Amerikanischer Spitzmull
Der Amerikanische Spitzmull (Neurotrichus gibbsii) ist eine Säugetierart aus der Familie der Maulwürfe (Talpidae). Sie kommt im westlichen Nordamerika vor, wo sie vom südwestlichen British Columbia in Kanada entlang der Pazifikküste bis in den mittleren Teil von Kalifornien in den Vereinigten Staaten verbreitet ist. Die Tiere bewohnen eine Vielzahl von Landschaften, die sowohl bewaldete Tieflande bis höhere Gebirgslagen einschließen. Bevorzugt werden Regionen mit mächtiger Humusdecke. Es handelt sich beim Amerikanischen Spitzmull um den kleinsten Vertreter der nordamerikanischen Maulwürfe. Er ähnelt äußerlich anderen Angehörigen der Familie, zeigt aber auch gewisse Ähnlichkeiten zu Spitzmäusen. Besondere Kennzeichen finden sich in seiner verhältnismäßig längeren Schnauze und dem längeren Schwanz. Seine Gliedmaßen weisen einen schlankeren Bau auf, wodurch die Tiere weniger gut an eine grabende Lebensweise angepasst sind. Das Fell ist dunkelgrau bis schwarz gefärbt.
| Amerikanischer Spitzmull | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Amerikanischer Spitzmull (Neurotrichus gibbsii) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name der Tribus | ||||||||||||
| Neurotrichini | ||||||||||||
| Hutterer, 2005 | ||||||||||||
| Wissenschaftlicher Name der Gattung | ||||||||||||
| Neurotrichus | ||||||||||||
| Günther, 1880 | ||||||||||||
| Wissenschaftlicher Name der Art | ||||||||||||
| Neurotrichus gibbsii | ||||||||||||
| (Baird, 1857) |
Die Lebensweise ist nur in Teilen gut erforscht. Der Amerikanische Spitzmull ist tag- und nachtaktiv mit relativ kurz wechselnden Aktivitäts- und Ruhephasen. Er legt Tunnel und Pfade an, die wenig tief in das Erdreich reichen oder durch Laubfallschichten verlaufen. Dabei entstehen keine Maulwurfshügel. Möglicherweise treten die Tiere in kleinen Gruppen auf. Die Nahrung besteht überwiegend aus Regenwürmern und Insekten zuzüglich einigem pflanzlichen Material. Ein Großteil der aktiven Zeit wird mit der Nahrungssuche verbracht. Die Fortpflanzungsphase ist ausgedehnt, werdende Muttertiere tragen bis zu vier Embryonen aus. Die Jungen werden in einem Nest aufgezogen.
Die wissenschaftliche Erstbeschreibung der Art erfolgte im Jahr 1857. Die heute gültige Gattungsbezeichnung wurde im Jahr 1880 eingeführt. Die genauen Verwandtschaftsverhältnisse des Amerikanischen Spitzmulls innerhalb der Maulwürfe waren lange Zeit ungeklärt. Genetische Untersuchungen sprechen für nähere Beziehungen zu den Japanischen Spitzmullen und zum Langschwanzmaulwurf. Innerhalb der Art werden drei Unterarten unterschieden. Fossilfunde liegen bisher nicht vor. Aus dem näheren Verwandtschaftsumfeld des Amerikanischen Spitzmulls sind jedoch Reste überliefert, die bis in das Untere Miozän zurückreichen. Sie stammen zum größten Teil aus Europa und Ostasien. Der Bestand der Art ist nicht gefährdet.
Merkmale
Habitus
Der Amerikanische Spitzmull ist der kleinste Vertreter der Maulwürfe Nordamerikas. Seine Kopf-Rumpf-Länge beträgt 6,3 bis 7,1 cm. Der Schwanz wird 2,9 bis 5,0 cm lang, was etwa 35 % der Länge des restlichen Körpers entspricht. Das Gewicht variiert von 6 bis 14,5 g. Ein ausgeprägter Geschlechtsdimorphismus besteht nicht. Das äußere Erscheinungsbild weist gewisse Ähnlichkeiten mit dem der Spitzmäuse auf. Der Schwanz ist verhältnismäßig lang, der Kopf groß und die Schnauze gestreckt sowie spitz. Die Augen sind sehr klein, ihr Durchmesser liegt bei 0,7 bis 1 mm. Sie werden von einer durchsichtigen Haut bedeckt, wodurch keine Lidspalte besteht. Als einzigartiges Merkmal kann die Pigmentierung der Pars ciliaris retinae hervorgehoben werden, die als vorderer Abschnitt der Netzhaut den Ziliarkörper bedeckt.[1] Äußere Ohrmuscheln sind nicht erkennbar. Der Gehörgang ist schlitzartig in Kopflängsrichtung orientiert mit Ausmaßen von 4 × 2 mm. Die Nasenlöcher öffnen seitwärts. An der Nasenspitze tritt ein Bündel an borstenartigen Haaren von nur 0,5 mm Länge auf. Kurz vor den Augen wiederum wachsen acht Paare an Vibrissen mit Längen bis zu 12 mm. Das weiche, dichte Körperfell ist einheitlich dunkelgrau oder schwarz bis rußig blaugrau gefärbt. Die einzelnen Haare des Deckfells erreichen 10 mm Länge und sind etwa doppelt so dick wie jene der Unterwolle. Der Schwanz beginnt dick und verjüngt sich zur Spitze. Er ist mit Schuppen in ringförmiger Anordnung bedeckt, trägt aber auch ein lockeres Fellkleid aus 2,5 mm langen Haaren. An der Schwanzspitze besteht eine Quaste aus 4,4 mm langen Haaren. Die Füße sind moderat klein und bestehen aus je fünf Strahlen. Die nackte Handfläche ist länger (rund 5,4 mm) als breit (rund 4 mm). Alle Strahlen tragen Krallen, im Gegensatz zu jenen der grabenden Maulwürfe sind diese beim Amerikanischen Spitzmull schmal und wenig gebogen. Die Rückseite der Füße ist mit Schuppen bedeckt. Die Länge der Hinterfüße liegt bei 1,3 bis 1,8 cm. Weibliche Tiere haben fünf Zitzenpaare, von denen sich eins im Brust- und je zwei im Bauch- und Lendenbereich befinden.[2][3][4]
Schädel- und Gebissmerkmale
Der Schädel wird 21,1 bis 23,7 mm lang und am Hirnschädel 9,7 bis 10,5 mm breit. Kurz hinter den Orbitae engt er sich auf 4,9 bis 5,5 mm ein. Insgesamt wirkt er in Seitenansicht flach, im Bereich der Stirnhöhlen ist er nur merklich geschwollen. An der Schädelbasis zeigen sich die Paukenblasen unvollständig. Am Mittelohr sind die Gehörknöchelchen im Vergleich zu einigen grabenden Maulwürfen kaum vergrößert, der Hammer ist nicht hypertrophiert. Die gesamte Konfiguration steht möglicherweise mit einem Hören im höheren Frequenzbereich in Verbindung.[5][6] Das Gebiss setzt sich aus 36 Zähnen mit folgender Zahnformel zusammen: Der vorderste obere Schneidezahn ist vergrößert, die nachfolgenden sind sehr klein. Gleiches kann zu den unteren Pendants gesagt werden. Der obere Eckzahn übertrifft den letzten Schneidezahn an Größe und hat einen dreieckigen Querschnitt, ist jedoch seitlich verschmälert. In der unteren Zahnreihe entspricht der Eckzahn etwa dem vordersten Schneidezahn. Im hinteren Gebiss fehlen zwei Prämolaren, unklar ist jedoch ob neben dem definitiven Verlust des ersten Vormahlzahns der zweite oder dritte reduziert wurde. Die verbliebenen Prämolaren und die Molaren sind relativ robust, wobei der jeweils letzte Mahlzahn sehr klein wirkt. Gelegentlich treten einzelne zusätzliche Zähne im Oberkiefer auf. Die Länge der oberen Zahnreihe variiert von 9.5 bis 10,5 mm.[7][8][3][4]

Skelettmerkmale
Im Skelettbau zeigt der Amerikanische Spitzmull einzelne Besonderheiten, die auf eine weniger starke Anpassung an eine grabende Lebensweise schließen lassen. Auffällig ist dies unter anderem am Schlüsselbein, das bei den grabenden Maulwürfen extrem kurz und hoch ist. Beim Amerikanischen Spitzmull weist es die zweieinhalbfache Länge der Höhe auf, im Vergleich dazu liegt der Längenwert bei den Westamerikanischen Maulwürfen (Scapanus) bei etwa drei Viertel der Höhe. Wiederum abweichend zu den grabenden Maulwürfen fehlt dem Schulterblatt des Amerikanischen Spitzmulls das Metacromion, ein Fortsatz am Acromion zur Aufnahme eines Teils des Musculus trapezius. Der Oberarmknochen ist deutlich schlanker als bei den Westamerikanischen Maulwürfen. Des Weiteren ist das Große Tuberkel neben dem Oberarmkopf weniger aufgebläht und der dort ansetzende Deltoid-Fortsatz verkleinert. Ebenso ist die innere Gelenkrolle am unteren Gelenkende weniger massiv. Elle und Speiche sind schlanker und länger als bei den Westamerikanischen Maulwürfen, sowie das Olecranon, der obere Gelenkfortsatz der Elle, kürzer.[9][10][3] Der für die grabenden Maulwürfe typische Präpollex („Vordaumen“), auch Os falciforme genannt, – ein stark vergrößertes Sesambein, das die Handfläche seitlich verbreitert – ist beim Amerikanischen Spitzmull eher klein ausgeprägt.[11]
Eimersches Organ
Auf der unbehaarten Nase des Amerikanischen Spitzmulles zeigen sich zahlreiche kleine buckelartige Aufwölbungen von rund 50 μm Durchmesser. Ihre höchste Erhebung wird durch eine Scheibe markiert. Die Buckel oder Papillen, die bienenwabenartig angeordnet sind, stellen Rezeptoren dar und werden als Eimersches Organ bezeichnet. Die jeweils darin endenden Nervenbahnen formen ein feinfühliges Tastwerkzeug, das vor allem bei grabenden Maulwürfen typisch ist. Insgesamt sind beim Amerikanischen Spitzmull rund 1560 Buckel ausgebildet. Sie verteilen sich regelhaft um die Nasenlöcher, ein jeweils kleiner seitlicher Fleck umfasst rund 50 Buckel.[12]
Genetische Merkmale
Der diploide Chromosomensatz lautet 2n = 38. Er setzt sich aus 15 zweiarmigen und 3 kleinen subtelozentrischen Autosomenpaaren zusammen. Das erste Chromosom ist nur unwesentlich größer als die nachfolgenden. Sowohl das Y-Chromosom als auch das X-Chromosom sind klein, ersteres ist fleckenartig, letzteres metazentrisch. Die Anzahl an Armen der Autosomen (fundamentale Anzahl) lautet 72.[13]
Verbreitung und Lebensraum
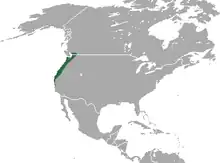
Das Verbreitungsgebiet des Amerikanischen Spitzmulls umfasst das westliche Nordamerika. Im Norden reicht die Grenze des Vorkommens bis in den südwestlichen Teil des kanadischen Bundesstaates British Columbia beidseits des Fraser Rivers. Das gesamte Verbreitungsareal nimmt hier etwa 10.000 km² ein.[14] Es zieht sich dann entlang der Westküste südwärts bis auf Höhe des Fremont Peak der Gabilan Range im Monterey County im mittleren Bereich des US-Bundesstaates Kalifornien. Im Osten begrenzen die Kaskadenkette und die Sierra Nevada in den Bundesstaaten Washington, Oregon und Kalifornien das Verbreitungsgebiet. Eine kleine Population lebt zudem auf Destruction Island, einer kleinen, nur 14 ha großen gras- und buschbewachsenen Insel rund 5,3 km vor der Westküste von Washington südwestlich der Mündung des Hoh Rivers.[15][3][4]
Der Lebensraum umfasst eine Vielzahl unterschiedlicher Habitate. Bevorzugt werden in tieferen Lagen Schwarzerden mit mächtiger Humusdecke von wenigstens 30 cm, die mit einer Schicht aus Blättern, Zweigen und Ästen bedeckt sein kann. Hierzu gehören vor allem Schluchten und Täler, deren Vegetation aus Ahorn-, Erlen- und Hartriegelgemeinschaften durchsetzt mit Pracht-Himbeeren oder Rotem Holunder besteht. Hinzu kommen Feuchtlandschaften in Flussufernähe bestanden mit Weidendickichten. Als weitere Landschaftsräume dienen Uferwälder aus Hartholz-Nadelbäumen, Douglasien und Gelbkiefern. Ebenso bewohnt der Amerikanische Spitzmull feuchtes Weiden- sowie Buschland und ist auch am unmittelbaren Flussufer zu finden. Generell meiden die Tiere aber trockene Gebiete. In Gebirgslagen können die Tiere in Gebieten mit dünner Flechtenbedeckung angetroffen werden. Hier reicht die Höhenverbreitung bis auf rund 2440 m über den Meeresspiegel. Aufgrund der Vielzahl an genutzten Landschaften kann der Amerikanische Spitzmull als Habitat-Generalist angesehen werden. Die Populationsdichte beträgt in zuträglichen Landschaften 12 bis 15 Individuen je Hektar.[16][3][14][4]
Lebensweise
Territorialverhalten
.jpeg.webp)
Die Lebensweise des Amerikanischen Spitzmulls ist nur bedingt gut untersucht. Die Tiere sind sowohl tag- als auch nachtaktiv. Hierbei wechseln sich kurze Perioden der Aktivität von 2 bis 18 Minuten Dauer von noch kürzeren Phasen der Ruhe mit 1 bis 8 Minuten Länge ab. Im Gegensatz zu vielen anderen Maulwürfen ist der Amerikanische Spitzmull häufiger an der Erdoberfläche anzutreffen. Sein Körperbau zeigt zwar weniger gute Grabeigenschaften als im Vergleich die teils sympatrisch vorkommenden Westamerikanischen Maulwürfen, dennoch legt er teils unterirdische Gänge an, die mitunter auch durch verrottendes Pflanzenmaterial am Erdboden verlaufen können. Die Eingänge sind stets offen. Oberflächennahe Pfade haben häufig eine Weite von 3,8 cm und sind nur 2,0 cm in den Boden eingetieft. Sie werden von Laub bedeckt und enthalten in gewissen Abständen kleine Löcher für die Luftzufuhr. In der Regel entstehen diese Gänge bei der Nahrungssuche und werden von den Tieren regelmäßig aufgesucht. Tiefer in den Untergrund reichende Tunnel sind mit 2,5 cm Durchmesser etwas schmaler. Insgesamt zeigen sich die Tunnelsysteme weniger komplex, maximale Tiefen werden bei 12,7 bis 30,5 cm erreicht. Häufig enthalten sie eine größere Kammer mit Ausmaßen von 12,7 × 7,5 cm. Diese befindet sich nahe dem Grundwasserspiegel, was eine gewisse Feuchtigkeit gewährt.[17][2][3][4]
In seiner Grabeweise ähnelt der Amerikanische Spitzmull den Westamerikanischen Maulwürfen. Die Erde wird beim Graben zur Seite gepresst, wobei lediglich ein Vorderfuß mit lateralen Bewegungen im Einsatz und der Körper um 45 ° gedreht ist. Der Aushub wird im Unterschied zu den Westamerikanischen Maulwürfen nicht an die Erdoberfläche gebracht, so dass kein Auswurfhügel (Maulwurfshügel) entsteht. Dennoch kann ein Individuum bis zu 209 g auf einmal an Erdmaterial bewegen, was gut dem 20fachen des Körpergewichts entspricht. An der Erdoberfläche läuft der Amerikanische Spitzmull relativ langsam und pausiert mehrfach. Die Krallen der Vorderfüße liegen auf der Fußsohle, so dass deren Rückseite den Boden berührt. Außerdem kann er gut schwimmen und setzt hierbei die kräftigen Beine alternierend paddelnd ein, wodurch undulierende Bewegungen entstehen. Außerdem klettert er in Sträuchern, geht hierbei bei Abwärtsrichtung mit dem Hinterteil voran. Kurze Ruhephasen verbringt ein Tier flach liegend. Zum Schlaf rollt es sich ein mit dem Kopf auf den Vorderbeinen liegend.[17][2][3][4]
Über das Sozialsystem des Amerikanischen Spitzmulls ist kaum etwas bekannt. Es wird angenommen, dass die Tiere in kleinen Gruppen leben und umherwandern. Größe und Stabilität von Aktionsräumen wurden bisher nicht genauer untersucht, einzelnen Angaben zufolge könnten sie aber bis zu 4100 m² groß sein.[18] Ebenso sind Lautäußerungen nicht dokumentiert, in Experimenten lösten Töne im höheren Frequenzbereich von 8 bis hin zum Ultraschallbereich von 30 kHz Reaktionen aus. Unter Bedrohung nehmen die Tiere eine Verteidigungsposition mit erhobenen Vorderbeinen und geöffnetem Maul ein. In der Regel fliehen sie jedoch unter eine Laubschicht und verhalten sich still.[17][2][3][4]
Ernährung und Energiehaushalt

Die Nahrung des Amerikanischen Spitzmulls setzt sich aus Wirbellosen und vereinzeltem Pflanzenmaterial zusammen. Nach Untersuchung von rund 40 Mageninhalten aus dem US-Bundesstaat Washington haben Regenwürmer mit 42 Volumenprozent den größten Anteil, gefolgt von Asseln und Larven von Insekten mit jeweils 36 beziehungsweise 12 Volumenprozent.[17] Ein ähnliches Ergebnis erbrachte die Analyse von nahezu einem Dutzend Mageninhalten aus Oregon. Hier dominierten Regenwürmer mit 49 Volumenprozent vor Insekten mit 20 Volumenprozent. Unter den Insekten konnten unter anderem Schnaken identifiziert werden. Weitere Beutetiere bestanden aus Hundertfüßern, Tausendfüßern und Weichtieren. Pflanzen erreichten einen Anteil von 0,6 Volumenprozent.[19] Tiere in Gefangenschaft fraßen auch Mäuse und verschiedene Lurchen, mieden aber Frösche und Salamander. Ebenso konsumierten sie zu gewissen Teilen Samen von Fichten, Kiefern und Douglasien sowie andere dargebotene Pflanzen wie Kräuter oder Sträucher.[20] Ein Großteil des Wasserbedarfs wird über die Nahrung gedeckt.[17][2][3][4]
Der Amerikanische Spitzmull verbringt nahezu die gesamte aktive Zeit mit der Nahrungssuche. Dabei hält er die lange bewegliche Nase in die Luft sowie auf den Boden und schwingt den Kopf hin und her. Aufgespürte Regenwürmer werden individuell durch die Vorderfüße gezogen, von einem Ende oder von der Seite aufgebissen oder in Stücke geteilt. Ein rund 10 g schweres Tier in menschlicher Obhut vertilgte einen 1,3 g schweren Regenwurm in gut 10 Minuten, in rund 12 Stunden konnte es etwa 14 g an Nahrung aufnehmen, was fast dem anderthalbfachen des eigenen Körpergewichts entspricht. Ein weiteres Individuum starb rund 11 Stunden nach dem Nahrungsentzug. Die Verdauung der Nahrung erfolgt rapide, bereits nach 45 Minuten finden sich Teile im ausgeschiedenen Kot wieder.[17][2][3][4]
Die durchschnittliche Körpertemperatur beträgt 38,4 °C, was höher ist als bei anderen untersuchten nordamerikanischen Maulwürfen. Sie ist jedoch abhängig von der Außentemperatur und kann entsprechend bei geringeren Umgebungswerten sinken. So wurde bei Außentemperaturen von 4,4 °C eine bis zu 6,8 °C niedrigere Körpertemperatur gemessen. Die Stoffwechselrate im Ruhezustand entspricht in etwa der der Spitzmäuse, liegt aber um das 2,3fache über dem Wert, der für ein vergleichbar großes Säugetier zu erwarten wäre. Entsprechend der Körpertemperatur kann sich auch der Umsatz bei fallenden Umgebungstemperaturen um bis zu 28 % verringern, so dass ein Hypometabolismus eintritt.[21] Die Atmungsfrequenz beträgt 3,4 Züge je Sekunde.[17][4]
Fortpflanzung
Über die Fortpflanzung liegen nur wenige Informationen vor. Die Paarungszeit ist möglicherweise ausgesprochen lang, in Washington wurden paarungswillige Männchen im Februar, stillende Muttertiere im September angetroffen. Feldbeobachtungen zufolge findet der überwiegende Teil der Paarungen aber möglicherweise zwischen den Monaten März und Mai statt. Männchen sondern dann einen moschusartigen Duft ab. Die Länge der Tragzeit ist nicht bekannt, währt aber wohl länger als 15 Tage. Trächtige Weibchen tragen zwischen einem und vier Embryonen aus, der Durchschnitt liegt bei drei. Neugeborene sind 26 bis 27 mm lang abzüglich eines rund 5 mm langen Schwanzes, das Gewicht variiert von 0,49 bis 0,67 g. Der Körper ist weitgehend nackt und besitzt eine dunkelviolette Färbung, Vibrissen fehlen und die Augen sind als dunkle Flecken erkennbar. Die Füße weisen noch keine ausgehärteten Krallen auf und sind eher fleischig.[22] Ein bisher singulär beobachtetes Nest mit Jungtieren in Washington bestand aus feuchten Weidenblättern und befand sich in einem Baumstumpf, gut 60 cm oberhalb der Erdoberfläche. Zum Nest führte ein Gang, der durch den Baumstumpf gegraben war.[17][2][3][4] Weibliche Individuen des Amerikanischen Spitzmulls besitzen neben einer penisartigen Klitoris auch Zwitterdrüsen und können daher als Hermaphroditen auftreten. Ähnliches ist bereits von den Eigentlichen Maulwürfen bekannt, nicht aber von den näher verwandten Japanischen Spitzmullen.[23]
Fressfeinde, Kommensalen und Parasiten
Aufgrund seiner teils oberirdischen Lebensweise wird der Amerikanische Spitzmull von zahlreichen Fressfeinden erbeutet. Als bedeutendster Prädator tritt die Schleiereule in Erscheinung. Nach Untersuchungen von Gewöllen aus dem zentralen Kalifornien ist der Maulwurfsvertreter ein regelmäßiges, wenn auch nicht sehr häufiges Opfer des Vogels, dessen Beutespektrum ansonsten von der Kalifornischen Wühlmaus und der Gebirgs-Taschenratte dominiert wird.[24] Ähnliches kann auch zum Virginia-Uhu, zum Sägekauz und zur Waldohreule gesagt werden.[25][26][27] Bei 380 analysierten Kotresten des Fischermarders aus Oregon fand sich der Amerikanische Spitzmull mit einem Anteil von 2,7 % unter den Beutetieren wieder, wobei er fast ausschließlich von weiblichen Tieren erlegt wurde.[28] Weitere Beutegreifer stellen der Rotschwanzbussard, der Kojote, der Waschbär und verschiedene Schlangen dar, zu letzteren gehört unter anderem die Nordwestliche Strumpfbandnatter.[2][3]
Der Amerikanische Spitzmull tritt mit verschiedenen anderen Säugetieren gemeinsam in Erscheinung. Dazu zählen Rotzahnspitzmäuse, Feldmäuse, Weißfußmäuse und Rötelmäuse, aber auch Stummelschwanzhörnchen und einige Vertreter der Westamerikanischen Maulwürfe. Zu einigen der Arten besteht ein Konkurrenzverhalten etwa um Nahrungsressourcen, bei anderen kommt es unter anderem zur gemeinsamen Nutzung von Gängen und Tunneln.[17][3]
Es sind zahlreiche Parasiten belegt, die den Amerikanischen Spitzmull befallen. Äußere Schmarotzer finden sich vor allem unter den Milben, so etwa Raubmilben wie Laelaps und Haemogamasus, aber auch Vertreter der Gattungen Eadiea und Pygmephorus aus der Gruppe der Prostigmata.[29][19][30] Des Weiteren sind Flöhe mit Corypsylla und Epitedia sowie Zecken mit Ixodes und Androlaelaps dokumentiert.[31] Als innere Parasiten können überwiegend verschiedene Würmer genannt werden. So ließen sich Saugwürmer wie Aspalacitrema identifizieren, zusätzlich auch Bandwürmer wie Liniscus und Hymenolepis sowie Fadenwürmer wie Porrocaecum.[32][33][3]
Systematik
Innere Systematik der Maulwürfe nach He et al. 2016[34]
|
Der Amerikanische Spitzmull ist eine Art aus der Gattung Neurotrichus. Er stellt das einzige rezente Mitglied dar, wodurch die Gattung als monotypisch gilt. Sowohl die Art als auch die Gattung werden zur Familie der Maulwürfe (Talpidae) verwiesen. Innerhalb der Maulwürfe formen sie die ebenfalls monotypische Tribus der Neurotrichini,[35][35][34] werden in einigen Systematiken aber auch zu den Japanischen Spitzmullen (Urotrichini) geordnet.[4] Laut molekulargenetischen Untersuchungen sind beide Gruppen eng miteinander verwandt, definierende Unterschiede finden sich jedoch vor allem im Gebissaufbau.[8][36][35] Gemeinsam mit den Japanischen Spitzmullen und dem Langschwanzmaulwurf (Scaptonychini) bildet der Amerikanische Spitzmull eine monophyletische Einheit. Alle drei Gruppen schließen Maulwürfe ein, die sich nur teilweise an ein unterirdisches Leben angepasst haben. Die Trennung von den anderen Linien der Maulwürfe fand bereits im Oberen Eozän vor rund 37 Millionen Jahren statt, während sich die Aufspaltung der Gruppe im Zeitraum von vor 30 bis 27 Millionen Jahren im Oligozän ereignete. Als nächstverwandte Gruppe kann eine gemeinsame Klade angesehen werden, die aus den Eigentlichen Maulwürfe (Talpini), den Desmanen (Desmanini) und dem Sternmull (Condylurini) besteht. Übergeordnet sind alle genannten Maulwurfslinien der Unterfamilie der Altweltmaulwürfe (Talpinae) zuzuweisen, letzterer gehören zusätzlich noch die Neuweltmaulwürfe (Scalopini) an. Die Altweltmaulwürfe vereinen sowohl unterirdisch lebende als auch an eine semi-aquatische Lebensweise angepasste Tiere aus Eurasien und aus Nordamerika.[34]
Dem Amerikanischen Spitzmull werden drei Unterarten zugewiesen. Aufgrund des monotypischen Charakters von Gattung und Tribus ergibt sich folgende Gliederung:[3][35][4][37]
- Tribus: Neurotrichini Hutterer, 2005
- Gattung: Neurotrichus Günther, 1880
- Neurotrichus gibbsii (Baird, 1857)
- N. g. gibbsii (Baird, 1857); im südwestlichen British Columbia sowie im westzentralen Washington, westlichen Oregon und nordwestlichen Kalifornien
- N. g. hyacinthinus Bangs, 1897; im westzentralen Kalifornien
- N. g. minor Dalquest & Burgner, 1941; im äußersten südwestlichen British Columbia und entlang der Küstenzone von Washington
Verschiedentlich wurden der Gattung Neurotrichus auch einzelne Fossilformen beigeordnet. Hierzu gehört etwa Neurotrichus columbianus, benannt im Jahr 1968 durch J. Howard Hutchison anhand eines Unterkiefers und einzelner Zähne vom McKay Reservoir im nördlichen Oregon.[38] Die in das Obere Miozän datierenden Funde werden nach weiteren Untersuchungen aber eher den Neuweltmaulwürfen zugerechnet und sind wohl mit Yanshuella näher verbunden.[36] Aus Europa stammen wiederum Neurotrichus polonicus und Neurotrichus scokzeni, beide Arten gehen auf Stanisław Skoczeń zurück. Erstere wurde von ihm im Jahr 1980 über einige Unterkieferreste, Zähne und weitere Skelettelemente aus der pliozänen Fundstelle von Rębielice Królewskie im südlichen Polen definiert.[39] Andere Autoren sahen vorläufig für Neurotrichus polonicus eher Affinitäten zu Quyania, einem entfernteren Verwandten des Amerikanischen Spitzmulls.[36][40][41] Die letztere Art war ursprünglich 1993 von Skoczeń als Neurotrichus minor ausgewiesen,[42] später aber aufgrund von Namenskonflikten mit der Unterart N. g. minor in Neurotrichus skoczeni abgeändert worden.[43] Sie basiert auf einem ebenfalls pliozänzeitlichen Obermknochen aus der Umgebung von Częstochowa, wiederum südliches Polen. Ihre Stellung war weitgehend umstritten, im Jahr 2016 kreierten Gabrielle Sansalone und Kollegen für beide Arten die Gattung Rzebikia, die wie Quyania zum weiteren Verwandtschaftsumfeld des Amerikanischen Spitzmulls gehört.[44]
Neben der rezenten Gattung Neurotrichus enthält die Tribus demzufolge zwei fossile Vertreter:[36][41][44]
- Quyania Storch & Qiu, 1983
- Rzebikia Sansalone, Kotsakis & Piras, 2016
Forschungsgeschichte



_Neurotrichus_gibbsii.jpg.webp)
Die wissenschaftliche Erstbeschreibung des Amerikanischen Spitzmulls erfolgte durch Spencer Fullerton Baird im Jahr 1857. Als Basis hierfür diente ein junges Individuum, das drei Jahre zuvor von George Gibbs am White River in der Kaskadenkette des US-Bundesstaates Washington gesammelt und via George Suckley an Baird übergeben worden war. Der exakte Fundort ist nicht eindeutig, wurde aber mit nördlich des Mount Rainier angegeben, später engten Walter W. Dalquest und Robert L. Burgner die Typuslokalität auf den Naches Pass ein, der zur damaligen Zeit den europäischen Siedlern bekannt gewesen war.[45] Baird selbst erkannte zahlreiche Ähnlichkeiten zu den Japanischen Spitzmullen und zum Sternmull, hob aber gegenüber letzterem die längere Schnauze und die fehlenden Hautanhängsel an der Nase beim Amerikanischen Spitzmull hervor. Er verwies seine neue Art daher in die Gattung Urotrichus und benannte sie zu Ehren des Entdeckers des Holotyp-Exemplars mit U. gibbsii. Die Spitzmulle sah Baird als eine Art Übergangsform zwischen den eigentlichen Maulwürfen und den Spitzmäusen an.[46] Dreiundzwanzig Jahre später, 1880, unterstrich Albert Günther in einem Aufsatz über japanischen Säugetiere die Unterschiede zwischen dem Amerikanischen Spitzmull und den Japanischen Spitzmullen, wobei er sich unter anderem auf abweichende äußerliche Merkmale sowie auf die differierende Gebissausprägung berief. Letzteres speziell veranlasste ihn, die neue Gattung Neürotrichus für den Amerikanischen Spitzmull einzuführen.[47] Der Gattungsname wird dabei vom griechischen Wort νέος (neos) für „neu“ und der Gattungsbezeichnung Urotrichus abgeleitet.[48] Günther bevorzugte die phonetische Abtrennung der Vorsilbe neos vom Gattungshauptwort, eine Schreibweise, die bis weit in das 20. Jahrhundert von einigen Autoren beibehalten wurde. Die erste, heute korrekte Schreibweise des Gattungsnamens Neurotrichus ist aber bereits für das Jahr 1881 verzeichnet und erfolgte durch William Alexander Forbes.[49][37]
Bereits im Jahr 1896 führte Frederick W. True eine erste Revision des Amerikanischen Spitzmulls durch, in der er die Art zwar als uniform ansah, allerdings einen auffälligen Größenunterschied zwischen den Tieren im nördlichen und südlichen Verbreitungsgebiet erwähnte.[50] Im Jahr darauf trennte Outram Bangs neben der Nominatform eine zweite Unterart ab, die er N. g. hyacinthinus nannte. Diese zeichnete sich durch ihre durchschnittlich größeren Dimensionen aus, zudem gab Bangs ein dunkleres Fell mit einer teils grün oder violett schimmernden Unterwolle an. Das Belegaexemplar bestand aus einem alten Weibchen aus Nicasio im Marin County im zentralen Kalifornien.[51] Kurz darauf, im Jahr 1899, nutzte Clinton Hart Merriam ein großes ausgewachsenes Männchen vom Mount Shasta im Shasta County des nördlichen Kalifornien zur Etablierung der Unterart N. g. major.[52] Die Form erhielt aber nur wenig Anerkennung, da sie im Jahr 1915 von Hartley H. T. Jackson während einer erneuten Revision des Amerikanischen Spitzmulls mit N. g. gibbsii synonymisiert wurde.[7] Die dritte, heute anerkannte Unterart N. g. minor stammt von Walter W. Dalquest und Robert L. Burgner aus dem Jahr 1941. Ihr liegt ein männliches Individuum vom Puget Sound im nordwestlichen Washington zu Grunde, das durch seine kleineren Dimensionen hervorstach.[45] Neben den beiden Revisionen der Art in den Jahren 1896 und 1915 wurden noch weitere durchgeführt. Die bisher letzte erfolgte durch Neal Woodman im Jahr 2018.[37]
In der forschungsgeschichtlichen Vergangenheit wurde der Amerikanische Spitzmull teilweise mit den Neuweltmaulwürfen in Verbindung gebracht, die dann in der Regel auch die Japanischen Spitzmulle und den Langschwanzmaulwurf aufnahmen, so unter anderem bei George Gaylord Simpson in seiner generellen Taxonomie der Säugetiere 1945.[53] In einer Überarbeitung der Systematik der Insektenfresser verschob Leigh Van Valen im Jahr 1971 den Amerikanischen Spitzmull und die Japanischen Spitzmulle in die Tribus der Urotrichini und zeigte somit ein näheres Verwandtschaftsverhältnis an. Gleichzeitig schloss er den Langschwanzmaulwurf aus und schlug ihn der neuen Tribus der Scaptonychini zu.[54] Die Urotrichini waren bereits 1883 von George Edward Dobson etabliert worden, beinhalteten in dieser ursprünglichen Version neben den verschiedenen Spitzmullen zusätzlich auch den Langschwanzmaulwurf.[55] In der dritten Ausgabe des Leitwerkes Mammal Species of the World aus dem Jahr 2005 kreierte Rainer Hutterer die Tribus der Neurotrichini für den Amerikanischen Spitzmull und sonderte die Form somit aus den Urotrichini aus.[35] Als Begründung für diesen Schritt gab Hutterer neben den anatomischen Differenzen auch eine zwei Jahre zuvor veröffentlichte molekulargenetische Studie an, die in ihrem Ergebnis den Amerikanischen Spitzmull aus einem Nahverhältnis zu den Japanischen Spitzmullen ausschloss.[56] Nachfolgende genetische Untersuchungen berücksichtigten den Amerikanischen Spitzmull nur selten und blieben bezüglich der tatsächlichen verwandtschaftlichen Beziehungen teilweise ambivalent.[57][58][59] Dies trifft auch auf einige anatomische Analyse zu, jedoch rückten diese die Art teils näher an den Langschwanzmaulwurf heran.[60][61][62][63] Eine im Jahr 2016 veröffentlichte molekulargenetische Studie, basierend auf nahezu allen, zu diesem Zeitpunkt bekannten Gattungen der Maulwürfe, ermittelte für den Amerikanischen Spitzmull ein näheres Verhältnis mit den Japanischen Spitzmullen und dem Langschwanzmaulwurf und sieht alle drei Gruppen in einer monophyletischen Beziehung. Unangetastet blieb hierbei die jeweilige Stellung in einer eigenständigen Tribus.[34]
Stammesgeschichte
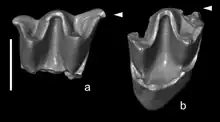
Der Fossilbericht der Neurotrichini ist bisher eher spärlich. Fast alle relevanten Funde sind aus Eurasien bekannt, wo die Gruppe möglicherweise ihren Ursprung hatte. Der bisher älteste Vertreter wurde mit Quyania berichtet, dessen bedeutendstes Material aus Ostasien stammt. Mehrere isolierte Zähne sind unter anderem aus der reichhaltigen Fossillandschaft von Aoerban im zentralen Bereich der chinesischen Autonomen Region Innere Mongolei entdeckt worden. Sie verteilen sich auf insgesamt vier fossilhaltige Lagen, das Untere und Obere Rote Muddestein-Schichtglied der Aoerban-Formation sowie das Balunhalagen-Bett und das Bilutu-Bett, welche einen Zeitraum vom Unteren zum Oberen Miozän abdecken.[64] Etwa zeitgleich zu den oberen Fundschichten datieren rund 50 Zähne aus Halajin Hushu im Norden der Inneren Mongolei.[65] Das für die Gattung definierende Material in Form von mehreren Unterkiefern, Zähnen und Langknochen erbrachte die Fundstelle Ertemte im Süden der Inneren Mongolei. Isolierte Zähne sind auch von der nahe gelegenen Lokalität Harr Obo beschrieben worden. Beide Fundstellen gehören in den Übergang vom Miozän zum Pliozän. Die kleinen Tiere besaßen noch ein umfangreicheres Gebiss als der heutige Amerikanische Spitzmull.[36] Abseits dieser ostasiatischen Funde konnte Quayania auch mit einigen wenigen Zähnen aus der Lokalität Tagay am Westufer der Insel Olchon im Baikalsee belegt werden. Mit einer Stellung im Übergang vom Unteren zum Mittleren Miozän entsprechen sie in etwa den unteren Fossillagen aus der Aoerban-Fundlandschaft.[66]
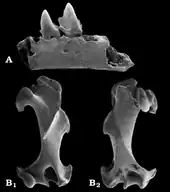
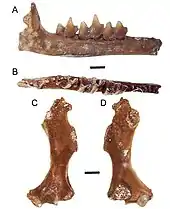
Weitere Fossilreste von Quyania liegen aus Europa vor. Hierunter fällt ein recht umfangreiches Ensemble von verschiedenen Fundstellen in Polen, deren bedeutendste sich mit Rębielice Królewskie im südlichen Landesteil befindet, hinzu kommen Podlesice, Kadzielnia oder Węże bei Częstochowa. Die ebenfalls aus Unterkieferteilen, Zähnen und postcranialen Skelettelementen bestehende Kollektion datiert in das ausgehende Pliozän und frühe Pleistozän.[41] Einzelne Funde lassen sich auch aus den Spaltenfüllungen von Petersbuch in Oberbayern der Gattung zuweisen. Mit einer Stellung in dem Beginn des Oberen Miozäns wäre dies der bisher älteste Beleg der Gattung in Europa.[67] Bereits im Jahr 1899 hatte Claude Gaillard einige Funde, darunter einen Oberarmknochen, aus der Fundstelle La Grive-Saint-Alban im südöstlichen Frankreich vorgestellt, sie aber dem Langschwanzmaulwurf zugeordnet.[68] Heute werden sie weitgehend zu Quyania verwiesen.[41] Die polnischen Fundstellen Rębielice Królewskie und Węże bargen mit Rzebikia noch die Reste einer weiteren Maulwurfform, die dem Amerikanischen Spitzmull nahesteht. Auch hierbei handelt es sich weitgehend um Gebissfragmente und Teile des Bewegungsapparates. Anhand der Konfiguration der Vorderbeine kann geschlossen werden, dass die Tiere über einige Grabungsbefähigungen verfügten.[39][42][44] Sehr ähnliches Material wurde des Weiteren aus Warschez im nördlichen Bulgarien berichtet.[40]
Aus Nordamerika liegen bisher nur wenige Reste vor. Einige Unterkieferteile und ein Oberarmknochen von der Gray Fossil Site im Washington County des US-Bundesstaates Tennessee stehen wahrscheinlich mit Neurotrichus in Verbindung, auch wenn einzelne Merkmale auf Quayania verweisen. Der fragmentierte Zustand der Funde erschwert eine exakte Beurteilung. Gebildet wurde die bedeutende Fossillagerstätte im Zeitraum des Oberen Miozäns und Unteren Pliozäns.[69][70] Vom Amerikanischen Spitzmull selbst hingegen sind keine Fossilfunde bekannt.[3]
Bedrohung und Schutz
Der Bestand des Amerikanischen Spitzmulls wird von der IUCN als „nicht gefährdet“ (least concern) eingestuft. Als Begründung gibt die Naturschutzorganisation die weite Verbreitung der Art an, zudem ist die Population relativ stabil. Größere Bedrohungen sind nicht bekannt. Lokal kam es aber zu einem Rückgang der Bestände durch die Einführung des Wildkaninchens, so etwa auf Destruction Island vor der Westküste des US-Bundesstaates Washington.[15] Der Amerikanische Spitzmull ist in mehreren Naturschutzgebieten präsent.[71][4]
Literatur
- L. N. Carraway und B. J. Verts: Neurotrichus gibbsii. Mammalian Species 387, 1991, S. 1–7
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 603) ISBN 978-84-16728-08-4
- Chris Maser, Bruce R. Mate, Jerry F. Franklin und C. T. Dyrness: Natural history of Oregon coast mammals. United States Department of Agriculture, Forest Service, General Technical Report PNW-133, 1981, S. 1–496 (S. 64–71)
- Ronald M. Nowak: Walker's Mammals of the World. 2 Bände. 6. Auflage. The Johns Hopkins University Press, Baltimore MD u. a. 1999, ISBN 0-8018-5789-9
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. A taxonomic and geographic Reference. 2 Bände. 3. Auflage. The Johns Hopkins University Press, Baltimore MD 2005, ISBN 0-8018-8221-4
Einzelnachweise
- Thomas H. Lewis: The anatomy and histology of the rudimentary eye of Neurotrichus. Northwest Science 57 (1), 1983, S. 8–15
- Chris Maser, Bruce R. Mate, Jerry F. Franklin und C. T. Dyrness: Natural history of Oregon coast mammals. United States Department of Agriculture, Forest Service, General Technical Report PNW-133, 1981, S. 1–496 (S. 64–71)
- L. N. Carraway und B. J. Verts: Neurotrichus gibbsii. Mammalian Species 387, 1991, S. 1–7
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 603) ISBN 978-84-16728-08-4
- Matthew J. Mason: Evolution of the Middle Ear Apparatus in Talpid Moles. Journal of Morphology 267, 2006, S. 678–695
- Daisuke Koyabu, Misato Hosojima und Hideki Endo: Into the dark: patterns of middle ear adaptations in subterranean eulipotyphlan mammals. Royal Society Open Science 4, 2017, S. 170608, doi:10.1098/rsos.170608
- Hartley H. T. Jackson: A review of the American moles. North American Fauna 38, 1915, S. 1–100 (S. 92–98)
- Alan C. Ziegler: Dental homologies and possible relationships of recent Talpidae. Journal of Mammalogy 52 (1), 1971, S. 50–68
- Berry Campbell: The shoulder anatomy of the moles. A study in phylogeny and adaptation. The American Journal of Anatomy 64, 1939, S. 1–39
- Charles A. Reed: Locomotion and Appendicular Anatomy in Three Soricoid Insectivores. The American Midland Naturalist 45 (3), 1951, S. 513–671
- Marcelo R. Sánchez-Villagra und Peter R. Menke: The mole’s thumb – evolution of the hand skeleton in talpids (Mammalia). Zoology 108, 2005, S. 3–12
- Kenneth C. Catania: Epidermal Sensory Organs of Moles, Shrew-Moles, and Desmans: A Study of the Family Talpidae with Comments on the Function and Evolution of Eimer’s Organ. Brain Behavior and Evolution 56, 2000, S. 146–174
- Shin-ichiro Kawada, Song Li, Ying-Xiang Wang, Orin B. Mock, Sen-ichi Oda und Kevin M. Campbell: Karyotype evolution of shrew moles (Soricomorpha: Talpidae). Journal of Mammalogy 89 (6), 2008, S. 1428–1434
- G. A. Zuleta und C. Galindo-Leal: Distribution and abundance of four species of small mammals at risk in a fragmented landscape. Wildlife Working Report No. WR-64, Ministry of Environment, Lands and Parks, Victoria, B. C., 1994, S. 1–34
- Keith B. Aubry und Stephen D. West: The Status of Native and Introduced Mammals on Destruction Island, Washington. The Murrelet 65 (3), 1984, S. 80–83
- Carol J. Terry: Habitat Differentiation Among Three Species of Sorex and Neurotrichus gibbsi in Washington. The American Midland Naturalist 106 (1), 1981, S. 119–125
- Walter W. Dalquest and Donald R. Orcutt: The Biology of the Least Shrew-Mole, Neurotrichus gibbsii minor. The American Midland Naturalist 27 (2), 1942, S. 387–401
- Nobuo Ishii: Size and distribution of home ranges of the Japanese shrew-mole Urotrichus talpoides. Journal of the Mammalian Society of Japan 18 (2), 1993, S. 87–98
- John O. Whitaker, Jr., Chris Maser und Richard J. Pedersen: Food and Ectoparasitic Mites of Oregon Moles. Northwest Science 53 (4), 1979, S. 268–273
- Carol J. Terry: Food habits of three sympatric species of insectivora in western Washington. Canadian Field-Naturalist 92, 1978, S. 38–44 ()
- Kevin L. Campbell und Peter W. Hochachka: Thermal biology and metabolism of the American Shrew-mole, Neurotrichus gibbsii. Journal of Mammalogy 81 (2), 2000, S. 578–585
- Ellen B. Kritzman: A Captive Shrew-Mole and Her Litter. The Murrelet 53 (3), 1972, S. 47–49
- N. M. Rubenstein, G. R. Cunha, Y. Z. Wang, K. L. Campbell, A. J. Conley, K. C. Catania, S. E. Glickman und N. J. Place: Variation in ovarian morphology in four species of New World moles with a peniform clitoris. Reproduction 126, 2003, S. 713–719
- Jack C. von Bloeker: Mammal remains from Detritus of raptorial birds in California. Journal of Mammalogy 18 (3), 1937, S. 360–361
- Chris Maser und Edmund D. Brodie, Jr.: A study of owl pellet contents from Linn, Benton and Polk counties, Oregon. The Murrelet 47, 1966, S. 9–14
- Eric Forsman und Chris Maser: Saw-Whet Owl Preys on Red Tree Mice. The Murrelet 51 (1), 1970, S. 10
- Carl D. Marti: A review of prey selection by the Long-eared owl. The Condor 78, 1976, S. 331–336
- Catherine M. Raley und Keith B. Aubry: The food habits of Fishers (Pekania pennati) in the Cascade Range of Southern Oregon. Northwestern Naturalist 101, 2020, S. 143–157
- A. Fain und J. O. Whitaker, Jr.: Two new species of Myobiidae from North American mammals (Acarina). Bulletin et Annales de la Société entomologique de Belgique 111, 1975, S. 57–65
- John O. Whitaker, Jr. und Chris Maser: Mites (excluding chiggers) of mammals of Oregon. Great Basin Naturalist 45 (1), 1985, S. 67–76
- Terry L. Yates, Danny B. Penc und Glenn K. Launchbaugh: Ectoparasites from seven species of North American moles (Insectivora: Talpidae). Journal of Medical Entomology 16, 1979, S. 166–168
- R. L. Rausch und V. R. Rausch: Capillaria maseri sp. n. (Nematoda) from Insectivores (Soricidae and Talpidae) in Oregon. Proceedings of the Helminthological Society of Washington 40, 1973, S. 107–112
- John M. Kinsella und Vasyl V. Tkach: Checklist of helminth parasites of Soricomorpha (= Insectivora) of North America north of Mexico. Zootaxa 1969, 2009, S. 36–58
- Kai He, Akio Shinohara, Kristofer M. Helgen, Mark S. Springer, Xue-Long Jiang und Kevin L. Campbell: Talpid Mole Phylogeny Unites Shrew Moles and Illuminates Overlooked Cryptic Species Diversity. Molecular Biology and Evolution 34 (1), 2016, S. 78–87
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. A taxonomic and geographic Reference. 2 Bände. 3. Auflage. Johns Hopkins University Press, Baltimore MD 2005, ISBN 0-8018-8221-4 ()
- Gerhard Storch und Shuding Qiu: The Neogene mammalian faunas of Ertemte and Harr Obo in Inner Mongolia (Nei Mongol), China. 2. Moles – Insectivora: Talpidae. Senckenbergiana lethaea 64, 1983, S. 89–127
- Neal Woodman: American Recent Eulipotyphla. Nesophontids, Solenodons, Moles, and Shrews in the New World. Smithsonian Institution Scholary Press, 2018, S. 1–107 (S. 17–19)
- J. Howard Hutchison: Fossil Talpidae (Insectivora, Mammalia) from the Later Tertiary of Oregon. Bulletin of the Museum of Natural History University of Oregon 11, 1968, S. 1–117 (S. 55–58)
- Stanisław Skoczeń: Scaptonychini Van Valen, 1967, Urotrichini and Scalopini Dobson, 1883 (Insectivora, Mammalia) in the Pliocene and Pleistocene of Poland. Acta zoologica cracoviensia 24, 1980, S. 411–448
- Vasil V. Popov: Late Pliocene Erinaceidae and Talpidae (Mammalia: Insectivora) from Varshets (North Bulgaria). Acta zoologica cracoviensia 47 (1–2), 2004, S. 61–80
- Barbara Rzebik-Kowalska: Review of the Pliocene and Pleistocene Talpidae (Soricomorpha, Mammalia) of Poland. Palaeontologia Electronica 17 (2), 2014, S. 17.2.26A, doi:10.26879/457
- Stanisław Skoczeń: New records of Parascalops, Neurotrichus and Condylura (Talpinae, Insectívora) from the Pliocene of Poland. Acta Theriologica 38 (2), 1993, S. 125–137
- Jelle S. Zijlstra: Neurotrichus skoczeni, new name for Neurotrichus minor Skoczeń, 1993, preoccupied. Journal of Vertebrate Paleontology 30, 2010, S. 1903
- Gabrielle Sansalone, Tassos Kotsakis und Paolo Piras: New systematic insights about Plio-Pleistocene moles from Poland. Acta Palaeontologica Polonica 61 (1), 2016, S. 221–229, doi:10.4202/app.00116.2014
- Walter W. Dalquest und Robert L. Burgner: The Shrew-Mole of Western Washington. The Murrelet 22 (1), 1941, S. 12–14
- Spencer Fullerton Baird: General Report upon the Zoology of the Several Pacific Railroad Routes. Part 1: Mammals. Reports of Explorations and Surveys, to Ascertain the Most Practicable and Economical Route for a Railroad from the Mississippi River to the Pacific Ocean 8 (1), 1857, S. 1–757 (S. 76–77) ()
- Albert Günther: Notes on some Japanese mammals. Proceedings of the Zoological Society of London 1880, S. 440–443 ()
- T. S. Palmer: Index Generum Mammalium: A List of the Genera and Families of Mammals. North American Fauna 23, 1904, S. 459 ()
- William Alexander Forbes: Mammalia. Zoological Record 17, 1881, S. 1–31 (S. 14) ()
- Frederick W. True: A revision of the American moles. Proceedings of the United States National Museum 19 (1101), 1896, S. 1–112 (S. 98–106)
- Outram Bangs: A new race of Gibb's mole. The American Naturalist 31 (363), 1897, S. 240–242
- Clinton Hart Merriam: Results of a biological survey of Mount Shasta, California. North American Fauna 16, 1899, S. 1–169 (S. 88) ()
- George Gaylord Simpson: The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History 85, 1945, S. 1–350 (S. 53)
- Leigh van Valen: New Paleocene Insectivores and Insectivore classification. Bulletin of the American Museum of Natural History 135, 1967, S. 217–284
- George Edward Dobson: A Monograph of the Insectivora, systematic and anatomical. Part II. London, 1883, S. 126–172 ()
- Akio Shinohara, Kevin L. Campbell und Hitoshi Suzuki: Molecular phylogenetic relationships of moles, shrew moles, and desmans from the new and old worlds. Molecular Phylogenetics and Evolution 27, 2003, S. 247–258, doi:10.1016/S1055-7903(02)00416-5
- Akio Shinohara, Hitoshi Suzuki, Kimiyuki Tsuchiya, Ya-Ping Zhang, Jing Luo, Xue-Long Jiang, Ying-Xiang Wang und Kevin L. Campbell: Evolution and Biogeography of Talpid Moles from Continental East Asia and the Japanese Islands Inferredfrom Mitochondrial and Nuclear Gene Sequences. Zoological Science 21 (12), 2004, S. 1177–1185, doi:10.2108/zsj.21.1 177
- María Teresa Cabria, Jonathan Rubines, Benjamín Gómez-Moliner und Rafael Zardoya: On the phylogenetic position of a rare Iberian endemic mammal, the Pyrenean desman (Galemys pyrenaicus). Gene 375, 2006, S. 1–13, doi:10.1016/j.gene.2006.01.038
- A. A. Bannikova, E. D. Zemlemerova, V. S. Lebedev, D. Yu. Aleksandrov, Yun Fang und B. I. Sheftel: Phylogenetic Position of the Gansu Mole Scapanulus oweni Thomas, 1912 and the Relationships Between Strictly Fossorial Tribes of the Family Talpidae. Doklady Biological Sciences 464, 2015, S. 230–234
- Masaharu Motokawa: Phylogenetic relationships within the family Talpidae (Mammalia: Insectivora). Journal of Zoology 263, 2004, S. 147–157
- Marcelo R. Sánchez-Villagra, Inés Horovitz und Masaharu Motokawa: A comprehensive morphological analysis of talpid moles (Mammalia) phylogenetic relationships. Cladistics 22, 2006, S. 59–88
- Nick Crumpton und Richard S. Thompson: The holes of moles: Osteological correlates of the trigeminal nerve in Talpidae. Journal of Mammalian Evolution 20, 2013, S. 213–225
- Achim H. Schwermann und Richard S. Thompson: Extraordinarily preserved talpids (Mammalia, Lipotyphla) and the evolution of fossoriality. Journal of Vertebrate Paleontology 35 (4), 2015, S. e934828, doi:10.1080/02724634.2014.934828
- Wang Xiao-Ming, Qiu Zhu-Ding, Li Qiang, Yukimitsu Tomida, Yuri Kimura, Zhijie Jack Tseng und Wang Hong-Jiang: A New Early to Late Miocene Fossiliferous Region In Central Nei Mongol: Lithostratigraphy and Biostratigraphy In Aoerban Strata. Vertebrata Palasiatica 47 (2), 2009, S. 111–134
- Qiu Zhu-Ding, Wang Xiao-Ming, Li Qiang, Li Lu, Wang Hong-Jiang und Chen Hai-Feng: Late Miocene mammalian fauna of Halajin Hushu in Nei Mongol, China. Vertebrata Palasiatica 59 (1), 2021, S. 19–44
- Leonid L. Voyta, Thomas Mörs und Margarita A. Erbajeva: Erinaceomorpha and Soricomorpha (Mammalia) of the Miocene Tagay fauna (Olkhon Island, Lake Baikal, Eastern Siberia): A preliminary report. Palaeobiodiversity and Palaeoenvironments 102, 2022, S. 897–914, doi:10.1007/s12549-022-00557-9
- Reinhard Ziegler: Moles (Talpidae) from the late Middle Miocene of South Germany. Acta Palaeontologica Polonica 48 (4), 2003, S. 617–648
- Claude Gaillard: Mammifères miocènes nouveaux ou peu connus de La Grive-St.-Alban. Archives du Muséum d’Histoire Naturelle de Lyon 7, 1899, S. 1–79 (S. 29–31) ()
- Danielle Oberg: Fossil Moles from the Gray Fossil Site, TN: Implications for Diversification and Evolution of North American Talpidae. East Tennessee State University, 2018, S. 1–95
- Danielle E. Oberg und Joshua X. Samuels: Fossil moles from the Gray fossil site (Tennessee): Implications for diversification and evolution of North American Talpidae. Palaeontologia Electronica 25 (3), 2022, S. a33, doi:10.26879/1150
- F. Cassola: Neurotrichus gibbsii (errata version published in 2017). The IUCN Red List of Threatened Species 2016. e.T41468A115188045 (); zuletzt aufgerufen am 25. September 2022
Weblinks
- Neurotrichus gibbsii in der Roten Liste gefährdeter Arten der IUCN 2016. Eingestellt von: F. Cassola, 2016. Abgerufen am 25. September 2022.