von visueller Rückwärtsmaskierung, d. h. die Sichtbarkeit des Primes wird durch die Präsentation eines nachfolgenden Reizes heruntergesetzt.[28][29][25]
Abb. 3 zeigt typische Maskierungsverläufe in Abhängigkeit von SOA, Prime und Zielreiz, wobei der Zielreiz gleichzeitig als Maske dient. Als Maß der bewussten Sichtbarkeit des Primes könnte hier etwa die Diskriminationsleistung einer Versuchsperson dienen, die die Aufgabe hat, in jedem Durchgang die Form des Primes (Raute oder Quadrat) zu erraten. Ohne Maskierung wäre die Leistung praktisch perfekt: die Versuchsperson könnte problemlos in jedem Durchgang den Prime korrekt als Raute oder Quadrat benennen. Bei vollständiger Maskierung des Primes würde die Leistung hingegen bei Zufallsniveau liegen (Abb. 3 links). In vielen Experimenten ergeben sich allerdings weniger extreme Maskierungsverläufe (Abb. 3 rechts). Die überwiegende Mehrheit der Experimente führt zu so genannter Typ-A-Maskierung, die bei kurzer SOA zwischen Prime und Maske am stärksten ist und mit wachsender SOA abnimmt, so dass der Prime immer leichter zu unterscheiden ist. Unter bestimmten Bedingungen kann es aber auch zu Typ-B-Maskierung kommen, bei der die Maskierungswirkung bei mittlerer SOA am stärksten ist, der Prime bei kürzeren oder längeren SOAs jedoch leichter zu unterscheiden ist. Typ-B-Maskierung kann vor allem bei Metakontrast-Maskierung vorkommen, hängt jedoch empfindlich von den Eigenschaften von Prime und Maske ab. Außerdem kann sich der Maskierungsverlauf von Person zu Person stark unterscheiden.[25][28][30]

Unabhängigkeit von Response Priming und visuellem Bewusstsein
Experimente zeigen, dass der Zeitverlauf des Response-Priming-Effekts (ansteigender Effekt mit steigender SOA) unabhängig vom Zeitverlauf der Maskierung ist. Klotz und Neumann (1999) demonstrierten Response-Priming-Effekte bei kompletter Maskierung des Primes.[4] Vorberg et al.[7] variierten den Zeitverlauf der Maskierung, indem sie die relative Dauer von Primes und Zielreizen veränderten. Zielreize waren Pfeile, die nach links oder rechts zeigen konnten, und Primes waren kleinere Pfeile, die durch die Zielreize metakontrast-maskiert wurden. Wenn die Versuchspersonen entscheiden mussten, in welche Richtung die Primes zeigten, konnten je nach Bedingung alle in Abb. 3 veranschaulichten Arten von Maskierungsverläufen erzeugt werden: komplette Sichtbarkeit, komplette Maskierung, Typ-A-Maskierung und Typ-B-Maskierung. Wenn die Versuchspersonen aber so schnell wie möglich auf die Richtung des Zielreizes reagieren sollten, ergaben sich in allen Bedingungen fast identische Priming-Effekte. Der Zeitverlauf dieser Effekte war immer gleich (mit der SOA ansteigend), egal ob die Primes komplett sichtbar oder komplett unsichtbar waren und egal ob die Sichtbarkeit mit der SOA zu- oder abnahm.[7] Priming-Effekte können also selbst dann zunehmen, wenn die Sichtbarkeit des Primes abnimmt. Solche entgegengesetzten Zeitverläufe von Priming und visuellem Bewusstsein des Primes zeigen, dass beide Prozesse auf unterschiedlichen Mechanismen beruhen.[31] Dieser Befund konnte in vielen weiteren Experimenten bestätigt werden, in denen sich zahlreiche Dissoziationen zwischen Maskierung und Priming-Effekten ergaben.[1][3][4][5][32][33][34][35][36][36][37][13][14][19] Die Unabhängigkeit von Priming und visuellem Bewusstsein spricht klar gegen die traditionelle Auffassung, nach der unbewusste Priming-Prozesse allenfalls eine Restfähigkeit sind, die zu einem kleinen Grade erhalten bleibt, wenn die Erkennbarkeit der Reize unter eine bestimmte Schwelle gesunken ist. Diese Auffassung hat immer wieder zu scharfer Kritik an der Forschung zur unbewussten oder subliminalen Wahrnehmung geführt,[38][39][40][41] aber sie ist vermutlich grundlegend falsch. Vielmehr erfolgt die motorische Aktivierung durch maskierte Primes offenbar unabhängig von Prozessen der Rückwärtsmaskierung, vorausgesetzt die Sichtbarkeit der Primes wird nur durch die Art der Maske bestimmt, während der Prime-Reiz selbst unverändert bleibt. Das heißt: visuell unbewusste (unsichtbare) Reize können, in einem kurzen Zeitintervall und unter bestimmten Bedingungen, motorische Reaktionen in der gleichen Weise beeinflussen wie bewusste Reize.
Varianten
Vorausgesetzt, man ist sich im Klaren über die Rolle bestimmter einflussreicher Variablen,[42] kann Response Priming in zahlreichen experimentellen Varianten und zur Untersuchung einer Vielzahl von Fragestellung der kognitiven Psychologie eingesetzt werden.[43] Die am häufigsten verwendete Form von benutzt einen Prime und einen Zielreiz an der gleichen Bildschirmposition, wobei der Zielreiz gleichzeitig als Maske dient (häufig nach dem Prinzip der Metakontrast-Maskierung). In vielen Experimenten werden auch zwei verschiedene Zielreize gleichzeitig gezeigt, denen zwei Primes an den gleichen Positionen vorausgehen.[3][4][13] Die Versuchsperson muss dann die beiden Zielreize unterscheiden und auf die Position des für die Aufgabe relevanten Zielreizes reagieren. Manchmal werden auch drei Reizarten eingesetzt (Prime, Maske, Zielreiz), vor allem wenn die SOA zwischen Prime und Zielreiz sehr lang sein muss.[21][22] Manchmal wird auch gar keine Maske verwendet.[19] Prime und Zielreiz müssen sich nicht an denselben Positionen befinden: Ein Reiz kann den anderen auch flankieren, wie es im Eriksen-Paradigma der Fall ist.[7][11][44][45] Der Eriksen-Effekt könnte nach heutigem Kenntnisstand ein Spezialfall des Response Priming sein.
Response-Priming-Effekte sind mit einer Vielzahl von Reizen und Diskriminationsaufgaben demonstriert worden, u. a. geometrischen Reizen,[3][4][15] Farbreizen[13][14][32] verschiedenen Arten von Pfeilen[7][9][10][46] natürlichen Bildern (Tiere vs. Objekte),[15] Vokalen und Konsonanten[47] Buchstaben[44] und Ziffern.[33] In einer Studie wurden Schachkonfigurationen als Primes und Zielreize präsentiert, und die Versuchspersonen mussten entscheiden, ob der König im Schach stand.[35] Ebenso wurden vielfältige Arten von Maskierung verwendet. Einige Experimente messen statt Tastenreaktionen (die meist mit zwei Antwortalternativen erfolgen) Sprechreaktionen[5] zielgerichtete Zeigebewegungen,[13][14][15] Blickbewegungen[48] oder so genannte Bereitschaftspotentiale, die die motorische Aktivierung im Gehirn erfassen und mit Methoden der Elektroenzephalografie gemessen werden.[8][9][12][49] Auch bildgebende Verfahren wie die Magnetresonanztomografie kommen zum Einsatz.[33] Wieder andere Experimente verwenden mehr als zwei Reaktionsalternativen. Mattler (2003) konnte zeigen, dass Response Priming nicht nur motorische Handlungen beeinflussen kann, sondern auch kognitive Operationen wie die Verlagerung der räumlichen Aufmerksamkeit oder den Wechsel zwischen Reaktionszeit-Aufgaben.[36]
Theorien
Im Folgenden werden drei Theorien zur Erklärung des positiven Response-Priming-Effekts vorgestellt. Für eine Übersicht der Erklärungsansätze zum negativen Kompatibilitätseffekt siehe Sumner (2007).[24]
Direkte Parameterspezifikation
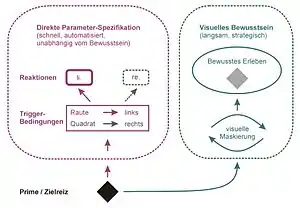
Die Theorie der direkten Parameterspezifikation („direct parameter specification“; Abb. 4) wurde von Odmar Neumann an der Universität Bielefeld entwickelt, um den Fehrer-Raab-Effekt und die frühesten Response-Priming-Studien zu erklären.[50] Diese Theorie geht davon aus, dass die Versuchsperson zu Beginn des Versuchs Regeln der Reiz-Reaktions-Zuordnung erwirbt, die nach kurzer Einübung in automatisierter Form vorliegen. Ist diese Übungsphase abgeschlossen, kann die Reaktion so weit vorbereitet werden, dass nur noch ein einzelnes kritisches Reizmerkmal (z. B. Raute vs. Quadrat) zur Auslösung der Reaktion nötig ist. Dieses eingehende Reizmerkmal definiert dann den letzten noch fehlenden Handlungsparameter („action parameter“, z. B. linke vs. rechte Reaktion). Die Handlungsauslösung erfolgt schnell und direkt, ohne dass eine bewusste Repräsentation des Reizes nötig wäre. Response Priming wird dadurch erklärt, dass der Prime mit seinen Reizeigenschaften genau dieselben Prozesse der direkten Parameterspezifikation auslöst, die instruktionsgemäß erst durch den Zielreiz erfolgen sollten. Parallel zur Antwortauslösung im visomotorischen System entsteht eine bewusste Repräsentation von Prime und Zielreiz, die aber Prozessen der visuellen Maskierung unterworfen sein kann. Die bewusste Repräsentation spielt allerdings keine Rolle für die motorische Reaktion auf Primes und Zielreize im aktuellen Versuchsdurchgang.
Action-Trigger-Ansatz
Der „Action-Trigger-Ansatz“ (engl. „action trigger account“) wurde von Wilfried Kunde, Andrea Kiesel und Joachim Hoffmann an der Universität Würzburg entwickelt.[47][51] Dieser Ansatz betont, dass Reaktionen auf unbewusste Reize weder durch semantische Analyse noch durch zuvor erlernte Reiz-Reaktions-Verknüpfungen ausgelöst werden. Stattdessen wird angenommen, dass ein Prime in eine existierende „action release condition“ passt und damit die Reaktion auslöst, ähnlich wie ein Schlüssel, der ein Schloss öffnet. Dies geschieht immer in zwei zeitlich aufeinander folgenden Schritten. Zuerst werden zu einer erwarteten oder bekannten Aufgabe „action triggers“ im Arbeitsgedächtnis aktiv gehalten, die eine spezifische motorische Reaktion auslösen sollen. Diese „action triggers“ werden in der Instruktions- und Übungsphase des Experiments gebildet. Im zweiten Schritt, „online stimulus processing“ genannt, wird verglichen, ob ein gezeigter Reiz in ein bekanntes Schema passt. Ist dies der Fall, so wird die verknüpfte Reaktion automatisch ausgelöst. Ein Beispiel wäre die Aufgabe zu sagen, ob eine dargebotene Zahl größer oder kleiner als fünf ist:[33] bei Präsentation der Zahlen „1“ bis „4“ soll die linke Taste gedrückt werden, bei Präsentation der Zahlen „6“ bis „9“ die rechte. Infolge der Instruktion werden „action triggers“ gebildet, die bei Präsentation des relevanten Reizes automatisch die entsprechende Reaktion hervorrufen. Eine wichtige Voraussage dieser Theorie ist, dass Reaktionen auch durch Primes ausgelöst werden können, die zwar selbst niemals als Target vorkommen, aber die Trigger-Bedingungen erfüllen.[51]
Auch hier spielt die bewusste Repräsentation des Reizes keine Rolle für die aktuelle motorische Aktivierung; sie kann aber dazu führen, dass in späteren Versuchsdurchgängen Reaktionskriterien strategisch geändert werden (etwa, um Antwortfehler zu vermeiden). Insgesamt kann diese Theorie als weitere Ausgestaltung des Konzeptes der Direkten Parameterspezifikation verstanden werden.
Rapid-Chase-Theorie
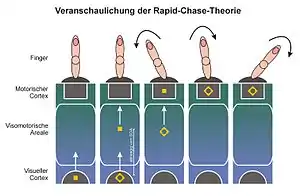
Die „Rapid-Chase-Theorie“[14][12][15] wurde 2006 von Thomas Schmidt, Silja Niehaus und Annabel Nagel vorgestellt. Sie verknüpft das Modell der direkten Parameterspezifikation mit der Beobachtung, dass neu erscheinende visuelle Reize eine Aktivitätswelle im visomotorischen System auslösen, die sich rasch von visuellen Arealen hin zu motorischen Arealen ausbreitet.[52][53][54][55] Da die Ausbreitungsgeschwindigkeit dieser Aktivitätswelle sehr hoch ist, haben Victor Lamme und Pieter Roelfsema von der Universität Amsterdam die These aufgestellt, dass hier zunächst ein reiner Feedforward-Prozess (Feedforward Sweep) vorliegt: Eine Zelle, die zuerst von der Wellenfront erreicht wird, muss ihre Aktivität weitergeben, ohne vorher Feedback von anderen Zellen integrieren zu können. Zugleich nehmen Lamme und Roelfsema an, dass eine solche Feedforward-Verarbeitung allein nicht ausreicht, um visuelles Bewusstsein eines Reizes entstehen zu lassen: Hierzu seien Feedback-Prozesse und rekurrente (wiederkehrende) Verarbeitungsschleifen nötig, die auch weit entfernte Hirnareale miteinander verbinden können.[53][29]
Nach der Rapid-Chase-Theorie lösen Prime und Zielreiz nacheinander solche „Feedforward Sweeps“ aus, die schließlich motorische Areale des Gehirns erreichen. Dort werden nacheinander motorische Prozesse ausgelöst; auch hier erfolgt die Auslösung automatisch und ohne Beteiligung des Bewusstseins. Da der Prime einen zeitlichen Vorsprung hat, liefern sich Prime und Zielreiz eine „Verfolgungsjagd“ („rapid chase“) durch das visomotorische System. Da das Prime-Signal den Motorkortex zuerst erreicht, leitet es die mit ihm verknüpfte motorische Reaktion ein. Je kürzer die SOA ist, desto früher kann das Zielreiz-Signal die Verfolgung aufnehmen. Erst wenn das Zielreiz-Signal ebenfalls im Motorkortex eingetroffen ist, kann es die motorische Reaktion weiterführen (wenn es konsistent zum Prime ist) oder sie umlenken (wenn es inkonsistent zum Prime ist). Damit lässt sich erklären, dass Response-Priming-Effekte mit der SOA ansteigen: Je länger die SOA ist, desto länger kann der Prime die Reaktion alleine steuern, und desto weiter kann er den Reaktionsprozess in eine bestimmte Richtung lenken. Gegebenenfalls kann der Prime auch einen Fehler provozieren (daraus ergeben sich die sehr häufig beobachteten Priming-Effekte in den Fehlerraten). Ein solcher Ablauf der motorischen Aktivierung durch Prime und Target ist bereits 2003 von Dirk Vorberg und Mitarbeitern in einem mathematischen Modell beschrieben worden[7] und stimmt auch mit frühen EEG-Befunden zum Response Priming überein.[8][56] Response-Priming-Effekte sind laut der Rapid-Chase-Theorie unabhängig vom visuellen Bewusstsein, weil sie von schnellen Feedforward-Prozessen getragen werden, während die Entstehung einer bewussten Repräsentation auf langsamere, rekurrente Prozesse angewiesen ist.[29][53]
Die wichtigste Voraussage der Rapid-Chase-Theorie ist, dass die Feedforward-Sweeps von Prime und Zielreiz streng sequentiell erfolgen sollten. Diese strenge Abfolge sollte sich im Zeitverlauf der motorischen Reaktionen niederschlagen, und es sollte einen frühen Zeitabschnitt geben, in der die Reaktion allein durch den Prime gesteuert wird, aber unabhängig von allen Eigenschaften des eigentlichen Zielreizes ist. Besonders gut lassen sich diese Voraussagen anhand des Zeitverlaufs von geprimten Zeigebewegungen untersuchen.[13] Hier zeigt sich, dass die Zeigebewegung zu einer festen Zeit nach Erscheinen des Primes (nicht des eigentlichen Zielreizes) beginnt und zunächst in Richtung des Primes erfolgt. Wenn Prime und Zielreiz inkonsistent sind, kann der Zielreiz die Zeigerichtung „im Flug“ umkehren und in die richtige Richtung lenken; je länger die SOA ist, desto länger und weiter bewegt sich der Finger in Richtung des irreführenden Primes.[13] Schmidt, Niehaus und Nagel konnten zeigen, dass die früheste Phase solcher geprimten Zeigebewegungen nur von Eigenschaften des Primes (hier dem Farbkontrast von roten versus grünen Primes) abhängt, nicht aber von Eigenschaften des Zielreizes (dem Zeitpunkt seines Erscheinens, seinem Farbkontrast oder seiner Maskierungswirkung).[14] Diese Befunde konnten mit verschiedenen Methoden und Reizen bestätigt werden.[12][15][19][18]
Da die Rapid-Chase-Theorie Response Priming als einen Feedforward-Prozess betrachtet, geht sie davon aus, dass die Priming-Effekte entstehen, bevor rekurrente und Feedback-Prozesse in die Verarbeitung eingreifen können. Die Theorie stellt deshalb die kontroverse These auf, dass Response-Priming-Effekte ein Maß für die vorbewusste Verarbeitung visueller Information darstellen, welche sich von der Repräsentation der Reize im visuellen Bewusstsein grundlegend unterscheiden kann.[37]
Einzelnachweise
- E. Fehrer, D. Raab: Reaction time to stimuli masked by metacontrast. In: Journal of Experimental Psychology. Nr. 63, 1962, S. 143–147.
- D. A. Rosenbaum, S. Kornblum: A priming method for investigating the selection of motor responses. In: Acta Psychologica. Nr. 51, 1982, S. 223–243.
- W. Klotz, P. Wolff: The effect of a masked stimulus on the response to the masking stimulus. In: Psychological Research. Nr. 58, 1995, S. 92–101.
- W. Klotz, O. Neumann: Motor activation without conscious discrimination in metacontrast masking. In: Journal of Experimental Psychology: Human Perception and Performance. Nr. 25, 1999, S. 976–992.
- U. Ansorge, W. Klotz, O. Neumann: Manual and verbal responses to completely masked (unreportable) stimuli: Exploring some conditions for the metacontrast dissociation. In: Perception. Nr. 27, 1998, S. 1177–1189.
- U. Ansorge, O. Neumann, S. I. Becker, H. Kälberer, H. Kruse: Sensorimotor supremacy: Investigating conscious and unconscious vision by masked priming. In: Advances in Cognitive Psychology. Nr. 3, 2007, S. 257–274.
- D. Vorberg, U. Mattler, A. Heinecke, T. Schmidt, J. Schwarzbach: Different time courses for visual perception and action priming. In: Proceedings of the National Academy of Sciences USA. Nr. 100, 2003, S. 6275–6280.
- H. Leuthold, B. Kopp: Mechanisms of priming by masked stimuli: Inferences from event-related brain potentials. In: Psychological Science. Nr. 9, 1998, S. 263–269.
- M. Eimer, F. Schlaghecken: Effects of masked stimuli on motor activation: Behavioral and electrophysiological evidence. In: Journal of Experimental Psychology: Human Perception and Performance. Nr. 24, 1998, S. 1737–1745.
- M. Eimer, F. Schlaghecken: Response facilitation and inhibition in subliminal priming. In: Biological Psychology. Nr. 64, 2003, S. 7–26.
- U. Mattler: Delayed flanker effects on lateralized readiness potentials. In: Experimental Brain Research. Nr. 151, 2003, S. 272–288.
- N. Vath, T. Schmidt: Tracing sequential waves of rapid visuomotor activation in lateralized readiness potentials. In: Neuroscience. Nr. 145, 2007, S. 197–208.
- T. Schmidt: The finger in flight: Real-time motor control by visually masked color stimuli. In: Psychological Science. Nr. 13, 2002, S. 112–118.
- T. Schmidt, S. Niehaus, A. Nagel: Primes and targets in rapid chases: Tracing sequential waves of motor activation. In: Behavioural Neuroscience. Nr. 120, 2006, S. 1005–1016.
- T. Schmidt, F. Schmidt: Processing of natural images is feedforward: A simple behavioral test. In: Attention, Perception & Psychophysics. Nr. 71, 2009, S. 594–606.
- U. Mattler: Flanker effects on motor output and the late-level response activation hypothesis. In: The Quarterly Journal of Experimental Psychology. Nr. 58A, 2005, S. 577–601.
- P. Sumner, P.-C. Tsai, K. Yu, P. Nachev: Attentional modulation of sensorimotor processes in the absence of perceptual awareness. In: Proceedings of the National Academy of Sciences USA. Nr. 103, 2006, S. 10520–10525.
- T. Schmidt, A. Seydell: Visual attention amplifies response priming of pointing movements to color targets. In: Perception & Psychophysics. Nr. 70, 2008, S. 443–455.
- F. Schmidt, T. Schmidt: Feature-based attention to unconscious shapes and colors. In: Attention, Perception, Psychophysics. Nr. 72, 2010, S. 1480–1494.
- P. Jaskowski: The negative compatibility effect with nonmasking flankers: A case for mask-triggered inhibition hypothesis. In: Consciousness & Cognition. Nr. 17, 2008, S. 765–777.
- R. Verleger, P. Jaskowski, A. Aydemir, R. H. J. van der Lubbe, M. Groen: Qualitative differences between conscious and nonconscious processing? On inverse priming induced by masked arrows. In: Journal of Experimental Psychology. Nr. 133, 2004, S. 494–515.
- A. Lingnau, D. Vorberg: The time course of response inhibition in masked priming. In: Perception & Psychophysics. Nr. 67, 2005, S. 545–557.
- S. T. Klapp, L. B. Hinkley: The negative compatibility effect: Unconscious inhibition influences reaction time and response selection. In: Journal of Experimental Psychology: General. Nr. 131, 2002, S. 255–269.
- P. Sumner: Negative and positive masked priming – implications for motor inhibition. In: Advances in Cognitive Psychology. Nr. 3, 2007, S. 317–326.
- B. G. Breitmeyer, H. Öğmen: Visual masking. In: Scholarpedia. 2(7), 2007, S. 3330.
- U. Ansorge, B. G. Breitmeyer, S. I. Becker: Comparing sensitivity across different processing measures under metacontrast masking conditions. In: Vision Research. Nr. 47, 2007, S. 3335–3349.
- U. Ansorge, S. I. Becker, B. Breitmeyer: Revisiting the metacontrast dissociation: Comparing sensitivity across different measures and tasks. In: Quarterly Journal of Experimental Psychology. Nr. 62, 2009, S. 286–309.
- B. G. Breitmeyer, H. Öğmen: Visual masking. Oxford University Press, Oxford 2006.
- V. DiLollo, J. T. Enns, R. A. Rensink: Competition for consciousness among visual events: The psychophysics of re-entrant visual processes. In: Journal of Experimental Psychology: General. Nr. 129, 2000, S. 481–507.
- T. Albrecht, S. Klapötke, U. Mattler: Individual differences in metacontrast masking are enhanced by perceptual learning. In: Consciousness and Cognition. Nr. 19, 2010, S. 656–666.
- T. Schmidt, D. Vorberg: Criteria for unconscious cognition: Three types of dissociation. In: Perception & Psychophysics. Nr. 68, 2006, S. 489–504.
- B. G. Breitmeyer, T. Ro, N. S. Singhal: Unconscious color priming occurs at stimulus- not percept-dependent levels of visual processing. In: Psychological Science. Nr. 15, S. 198–202.
- S. Dehaene, L. Naccache, G. Le Clec'H, E. Koechlin, M. Mueller, G. Dehaene-Lambertz, P. F. van de Moortele, D. Le Bihan: Imaging unconscious semantic priming. In: Nature. Nr. 395, 1998, S. 597–600.
- S. Fellows, R. Tabaza, M. Heumann, W. Klotz, O. Neumann, M. Schwarz, J. Noth, R. Topper: Modification of a functional motor task by non-consciously perceived sensory stimuli. In: Neuroreport. Nr. 13, 2002, S. 637–640.
- A. Kiesel, W. Kunde, C. Pohl, M. P. Berner, J. Hoffmann: Playing chess unconsciously. In: Journal of Experimental Psychology: Learning, Memory, and Cognition. Nr. 35, 2009, S. 292–298.
- U. Mattler: Priming of mental operations by masked stimuli. In: Perception & Psychophysics. Nr. 65, 2003, S. 167–187.
- T. Schmidt, S. Miksch, L. Bulganin, F. Jäger, F. Lossin, J. Jochum, P. Kohl: Response priming driven by local contrast, not subjective brightness. In: Attention, Perception & Psychophysics. Nr. 72, 2010, S. 1556–1568.
- C. W. Eriksen: Discrimination and learning without awareness: A methodological survey and evaluation. In: Psychological Review. Nr. 67, 1960, S. 279–300.
- D. Holender: Semantic activation without conscious identification in dichotic listening, parafoveal vision, and visual masking: A survey and appraisal. In: Behavioral and Brain Sciences. Nr. 9, 1986, S. 1–23.
- D. Holender, K. Duscherer: Unconscious perception: The need for a paradigm shift. In: Perception & Psychophysics. Nr. 66, 2004, S. 872–881.
- P. Cardoso-Leite, A. Gorea: On the perceptual/motor dissociation: A review of concepts, theory, experimental paradigms and data interpretations. In: Seeing and Perceiving. Nr. 23, 2010, S. 89–151.
- F. Schmidt, A. Haberkamp, T. Schmidt: Dos and don'ts in response priming research. In: Advances in Cognitive Psychology. Nr. 7, 2011, S. 120–131.
- T. Schmidt, A. Haberkamp, G. M. Veltkamp, A. Weber, A. Seydell-Greenwald, F. Schmidt: Visual processing in rapid-chase systems: Image processing, attention, and awareness. In: Frontiers in Psychology. Nr. 2, 2011, S. 1–16.
- B. A. Eriksen, C. W. Eriksen: Effects of noise letters upon the identification of a target letter in a nonsearch task. In: Perception & Psychophysics. Nr. 16, 1974, S. 143–149.
- W. Schwarz, A. Mecklinger: Relationship between flanker identifiability and compatibility effect. In: Perception & Psychophysics. Nr. 57, 1995, S. 1045–1052.
- P. Jaskowski, M. Slosarek: How important is the prime's gestalt for subliminal priming? In: Consciousness & Cognition. Nr. 16, 2007, S. 485.497.
- W. Kunde, A. Kiesel, J. Hoffmann: Conscious control over the content of unconscious cognition. In: Cognition. Nr. 88, 2003, S. 223–242.
- J. Schwarzbach, D. Vorberg: Response priming with and without awareness. In: H. Öğmen, B. G. Meyer (Hrsg.): The first half second: The microgenesis and temporal dynamics of unconscious and conscious visual processes. MIT Press, Cambridge, MA.
- P. Jaskowski, A. Bialunska, M. Tomanek, R. Verleger: Mask- and distractor-triggered inhibitory processes in the priming of motor responses: An EEG study. In: Psychophysiology, Nr. 45, 2008, S. 70–85.
- O. Neumann: Direct parameter specification and the concept of perception. In: Psychological Research. Nr. 52, 1990, S. 207–215.
- A. Kiesel, W. Kunde, J. Hoffmann: Mechanisms of subliminal response priming. In: Advances in Cognitive Psychology. Nr. 1–2, 2007, S. 307–315.
- J. Bullier: Integrated model of visual processing. In: Brain Research Reviews. Nr. 36, 2001, S. 96–107.
- V. A. F. Lamme, P. R. Roelfsema: The distinct modes of vision offered by feedforward and recurrent processing. In: Trends in Neurosciences. Nr. 23, 2000, S. 571–579.
- S. J. Thorpe, D. Fize, C. Marlot: Speed of processing in the human visual system. In: Nature. Nr. 381, 1996, S. 520–522.
- R. VanRullen, S. J. Thorpe: Surfing a spike wave down the ventral stream. In: Vision Research. Nr. 42, S. 2593–2615.
- W. Klotz, M. Heumann, U. Ansorge, O. Neumann: Electrophysiological activation by masked primes: Independence of prime-related and target-related activities. In: Advances in Cognitive Psychology. Nr. 3, 2007, S. 449–465.