Mesosaurus
Mesosaurus ist eine ausgestorbene Reptilien-Gattung aus der Zeit des frühen Perms (Artinskium, vor 290 bis 279 Millionen Jahren). Mesosaurus war aquatisch und konnte sich wahrscheinlich nicht besonders gut an Land fortbewegen. Er lebte an und in einem ausgedehnten See oder Binnenmeer, der oder das sich über das heutige südliche Afrika und östliche Südamerika erstreckte.
| Mesosaurus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelett in Kalkstein der Irati-Formation (Paraná-Becken, Brasilien) im Museo Geominero in Madrid | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Unterperm (Artinskium) | ||||||||||||
| 290 bis 279 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Mesosaurus | ||||||||||||
| Gervais, 1865[1] | ||||||||||||
| Art | ||||||||||||
|
Die Reptilien-Familie, in welche Mesosaurus gestellt wird, die Mesosauriden, sind nach heutigem Kenntnisstand die ersten amniotischen, d. h., von Gewässern prinzipiell unabhängig fortpflanzungsfähigen Landwirbeltiere, die zu einer nahezu rein aquatischen Lebensweise zurückkehrten – eine Entwicklungsrichtung, die im weiteren Verlauf der Erdgeschichte noch zahlreiche andere Linien der Amnioten einschlugen (u. a. Sauropterygier, Thalattosaurier, Pinguine, Robben). Zudem sind sie die einzigen sekundär aquatischen Amnioten des frühen Perms. Erst im späten Perm, etwa 20 Millionen Jahre nach den Mesosauriern, erscheinen mit einigen Vertretern der Younginiformes (z. B. Acerosodontosaurus und Hovasaurus), einer möglicherweise paraphyletischen Gruppe früher Diapsiden, die geologisch nächstjüngeren Formen im Fossilbericht.
Etymologie
Der Name „Mesosaurus“ ist zusammengesetzt aus den altgriechischen Wörtern μέσος mésos ‚mittel‘ und σαῦρος sauros ‚Eidechse‘, bedeutet folglich soviel wie „mittlere Echse“. Der Name wurde 1865 vom französischen Naturforscher Paul Gervais geprägt,[1] der damit zum Ausdruck bringen wollte, dass er einzelne Merkmale des Skelettes von Mesosaurus bei zahlreichen sowohl seinerzeit bereits bekannten fossilen als auch rezenten Reptilien wiederfand, und die Gattung daher eine Zwischenstellung einnehme.
Merkmale

Mesosaurus war ein verhältnismäßig kleines, schlankes Reptil, das eine Länge von bis zu einem Meter erreichte. Die meisten gefundenen Exemplare weisen jedoch nur eine Länge von etwa 40 Zentimetern auf. Die Kiefer sind sehr schmal und stark verlängert, etwa so wie bei heutigen Gavialen. Sie sind mit einer Vielzahl nadelförmiger und z. T. sehr langer Zähne bestückt, deren Spitzen zu Lebzeiten relativ stark nach außen gerichtet waren, sodass die Zähne schräg im Kiefer standen. Der Hals besitzt 12 Wirbel und ist verlängert, was vermutlich die Bewegungsfreiheit des Kopfes erhöhte.
Als Anpassung an die aquatische Lebensweise besaßen sie einen abgeplatteten Ruderschwanz für den Vortrieb im Wasser, ähnlich wie ihn die heutigen Krokodile besitzen. Da die Gliedmaßen nicht, wie bei Landbewohnern, vorwiegend dazu dienten, den Rumpf an Land zu tragen, sind die Knochen in den Hand- und Fußwurzeln der Mesosauriden leicht reduziert. Zudem sind Hände und Füße paddelartig ausgebildet und die Füße sind relativ groß und dienten vermutlich ebenfalls dem Vortrieb. Wahrscheinlich spannten sich zwischen den Fingern und Zehen Schwimmhäute, möglicherweise bildeten die Weichteile von Hand und Fuß sogar regelrechte „Flossen“, ähnlich wie bei den heutigen Ohrenrobben.
Die Rippen sind im Vergleich zu ähnlich großen landbewohnenden Reptilien verdickt – eine Abwandlung, die als Pachyostose bezeichnet wird. Pachyostose tritt als charakteristisches Element auch bei vielen anderen sekundär aquatischen Wirbeltieren auf (z. B. besonders deutlich bei den heutigen Seekühen). Die dickeren und somit auch schwereren Knochen sorgen für einen verringerten Auftrieb im Wasser.
Schläfenfenster
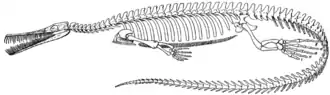
Viele ausgestorbene und lebende Reptiliengruppen zeichnen sich durch charakteristische Öffnungen in den hinteren oberen und seitlichen Partien des Schädeldaches aus, sogenannte Temporal- oder Schläfenfenster. Auch bei Mesosaurus wurden solche Schläfenfenster bereits in der ersten Hälfte des 20. Jahrhunderts beschrieben.[3] Allerdings ist im Rahmen späterer Untersuchungen festgestellt worden, dass die Schädelknochen von Mesosaurus in diesem Bereich sehr dünnwandig und fragil sind, und dass frühere Beobachtungen von Temporalfenstern darauf beruhten, dass diese dünnwandigen Schädelpartien während der Fossilisierung beschädigt worden seien, Mesosaurus also keine Temporalfenster besäße.[4] Vor kurzem wurden jedoch erneut einzelne Mesosaurus-Exemplare beschrieben, die ein Temporalfenster besitzen sollen.[3]
Lebensweise
Ernährung
Mesosaurus war ein Jäger, der seiner Beute aktiv im freien Wasser nachstellte, ähnlich dem Jagdverhalten heutiger Robben oder Zahnwale. Die nadelförmigen Zähne dienten aber wahrscheinlich nicht dem Aufspießen von Beutetieren, sondern als Reusenapparat zum Einfangen einzelner kleinerer frei schwimmender Krebstiere und Fische, die im Maul festgehalten wurden, wenn das Tier die Kiefer schloss und das Wasser zwischen den ineinandergreifenden Zähnen hindurch aus dem Maul presste. Die Hauptnahrung von Mesosaurus bestand vermutlich aus Krill-ähnlichen Kleinkrebsen der ausgestorbenen Familie Notocarididae.[4]

Fortpflanzung
Ein sehr kleines Mesosaurus-Individuum mit weitgehend verknöchertem Skelett, das innerhalb des Rippenkorbs eines ausgewachsenen Tieres entdeckt wurde, weist keine Anzeichen von magensäurebedingten Lösungserscheinungen auf. Daher handelt es sich wahrscheinlich um einen Embryo bzw. Fötus im Mutterleib. Dies ließe den Schluss zu, dass Mesosaurus, wie einige Meeresreptilien des Mesozoikums, lebendgebärend (vivipar) war. Nicht weniger wahrscheinlich ist aber, dass er Eier am Strand ablegte, ganz ähnlich wie es die heutigen Meeresschildkröten tun. Im Gegensatz zu den Schildkröten war der Mesosaurus-Embryo jedoch zum Zeitpunkt der Eiablage bereits relativ weit entwickelt, wurde also bereits im Muttertier weitgehend ausgebrütet (sogenannte Ovoviviparie). Da diese Eier im Vergleich zum ausgewachsenen Tier recht groß waren, konnte ein Weibchen nur ein bis zwei Eier in sich tragen, weswegen aus einem Fortpflanzungszyklus nur wenige Nachkommen resultierten. Daher ist es nicht auszuschließen, dass Mesosaurus so etwas wie Brutpflege betrieb, um die Überlebenschancen dieser wenigen Nachkommen zu verbessern.[5]
Systematik

Aufgrund verschiedener Interpretationen des Schädelbaus hinsichtlich des Vorhandenseins von Schläfenfenstern wurde Mesosaurus im 20. Jahrhundert zunächst als Diapside und damit als Verwandter der meisten heute lebenden Reptilien und Dinosaurier, dann als Synapside und damit als ein entfernter Verwandter der Säugetiere und in jüngerer Vergangenheit als „Anapside“ (siehe klassisches Konzept der Anapsida nach Caroll, 1988) betrachtet.[4] Letztgenannte Gruppe und das zugrunde liegende Gliederungskonzept bilden jedoch den tatsächlichen Verlauf der Evolution und damit die tatsächlichen Verwandtschaftsverhältnisse der Amnioten nicht korrekt ab bzw. hat die Bedeutung des nicht-vorhandenseins von Schläfenöffnungen für die Systematik der Amnioten abgenommen.[6] Neuere, kladistische Untersuchungen ergaben, dass Mesosaurus offenbar mit einer Reihe anderer, überwiegend permischer und heute gänzlich ausgestorbener Reptilien verwandt ist, die, einschließlich der Mesosauriden, als Parareptilien bezeichnet werden.[4][6] Die Gemeinsamkeiten im Körperbau, die Mesosaurus aufgrund seiner aquatischen Lebensweise mit sowohl ausgestorbenen als auch heute lebenden aquatischen Reptilien, wie zum Beispiel den Thalattosauriern oder Krokodilen, aufweist, sind das Ergebnis einer konvergenten Evolution.
Die engsten Verwandten von Mesosaurus sind die anderen Gattungen in der Familie Mesosauridae: Brazilosaurus und Stereosternum. Von Mesosaurus selbst wurden zwei Arten beschrieben: M. tenuidens und M. brasiliensis. Eine dritte Art, M. capensis, wurde ursprünglich als Vertreter einer eigenen Gattung (Ditrochosaurus) beschrieben. Heute werden M. (= „Ditrochosaurus“) capensis, und M. brasiliensis als jüngere Synonyme von M. tenuidens betrachtet. Teilweise wird auch Stereosternum tumidum, die einzige Art dieser Gattung, als Mesosaurus-Art aufgefasst.[7]
Der Holotyp der Art Mesosaurus tenuidens, die wiederum Typus-Art der Gattung Mesosaurus ist, wurde in den 1840er Jahren im „Griqualand“ (sehr wahrscheinlich handelte es sich dabei um das West-Griqualand – heute Teil der Nordkap-Provinz Südafrikas) vom Bruder des französischen Naturforschers Édouard Verreaux in einer Griqua-Hütte entdeckt. Ihm zufolge habe die Gesteinsplatte mit dem Fossil den Bewohnern als Deckel für ihren Kochtopf gedient.[1]
Vorkommen und Fossilerhaltung


Mesosaurus-Fossilien kommen im südlichen Afrika überwiegend in der unterpermischen Whitehill-Formation des Karoo-Systems und in Brasilien und Uruguay überwiegend in der ebenfalls unterpermischen Irati- bzw. Mangrullo-Formation des Paraná-Beckens vor. Sie treten in diesen Schichten z. T. massenhaft, meist in Form ganzer oder partieller, flachgedrückter Skelette auf.[9]
Whitehill-, Irati- und Mangrullo-Formation bestehen aus dunklen Tonsteinen. Die Skelette von Mesosaurus sind in diesen Tonsteinen zu einem sehr weichen Material umgewandelt worden und sind oft bereits weitgehend herausgewittert, sodass nur Abdrücke im Gestein übrig bleiben. Einige Abschnitte der Irati-Formation (Assistência-Subformation) bestehen aus gelblich-weißem Kalkstein. In diesen Kalksteinen ist die Fossilüberlieferung der Knochensubstanz deutlich besser. Allerdings gelten nicht Mesosaurus, sondern die beiden anderen Mesosaurier-Gattungen Stereosternum und Brazilosaurus als typische Vertreter der Kalksteinfazies.[8][9]
Bei den Ton- und Kalksteinen der Whitehill-, Irati- und Mangrullo-Formation handelt es sich um die Ablagerungen eines ausgedehnten Sees oder Binnenmeeres („Mesosaurus-Meer“), wobei die Kalksteine küstennahe, flache Gewässer und die Tonsteine küstenfernere, tiefere Gewässer repräsentieren.[8][9]
Mesosaurus und die Plattentektonik

Die Verbreitung der Gattung Mesosaurus in den beiderseits des Süd-Atlantiks befindlichen Regionen des östlichen Südamerikas und südlichen Afrikas, die heute durch tausende Kilometer offenen Ozeans voneinander getrennt sind, war bereits dem deutschen Naturwissenschaftler Alfred Wegener bekannt. Sie diente ihm als wichtiges Argument zur Untermauerung der von ihm 1915 aufgestellten Theorie der Kontinentalverschiebung. Die Verbreitung der Mesosaurus-Fossilien beiderseits des Atlantiks und die Ähnlichkeit der Gesteine die sie enthalten, sind ein deutlicher Hinweis darauf, dass zu Lebzeiten von Mesosaurus Afrika und Südamerika im Urkontinent Gondwana vereint waren. Gondwana brach erst nach dem Aussterben der Mesosaurier infolge der durch die Mechanismen der Plattentektonik bewirkten Bewegungen der Lithosphärenplatten auseinander und Afrika und Südamerika driften seither voneinander weg.
Literatur
- Martin Sander: Reptilien. 220 Einzeldarstellungen (= Haeckel-Bücherei. Bd. 3). Ferdinand Enke Verlag, Stuttgart 1994, ISBN 3-432-26021-0.
- Michael J. Benton: Vertebrate Palaeontology. 3. Auflage. Wiley-Blackwell Publishing, Malden MA 2005, ISBN 0-632-05637-1.
Einzelnachweise
- Paul Gervais: Description du Mesosaurus tenudiens. Reptile fossile de l'Afrique australe. Académie des Sciences et Lettres de Montpellier. Mémoires de la Section des Sciences. 6. Band, Teil II, 1865, S. 169–175 (MDZ-Reader).
- Samuel Wendell Williston: Water Reptiles of the Past and Present. The University of Chicago Press, Chicago IL 1914, S. 128 (archive.org).
- Graciela Piñeiro, Jorge Ferigolo, Alejandro Ramos, Michel Laurin: Cranial morphology of the Early Permian mesosaurid Mesosaurus tenuidens and the evolution of the lower temporal fenestration reassessed. Comptes Rendus Palevol. Bd. 11, Nr. 5, 2012, S. 379–391, doi:10.1016/j.crpv.2012.02.001.
- Sean Patrick Modesto: The cranial skeleton of the Early Permian aquatic reptile Mesosaurus tenuidens: implications for relationships and palaeobiology. Zoological Journal of the Linnean Society. Bd. 146, Nr. 3, 2006, S. 345–368, doi:10.1111/j.1096-3642.2006.00205.x.
- Die Informationen des kompletten Absatzes stammen aus: Graciela Piñeiro, Jorge Ferigolo, Melitta Meneghel, Michel Laurin: The oldest known amniotic embryos suggest viviparity in mesosaurs. Historical Biology. Bd. 24, Nr. 6, 2012, S. 620–630, doi:10.1080/08912963.2012.662230.
- Linda A. Tsuji, Johannes Müller: Assembling the history of the Parareptilia: phylogeny, diversification, and a new definition of the clade. Fossil Record. Bd. 12, Nr. 1, 2009, S. 71–81, doi:10.1002/mmng.200800011.
- Hans-Volker Karl, Elke Gröning, Carsten Brauckmann: The Mesosauria in the collections of Göttingen and Clausthal: implications for a modified classification. Clausthaler Geowissenschaften. Bd. 6, 2007, ISSN 1611-0609, S. 63–78.
- Burger W. Oelofsen, Dina C. Araújo: Mesosaurus tenuidens and Stereosternum tumidum from the Permian Gondwana of both Southern Africa and South America. South African Journal of Science. Bd. 83, Nr. 6, 1987, ISSN 0038-2353, S. 370–372.
- Sean Patrick Modesto: The Postcranial Skeleton of the Aquatic Parareptile Mesosaurus tenuidens from the Gondwanan Permian. Journal of Vertebrate Paleontology. Bd. 30, Nr. 5, 2010, S. 1378–1395, doi:10.1080/02724634.2010.501443.
Weblinks
- Kladogramm der Mesosauriden und weiterer Parareptilien (Mikko's Phylogeny Archive)