Adaptation (Auge)
Unter Adaptation (lat. adaptare „anpassen“) versteht man beim Auge dessen Anpassung an die im Gesichtsfeld vorherrschenden Leuchtdichten.

Bei plötzlichen Helligkeitsunterschieden kann über den Pupillenlichtreflex vermittelt durch Verengung oder Erweiterung der Pupillen mit der Irismuskulatur die einfallende Lichtmenge rasch angepasst werden, jedoch ist hierdurch nur eine Veränderung innerhalb eines Spielraums von etwa 1:10 erreichbar.
Die weitergehende Anpassung an unterschiedliche Umgebungshelligkeiten kommt durch eine Änderung der Lichtempfindlichkeit der Netzhaut (Retina) zustande und ist erst nach einer gewissen Verzögerung optimal. Erst diese retinale Adaptation macht es möglich, Lichtreize sehr verschiedener Stärke zu verarbeiten, um etwa schwach scheinende Sterne am mondlosen Nachthimmel oder aber Spuren im Schnee bei Sonnenlicht zu sehen (Spielraum etwa 1:1012).[1] Hierzu tragen außer biochemisch angepassten Transduktionsprozessen in den Sinneszellen (Photorezeptoren) auch unterschiedlich gewichtete Verschaltungen innerhalb des rezeptiven Feldes nachgeordneter retinaler Nervenzellen bei.[2]
Pupillenlichtreflex
Die Iris (Regenbogenhaut) stellt die Begrenzung des Sehlochs (Pupille) dar. Das Ergebnis des Pupillenlichtreflexes, kurz Pupillenreflex, ist eine Tonusänderung der glatten Irismuskulatur. Dies bewirkt eine Veränderung der Pupillenweite, wodurch die relative Menge des in das Auge einfallenden Lichts angepasst werden kann. Der Mechanismus ist vergleichbar mit der Regulierung der Blendenöffnung bei Fotoapparaten. Dafür besitzt die Iris zwei Muskeln:
- Der Musculus dilatator pupillae („Pupillenerweiterer“) wird von sympathischen Nervenfasern aus dem Centrum ciliospinale (Rückenmarkssegmente C8–Th3) innerviert. Die Erweiterung der Pupille wird als Mydriasis bezeichnet.
- Der Musculus sphincter pupillae („Pupillenverenger“) wird von parasympathischen Fasern des Nervus oculomotorius (III. Hirnnerv) aus dem Ncl. Edinger-Westphal über das Ganglion ciliare innerviert. Er wird bei hohem Lichteinfall aktiviert. Eine Verengung der Pupille wird als Miosis bezeichnet.
Die reflektorische Regelung des Lichteinfalls durch die Pupille bewirkt eine rasche Anpassung an plötzliche Wechsel der Helligkeit. Bei Zunahme des Pupillendurchmessers um das Dreifache liegt die Vergrößerung der Öffnungsfläche in der Größenordnung von Faktor 10 (101). Da der Gesamtbereich der Hell-/Dunkeladaptation jedoch mehr als 11 Größenordnungen beträgt (um 1012), spielt der Pupillenreflex in diesem Zusammenhang nur eine untergeordnete Rolle.
Reflexkette
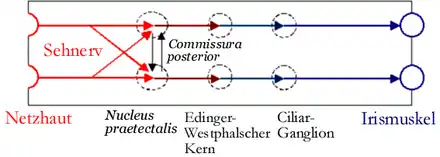
Afferenz: Die Information über den erhöhten Lichteinfall wird von lichtempfindlichen Photorezeptoren in der Retina über den Sehnerv (Nervus opticus) und Tractus opticus in den Epithalamus zu den Nuclei praetectales geleitet. Deren Efferenzen leiten die Helligkeits-Information beidseitig in die Edinger-Westphal-Kerne (Nuclei accessorii nervi oculomotorii).
Efferenz: In den Edinger-Westphal-Kernen findet eine Verschaltung auf den parasympathischen Anteil des Nervus oculomotorius statt. Über das Ziliarganglion wird der Musculus sphincter pupillae zur Kontraktion angeregt und damit die Pupille verengt. Da einerseits beide prätektalen Kerne über die Commissura posterior verbunden sind und von jedem Auge eine Verknüpfung zu beiden prätektalen Kernen besteht, wird der Reflex von beiden Augen gleichzeitig durchgeführt, auch wenn nur ein Auge plötzlich beleuchtet wird.[3] Daher kann auch an einem blinden Auge durch Beleuchtung des anderen gesunden Auges eine Pupillenverengung ausgelöst werden, solange der Reflexbogen intakt ist (konsensuelle Lichtreaktion).
Bei Vögeln und Reptilien bestehen die Irismuskeln vorwiegend aus quergestreifter Muskulatur und sind willkürlich beeinflussbar. Da bei diesen Wirbeltierklassen alle Sehnervenfasern kreuzen, zeigen sie auch keinen konsensuellen Pupillenlichtreflex.[4]
Anpassungsvorgänge der Netzhaut

Über einen weiten Bereich können die lichtempfindlichen Photorezeptoren der Netzhaut ihre Empfindlichkeit in Abhängigkeit von der Beleuchtungsstärke ändern. Die Dunkeladaptation ist dabei ein langsamer Prozess, da der Sehfarbstoff in seinen aktiven Zustand gewandelt werden muss; es dauert bei Zapfen etwa 10 Minuten und bei Stäbchen rund 30 Minuten, bis sie vollständig an dunkle Lichtverhältnisse angepasst sind. Die Anpassung an helle Lichtverhältnisse wird dagegen schon in Sekundenbruchteilen[5] wirksam und ist je nach Adaptionstyp in bis zu 6 Minuten optimal;[6] sie ist auch als Schutz vor Netzhautschäden durch übermäßig starkes Licht zu verstehen.
Einen zusätzlichen Anpassungsvorgang stellt die veränderte räumliche Summation dar, bei der die Fläche der Netzhaut, aus der eine Ganglienzelle der Netzhaut erregende Impulse erhalten kann, unter dem Einfluss retinaler Verschaltungen (z. B. durch laterale Hemmung) bei ansteigender Leuchtdichte abnimmt. Umgekehrt kann bei sinkender Leuchtdichte eine höhere Anzahl von Photorezeptoren des rezeptiven Feldes zur Bildung von Aktionspotentialen beitragen, die über den Neuriten im Sehnerv weitergeleitet werden.
Bei niedrigen Leuchtdichten können daneben eine Verlangsamung der Augenbewegungen bzw. eine verlängerte Fixationsdauer, die zu einer zeitlichen Summation führen, ebenfalls als Anpassungmodus verstanden werden.
Retinale Adaptationsprozesse, deren Wirkung auf bestimmte Netzhautareale beschränkt betrachtet wird, werden oft als lokale Adaptation bezeichnet und liegen beispielsweise dem Troxler-Effekt zugrunde. Bei besonderer Ausprägung führen sie nachwirkend zu einem Nachbild, das auch beim sogenannten Sukzessivkontrast beobachtet werden kann – bis dieses aufgrund retinaler Adaptation wieder verschwindet.
Chromatische Adaptation
Da die Netzhaut mit verschiedenen Typen von lichtempfindlichen Zellen ausgestattet ist, die für unterschiedliche Spektralbereiche empfänglich sind, kann durch Adaption auch der „Weißabgleich“ des Auges erledigt werden, die chromatische Adaptation. Wenn in der neuen Lichtsituation eine andere Farbtemperatur vorherrscht, z. B. durch einen verstärkten Rotanteil, dann werden die rotempfindlichen Zellen ihre Empfindlichkeit im Verhältnis zu den anderen verringern. Als Resultat empfindet der Betrachter eine weiße Fläche anschließend ebenfalls wieder als weiß, obwohl sie eine proportional erhöhte Menge roten Lichtes reflektiert. Adaptive Farbverschiebung ist der Unterschied in der wahrgenommenen Objektfarbe aufgrund einer Änderung der chromatischen Adaptation.
Hell- und Dunkeladaptation
Hell- und Dunkeladaptation der Wirbeltiere sind an die Retinomotorik gebunden (Bewegung der Pigmentepithelzellfortsätze und der Außenglieder der Photorezeptoren). Diese Wanderungsprozesse sind wahrscheinlich nur bei Tieren und nicht beim Menschen nachweisbar.[7] Helladaptation ist der Spezialfall des Tagsehens, wenn das gesamte visuelle System sich an Leuchtdichten oberhalb 3,4 cd/m2 angepasst hat. Dunkeladaptation ist der Spezialfall, wenn das visuelle System sich an Leuchtdichten unter 0,034 cd/m2 angepasst hat. Ein sehr offensichtliches Beispiel der (quantitativen) Adaptation kann beobachtet werden, wenn eine Person sich aus der vollen Sonne in ein Gebäude hineinbewegt. Die visuelle Umgebung im Gebäude wird zuerst nahezu schwarz erscheinen. Nach einigen Minuten ist die Person dann wieder in der Lage, Details zu erkennen (z. B. Zeitungstext zu lesen). Allerdings ist der Blick aus dem Fenster dann wieder unangenehm, da die großen Leuchtdichten draußen nun starke Blendung verursachen.
Die Dunkeladaptation beruht in erster Linie darauf, dass sowohl in den Zapfen als auch in den Stäbchen der Sehfarbstoff resynthetisiert wird. Da der Wiederaufbau jeweils langsamer als der Zerfall vor sich geht, bedarf die Dunkeladaptation eines längeren Zeitraums als die Helladaptation.[8]
Transiente Adaptation
Transiente Adaptation ist ein Begriff für den Spezialfall, der dann eintritt, wenn das Auge wiederholt zwischen einem hohen und einem niedrigen Lichtniveau hin und her wechseln muss. Dieses ist der Fall, wenn die Umgebung sehr hohe Kontraste aufweist, z. B. wenn ein Computermonitor (140–300 cd/m2) und eine sonnenbeschienene Fläche im Fenster (> 5000 cd/m2) ohne Kopfdrehung nebeneinander sichtbar sind. Dieser Zustand wird eine baldige Ermüdung der Augen zur Folge haben. Der Transient adaptation factor (TAF) ist ein englischsprachiger Begriff und bezeichnet die relative Reduktion des wahrnehmbaren Kontrastes durch die Readaptation zwischen unterschiedlich hellen Umgebungen.
Siehe auch
Einzelnachweise
- Stefan Silbernagl, Agamemnon Despopoulos: Taschenatlas Physiologie, 8. Auflage, Thieme Verlag, 2012, ISBN 978-3-13-567708-8, S. 374.
- Josef Dudel, Randolf Menzel, Robert F. Schmidt: Neurowissenschaft: vom Molekül zur Kognition, 2. Auflage, Springer-Verlag, 2001, ISBN 3-540-41335-9, Kapitel 17, S. 394ff.
- Werner Kahle u. a.: dtv-Atlas der Anatomie. Band 3, Deutscher Taschenbuchverlag, München 1978, ISBN 3-423-03019-4, S. 312.
- Walter Baumgartner: Klinische Propädeutik der Haus- und Heimtiere. Georg Thieme, Stuttgart 2009, ISBN 978-3-8304-4175-5, S. 413.
- James T. Fulton: Light & Dark Adaptation in human vision. auf: neuronresearch.net
- Theodor Axenfeld (Begr.); Hans Pau (Hrsg.): Lehrbuch und Atlas der Augenheilkunde. Gustav Fischer Verlag, Stuttgart 1980, ISBN 3-437-00255-4, S. 54.
- Wolf D. Keidel: Sinnesphysiologie. Teil I: Allgemeine Sinnesphysiologie; Visuelles System. Springer, Berlin / Heidelberg / New York 1976, ISBN 3-540-07922-X, S. 160.
- Gerhard Thews, Ernst Mutschler, Peter Vaupel: Anatomie, Physiologie, Pathophysiologie des Menschen. 5. Auflage. Wissenschaftliche Verlagsgesellschaft, Stuttgart 1999, ISBN 3-8047-1616-4, S. 738f.