Mendelsche Regeln
Die Mendelschen Regeln beschreiben den Vererbungsvorgang bei Merkmalen, deren Ausprägung von jeweils nur einem Gen bestimmt wird (monogener Erbgang). Sie sind nach ihrem Entdecker Gregor Mendel benannt, der sie 1865/1866 publizierte, die aber zunächst kaum zur Kenntnis genommen und erst 1900, lange nach seinem Tod, „wiederentdeckt“ wurden.

Die Mendelschen Regeln gelten nur für Lebewesen, bei denen sich im Zuge der sexuellen Fortpflanzung diploide und haploide Stadien (mit zwei bzw. einem Chromosomensatz im Zellkern) abwechseln (siehe Kernphasenwechsel). Für Organismen mit höherem Ploidiegrad (Polyploidie) lassen sich entsprechende Regeln ableiten. Viele Merkmale werden allerdings polygen vererbt durch das Zusammenwirken mehrerer Gene.
Die alternativen Bezeichnungen Vererbungsgesetze[1] und Mendelsche Gesetze sind ungebräuchlich geworden, da schon bald genetische Phänomene entdeckt wurden, bei denen ein Erbgang von den „Regeln“ abweichen kann. Beispiele für eine nicht-mendelsche Vererbung sind die Genkopplung, die extrachromosomale Vererbung, die nicht-zufällige Segregation von Chromosomen und der Meiotic Drive.
Forschungsgeschichte
Die Mendelschen Regeln wurden Anfang der 1860er-Jahre von dem Augustinermönch und Hilfslehrer Gregor Mendel erkannt und durch Kreuzungsversuche an Erbsenpflanzen bestätigt. 1865 berichtete er darüber in zwei Vorträgen im Naturforschenden Verein in Brünn ([Ü]ber Hybriden im Pflanzenreiche am 8. Februar bzw. im März 1865).[2][3][4] 1866 folgte eine lange Zeit wenig beachtete gedruckte Publikation.[5]
Mendel entdeckte Gesetzmäßigkeiten, die Anderen zuvor und noch Jahrzehnte danach verborgen blieben. Der Erfolg seiner Untersuchungen an Erbsenpflanzen lässt sich im Nachhinein mit folgenden Faktoren begründen:[6]
- die Beschränkung auf wenige, klar unterscheidbare Merkmale,
- die Auswahl reinerbiger Stämme,
- die künstliche Bestäubung unter Schutz vor natürlicher Fremdbestäubung und Selbstbestäubung[7]
- die großangelegten Versuchsreihen sowie
- die statistische Auswertung.
Insbesondere die Beschränkung auf einzelne Merkmale stand im Gegensatz zu damals herrschenden Vorstellungen, wonach der ganze elterliche Organismus auf die Eigenschaften der Nachkommen Einfluss nimmt und dies durch eine Flüssigkeit (beim Menschen das Blut) vermittelt wird. Entsprechend wurden die Nachkommen als Mischlinge oder Bastarde betrachtet und als solche kategorisiert.[8]
Mendel wählte sieben Merkmale aus und kreuzte jeweils reinerbige Pflanzen, die sich in einem dieser Merkmale, z. B. der Blütenfarbe, unterschieden. Er stellte fest, dass alle Nachkommen der ersten Generation nur eine Ausprägung des jeweiligen Merkmals aufwiesen, z. B. violette Blüten. Kreuzte er nun Pflanzen dieser Generation erneut miteinander, so trat in der zweiten Tochtergeneration das verschwundene Merkmal (weiße Blüten) wieder auf, und zwar bei einem Viertel der Nachkommen. Dieses 3:1–Verhältnis fand Mendel in allen sieben untersuchten Fällen, und er konnte es durch eine große Zahl von Versuchen statistisch absichern. Zur Erklärung führte er die Begriffe Dominanz und Rezessivität ein: Die dominante violette Farbe verdeckte in der ersten Generation die rezessive weiße, diese trat aber in der zweiten Generation wieder auf. Aus diesen Beobachtungen schloss Mendel, dass jede Pflanze den „Erbfaktor“ für die Blütenfarbe oder ein anderes Merkmal in zweifacher Ausfertigung besitzt. Eine Bestätigung dieser Annahme lieferten Rückkreuzungsversuche mit weißblütigen Pflanzen, bei denen entweder nur violettblühende Tochterpflanzen auftraten oder beide Blütenfarben zu gleichen Teilen, womit sich die violettblühende Mutterpflanze als reinerbig bzw. mischerbig erwies.[9]
Der niederländische Botaniker Hugo de Vries kam in den 1880er Jahren bei Kreuzungsexperimenten zu der Überzeugung, dass der bisherige ganzheitliche Erklärungsansatz falsch sei und man stattdessen die Vererbung einzelner Merkmale untersuchen müsse. In zahlreichen Versuchsreihen mit vielen verschiedenen Pflanzenarten, die er quantitativ auswertete, fand er durchweg wie Mendel eine Aufspaltung im Verhältnis 3:1. In einer Publikation mit dem Titel Das Spaltungsgesetz der Bastarde forderte er 1900 „eine vollständige Umwandlung der Ansichten“ und griff dabei Mendels Arbeit über Erbsen auf, deren Ergebnisse er als allgemein gültig bezeichnete. Parallel begann der deutsche Botaniker Carl Correns 1894 mit entsprechenden Versuchen, deren Ergebnisse er ebenfalls 1900 unter Bezug auf Mendel veröffentlichte. Als dritter „Wiederentdecker“ Mendels im selben Jahr galt lange Zeit der österreichische Pflanzenzüchter Erich Tschermak; bei ihm bestehen jedoch Zweifel, ob er Mendels Ergebnisse zu diesem Zeitpunkt korrekt verstanden hatte.[10][11][12] Im Jahr 1902 hatte Archibald E. Garrod an einer Familie mit Alkaptonurie die Gültigkeit der Mendelschen Regeln nachgewiesen.[13] Zwischenzeitlich waren die Chromosomen und ihre Verteilung an die Nachkommen beschrieben worden, so dass die mendelschen Regeln jetzt mit diesen Beobachtungen zur Chromosomentheorie der Vererbung vereinigt werden konnten.
Die drei Regeln
Gültigkeit
Die Mendelschen Regeln beziehen sich auf Merkmale, die von einem einzigen Gen festgelegt werden. Jedes Gen liegt in der diploiden Kernphase in zwei Kopien („Allelen“) vor, von denen je eines von jedem Elternteil stammt. Die diploide Phase umfasst bei den Metazoa (Vielzellige Tiere einschließlich des Menschen) alle Zellen mit Ausnahme der Gameten (Eizelle und Spermium). Pflanzen weisen dagegen einen Wechsel von diploiden und haploiden Generationen auf, wobei bei „höheren“ Pflanzen die diploide Generation in den Vordergrund tritt und die haploide Phase bei den Samenpflanzen stark reduziert ist (Embryosack und Pollenkorn). Mendel und seine „Wiederentdecker“ befassten sich mit den Merkmalen diploider Pflanzen.
Regel 1: Uniformität

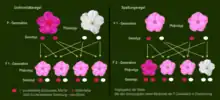
Werden zwei Individuen miteinander gekreuzt, die sich in einem Merkmal unterscheiden, für das sie beide jeweils homozygot (reinerbig) sind, dann sind die Nachkommen der ersten Generation (Tochtergeneration F1) uniform, d. h. gleich bezogen auf das untersuchte Merkmal. Diese Uniformitätsregel oder Reziprozitätsregel gilt sowohl für den Phänotyp als auch für den Genotyp, welcher bei allen Individuen der F1-Generation heterozygot (mischerbig) ist.
Für die Ausprägung des Merkmals tritt je nach dessen Erbgang eine von drei Möglichkeiten ein:
- Beim dominant-rezessiven Erbgang haben alle Mitglieder der F1-Generation denselben Phänotyp wie ein Elternteil. Beispiel: Bei Erbsen ist die rote Blütenfarbe dominant gegenüber der weißen, die Anlage für weiße Blüten wird daher als rezessiv bezeichnet. Wenn reinerbige rotblühende und reinerbige weißblühende Individuen gekreuzt werden, haben alle Mitglieder der F1-Generation ein Allel für weiß und ein Allel für rot vererbt bekommen, sie sind heterozygot. Trotzdem sind sie alle rotblühend, weil rot gegenüber weiß dominant ist.
- Beim intermediären Erbgang haben alle Mitglieder der F1-Generation eine Mischform der elterlichen Merkmale, man spricht auch von unvollständiger Dominanz. Ein Beispiel ist die Blütenfarbe der Wunderblume Mirabilis jalapa: Wenn rot- und weißblütige Exemplare gekreuzt werden, so haben alle Nachkommen rosafarbene Blüten. Die vollständige Dominanz ist allerdings ein idealer Grenzfall; es gibt alle Abstufungen bis hin zum perfekten intermediären Erbgang.[14][15]
- Beim kodominanten Erbgang bilden alle Mitglieder der F1-Generation beide Merkmale der Eltern separat aus. Ein Beispiel dafür sind die Allele A und B im AB0-System der menschlichen Blutgruppen.
Ausnahmen von der 1. Regel können auftreten, wenn sich das Gen für ein untersuchtes Merkmal auf einem Geschlechtschromosom (Gonosom) befindet. Dann kann es sein, dass die F1-Generation nicht uniform ist.
Regel 2: Spaltung
Die Spaltungsregel oder Segregationsregel gilt, wenn zwei Individuen gekreuzt werden, die beide gleichartig heterozygot sind, also z. B. zwei Pflanzen, die für die Blütenfarbe "Weiß" und "Rot" Erbanlagen haben. Das kann etwa die F1-Generation des vorherigen Abschnitts sein. In Beschreibungen der mendelschen Regeln werden die Nachkommen einer solchen Heterozygoten-Kreuzung daher als Enkel- oder zweite Filialgeneration (F2) bezeichnet. Die Nachkommen aus dieser Paarung sind untereinander nicht mehr uniform, sondern spalten sich sowohl im Genotyp als auch im Phänotyp auf.
- Handelt es sich um eine dominant-rezessive Vererbung, so sind durchschnittlich ein Viertel der F2-Individuen reinerbig mit zwei rezessiven Allelen und zeigen eine entsprechende Merkmalsausprägung (z. B. weiße Erbsenblüten). Die anderen drei Viertel zeigen im Phänotyp die dominante Erbanlage. Diese drei Viertel setzen sich zusammen aus einem Viertel reinerbige und zwei Vierteln mischerbige Individuen. Insgesamt besteht also im Phänotyp ein Verhältnis von 3:1, im Genotyp ein Verhältnis von 1:2:1. Bei dominant-rezessiver Vererbung sind also in der zweiten Nachkommengeneration, der F2-Generation, 25 Prozent der Individuen homozygote Träger des dominanten Merkmals und 50 Prozent heterozygote Träger des dominanten Merkmals. Das fand Gregor Mendel durch Rückkreuzungen heraus. Die mischerbigen Individuen, die das dominante Merkmal ausprägen, aber auch die rezessive Erbanlage vererben können, nennt man Konduktoren.
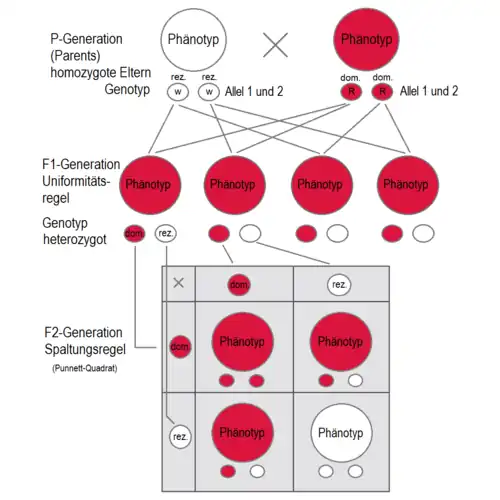
Elterngeneration P mit jeweils reinerbigen Anlagen für weiß (w/w) bzw. rot (R/R).
F1-Generation: Alle Individuen sehen gleich aus. Auch bei den mischerbigen Exemplaren reicht die dominante Erbanlage für die Bildung des roten Blütenpigments auf nur einem Allel aus, um eine volle Ausprägung des Merkmals rotblütig zu erreichen, obwohl das andere Allel die Erbanlage weiß, also das Fehlen des Blütenpigments beinhaltet.
F2-Generation: Dominante (rot) und rezessive (weiße) Erscheinungsformen zeigen ein Verhältnis von 3:1.
Der erste Vererbungsschritt von der P-Generation zur F1-Generation wird hier in einem anderen Erbschema dargestellt als die darauf folgende Vererbung von der F1- zur F2-Generation. Das untere ist ein Punnett-Quadrat
- Bei intermediärer Vererbung weisen jeweils durchschnittlich 25 % der Nachkommen das Merkmal eines der beiden reinerbigen Individuen auf, etwa 50 % der Nachkommen bilden eine Mischform der beiden Merkmale aus (unvollständige Dominanz). Das Mengenverhältnis ist beim Phänotyp und beim Genotyp jeweils 1:2:1. Der Genotyp ist in diesem Falle am Phänotyp erkennbar.

Elterngeneration P mit jeweils reinerbigen Anlagen für weiß (w/w) bzw. pinkrot (r/r).
F1-Generation: Alle Individuen sehen gleich aus. Die „roten“ und „weißen“ Erbanlagen der mischerbigen Pflanzen ergeben eine rosa Blütenfarbe.
F2-Generation. Rote, rosa und weiße Blütenfarben treten mit einem 1:2:1 Verhältnis auf.
- Bei kodominanter Vererbung gilt die Aufspaltung im Verhältnis 1:2:1 analog.
Zu Abweichungen von der Spaltungsregel siehe jedoch: Nicht-zufällige Segregation von Chromosomen.
Regel 3: Unabhängigkeit

Die Unabhängigkeitsregel oder Neukombinationsregel beschreibt die Vererbung von zwei Merkmalen (dihybrider Erbgang), die gleichzeitig vorhanden sind, bei der Kreuzung reinerbiger Individuen und deren Nachkommen. Beide Merkmale werden unabhängig voneinander vererbt (daher der Name der Regel), sodass ab der F2-Generation neue, reinerbige Kombinationen auftreten. Erst Zytogenetiker zeigten, dass die Gene unabhängiger Merkmale auf verschiedenen Chromosomen liegen.
Beispiel im nebenstehenden Erbschema: P-Generation: Jedes Elterntier besitzt je ein dominantes und ein rezessives Merkmal jeweils reinerbig (homozygot). F1: Nach der Uniformitätsregel sind alle Individuen in Genotyp und Phänotyp gleich und mischerbig (heterozygot). Sie prägen die dominanten Merkmale im Phänotyp aus. F2: Nun sind alle Allelkombinationen möglich. Die Merkmale werden unabhängig voneinander vererbt, so dass neue Kombinationen auftreten können. Durchschnittliches Zahlenverhältnis der Phänotypen 9:3:3:1[16] Bei diesem Beispiel in der F2-Generation: 9 voll pigmentierte mit kurzem, 3 voll pigmentierte Katzen mit langem Fell und 3 weiß gescheckte mit kurzem Fell, sowie eine weiß gescheckte Katze mit langem Fell entstanden durch Neukombination der beiden rezessiven Merkmale. Die Spaltungsregel und die Unabhängigkeitsregel stehen in Einklang mit der Chromosomentheorie der Vererbung.

Die Unabhängigkeitsregel gilt allerdings nur dann, wenn sich die für die Merkmale verantwortlichen Gene auf verschiedenen Chromosomen befinden, die durch die Meiose voneinander getrennt werden, oder wenn sie auf dem gleichen Chromosom so weit voneinander entfernt liegen, dass sie während der Meiose durch Crossing-over regelmäßig getrennt voneinander vererbt werden. Befinden sich Gene auf dem gleichen Chromosom nahe beieinander, so werden sie in Kopplungsgruppen vererbt.
Bei den beiden hier beispielhaft dargestellten Merkmalen handelt es sich um dominant-rezessive Erbgänge. Werden jedoch ein oder beide Merkmale intermediär vererbt, so werden die Phänotypen nicht im Zahlenverhältnis 9:3:3:1 ausgebildet. Im Genotyp bleiben die Zahlenverhältnisse bei allen Erbformen gleich. Alle Verhältnisse lassen sich durch Ausrechnen der obigen Matrix (Punnett-Quadrat) leicht ermitteln.

Literatur
- Uwe Hoßfeld, Michael V. Simunek: 150 Jahre Mendels Vortrag „Versuche über Pflanzen-Hybriden“. In: BIOspektrum. Band 21, Nr. 2, 2015, S. 238 f. (Biospektrum: Online-Version).
- Floyd V. Monaghan und Alain F. Corcos: Reexamination of the fate of Mendel's paper. In: The Journal of Heredity. Band 78, Nr. 2, 1987, S. 116–118.
- Jane B. Reece & al.: Campbell Biologie. 10. Aufl., Pearson Deutschland, Hallbergmoos 2016. Kap. 14: Mendel und das Genkonzept, S. 345–373.
- Ulrike Roll: Mendelsche Gesetze. In: Werner E. Gerabek, Bernhard D. Haage, Gundolf Keil, Wolfgang Wegner (Hrsg.): Enzyklopädie Medizingeschichte. De Gruyter, Berlin/New York 2005, ISBN 3-11-015714-4, S. 968 f.
Weblinks
- Versuche über Pflanzen-Hybriden (1865): Mendels Original-Veröffentlichung (vorgelegt in den Sitzungen vom 8. Februar und 8. März 1865) bei mendelweb.org, Roger B. Blumberg
Einzelnachweise
- Vgl. etwa Paul Diepgen, Heinz Goerke: Aschoff/Diepgen/Goerke: Kurze Übersichtstabelle zur Geschichte der Medizin. 7., neubearbeitete Auflage. Springer, Berlin/Göttingen/Heidelberg 1960, S. 41.
- Vom naturforschenden Vereine. In: Mährischer Correspondent, 7. Februar 1865, S. 3 (online bei ANNO).
- Monatsversammlung des naturforschenden Vereines. In: Mährischer Correspondent, 10. März 1865, S. 4 (online bei ANNO).
- Uwe Hoßfeld, Michael V. Simunek: 150 Jahre Mendels Vortrag „Versuche über Pflanzen-Hybriden“. 2015, S. 238.
- Gregor Mendel: Versuche über Pflanzenhybriden. Verhandlungen des Naturforschenden Vereines in Brünn. Bd. IV. 1866. S. 3–47.
- Abiturwissen Biologie. Bibliographische Informationen der Deutschen Bibliothek, 2004, ISBN 3-411-00222-0.
- Neil A. Campbell, Jane B. Reece: Biologie. Spektrum-Verlag 2003, Seite 294, ISBN 3-8274-1352-4.
- Ilse Jahn, Rolf Löther, Konrad Senglaub (Hrsg.): Geschichte der Biologie. 2., durchgesehene Auflage. VEB Fischer, Jena 1985, S. 554–555.
- Jane B. Reece & al.: Campbell Biologie, 10. Aufl., Pearson Deutschland, Hallbergmoos 2016, S. 347–352.
- Ilse Jahn (Hrsg.): Geschichte der Biologie. 3. Auflage. Sonderausgabe Nikol, Hamburg 2004, S. 415–418.
- Ernst Mayr: The Growth of Biological Thought, Belknap Press, S. 730 (1982)
- Floyd Monaghan, Alain Corcos: Tschermak: A non-discoverer of Mendelism, Journal of Heredity 77: 468f (1986) und 78: 208-210 (1987) (Abstract)
- Paul Diepgen, Heinz Goerke: Aschoff/Diepgen/Goerke: Kurze Übersichtstabelle zur Geschichte der Medizin. 7., neubearbeitete Auflage. Springer, Berlin/Göttingen/Heidelberg 1960, S. 56.
- Jane B. Reece & al.: Campbell Biologie. 10. Aufl., Pearson Deutschland, Hallbergmoos 2016. S. 358.
- C. Bresch, R. Hausmann: Klassische und molekulare Genetik. 3. Aufl., Springer, Berlin / Heidelberg / New York 1972. S. 41.
- Spektrum Lexikon Biologie Mendel-Regeln