MDCK-Zellen
MDCK-Zellen (von englisch Madin-Darby Canine Kidney) sind eine Zelllinie, die aus der Niere eines Hundes abstammt.

Eigenschaften
Es existieren verschiedene Stämme von MDCK-Zellen: die parentale Linie, MDCK I, MDCK II, MDCK.1, MDCK.2, Superdome und Supertube.[1] Sie sind aufgrund einer verzweigenden Morphogenese eine der wenigen Zellkultur-Modelle für die Entstehung von Gewebe und geeignet für die 3D-Zellkultur.[2] MDCK-Zellen werden für verschiedene Versuche von Epithelzellen in Zellkultur verwendet, wie Proteintransport, Zellpolarität und Zellkontakte (Tight Junctions, Adherens Junctions, Desmosomen und Gap Junctions).[1] Weiterhin werden sie für Untersuchungen des Transports von Arzneistoffen in und durch Zellen verwendet.[3] Sie sind in der Virologie typische Zellen, die zum Plaque-Assay und zum Neutralisationstest verwendet werden.[4] Ebenso können sie zur Virusanzucht bei der Produktion von Influenzaimpfstoffen eingesetzt werden.[5] Der Influenzaimpfstoff Optaflu wird in MDCK-Zellen hergestellt.[6] Die Infizierbarkeit (Permissivität) durch Viren umfasst bei MDCK-Zellen Influenzaviren, Humanes Coxsackie-Virus B3, Humanes Coxsackie-Virus B4, Humanes Coxsackie-Virus B 5, Reovirus Typ 2, Adeno-assoziiertes Virus 4, Adeno-assoziiertes Virus 5, Vacciniavirus, Vesicular stomatitis Indiana virus und Humanes Poliovirus 2.[7]
Geschichte

Nachdem Stewart H. Madin und Norman B. Darby, Jr. 1958 erstmals Epithelzellen aus dem Nierentubulus eines erwachsenen weiblichen Cocker Spaniels isoliert hatten,[7] wurde die Zelllinie als Modell für die Virusinfektion von Säugetierzellen verwendet.[8][9][10] Tatsächlich wählten sie die Isolierung von Nierentubuli mit genau diesem Ziel, da sie bereits zuvor mit der Virusinfektion von Zellen aus Nierentubuli anderer Säugetiere erfolgreich gewesen waren, und nicht ein neues Modellsystem für die Epithelzellbiologie zu schaffen. Erst 1970 veröffentlichte das Labor von Zbynek Brada eine Arbeit, in der MDCK-Zellen als repräsentative Zelllinie beschrieben wurden, die Merkmale von Nierentubulusepithelzellen aufwiesen.[11] Sie begründeten diese Schlussfolgerung mit der Flüssigkeitstransportaktivität der aus MDCK-Zellen gebildeten Monoschichten, dem Vorhandensein von Mikrovilli auf ihrer apikalen (oberen) Oberfläche und ihrer Fähigkeit, sich selbst zu organisieren, wenn sie in 3D zu Hohlkugeln gezüchtet werden. In ihrem Bericht spekulierten die Autoren, dass die „histotypische Expression“, durch die MDCK-Zellen Strukturen bildeten, die an ihr Ursprungsgewebe erinnerten, auch für die Untersuchung anderer Gewebe fruchtbar gemacht werden könnte. Die folgenden Jahrzehnte haben ihnen weitgehend Recht gegeben, auch wenn sich das Repertoire zur Untersuchung der Organisation und des Verhaltens von Zellen innerhalb von Geweben erheblich erweitert hat.[12]
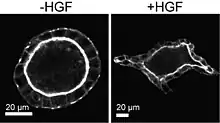
In den 1970er Jahren fand die MDCK-Zelllinie neue Verwendung als Modell für Säugetierepithelgewebe. Im Jahr 1982 zeigten Mina Bissell und Kollegen, dass MDCK-Monolayer auf die Zugabe einer Kollagenschicht (eine so genannte „Sandwich-Kultur“) mit Proliferation und der Bildung hohler Röhren reagierten,[13] was zum ersten Mal darauf hindeutete, dass die Zelllinie auf eine 3D-Umgebung reagieren würde, indem sie sich selbst zu einer entsprechenden 3D-Struktur organisierte, die an Nierentubuli erinnerte. In den folgenden Jahren wurde gezeigt, dass die Kultur von MDCK-Zellen, die vollständig in Kollagen eingebettet waren, Hohlkugeln oder Acini hervorbrachte[14] – einfache epitheliale Monoschichten mit einem definierten Inneren und Äußeren. Die Tatsache, dass MDCK-Zellen unter diesen Bedingungen keine Tubuli bildeten, blieb jedoch bis zu einem späteren Zeitpunkt unerklärt.
Zur gleichen Zeit in den 1980er Jahren waren Biologen, die sich mit der Zellmotilität befassten, auf ein interessantes und reproduzierbares Verhalten von Zellen in Kultur gestoßen: die Streuungsreaktion. Epithelzellen wachsen in der Kultur normalerweise in engen Clustern. Sie konnten jedoch dazu gebracht werden, Zell-Zell-Kontakte zu unterbrechen und sich zu strecken und zu bewegen, wenn sie einem Streufaktor ausgesetzt waren, der von mesenchymalen Zellen wie den Schweizer 3T3-Fibroblasten abgesondert wurde.[15] Dies wurde 1987 von der Gruppe von Julia Gray am besten beschrieben.[16] Zur gleichen Zeit, Mitte der 1980er Jahre, wurde von der Gruppe von Walter Birchmeier berichtet, dass ein monoklonaler Antikörper Zell-Zell-Kontakte unterbricht und die Polarität von Zellen in Kultur von vorne nach hinten verändert.[17][18] Das Ziel dieses Antikörpers wurde später als eine Komponente von Zell-Zell-Verbindungen, E-Cadherin, identifiziert.[19] Diese unterschiedlichen Beobachtungen fügten sich schließlich zu einem widerstandsfähigen Paradigma für Zellmotilität und Zellpolarität zusammen. Epithelzellen sind in der Regel nicht beweglich, können aber durch Hemmung der Zell-Zell-Verbindungen oder durch Zugabe von Wachstumsfaktoren, die eine Streuung bewirken, beweglich werden.[20] Beide Vorgänge sind reversibel, und in beiden Fällen werden die Zell-Zell-Verbindungen aufgebrochen.
1991 berichteten Lelio Orci und Kollegen erstmals über die Reaktion von MDCK-Acini in 3D-Kultur auf den Streufaktor.[21] Sie kultivierten Acini von MDCK-Zellen in Kollagengelen mit oder ohne Schweizer 3T3-Fibroblasten, in denen Medien ausgetauscht werden konnten, die Zelltypen aber nicht in direktem Kontakt standen. Diese als Kokultur bezeichnete Zellkulturstrategie veranlasste die MDCK-Acini zu einer verzweigten Morphogenese, bei der sich die Zellen zu einem Netzwerk miteinander verbundener Röhren anordnen, das der Entwicklung vieler Gewebe ähnelt.[22] Im selben Jahr wurde nachgewiesen, dass es sich bei dem Streufaktor um ein bereits zuvor beschriebenes Protein handelt, das von Fibroblasten sezerniert wird, den Hepatozyten-Wachstumsfaktor (HGF).[23] Diese Arbeit löste ein herausragendes Rätsel der MDCK-Kultur, da das Gewebe, aus dem diese Zellen stammen, röhrenförmig ist, sie sich jedoch zuvor nur in 3D-Kultur zu kugelförmigen Azini entwickelt hatten. Über dieses unmittelbare Paradoxon hinaus wurde ein entscheidender Zusammenhang zwischen der akuten Induktion der Zellmotilität in der 2D-Kultur durch den Streufaktor und seiner Auswirkung auf die räumliche Organisation von Geweben in 3D hergestellt. Diese Verbindung ist nach wie vor von Bedeutung, da sie ein Bindeglied zwischen genau definierten Mechanismen der Zellmotilität in 2D und komplexen Umstrukturierungen in 3D darstellt, deren Regulierung noch nicht vollständig geklärt ist.
Verzweigende Zellentwicklung
Um das Jahr 2000 wurde das Verständnis der Biologie von MDCK-Zellen in 3D-Kultur vor allem durch das Labor von Keith Mostov gefördert. Diese Gruppe hat sich auf die Regulierung der Zellpolarität und ihre Auswirkungen auf die Verzweigungsmorphogenese konzentriert.[24][25] In der Tat haben die Arbeiten der Mostov-Gruppe jahrzehntelanges Wissen über die räumliche Trennung von Zellfunktionen und ihre molekularen Marker zu einem bemerkenswerten Modell für die Entstehung und Homöostase der Zellpolarität in Geweben zusammengefasst.[26][27] Im Jahr 2003 berichtete die Mostov-Gruppe über die erste umfassende Darstellung, die die Verzweigungsmorphogenese mit den Merkmalen der apikal-basalen Polarität in Verbindung bringt[28] und stellte fest, dass MDCK-Zellen während des Beginns der Verzweigungsmorphogenese nicht den Kontakt zu ihren Nachbarn verlieren, sondern dass die kanonischen Marker der Zellpolarität vorübergehend verloren gehen. Ein Ergebnis dieser Polaritätsverschiebung ist die Neuausrichtung der Zellteilung entlang eines neu wachsenden Zellzweigs, um die Tochterzellen korrekt zu positionieren und die Verzweigung fortzusetzen. Die Zellmotilität, durch die MDCK-Zellen Verzweigungen erzeugen und verlängern, wurde mit diesen Polaritätsänderungen in Verbindung gebracht.
Diese Erkenntnisse wurden in ein Modell für die Verzweigungsmorphogenese integriert, das sich auf die vorübergehende Neuordnung der Zellpolaritätssignalgebung konzentriert. Dieses Modell wurde informell als Mostov-Weg bezeichnet. Auf diese Weise können normalerweise unbewegliche Zellen Ausstülpungen erzeugen und gemeinsam wandern, gefolgt von einer Redifferenzierung und der Bildung von Hohlröhren. Zur Untermauerung dieses Modells haben Mostov und Kollegen festgestellt, dass die Wirkung von HGF auf MDCK-Acini einen teilweisen Übergang von epithelialen zu mesenchymalen Zellphänotypen auslöst.[29] Dieses Argument stützt sich auf ein etabliertes Signalprogramm, das als epithelialer zu mesenchymaler Übergang (EMT) bezeichnet wird und durch das sessile Epithelzellen beweglich werden und Zell-Zell-Kontakte abbrechen.[30] EMT wurde als die transkriptionelle Signalkaskade vorgeschlagen, die die Zellstreuung antreibt, obwohl frühere Forscher die beiden nicht miteinander in Verbindung brachten[31][32] Angesichts des Unterschieds, dass bei den Azinien in 3D die Zell-Zell-Verbindungen nicht reißen, ist es unklar, wie das EMT-Konzept genau mit der Verzweigungsmorphogenese in Verbindung gebracht werden kann.
Die Mostov-Gruppe hat auch untersucht, wie HGF die Zellmotilität während der Verzweigungsmorphogenese bei MDCK aktiviert.[33][34] Ihre Studien haben gezeigt, dass die Verzweigungsmorphogenese den Transkriptionsfaktor Erk benötigt, der der Mitogen-aktivierten Proteinkinase-Kaskade nachgeschaltet ist, einem gut definierten Signaltransduktionsweg, der an der Zellmotilität und -proliferation beteiligt ist.[35] Die genaue Zellmotilitätsmaschinerie, die für die Verzweigungsmorphogenese der MDCK verantwortlich ist, wurde von der Mostov-Gruppe nicht spezifiziert, abgesehen von der Notwendigkeit eines Signalproteins, das an der Regulierung der kleinen GTPase Rho beteiligt ist.[30] Darüber hinaus hat das Gardel-Labor gezeigt, dass die invasive Motilität von MDCK-Zellen in den Azinien Dia1 erfordert, das die Zelladhäsion an einzelnen Kollagenfibrillen reguliert.
In der Zwischenzeit haben andere Gruppen die Notwendigkeit von Zell-ECM-Adhäsionsproteinen oder deren Regulatoren in der MDCK-Verzweigungsmorphogenese nachgewiesen.[36][37] Unter Verwendung eines modifizierten Protokolls für die MDCK-Zellkultur und die Verzweigungsmorphogenese wiesen Gierke und Wittman nach, dass die Mikrotubuli-Dynamik für die Regulierung der frühen Schritte der Verzweigung erforderlich ist.[38] Sie beobachteten eine mangelhafte adhäsive Kopplung der Zellen an die Kollagenmatrix, wenn die Mikrotubuli dereguliert waren. Dieser Phänotyp deutet darauf hin, wie wichtig es ist, dass die entsprechenden Zelladhäsions- und Protrusionsproteine an die Zellfront transportiert werden, wenn die Verzweigungsmorphogenese eingeleitet wird. Zusammen mit den Beobachtungen der Mostov-Gruppe bestätigt diese Arbeit, dass die Zellpolarität für die Homöostase der MDCK-Azinuszellen sowie für das Migrationsverhalten während der Verzweigungsmorphogenese unerlässlich ist.
Weblinks
Einzelnachweise
- J. D. Dukes, P. Whitley, A. D. Chalmers: The MDCK variety pack: choosing the right strain. In: BMC cell biology. Band 12, Oktober 2011, S. 43, doi:10.1186/1471-2121-12-43, PMID 21982418, PMC 3209442 (freier Volltext).
- L. E. O’Brien, M. M. Zegers, K. E. Mostov: Opinion: Building epithelial architecture: insights from three-dimensional culture models. In: Nature reviews. Molecular cell biology. Band 3, Nummer 7, Juli 2002, S. 531–537, doi:10.1038/nrm859, PMID 12094219.
- D. A. Volpe: Drug-permeability and transporter assays in Caco-2 and MDCK cell lines. In: Future Medicinal Chemistry. Band 3, Nummer 16, Dezember 2011, S. 2063–2077, doi:10.4155/fmc.11.149, PMID 22098353.
- A. J. Eisfeld, G. Neumann, Y. Kawaoka: Influenza A virus isolation, culture and identification. In: Nature protocols. Band 9, Nummer 11, November 2014, S. 2663–2681, doi:10.1038/nprot.2014.180, PMID 25321410.
- J. P. Gregersen, H. J. Schmitt, H. Trusheim, M. Bröker: Safety of MDCK cell culture-based influenza vaccines. In: Future microbiology. Band 6, Nummer 2, Februar 2011, S. 143–152, doi:10.2217/fmb.10.161, PMID 21366415.
- A. Doroshenko, S. A. Halperin: Trivalent MDCK cell culture-derived influenza vaccine Optaflu (Novartis Vaccines). In: Expert review of vaccines. Band 8, Nummer 6, Juni 2009, S. 679–688, doi:10.1586/erv.09.31, PMID 19485748.
- ATCC. In: ATCC. Abgerufen am 28. August 2017.
- Irving J. Green: Serial Propagation of Influenza B (Lee) Virus in a Transmissible Line of Canine Kidney Cells. In: Science. 1962, Band 138, Nummer 3536, S. 42–43 doi:10.1126/science.138.3536.42.
- Charles R. Gaush, Walter L. Hard, Thomas F. Smith: Characterization of an Established Line of Canine Kidney Cells (MDCK). In: Proceedings of the Society for Experimental Biology and Medicine. 122. Jahrgang, Nr. 3, 1. Juli 1966, ISSN 0037-9727, S. 931–935, doi:10.3181/00379727-122-31293, PMID 5918973 (englisch, sagepub.com).
- J. E. Moulton, Linda M. Frazier: Deoxyribonucleic acid and protein changes in dog kidney cells infected with infectious canine hepatitis virus. In: Virology. 1961 doi:10.1016/0042-6822(61)90226-4.
- Joseph Leighton, Larry W. Estes, Sunder Mansukhani, Zbynek Brada: A cell line derived from normal dog kidney (MDCK) exhibiting qualities of papillary adenocarcinoma and of renal tubular epithelium. In: Cancer. 1970, Band 26, Nummer 5, S. 1022–1028 doi:10.1002/1097-0142(197011)26:5<1022::aid-cncr2820260509>3.0.co;2-m.
- E. R. Shamir, A. J. Ewald: Three-dimensional organotypic culture: experimental models of mammalian biology and disease. In: Nature reviews. Molecular cell biology. Band 15, Nummer 10, Oktober 2014, S. 647–664, doi:10.1038/nrm3873, PMID 25237826, PMC 4352326 (freier Volltext).
- H. G. Hall, Deborah A. Farson, Mina J. Bissell: Lumen formation by epithelial cell lines in response to collagen overlay: a morphogenetic model in culture. In: Proceedings of the National Academy of Sciences. 1982, Band 79, Nummer 15, S. 4672–4676 doi:10.1073/pnas.79.15.4672.
- Eliah R. Shamir, Andrew J. Ewald: Three-dimensional organotypic culture: experimental models of mammalian biology and disease. In: Nature Reviews Molecular Cell Biology. 2014, Band 15, Nummer 10, S. 647–664 doi:10.1038/nrm3873.
- Michael Stoker, Marion Perryman: An epithelial scatter factor released by embryo fibroblasts. In: Journal of Cell Science. 1985, Band 77, Nummer 1, S. 209–223 doi:10.1242/jcs.77.1.209.
- Michael Stoker, Ermanno Gherardi, Marion Perryman, J. A. Gray: Scatter factor is a fibroblast-derived modulator of epithelial cell mobility. In: Nature. 1987, Band 327, Nummer 6119, S. 239–242 doi:10.1038/327239a0.
- J. Behrens, Walter Birchmeier, Steven L. Goodman, Beat A. Imhof: Dissociation of Madin-Darby canine kidney epithelial cells by the monoclonal antibody anti-arc-1: mechanistic aspects and identification of the antigen as a component related to uvomorulin. In: Journal of Cell Biology. 1985, Band 101, Nummer 4, S. 1307–1315 doi:10.1083/jcb.101.4.1307.
- Beat A. Imhof, H. Peter Vollmers, Simon J. Goodman, Walter Birchmeier: Cell-cell interaction and polarity of epithelial cells: Specific perturbation using a monoclonal antibody. In: Cell. 1983 doi:10.1016/0092-8674(83)90099-5.
- J. Behrens, Marcus Mareel, F. Van Roy, Walter Birchmeier: Dissecting tumor cell invasion: epithelial cells acquire invasive properties after the loss of uvomorulin-mediated cell-cell adhesion. In: Journal of Cell Biology. 1989, Band 108, Nummer 6, S. 2435–2447 doi:10.1083/jcb.108.6.2435.
- Jean Paul Thiery, Hervé Acloque, Ruby Yun-Ju Huang, M. Angela Nieto: Epithelial-Mesenchymal Transitions in Development and Disease. In: Cell. 2009, Band 139, Nummer 5, S. 871–890 doi:10.1016/j.cell.2009.11.007.
- Ruggero Montesano, Gerhard Schaller, Lelio Orci: Induction of epithelial tubular morphogenesis in vitro by fibroblast-derived soluble factors. In: Cell. 1991, Band 66, Nummer 4, S. 697–711 doi:10.1016/0092-8674(91)90115-F.
- Markus Affolter, Rolf Zeller, Emmanuel Caussinus: Tissue remodelling through branching morphogenesis. In: Nature Reviews Molecular Cell Biology. 2009, Band 10, Nummer 12, S. 831–842 doi:10.1038/nrm2797.
- Kerstin Weidner, Naokatu Arakaki, Gerd K. Hartmann, Joël Vandekerckhove, S. Weingart, Hans L. Rieder, C. Fonatsch, Hirohito Tsubouchi, Tomoyuki Hishida, Yasushi Daikuhara: Evidence for the identity of human scatter factor and human hepatocyte growth factor. In: Proceedings of the National Academy of Sciences. 1991, Band 88, Nummer 16, S. 7001–7005 doi:10.1073/pnas.88.16.7001.
- DM Bryant, KE Mostov: From cells to organs: building polarized tissue. In: Nat Rev Mol Cell Biol. 9. Jahrgang, Nr. 11, 2008, S. 887–901, doi:10.1038/nrm2523, PMID 18946477, PMC 2921794 (freier Volltext).
- Lucy Erin O’Brien, Mirjam M. P. Zegers, Keith E. Mostov: Building epithelial architecture: insights from three-dimensional culture models. In: Nature Reviews Molecular Cell Biology. 3. Jahrgang, Nr. 7, 2002, S. 531–537, doi:10.1038/nrm859, PMID 12094219.
- Fernando Martin-Belmonte, Ama Gassama, Anirban Datta, Wei Yu, Ursula Rescher, Volker Gerke, Keith Mostov: PTEN-mediated apical segregation of phosphoinositides controls epithelial morphogenesis through Cdc42. In: Cell. 128. Jahrgang, Nr. 2, 2007, S. 383–397, doi:10.1016/j.cell.2006.11.051, PMID 17254974, PMC 1865103 (freier Volltext).
- David M. Bryant, Julie Roignot, Anirban Datta, Arend W. Overeem, Minji Kim, Wei Yu, Xiao Peng, Dennis J. Eastburn, Andrew J. Ewald, Zena Werb, Keith E. Mostov: A molecular switch for the orientation of epithelial cell polarization. In: Developmental Cell. 31. Jahrgang, Nr. 2, 2014, S. 171–187, doi:10.1016/j.devcel.2014.08.027, PMID 25307480, PMC 4248238 (freier Volltext).
- Wei Yu, Lucy E. O’Brien, Fei Wang, Henry Bourne, Keith E. Mostov, Mirjam M.P. Zegers: Hepatocyte growth factor switches orientation of polarity and mode of movement during morphogenesis of multicellular epithelial structures. In: Molecular Biology of the Cell. 14. Jahrgang, Nr. 2, 2003, S. 748–763, doi:10.1091/mbc.E02-06-0350, PMID 12589067, PMC 150005 (freier Volltext).
- Mirjam M.P. Zegers, Lucy E. O’Brien, Wei Yu, Anirban Datta, Keith E. Mostov: Epithelial polarity and tubulogenesis in vitro. In: Trends in Cell Biology. 13. Jahrgang, Nr. 4, 2003, S. 169–176, doi:10.1016/S0962-8924(03)00036-9, PMID 12667754.
- J. P. Thiery, H. Acloque, R. Y. Huang, M. A. Nieto: Epithelial-mesenchymal transitions in development and disease. In: Cell. Band 139, Nummer 5, November 2009, S. 871–890, doi:10.1016/j.cell.2009.11.007, PMID 19945376.
- Elzbieta Janda, Kerstin Lehmann, Iris Killisch, Martin Jechlinger, Michaela Herzig, Julian Downward, Hartmut Beug, Stefan Grünert: Ras and TGFβ cooperatively regulate epithelial cell plasticity and metastasis. In: The Journal of Cell Biology. 156. Jahrgang, Nr. 2, 2002, S. 299–314, doi:10.1083/jcb.200109037, PMID 11790801, PMC 2199233 (freier Volltext).
- Raghu Kalluri: Epithelial-mesenchymal transition and its implications for fibrosis. In: Journal of Clinical Investigation. 112. Jahrgang, Nr. 12, 2003, S. 1776–1784, doi:10.1172/JCI20530, PMID 14679171, PMC 297008 (freier Volltext).
- Lucy Erin O’Brien: ERK and MMPs sequentially regulate distinct stages of epithelial tubule development. In: Developmental Cell. 7. Jahrgang, Nr. 1, 2004, S. 21–32, doi:10.1016/j.devcel.2004.06.001, PMID 15239951.
- M. Kim, A. M. Shewan, A. J. Ewald, Z. Werb, K. E. Mostov: p114RhoGEF governs cell motility and lumen formation during tubulogenesis through a ROCK–myosin-II pathway. In: Journal of Cell Science. 128. Jahrgang, Nr. 23, 2015, S. 4317–4327, doi:10.1242/jcs.172361, PMID 26483385, PMC 4712812 (freier Volltext).
- Emmanuel Vial, Erik Sahai, Christopher J. Marshall: ERK-MAPK signaling coordinately regulates activity of Rac1 and RhoA for tumor cell motility. In: Cancer Cell. 4. Jahrgang, Nr. 1, 2003, S. 67–79, doi:10.1016/S1535-6108(03)00162-4, PMID 12892714.
- Michael P. Hunter, Mirjam M. Zegers: Pak1 regulates branching morphogenesis in 3D MDCK cell culture by a PIX and β1-integrin-dependent mechanism. In: American Journal of Physiology. Cell Physiology. 299. Jahrgang, Nr. 1, 2010, S. C21–C32, doi:10.1152/ajpcell.00543.2009, PMID 20457839, PMC 2904258 (freier Volltext).
- Si-Tse Jiang, Sue-Jean Chiu, Hong-Chen Chen, Woei-Jer Chuang, Ming-Jer Tang: Role of α 3 β 1 integrin in tubulogenesis of Madin-Darby canine kidney cells. In: Kidney International. 59. Jahrgang, Nr. 5, 2001, S. 1770–1778, doi:10.1046/j.1523-1755.2001.0590051770.x, PMID 11318947.
- Sarah Gierke, Torsten Wittmann: EB1-recruited microtubule+ TIP complexes coordinate protrusion dynamics during 3D epithelial remodeling. In: Current Biology. 22. Jahrgang, Nr. 9, 2012, S. 753–762, doi:10.1016/j.cub.2012.02.069, PMID 22483942, PMC 3350573 (freier Volltext).