Japanischer Bergmaulwurf
Der Japanische Bergmaulwurf (Oreoscaptor mizura) ist eine Säugetierart aus der Familie der Maulwürfe (Talpidae). Es handelt sich um den kleinsten Vertreter der Altweltmaulwürfe in Asien. Äußerlich kennzeichnet er sich durch ein schiefergraues Fell, einen vergleichsweise langen Schwanz und einen nackten Hautfleck hinter dem Nasenspiegel. Sein Verbreitungsgebiet umfasst die Bergregionen der japanischen Insel Honshū. Die Tiere bewohnen dort Waldlandschaften in mittleren bis höheren Lagen. Allgemein gilt die Art als selten. Dadurch liegen nur wenige Informationen zur Lebensweise vor. Die Hauptnahrung besteht vornehmlich aus Insekten. Außerdem legen die Tiere Nester aus Pflanzen an. Der Japanische Bergmaulwurf wurde im Jahr 1880 wissenschaftlich beschrieben. Im Verlauf des 20. Jahrhunderts galt er zumeist als Mitglied der Südostasiatischen Maulwürfe. Morphologische und genetische Untersuchungen sprechen aber für einen Verweis in eine eigenständige Gattung innerhalb der Altweltmaulwürfe. Aus diesem Grund wurde im Jahr 2016 die Gattung Oreoscaptor eingeführt. Der Bestand des Japanischen Bergmaulwurfs ist nicht bedroht.
| Japanischer Bergmaulwurf | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Japanischer Bergmaulwurf (Oreoscaptor mizura) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name der Gattung | ||||||||||||
| Oreoscaptor | ||||||||||||
| Kawada, 2016 | ||||||||||||
| Wissenschaftlicher Name der Art | ||||||||||||
| Oreoscaptor mizura | ||||||||||||
| (Günther, 1880) |
Merkmale
Habitus
Der Japanische Bergmaulwurf ist mit einer Kopf-Rumpf-Länge von 8,2 bis 11,1 cm, einer Schwanzlänge von 2,0 bis 2,7 cm und einem Gewicht von 20,5 bis 32,0 g ein kleiner Vertreter der Maulwürfe und der kleinste aller asiatischen Altweltmaulwürfe. Der Schwanz weist eine rutenförmige Gestalt auf und ist nur spärlich behaart. Seine Länge entspricht etwa 19 bis 29 % der Länge des restlichen Körpers. Dadurch ist er verhältnismäßig lang, außerdem übertrifft er die Länge der Vorder- und Hinterfüße, letztere besitzen Maße von 1,3 bis 1,6 cm, erstere sind durchschnittlich 1,4 cm lang und 1,3 cm breit. Das Rückenfell zeigt einen grauen bis dunkel schieferfarbenen Ton. Am Rücken setzt es sich aus über 1000 Einzelhaaren zusammen, darunter sind aber nur sehr wenige Leithaare (0,4 %).[1] Die Schnauze ist lang und breit im Vergleich zu den Südost- und Ostasiatischen Maulwürfen (Euroscaptor und Mogera) und von weißlichen Haaren bedeckt. Auf der Oberseite hebt sich ein nackter, dreieckiger bis rautenförmiger Bereich ab, den eine langgestreckte Grube in der Mitte kennzeichnet. Hervorgerufen wird diese durch die seitlich stark angeschwollenen Lippen. Der Penis ist kurz und dick, etwa 5,5 mm lang und 2,2 mm breit und dadurch deutlich unterschiedlich zu dem der Eurasischen Maulwürfen (Talpa).[2][3][4]
Schädel- und Gebissmerkmale
Der Schädel ist klein und grazil gebaut. Seine Länge variiert zwischen 25,0 und 28,0 mm. Das Rostrum besitzt eine schlanke Gestalt auf. Es verläuft zwischen dem Eckzahn und dem hinteren Prämolaren relativ parallel, verjüngt sich nach vorn und verbreitert sich nach hinten. Das Foramen infraorbitale öffnet sich oberhalb des zweiten Molaren. Die Jochbögen sind schwach entwickelt und kurz. Im Vergleich zu den Südostasiatischen Maulwürfen ist der hintere Schädelabschnitt breit mit einer Hirnschädelbreite, die gut 50 % der Länge aufweist. Der Unterkiefer ist ebenfalls schlank. Gut zwei Drittel seiner Länge werden von den Zähnen beansprucht. Dadurch erscheint der hintere Abschnitt sehr kurz, was wiederum einen Unterschied zu den Südostasiatischen Maulwürfen darstellt. Der Kronen- und der Gelenkfortsatz sind jeweils schmal ausgebildet, der Winkelfortsatz dagegen ist breit und dünn. Das Gebiss besteht aus 44 Zähnen, die Zahnformel lautet: . Die obere Reihe der Schneidezähne formt ein V. Hier ist der innerste Schneidezahn am größten ausgebildet, unter den oberen Prämolaren stellt der zweite den kleinsten, der vierte den größten dar. In der unteren Gebissreihe sind die Schneidezähne und der Eckzahn relativ gleich groß und der vorderste Prämolar übertrifft den zweiten in seinen Ausmaßen. Die Molaren zeigen sich allgemein verhältnismäßig groß und zeichnen sich im unteren Gebiss durch fünf Haupthöcker aus. Die obere Zahnreihe ist 10,4 bis 12,1 mm lang, die untere 9,4 bis 11,5 mm. Davon nehmen die Mahlzähne oben etwa 44 %, unten über 50 % der Gesamtlänge ein.[3][4]

Skelettmerkmale
Der Japanische Bergmaulwurf besitzt einen sogenannten mogeriden Beckenaufbau, das bedeutet, dass am Kreuzbein die Öffnungen des vierten und fünften Wirbels durch eine Knochebrücke auf der hinteren Seite überdeckt werden. Das stimmt mit den Ostasiatischen Maulwürfen (Mogera) überein, weicht aber von den Südostasiatischen Maulwürfen (Euroscaptor) ab, die ein caecoidales Becken besitzen (die Öffnungen der hinteren Kreuzbeinwirbel sind zumeist frei).[5][6][7]
Eimersches Organ
Das Eimersche Organ als feinfühliges Tastwerkzeug, das bei den meisten grabenden Maulwürfen vorkommt und aus zahlreichen Rezeptoren auf der unbehaarten Nasenspitze besteht, ist beim Japanischen Bergmaulwurf nur wenig untersucht. Allerdings sind auch bei diesem buckelartige Aufwölbungen dokumentiert, die rund 50 μm Durchmesser haben und in die jeweils Nervenbahnen enden.[8]
Genetische Merkmale
Der diploide Chromosomensatz lautet 2n = 36. Es treten 4 metazentrische, 3 submetazentrische, 2 subteleozentrische und 8 acrozentrische Chromosomenpaare auf. Das X-Chromosom ist submetazentrisch, das Y-Chromosom fleckenartig.[9][4]
Verbreitung und Lebensraum
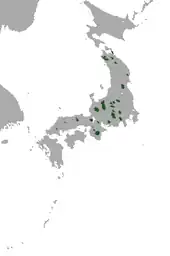
Das Vorkommen des Japanischen Bergmaulwurfs liegt auf der größten japanischen Insel, Honshū, etwa von den Regionen Chūgoku im Süden bis Tōhoku im Norden. Dort bewohnt er Nadel- und Laubwälder der gemäßigten alpinen und subalpinen Zone in Höhenlagen von 500 bis 2400 m. In Einzelfällen wurden Tiere auch in nur 270 m Höhenlage beobachtet.[10] Aufgrund der nur geringen Pflanzendecke und dem eher eingeschränkten Nahrungsangebot in diesen Landschaften ist die Populationsdichte gering. Die Art wird dadurch selten gesichtet. Im westlichen Teil seines Verbreitungsgebietes tritt sie sympatrisch mit dem Japanischen Maulwurf (Mogera wogura) auf.[11][4]
Lebensweise
Territorialverhalten
Aufgrund der seltenen Sichtungen des Japanischen Bergmaulwurfs sind Informationen zu seiner Lebensweise eher rar. Es wird vermutet, dass er einzelgängerisch auftritt und teils unterirdisch (semi-fossorial) lebt mit häufigen Aufenthalten an der Erdoberfläche. Die Tiere graben Kammern von bis zu 40 cm Durchmesser, die in der Regel nur einen Eingang haben. Die zugehörigen Tunnel und Gänge haben Weiten von 2,4 bis 4,3 cm. In den Kammern legen sie Nester aus breitblättrigem Pflanzenmaterial an, das nicht zerkleinert wird. Ein bei Kyoto untersuchtes Nest hatte einen Durchmesser von 18 cm und eine Höhe von 15 cm. Zu den bevorzugten Lagen der Nester gehören Hänge unter Buchen-, Weiden- und Birkengewächsen. Häufig schnüffelt der Japanische Bergmaulwurf hörbar beim Betreten des Unterschlupfes. Teilweise wird die Anwesenheit eines Tieres durch Pilze aus der Gattung der Fälblinge angezeigt, welche bevorzugt auf den Latrinen in der Nähe der Unterschlüpfe wachsen. Untersuchungen in der gleichen Region zwischen 1977 und 1992 erbrachten, dass der Japanische Bergmaulwurf die immer gleichen Nestplätze aufsucht, wahrscheinlich in einer Abfolge von mehreren Generationen. Eventuell resultiert dieses Verhalten aus der Minimierung der energieraufwendigen Grabetätigkeiten. Eine weitere Erklärung dafür könnte die reinigende Funktion der Pilze auf den Latrinen sein, die somit die unmittelbare Umgebung der Nester für die Maulwürfe über längere Zeiträume bewohnbar halten, wodurch eine Art symbiotischer Gemeinschaft entsteht.[12][13][14][4]
Ernährung
Der Japanische Bergmaulwurf ernährt sich überwiegend von Wirbellosen. Untersuchungen von drei Mageninhalten aus den Japanischen Alpen erbrachten vornehmlich Reste von Insekten. Sie stellten insgesamt zwei Drittel der Nahrungsmenge. Darunter befanden sich Larven von Laufkäfern und Zweiflüglern sowie größere Mengen unbestimmbaren Materials. Des Weiteren wurden auch Teile von Hundertfüßern, Wenigborstern und Egeln gefunden.[11] Tiere in Gefangenschaft fraßen zudem Regenwürmer und Mehlwürmer.[4]
Fortpflanzung
Über die Fortpflanzung ist kaum etwas bekannt. Drei Neugeborene wurden im Mai 2007 in einem Nest aufgefunden, es war die erste Beobachtung von Nachwuchs beim Japanischen Bergmaulwurf.[15] Dadurch besteht ein Wurf möglicherweise aus bis zu drei Jungen. Wahrscheinlich verlässt der Nachwuchs im Zeitraum von Juni bis August das mütterliche Nest, was Kadaverfunde befürworten.[4]
Systematik
Innere Systematik der Eigentlichen Maulwürfe nach He et al. 2016[18]
|
Der Japanische Bergmaulwurf ist eine Art aus der Gattung Oreoscaptor und gilt als deren einziger Vertreter. Die Gattung gehört in das nähere Verwandtschaftsumfeld der Ostasiatischen Maulwürfe (Mogera) sowie der Südostasiatischen Maulwürfe (Euroscaptor). Alle drei Gattungen sind wiederum in die Tribus der Eigentlichen Maulwürfe (Talpini) und der Familie der Maulwürfe (Talpidae) eingebettet. Die Eigentlichen Maulwürfe umfassen dabei die zumeist grabenden Angehörigen der Maulwürfe, während andere Vertreter der Familie dem gegenüber nur teilweise unterirdisch leben, sich oberirdisch fortbewegen oder eine semi-aquatische Lebensweise verfolgen. Laut molekulargenetischen Analysen spalteten sich die Eigentlichen Maulwürfe im Oberen Eozän vor rund 34 bis 36 Millionen Jahren von den anderen Triben der Maulwürfe ab. Eine stärkere Diversifizierung der Talpini ist seit dem Mittleren Miozän vor rund 12 Millionen Jahren zu verzeichnen. Von den Ostasiatischen Maulwürfen trennte sich der Japanische Bergmaulwurf gemäß der genetischen Daten wahrscheinlich im Oberen Miozän vor 7 bis 8 Millionen Jahren ab.[19][20][21][22][18]
Teilweise werden innerhalb der Art des Japanischen Bergmaulwurfs mehrere Unterarten unterschieden:[3][4]
- O. m. hiwaensis (Imaizumi, 1955); von Hiwa-cho in der Präfektur Hiroshima, teilweise auch aus den Präfekturen Shimane, Okayama und Hyogo berichtet
- O. m. mizura (Günther, 1880); Nominatform
- O. m. ohtai (Imaizumi, 1955); vom Berg Chogatake in der Präfektur Nagano, teilweise auch aus der Präfektur Aomori berichtet
Die einzelnen Unterarten werden nicht vollständig anerkannt. Ihre Unterscheidung erfolgt auf der Grundlage der Fellfärbung, der Größe und der relativen Schwanzlänge. Die genauen Verbreitungsgrenzen sind allerdings unklar. Einige japanische Forscher wie Yoshinori Imaizumi erhoben O. m. hiwaensis Ende des 20. Jahrhunderts auch in den Artstatus, allerdings sind bisher nur insgesamt wenige Individuen bekannt beziehungsweise nur ungenügende Merkmalsstudien durchgeführt worden, um dies zu validieren.[3] Einzelne genetische Analysen weisen den Japanischen Bergmaulwurf als stark divers aus, die Aufsplitterung setzte im Unteren Pliozän vor 4,1 Millionen Jahren ein.[20] In der Regel wird die Art als monotypisch geführt.[4]
Forschungsgeschichte

Die Erstbeschreibung der Art stammt von dem deutschen Zoologen Albert Günther und wurde im Jahr 1880 verfasst. Hierin führte Günther den Japanischen Bergmaulwurf mit der wissenschaftlichen Bezeichnung Talpa mizura und verwies ihn damit zu den Eurasischen Maulwürfen (Talpa). Ihm lag dafür ein Individuum vor, das ihm neben der einheitlich schiefergrauen Fellfärbung besonders durch den langen Schwanz im Vergleich zum damals bekannten Japanischen Maulwurf (Mogera wogura) auffiel. In seiner kurzen Notiz konnte er aber aufgrund der schlechten Erhaltung keine weiteren artbestimmenden Merkmale feststellen.[23] Als Herkunftsgebiet nannte Günther die Umgebung von Yokohama. Spätere Autoren nahmen dies als Verwechslung an, da der Japanische Bergmaulwurf dort nicht vorkommt. Teilweise werden aber auch das Tanzawa-Bergland oder die Region um Hakone, beides in der Präfektur Kanagawa, als Typusgebiet der Art angegeben.[3]
Nahezu ein halbes Jahrhundert wurde der Japanische Bergmaulwurf danach nicht mehr gesichtet, einige Forscher vermuteten aber, dass es sich um einen Vertreter der Ostasiatischen Maulwürfe handelt. Erst in den 1920er bis 1940er wurden mehrere weitere Individuen teils in Museumsbeständen entdeckt. Yoshinori Imaizumi untersuchte diese und verwies die Art 1948 zur Gattung der Südostasiatischen Maulwürfe, die sich von den Ostasiatischen Maulwürfen durch die Anzahl der Zähne unterscheidet. Im gleichen Jahr vereinte Ernst Schwarz alle talpinen Maulwürfe Ost- und Südostasiens mit den Eurasischen Maulwürfen. Zudem ordnete er alle Formen der Region einer Art zu, die er mit Talpa micrura auswies, diese aber in mehrere Unterarten aufteilte. Den Japanischen Bergmaulwurf setzte er mit dem Japanischen Maulwurf gleich (Talpa micrura wogura nach Schwarz).[24] Dem schloss sich Imaizumi in den 1950er Jahren an, hob den Japanischen Bergmaulwurf jedoch auf Artniveau an. Der Eingliederung in die Südostasiatischen Maulwürfe folgten indes aber zahlreiche Fachwissenschaftler, so auch Hisashi Abe, der in den 1960er Jahren die Insektenfresser Japans studierte.[2][11] Dieser Status quo blieb für den Rest des 20. Jahrhunderts weitgehend erhalten und wurde unter anderem auch durch Ivo Grulichs Analysen unterstützt.[5][6] Verschiedene molekulargenetische Studien in den 2000er Jahren und später deckten auf, dass der Japanische Bergmaulwurf tatsächlich den Ostasiatischen Maulwürfen näher steht und deren Schwestergruppe bildet. Die Trennung der beiden Gruppen reicht dabei bis in das Obere Miozän zurück.[25][20][19][18][26] Der weit in die Vergangenheit datierende Split befürwortete nach Meinung der Wissenschaftler eine eigene generische Stellung für den Japanischen Bergmaulwurf. Dies zum Anlass nehmend und unter Einbeziehung weiterer morphologischer Studien führte der japanische Zoologen Shin-ichiro Kawada im Jahr 2016 die neue Gattung Oreoscaptor ein. Die Bezeichnung leitet sich aus den griechischen Wörtern ὄρος (oros für „Gebirge“ oder „Berg“) und σκάπτηρ (skapter für „Grabender“) her, kann also mit „Gebirgsgräber“ übersetzt werden. Sie bezieht sich somit auf den bevorzugten gebirgigen Lebensraum der Art.[3] Einige modernere Systematiken übernahmen im Folgenden die neue Zuweisung, so auch der achte Band des Standardwerks Handbook of the Mammals of the World, welcher die insektenfressenden Säugetiere behandelt.[4]
Stammesgeschichte
Es sind nur wenige Fossilfunde des Japanischen Bergmaulwurfs berichtet worden. Die ältesten datieren in das jüngere Mittelpleistozän, absolute Daten liegen bei 190.000 bis 180.000 Jahren vor heute. Einige Skelettelemente traten in der Ikumo-Höhle zu Tage, einer reichhaltigen Fundstelle in der Präfektur Yamaguchi im äußersten Süden von Honshū. Als Begleitfauna kommen hier weitere Insektenfresser, Nagetiere, Raubtiere und verschiedene Huftiere vor. Nahezu gleichalt sind einzelne Gebissreste aus den nahe gelegenen Ubi-Kosan-Steinbrüchen. Ein spät-jungpleistozänes Alter weisen wiederum einige wenige Funde aus den Aisawa-Steinbrüchen in der Präfektur Tochigi im zentralen Honshū auf. Zusätzliches Material stammt weitgehend aus holozänen Ablagerungen wie aus der Tanuki-ana-Höhle wiederum in der Präfektur Yamaguchi.[27]
Gefährdung
Die IUCN listet den Japanischen Bergmaulwurf als „nicht bedroht“ (least concern). Zwar gibt die Naturschutzorganisation die Art als selten und das Verbreitungsgebiet als teilweise fragmentiert an, doch geht sie nicht von einem gravierenden Rückgang des Gesamtbestandes aus. Allerdings kann es im westlichen Teil des Verbreitungsgebietes durch Landschaftszerstörung in Folge ackerbaulicher oder wasserwirtschaftlicher Tätigkeiten zu Beeinträchtigungen kommen. Der Japanische Bergmaulwurf kommt in mehreren Naturschutzgebieten vor, etwa im Nikkō-Nationalpark.[28]
Literatur
- Shin-ichiro Kawada: Morphological Review of the Japanese Mountain Mole (Eulipotyphla, Talpidae) with the Proposal of a New Genus. Mammal Study 41 (4), 2016, S. 191–205, doi: 10.3106/041.041.0404
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 614) ISBN 978-84-16728-08-4
Einzelnachweise
- Naohiko Sagara: Hairtypes of Japanese Insectivora, with special reference to the presence of straightguard hairs. Journal of Mammalogical Society of Japan 11, 1986, S. 57–64
- Hisashi Abe: Classification and biology of Japanese Insectivora (Mammalia). I. Studies on variation and classification. Journal of the Faculty of Agriculture, Hokkaido University 55, 1967, S. 191–265 ()
- Shin-ichiro Kawada: Morphological Review of the Japanese Mountain Mole (Eulipotyphla, Talpidae) with the Proposal of a New Genus. Mammal Study 41 (4), 2016, S. 191–205, doi:10.3106/041.041.0404
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 614) ISBN 978-84-16728-08-4
- Ivo Grulich: Zum Bau des Beckens (Pelvis), eines systematisch-taxonomischen Merkmales, bei der Unterfamilie Talpinae. Zoologické Listy 20, 1971, S. 15–28 ()
- Ivo Grulich: Zur Kenntnis der Gattungen Scaptochirus und Parascaptor (Talpini, Mammalia). Folia Zoologica 31, 1982, S. 1–20 ()
- Shin-Ichiro Kawada, Nguyen Truong Son und Dang Ngoc Can: A new species of mole of the genus Euroscaptor (Soricomorpha, Talpidae) from northern Vietnam. Journal of Mammalogy 93 (3), 2012, S. 839–850, doi:10.1644/11-MAMM-A-296.1
- Kenneth C. Catania: Epidermal Sensory Organs of Moles, Shrew-Moles, and Desmans: A Study of the Family Talpidae with Comments on the Function and Evolution of Eimer’s Organ. Brain Behavior and Evolution 56, 2000, S. 146–174
- Shin-ichiro Kawada, Masashi Harada, Yoshitaka Obara, Shuji Kobayashi, Kazuhiro Koyasu und Sen-ichi Oda: Karyosystematic Analysis of Japanese T alpine Moles in the Genera Euroscaptor and Mogera (Insectivora, Talpidae). Zoological Science 18 (7), 2001, S. 1003–1010, doi:10.2108/zsj.18.1003
- Naohiko Sagara, Toshiho Ueda, Fujio Nishida und Toshiro Masai: The existence of Euroscaptor mizura in Kutsuki, Shiga Prefecture, with special reference to its dwelling at lower altitudes. Honyurui Kagaku 48 (1), 2008, S. 31–38 ()
- Hisashi Abe: Classification and biology of Japanese Insectivora (Mammalia). II. Biological aspects. Journal of the Faculty of Agriculture, Hokkaido University 55 (4), 1968, S. 429–458 ()
- Naohiko Sagara, Shuji Kobayashi, Hidetoshi Ota, Toyoaki Itsubo und Hiroaki Okabe: Finding Euroscaptor mizura (Mammalia: Insectivora) and Its Nest from under Hebeloma radicosum (Fungi: Agaricales) in Ashiu, Kyoto, with Data of Possible Contiguous Occurrences of Three Talpine Species in This Region. Contributions from the Biological Laboratory, Kyoto University 27, 1989, S. 261–272 ()
- Naohiko Sagara, Hisashi Abe und Hiroaki Okabe: The persistence of moles in nesting at the same site as indicated by mushroom fruiting and nest reconstruction. Canadian Journal of Zoology 71, 1993, S. 1690–1693
- Naohiko Sagara und Yu Fukasawa: Inhabitant changes in long-term mole nesting at the same site, revealed by observing mushroom fruiting at the site. Mammalia 78 (3), 2014, S. 383–391, doi:10.1515/mammalia-2013-0073
- Naohiko Sagara: First observation of Euroscaptor mizura nestlings, and success in transferring one into captive-rearing. Honyurui Kagaku 49, 2009, S. 45–52 ()
- Donald W. Duszynski und Alice R. Wattam: Coccidian Parasites (Apicomplexa: Eimeriidae) from Insectivores. V. Ten Forms from the Moles of Japan (Euroscaptor, Mogera spp.) Journal of Protozoology 35 (1), 1988, S. 55–57
- Yasushi Yokohata: A brief review of the biology of moles in Japan. Mammal Study 30 (sp. 1), 2005, S. S25–S30
- Kai He, Akio Shinohara, Kristofer M. Helgen, Mark S. Springer, Xue-Long Jiang und Kevin L. Campbell: Talpid Mole Phylogeny Unites Shrew Moles and Illuminates Overlooked Cryptic Species Diversity. Molecular Biology and Evolution 34 (1), 2016, S. 78–87
- Kai He, Akio Shinohara, Xue-Long Jiang und Kevin L. Campbell: Multilocus phylogeny of talpine moles (Talpini, Talpidae, Eulipotyphla) and its implications for systematics. Molecular Phylogenetics and Evolution 70, 2014, S. 513–521, doi:10.1016/j.ympev.2013.10.002
- Akio Shinohara, Shin-ichiro Kawada, Nguyen Truong Son, Chihiro Koshimoto, Hideki Endo, Dang Ngoc Can, Hitoshi Suzuki: Molecular phylogeny of East and Southeast Asian fossorial moles (Lipotyphla, Talpidae). Journal of Mammalogy 95 (3), 2014, S. 455–466, doi:10.1644/13-MAMM-A-135
- Akio Shinohara, Shin-ichiro Kawada, Nguyen Truong Son, Dang Ngoc Can, Shinsuke H. Sakamoto und Chihiro Koshimoto: Molecular phylogenetic relationships and intra-species diversities of three Euroscaptor spp. (Talpidae: Lipotyphla: Mammalia) from Vietnam. Raffles Bulletin of Zoology 63, 2015, S. 366–375
- Anna A. Bannikova, Elena D. Zemlemerova, Paolo Colangelo, Mustafa Sözen, M. Sevindik, Artem A. Kidov, Ruslan I. Dzuev, Boris Kryštufek und Vladimir S. Lebedev: An underground burst of diversity – a new look at the phylogeny and taxonomy of the genus Talpa Linnaeus, 1758 (Mammalia: Talpidae) as revealed by nuclear and mitochondrial genes. Zoological Journal of the Linnean Society 175, 2015, S. 930–948
- Albert Günther: Notes on some Japanese Mammalia. Proceedings of the Zoological Society of London, 1880, S. 440–443 ()
- Ernst Schwarz: Revision of the Old‐World Moles of the Genus Talpa Linnaeus. Proceedings of the Zoological Society of London 118 (1), 1948, S. 36–48
- Akio Shinohara, Shin-ichiro Kawada, Masashi Harada, Kazuhiro Koyasu, Sen-ichi Oda und Hitoshi Suzuki: Phylogenetic relationships of the short-faced mole, Scaptochirus moschatus (Mammalia: Eulipotyphla), among Eurasian fossorial moles, as inferred from mitochondrial and nuclear gene sequences. Mammal Study 33, 2008, S. 77–82
- E. D. Zemlemerova, A. A. Bannikova, V. S. Lebedev, V. V. Rozhnov und A. V. Abramov: Secrets of the underground Vietnam: an underestimated species diversity of Asian moles (Lipotyphla: Talpidae: Euroscaptor). Proceedings of the Zoological Institute RAS 320 (2), 2016, S. 193–220
- Yoshinari Kawamura: Quaternary Rodent Faunas in the Japanese Islands (Part 1). Memoirs of the Faculty of Science, Kyoto University, Series of Geology and Mineralogy 53 (1/2), 1988, S. 31–348 ()
- Laginha Pinto Correia: Euroscaptor mizura. The IUCN Red List of Threatened Species 2016. e.T8385A22320533 (); zuletzt aufgerufen am 23. Juli 2020
Weblinks
- Euroscaptor mizura in der Roten Liste gefährdeter Arten der IUCN 2016. Eingestellt von: Laginha Pinto Correia, 2016. Abgerufen am 23. Juli 2020.