Langnasengürteltiere
Die Langnasen- oder Weichgürteltiere (Dasypus) sind eine Säugetiergattung mit sieben Arten aus der Gruppe der Gürteltiere (Dasypoda). Zu dieser Gattung gehört auch mit dem Neunbinden-Gürteltier die wohl bekannteste und als einzige dieser Gruppe auch in Nordamerika vorkommende Art, alle anderen Vertreter sind nur in Südamerika verbreitet. Sie repräsentieren die einzigen, heute noch bestehenden Vertreter der Familie der Dasypodidae. Diese Gürteltiere leben in offenen Gras- und Busch- beziehungsweise in geschlossenen Waldlandschaften und ernähren sich überwiegend von Insekten. Sie legen teils weit verzweigte, unterirdische Baue an und sind je nach Art tag- oder nachtaktiv. Die Stammesgeschichte der Langnasengürteltiere als Gattung reicht gesichert bis ins Pliozän vor mehr als 2,5 Millionen Jahren zurück, möglicherweise aber auch bis in das Obere Miozän. Es gibt aber nur wenige Fossilfunde. Frühe Verwandte traten hingegen schon im Paläogen auf.
| Langnasengürteltiere | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Neunbinden-Gürteltier (Dasypus novemcinctus) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name der Familie | ||||||||||||
| Dasypodidae | ||||||||||||
| J. E. Gray, 1821 | ||||||||||||
| Wissenschaftlicher Name der Unterfamilie | ||||||||||||
| Dasypodinae | ||||||||||||
| J. E. Gray, 1821 | ||||||||||||
| Wissenschaftlicher Name der Gattung | ||||||||||||
| Dasypus | ||||||||||||
| Linnaeus, 1758 |
Beschreibung

Langnasengürteltiere sind kleine bis große Gürteltiere mit Kopf-Rumpf-Längen von 24 bis 57 cm und einem Gewicht von 1 bis 10 kg, fossile Arten waren teilweise auch größer. Die Schwanzlänge erreicht in der Regel mindestens die Hälfte der Körperlänge. Charakteristisch ist der Kopf mit einer auffallend langen, spitzen Schnauze, die oft mehr als 50 % der gesamten Schädellänge einnimmt. Das Gebiss besteht aus untypischen, nicht für Säugetiere charakteristischen Zahnbildungen, die kein Zahnschmelz und nur eine Wurzel aufweisen. Sowohl im Ober als auch im Unterkiefer befinden sich je Kieferhälfte 7 bis 9 solcher Zähne, insgesamt also 28 bis 36. Die Rückenpanzerung, die überwiegend glatt, hoch und deutlich gerundet sowie meist gelblich-braun gefärbt ist, besitzt sechs bis elf bewegliche Segmente oder Bänder zwischen einem festen Schulter- und einen ebensolchen Beckenteil. Sie besteht an den starren Panzerteilen aus rundlichen bis rosettenförmigen, an den beweglichen aus viereckigen Knochenplättchen, sogenannte Osteoderme. Überdeckt wird die Panzerstruktur von kleinen Hornschildchen von rundlicher (fester Panzer) oder dreieckiger Gestalt (Bänder) mit nach hinten weisender Spitze. Am festen Panzer fassen die Hornschildchen bei den Langnasengürteltieren mehrere Knochenplättchen ein, wodurch ein mitunter irreguläres und nicht strikt lineares Muster entsteht.[1][2] Darüber hinaus haben die Langnasengürteltiere eine Panzerung an der Oberseite des Kopfes und im Beckenbereich. Der lange Schwanz ist meist von 12 bis 15 Knochenringen umgeben, bestehend aus häufig zwei Reihen kleiner Knochenschildchen; diese Knochenringe nehmen zwei Drittel der Schwanzlänge ein, das Ende des Schwanzes ist spitz geformt. Die Beine sind verhältnismäßig kurz, an den Vorderfüßen haben die Langnasengürteltiere vier und an den Hinterfüßen fünf Zehen, alle mit scharfen Krallen, nur das Kappler-Gürteltier besitzt noch eine rudimentäre vordere fünfte Zehe. Insgesamt treten vier Milchdrüsen auf, ein Paar hinter den Vorder- und ein zweites vor den Hinterbeinen.[3][4]
Verbreitung
Das Verbreitungsgebiet der Langnasengürteltiere reicht von den südlichen USA bis ins südliche Südamerika, sie sind damit die am weitesten verbreitete Gürteltiergattung. Als Lebensraum dienen häufig offene Landschaften wie Savannen und Steppen, aber auch tropische Regenwälder und Bergnebelwälder.[3]
Lebensweise
.jpg.webp)
Territorialverhalten
Diese Tiere sind in der Regel einzelgängerisch und sowohl tag- als auch nachtaktiv. Sie leben in selbst gegrabenen, bis zu 7 m langen, unterirdischen Bauen und verlassen diesen vorwiegend zur Nahrungssuche. Mehrere Tiere können sich einen Bau teilen, generell sind sie nicht territorial und haben kein festes Revier, jedoch gibt es Heimatgebiete (home ranges) von bis zu 12 ha Größe. Die Langnasengürteltiere können schnell laufen und teilweise gute Schwimmer, im Falle eines Angriffs versuchen sie, in ihren Bau zu flüchten. Ist dies nicht möglich, legen sie sich auf den Boden, um den weichen Bauch vor Angreifern zu schützen und rammen die Krallen in den Boden, wodurch sie kaum bewegt werden können.[3]
Ernährung
Langnasengürteltiere ernähren sich in erster Linie von Ameisen, Termiten und Käfern, darüber hinaus nehmen sie auch andere Insekten und Spinnen zu sich, insgesamt sind sie eher opportunistische Insektenfresser. Gelegentlich werden auch kleine Wirbeltiere und Vogeleier vertilgt. Weiterhin ist die Aufnahme von Sand und Lehm bekannt, was zum Ausgleich des Mineralienhaushaltes dient.[3]
Fortpflanzung
Nach bis zu 140-tägiger Tragzeit bringt das Weibchen vier bis zwölf Jungtiere zur Welt, welche zwischen 40 und 85 g wiegen. Diese werden nach rund drei Monaten entwöhnt und sind mit etwa einem Jahr geschlechtsreif. Die Lebenserwartung der Langnasengürteltiere dürfte bei zwölf bis fünfzehn Jahren liegen. Bemerkenswert bei diesen Gürteltiervertretern ist die verzögerte Implantation der Eizelle, die unter Stressbedingungen einsetzt. Dies führt dazu, dass die Jungen teilweise erst bis zu 32 Monate nach der letzten Befruchtung geboren werden können. Außerdem ist Polyembryonie bekannt, unter anderem beim Neunbinden-Gürteltier, das immer vier genetisch identische Jungtiere zur Welt bringt.[3]
Systematik
Innere Systematik der Gürteltiere nach Gibb et al. 2015[5]
|
Die Langnasengürteltiere (Dasypus) stellen eine Gattung innerhalb der Familie der Dasypodidae dar, die Untersuchungen aus dem Jahr 2015 zufolge eine der beiden Familien der Gürteltiere (Dasypoda) bildet. Die Gürteltiere wiederum gehören zur Ordnung der Gepanzerten Nebengelenktiere (Cingulata). Die Dasypodidae gelten als rezent monotypisch, enthalten aber zahlreiche ausgestorbenen Formen. Zu diesen fossilen Verwandten zählen unter anderem Stegotherium, das weitgehend aus dem Miozän bekannt ist und mehrere Arten umfasste,[6][7] ebenso wie Anadasypus aus dem Mittelmiozän. Weitere Gattungen stellen Pliodasypus aus dem Pliozän und Propraopus aus dem Pleistozän dar.[8] Anadasypus, Propraopus und Pliodasypus sind dabei wesentlich enger mit Dasypus verwandt, die beiden letztgenannten bilden die Schwestergruppe, und formen mit diesem zusammen die Tribus der Dasypodini, Stegotherium steht etwas außerhalb in der Verwandtschaftsfolge und gehört der Tribus der Stegotheriini an.[9][10] Laut molekulargenetischen Untersuchungen hatten sich die Dasypodidae bereits im Mittleren Eozän vor rund 45 Millionen Jahren von den Linien der anderen Gürteltiere abgetrennt. Als nächstverwandte Gruppe werden die Chlamyphoridae angesehen, die alle anderen Gattungen der heutigen Gürteltiere einschließen.[5] Insgesamt können die Langnasengürteltiere und ihre näheren Verwandten aufgrund morphologischer Merkmale und der genetischen Analysen als eine der urtümlichsten Gruppen innerhalb der Gürteltiere und der Gepanzerten Nebengelenktiere angesehen werden.[11]
In der Regel werden heute sieben noch lebende Arten unterschieden, die zu drei verschiedenen Untergattungen gestellt werden:[12][13][14]
- Untergattung Dasypus Linnaeus, 1758
Innere Systematik der Langnasengürteltiere nach Gibb et al. 2015[5]
|
- Südliches Siebenbinden-Gürteltier (Dasypus hybridus (Desmarest, 1804)); Paraguay, Uruguay und nördliches und mittleres Argentinien
- Neunbinden-Gürteltier (Dasypus novemcinctus Linnaeus, 1758); vom Südosten der USA bis Argentinien
- Savannen-Gürteltier (Dasypus sabanicola Mondolfi, 1968); Llanos-Region in Kolumbien und Venezuela
- Siebenbinden-Gürteltier oder Nördliches beziehungsweise Brasilianisches Siebenbinden-Gürteltier (Dasypus septemcinctus Linnaeus, 1758); südliches Brasilien, Bolivien und Paraguay
- Yungas-Gürteltier (Dasypus mazzai Yepes, 1933); Nordosten von Argentinien
- Untergattung Hyperoambon Peters, 1864
- Kappler-Gürteltier oder Kappler-Weichgürteltier (Dasypus kappleri Krauss, 1862); Kolumbien und Nordost-Brasilien
- Untergattung Cryptophractus Fitzinger, 1856
- Pelzgürteltier (Dasypus pilosus (Fitzinger, 1856)); Peru
Des Weiteren werden folgende fossilen Arten unterschieden:
- Dasypus bellus Simpson, 1930
- Dasypus punctatus Lund, 1840
Teilweise wird diskutiert, ob das Pelzgürteltier aufgrund abweichender morphologischer und anatomischer Merkmale eventuell einer eigenständigen Gattung angehört, die dann mit Cryptophractus zu bezeichnen wäre, was auch der ursprünglichen Benennung von Leopold Fitzinger entspräche.[15] Die Forderung nach einem eigenen Gattungsstatus kam schon mehrfach auf,[16] im Jahr 2015 vorgestellte Untersuchungen zur Phylogenese mit Hilfe skelettanatomischer Merkmale unterstützen diese.[11] Eine molekulargenetische Studie aus dem gleichen Jahr widerspricht dem jedoch und sieht das Pelzgürteltier tief in der Gattung Dasypus verankert. Bezüglich des Neunbinden-Gürteltiers könnte laut der gleichen genetischen Studie eine Population aus Französisch-Guayana eine eigenständige Art repräsentieren, die sich bereits vor rund 3,7 Millionen Jahren von der eigentlichen Linie des Neunbinden-Gürteltiers und dessen unmittelbarer Verwandtschaft abgetrennt hatte.[5] Die Eigenständigkeit der Guayana-Gruppe ließ sich auch durch die Auswertung schädelmorphologischer Daten belegen.[17] Weiteren genetischen Untersuchungen zufolge ist das Yungas-Gürteltier identisch mit dem Savannen-Gürteltier, so dass letztere Form in erstere eingegliedert werden müsste. Die neue, umfassendere Art bestünde dann aus mehreren, bis zu 3000 km voneinander getrennten Populationen.[18]
Eine auf anatomischen Merkmalen basierende systematische Revision der Langnasengürteltiere aus dem Jahr 2018 unter Berücksichtigung von über 2120 Individuen der Gattung erkennt entgegen der oben dargestellten klassischen Gliederung insgesamt acht Arten an. Dabei wurde das Kappler-Gürteltier in drei Arten aufgespalten (Der Vorschlag wurde bereits im Jahr 2016 unterbreitet,[19] der überwiegende Teil der Fachwissenschaftler folgte ihm aber bisher nicht.[20]). Das Südliche Siebenbinden-Gürteltier hingegen schlugen die Autoren der Revision dem Siebenbinden-Gürteltier als Unterart zu. Im Ergebnis dieser 2018er Studie würden sich die Langnasengürteltiere folgendermaßen unterteilen:[21]
- Dasypus beniensis Lönnberg, 1942
- Dasypus kappleri Krauss, 1862
- Dasypus mazzai Yepes, 1933
- Dasypus novemcinctus Linnaeus, 1758
- Dasypus pastasae (Thomas, 1901)
- Dasypus pilosus (Fitzinger, 1856)
- Dasypus sabanicola Mondolfi, 1968
- Dasypus septemcinctus Linnaeus, 1758
Stammesgeschichte
Ursprung und frühe Entwicklungslinien

Die Gepanzerten Nebengelenktiere sind die ältesten bekannten Vertreter der Nebengelenktiere (Xenarthra) und schon aus dem Paläogen des heutigen Brasilien nachgewiesen. Die bisher frühesten bekannten Funde werden gemeinhin der Gattung Riostegotherium zugewiesen. Sie bildet einen Teil der Tribus der Astegotheriini und gehört damit in das nähere Verwandtschaftsumfeld der Langnasengürteltiere. Die Funde stammen aus einer Spaltenfüllung der Itaboraí-Formation nahe São José de Itaboraí in Itaboraí im südöstlichen Brasilien. Dokumentiert wurden einzelne Knochenplättchen und postcraniales Skelettmaterial. Ihr Alter beträgt rund 58 Millionen Jahre.[22][23] Die Osteoderme oder Knochenplättchen dieser frühen Formen glichen in ihrem Aufbau mit ihrer oberen und unteren Knochenschicht sowie einer mittleren weicheren Schicht schon weitgehend denen der Langnasengürteltiere. Von denen der anderen Gürteltiere aus der Familie der Chlamyphoridae unterscheiden sie sich durch die wenig entwickelte Mittelschicht, während sie bei letzteren deutlicher aufgelockert ist. Bei den Glyptodontidae wiederum war diese von luftgefüllten Kammern durchsetzt.[24][25] Die weitere Entwicklung der frühen Verwandtschaft der Langnasengürteltiere findet dann überwiegend im südlichen Teil von Südamerika statt. So kommen mit Prostegotherium und Stegosimpsonia im Verlauf des Unteren Eozäns Gattungen hinzu, die unter anderem von den Fundstellen Laguna Fría und La Barda am Mittellauf des Río Chubut in Patagonien aufgedeckt wurden.[26] Zeitlich folgend kann die Geste-Formation im nordwestlichen Argentinien eingeordnet werden, die in den Übergang vom Mittleren zum Oberen Eozän gehört. Hier traten Astegotherium und Parastegosimpsonia auf.[27] Ein rund 45 Millionen Jahre alter Unterkiefer von Astegotherium, einer der ältesten derartigen Belege für die Gürteltiere, aus fossilführenden Schichten eines Nebenflusses des Río Chico im südlichen Argentinien verweist darauf, dass die frühen Formen damals noch einen deutlichen Zahnschmelzüberzug auf den Zähnen besaßen.[28] Die Fundstelle Gran Barranca im zentralen Patagonien hat eine hohe Bedeutung für die frühe Stammesgeschichte der Gürteltiere. Ihre Ablagerungen datieren vom Unteren Eozän bis in das Untere Miozän. Die darin verstreuten Fundlagen erbrachten Formen wie Stegosimpsonia oder Pseudostegotherium, deren Belegmaterial aber zumeist aus isolierten Knochenplättchen besteht. Aus den oberen Fundeinheiten ist mit Stegotherium erstmals ein Vertreter der Tribus der Stegotheriini nachweisbar.[29] Bereits im Verlauf des Paläogen lassen sich frühe Formen aus dem Verwandtschaftskreis der Langnasengürteltiere in nördlicheren Bereichen von Südamerika nachweisen. Dazu gehören Knochenplättchen von Parastegosimpsonia vom Río Yurúa im zentral-östlichen Peru, die biostratigraphisch zwischen dem Oberen Eozän und dem Unteren Oligozän einzuordnen sind.[30]

Stegotherium erlangt dann in der Folgezeit große Bedeutung und tritt sehr variantenreich in Erscheinung. Zu nennen sind hier Panzerteile aus der Santa-Cruz-Formation im südlichen Patagonien, die sich im Übergang vom Unteren zum Mittleren Miozän ablagerte. Die Tiere waren möglicherweise aufgrund ihrer stark verlängerten Schnauze und den verkleinerten Zähnen an eine deutlicher auf Ameisen und Termiten basierende Lebensweise angepasst.[31][7][32] In das ausgehende Mittlere Miozän gehört die gleichfalls bedeutende Fundstelle La Venta am Mittellauf des Río Magdalena in Kolumbien. Die hier aufgeschlossene Honda-Gruppe besteht aus mehreren Gesteinseinheiten. Mit Nanoastegotherium kommt hier noch ein Angehöriger der Astegotheriini vor. Gleichzeitig sind mit Plesiodasypus und Anadasypus zwei Gattungen belegt, die in das unmittelbar nähere Verwandtschaftsumfeld der heutigen Langnasengürteltiere einzugliedern sind. Sie markieren gleichzeitig den ältesten Nachweis für das Auftreten der Tribus der Dasypodini. Ihr Alter wurde auf 13,3 beziehungsweise 12,5 Millionen Jahre bestimmt. Von Plesiodasypus sind unter anderem ein Schädel und Panzerfragmente aufgefunden worden, von Anadasypus der Rest eines Rückenpanzers.[33][34] Die gesamte weitere Entwicklung der Dasypodini war zunächst auf die tropischen und subtropischen Landschaftsräume Südamerikas beschränkt.[35] Im folgenden Pliozän und Pleistozän erweitert sich das Formenspektrum der Dasypodini. So erscheint Pliodasypus in der pliozänen San-Gregorio-Formation in Venezuela, wobei die Gattung nur auf einigen wenigen Knochenplättchen beruht.[10] Eine weitere Verbreitung und einen größeren Formenreichtum wies Propraopus im Pleistozän auf. Deren Angehörige kamen dann sowohl in den heute tropisch geprägten als auch gemäßigt-klimatischen Gebieten Südamerikas vor und sind vom Meeresspiegelniveau bis in höhere Lagen um 2800 m nachgewiesen. Die Gattung starb vermutlich erst im Unteren Holozän aus.[36][9][37]
Entwicklungsgeschichte der Gattung Dasypus
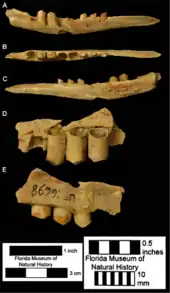
Gegenüber der langen Entwicklungsgeschichte der Verwandtschaft der heutigen Langnasengürteltiere ist die Gattung Dasypus deutlich jünger und zudem allgemein ein eher seltenes fossiles Faunenelement. Molekulargenetischen Untersuchungen zufolge trennte sich das Kappler-Gürteltier vor rund 12 Millionen Jahren im Oberen Miozän von der gemeinsamen Linie mit den anderen Vertretern der Langnasengürteltiere, die weitere Diversifizierung der Gattung folgte dann vor etwa 5 Millionen Jahren im Übergang vom Miozän zum Pliozän.[38][5] Der Ursprung der Gattung Dasypus wird im nördlichen Teil Südamerikas vermutet, da zum fraglichen Entstehungszeitraum mehrere nahe verwandte Formen hier verbreitet waren. Problematisch erscheint ein Fund eines einzelnen Knochenplättchens von einem beweglichen Band aus dem Conglomerado osífero der Ituzaingó-Formation in der argentinischen Provinz Entre Ríos. Der Conglomerado osífero wird in das ausgehende Miozän gestellt, aber sowohl in seiner Zuweisung zu Dasypus als auch in der exakten Alterseinstufung ist das Knochenplättchen umstritten, da es möglicherweise auch aus höheren Schichten verlagert sein könnte. Eindeutigere Nachweise für das Auftreten von Dasypus bereits im ausgehenden Miozän lieferte die Guanaco-Formation im äußersten Nordwesten von Argentinien. Die hier entdeckten Knochenplättchen konnten aber noch keiner genauen Art zugewiesen werden.[39] Die ältesten genauer bestimmten Überreste stammen bisher aus dem Pliozän vor über 2,5 Millionen Jahren und werden allgemein Dasypus bellus, dem „Schönen Gürteltier“, zugewiesen. Sie sind unter anderem aus Florida und Nebraska belegt.[40][10] Dasypus bellus, eine heute ausgestorbene Art, war über weite Teile Südamerikas und den Osten Nordamerikas verbreitet und wird häufig mit dem Neunbinden-Gürteltier in Verbindung gebracht, war aber deutlich größer und erreichte in der Spätphase seines Auftretens bis zu 18 kg Körpergewicht.[41][42] Neuere Untersuchungen sehen auch eine nähere Verbindung zum Kappler-Gürteltier. Im späten Pleistozän ist dann noch die ausgestorbene Art Dasypus punctatus nachgewiesen, vornehmlich in Brasilien, wo sie den bisher frühesten sicheren Nachweis der gesamten Gattung in Südamerika bildet. Dieser Art wird ebenfalls eine nahe Stellung zum Kappler-Gürteltier bescheinigt.[9] Beide heute fossilen Arten verschwanden gegen Ende des Pleistozäns vor 10.000 Jahren.
Obwohl die heutigen Langnasengürteltiere die vielfältigste Gruppe der Gürteltiere stellen, sind Überreste der rezenten Vertreter aus dem Pleistozän kaum bekannt, mit Ausnahme des Savannen-Gürteltiers, das in Venezuela bereits vor rund 25.000 Jahren lebte.[8] Ein ähnliches Alter weisen einzelne, sehr robuste Reste aus dem südlichen Brasilien auf, die wahrscheinlich mit dem Kappler-Gürteltier in Verbindung stehen.[43] Aus dem zentralen Brasilien wiederum wurden einzelne Knochenplättchen des Neunbinden-Gürteltiers aus Höhlenfundstellen des Spätpleistozäns berichtet.[44][45] Untersuchungen an Fossilmaterial, unter anderem aus der Medford-Höhle in Florida, belegen zudem die Anwesenheit des Neunbinden-Gürteltiers im ausgehenden Pleistozän in Nordamerika, möglicherweise trat die Art auch noch im frühen Holozän hier auf.[46] Die heutige Verbreitung der Langnasengürteltiere ist zum Teil ein Resultat der jüngsten Zeit. Die ersten Nachweise des Neunbinden-Gürteltiers in den USA sind aus dem Jahr 1849 bekannt. Seitdem hat sich diese Art über weite Teile des südöstlichen Nordamerikas ausgebreitet und besiedelt als erster Vertreter der Gürteltiere seit 10.000 Jahren, nach dem Ende der letzten Eiszeit, wieder den Nordkontinent Amerikas. Warum diese Gürteltierart erst so spät hier wieder auftauchte, ist bisher ungeklärt, da sie bereits wesentlich früher in Mexiko nachgewiesen ist.[3]
Taxonomie und Etymologie
Der Gattungsname Dasypus ist griechischer Herkunft und wurde von Linnaeus im Jahr 1758 eingeführt. Er stellt eine Übersetzung des aztekischen Wort Azotochtli dar, was so viel wie „Schildkrötenhase“ bedeutet. Das Wort ist über den spanischen Conquistador Francisco Hernández de Córdoba als Bezeichnung für das Neunbinden-Gürteltier überliefert und bezieht sich auf das Aussehen der Tiere. Linnaeus befand das Wort aber als nicht geeignet für eine Latinisierung, was eine Voraussetzung ist für die von ihm in seinem Werk Systema Naturae eingeführten binären wissenschaftlichen Namensbildung, und schuf deshalb ein griechisches Wort mit gleicher Bedeutung (Tautologie). Das regional häufig verwendete spanische Wort mulita bedeutet „Eselchen“ und erinnert an die eselartigen Ohren.[47][4]
Langnasengürteltiere und der Mensch
Krankheitsüberträger
Die Langnasengürteltiere gelten teilweise als Krankheitsüberträger. Bedeutend sind das Neunbinden-, das Südliche Siebenbinden und das Savannen-Gürteltier als Wirte von Mycobacterium leprae, welches die Lepra beim Menschen hervorrufen kann. In Nordamerika sind 4,1 bis 4,7 % der einzelnen Populationen betroffen. Ob die Gürteltierart aber die Krankheit tatsächlich auf den Menschen überträgt ist umstritten, im US-Bundesstaat Louisiana ist die Lepra seit etwa den 1850er Jahren endemisch, das Neunbinden-Gürteltier wurde aber erstmals in den 1920er Jahren dort beobachtet. Weiterhin infizieren sich die Langnasengürteltiere mit Trypanosoma cruzi, der als Verursacher der in Südamerika häufigen Chagas-Krankheit gilt. Auch hier ist ein Übertragungsweg über Gürteltiere nicht sicher, da bei diesen eine andere Form auftritt, welche beim Menschen bisher so gut wie nie nachgewiesen wurde. Zur Untersuchung werden Langnasengürteltiere häufig als Labortiere gehalten.[47][3]
Bedrohung und Schutz
Die verschiedenen Arten der Langnasengürteltiere werden häufig bejagt und dienen als Nahrungsressource. Teilweise gelten sie auch als Plage, die in landwirtschaftlich genutzten Flächen auf ihrer Nahrungssuche Löcher oder Baue graben, durch die sich Nutztiere möglicherweise verletzen können. Gravierender für die Langnasengürteltiere sind die Zerstörung der Lebensräume durch Ausbreitung menschlicher Siedlungen und bewirtschafteter Areale, die so häufig für schwindende Populationen sorgen. Ein Teil der Arten, so das Neunbinden-, Siebenbinden-, Kappler- und Savannen-Gürteltier ist gegenwärtig nicht ihrem Bestand bedroht, während das Südliche Siebenbinden-Gürteltier von der IUCN als gering gefährdet eingestuft wird. Allerdings gilt das Pelzgürteltier als gefährdet, während beim Yungas-Gürteltier aufgrund von Datenlücken keine Angaben gemacht werden können. Die Langnasengürteltiere sind in zahlreichen geschützten Gebieten auf den beiden amerikanischen Kontinenten vertreten.[13]
Literatur
- C. M. McDonough und W. J. Laughry: Dasypodidae (Long-nosed armadillos). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos. Lynx Edicions, Barcelona 2018, S. 30–47 ISBN 978-84-16728-08-4
Einzelnachweise
- Alfredo A. Carlini, Martín Ricardo Ciancio, John J. Flynn, Gustavo J. Scillato‐Yané und André R. Wyss: The phylogenetic and biostratigraphic significance of new armadillos (Mammalia, Xenarthra, Dasypodidae, Euphractinae) from the Tinguirirican (early oligocene) of Chile. Journal of Systematic Palaeontology 7 (4), 2009, S. 489–503
- Matthew K. Vickaryous und Brian K. Hall: Osteoderm Morphology and Development in the Nine-Banded Armadillo, Dasypus novemcinctus (Mammalia, Xenarthra, Cingulata). Journal of Morphology 267 (11), 2006, S. 1273–1283
- Mariella Superina: Biologie und Haltung von Gürteltieren (Dasypodidae). Universität Zürich, 2000, S. 1–248
- Paul Smith: The Xenarthra famalies Myrmecophagidae and Dasypodidae. Fauna Paraguay Handbook of the Mammals of Paraguay 2012, S. 1–35
- Gillian C. Gibb, Fabien L. Condamine, Melanie Kuch, Jacob Enk, Nadia Moraes-Barros, Mariella Superina, Hendrik N. Poinar und Frédéric Delsuc: Shotgun Mitogenomics Provides a Reference Phylogenetic Framework and Timescale for Living Xenarthrans. Molecular Biology and Evolution 33 (3), 2015, S. 621–642
- Timothy J. Gaudin und John R. Wible: The phylogeny of living and extinct armadillos (Mammalia, Xenarthra, Cingulata): a craniodental analysis. In: Matthew T. Carrano, Timothy J. Gaudin, Richard W. Blob und John R. Wible (Hrsg.): Amniote Paleobiology: Phylogenetic and Functional Perspectives on the Evolution of Mammals, Birds and Reptiles. Chicago 2006, University of Chicago Press, S. 153–198
- Laureano Raúl González Ruiz und Gustavo Juan Scillato-Yané: A new Stegotheriini (Mammalia, Xenarthra, Dasypodidae) from the “Notohippidian” (early Miocene) of Patagonia, Argentina. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 252 (1), 2009, S. 81–90
- Ascanio D. Rincón, Richard S. White und H. Gregory McDonald: Late Pleistocene Cingulates (Mammalia: Xenarthra) from Mene De Inciarte Tar Pits, Sierra De Perijá, Western Venezuela. Journal of Vertebrate Paleontology 28 (1), 2008, S. 197–207
- Mariela C. Castro, Ana Maria Ribeiro, Jorge Ferigolo und Max C. Langer: Redescription of Dasypus punctatus Lund, 1840 and considerations on the genus Propraopus Ameghino, 1881 (Xenarthra, Cingulata). Journal of Vertebrate Paleontology 33 (2), 2013, S. 434–447
- Mariela C. Castro, Alfredo A. Carlini, Rodolfo Sánchez und Marcelo R. Sánchez-Villagra: A new Dasypodini armadillo (Xenarthra: Cingulata) from San Gregorio Formation, Pliocene of Venezuela: affinities and biogeographic interpretations. Naturwissenschaften 101, 2014, S. 77–86, doi:10.1007/s00114-013-1131-5
- Mariela C. Castro, Martín R. Ciancio, Victor Pacheco, Rodolfo M. salas-Gismondi, J. Enrique Bostelmann und Alfredo A. Carlini: Reassessment of the hairy long-nosed armadillo “Dasypus” pilosus (Xenarthra, Dasypodidae) and revalidation of the genus Cryptophractus Fitzinger, 1856. Zootaxa 3947 (1), 2015, S. 30–48, doi:10.11646/zootaxa.3947.1.2
- Mariella Superina und John M. Aguiar: A Reference List of Common Names for the Edentates. Edentata 7, 2006, S. 33–44
- Agustín Manuel Abba und Mariella Superina: The 2009/2010 Armadillo Red List Assessment. Edentata 11 (2), 2010, S. 135–184
- Sergio F. Vizcaíno: Identificación específica de las mulitas, género Dasypus L. (Mammalia, Dasypodidae), del noroeste argentino. Descripción de una nueva especie. Mastozoologia Neotropical 2 (1), 1995, S. 5–13
- Leopold Joseph Fitzinger: Tageblatt der 32. Versammlung Deutscher Naturforscher und Ärzte in Wien, 1856, S. 123 ()
- Edentate Specialist Group: The 2004 Edentata species assessment workshop, Belo Horizonte, Minas Gerais, Brazil, December 16–17, 2004. Edentata 5, 2004, S. 3–26
- Lionel Hautier, Guillaume Billet, Benoit de Thoisy und Frédéric Delsuc: Beyond the carapace: skull shape variation and morphological systematics of long-nosed armadillos (genus Dasypus). PeerJ 5, 2017, S. e3650, doi:10.7717/peerj.3650
- Agustín M. Abba, Guillermo H. Cassini, Juan I. Túnez und Sergio F. Vizcaíno: The enigma of the Yepes’ armadillo: Dasypus mazzai, D. novemcinctus or D. yepesi? Revista del Museo Argentino de Ciencias Naturales, N. S. 20 (1), 2018, S. 83–90
- Anderson Feijó und Pedro Cordeiro-Estrela: Taxonomic revision of the Dasypus kappleri complex, with revalidations of Dasypus pastasae (Thomas, 1901) and Dasypus beniensis Lönnberg, 1942 (Cingulata, Dasypodidae). Zootaxa 4170 (2), 2016, S. 271–297 ()
- C. M. McDonough und W. J. Laughry: Dasypodidae (Long-nosed armadillo). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos. Lynx Edicions, Barcelona 2018, S. 30–47 ISBN 978-84-16728-08-4
- Anderson Feijó, Bruce D. Patterson und Pedro Cordeiro-Estrela: Taxonomic revision of the long-nosed armadillos, Genus Dasypus Linnaeus, 1758 (Mammalia, Cingulata). PLoS ONE 13 (4), 2018, S. e0195084, doi:10.1371/journal.pone.0195084
- Édison Vicente Oliveira und Lílian Paglarelli Bergqvist: A new Paleogene armadillo (Mammalia, Dasypodoidea) from the Itaboraí basin, Brazil. Associatión Paleontológica Argentina, Publicatión Especial 5, 1998, S. 35–40
- Richard L. Cifelli: Eutherian Tarsals from the Late Paleocene of Brazil. American Museum Novitates 2761, 1983, S. 1–31
- M. R. Ciancio, C. M. Krmpotic, A. C. Scarano und M. B. Epele: Internal Morphology of Osteoderms of Extinct Armadillos and Its Relationship with Environmental Conditions. Journal of Mammalian Evolution 26, 2019, S. 71–83, doi:10.1007/s10914-017-9404-y
- Lílian P. Bergqvist, Paulo Victor Luiz G. C. Pereira, Alessandra S. Machado, Mariela C. de Castro, Luiza B. Melki und Ricardo T. Lopez: Osteoderm microstructure of Riostegotherium yanei, the oldest Xenarthra. Anais da Academia Brasileira de Ciências 91 (Suppl. 2), 2019, S. e20181290, doi:10.1590/0001-3765201920181290
- Marcelo F. Tejedor, Francisco J. Goin, Javier N. Gelfo, Guillermo López, Mariano Bond, Alfredo A. Carlini, Gustave J. Scillato-Yane, Michael O. Woodburne, Laura Chornogubsky, Eugenio Aragón, Marcelo A. Reguero, Nicholas J. Czapkewski, Sergio Vincon, Gabriel M. Martin und Martin R. Ciancio: New Early Eocene Mammalian Fauna from Western Patagonia, Argentina. American Museum Novitates 3638, 2009, S. 1–43
- Martín R. Ciancio, Claudia Herrera, Alejandro Aramayo, Patricio Payrola und María J. Babot: Diversity of cingulate xenarthrans in the middle-late Eocene of Northwestern Argentina. Acta Palaeontologica Polonica 61 (3), 2016, S. 575–590, doi:10.4202/app.00208.2015
- Martín R. Ciancio, Emma C. Vieytes und Alfredo A. Carlini: When xenarthrans had enamel: insights on the evolution of their hypsodonty and paleontological support for independent evolution in armadillos. Naturwissenschaften 101, 2014, S. 715–725
- Alfredo A. Carlini, Martín R. Ciancio und Gustavo J. Scillato-Yané: Middle Eocene – Early Miocene Dasypodidae (Xenarthra) of Southern South America: faunal succession at Gran Barranca – biostratigraphy and paleoecology. In: Richard H. Madden, Alfredo A. Carlini, Maria Guiomar Vucetich und Richard F. Kay (Hrsg.): The Paleontology of Gran Barranca. Evolution and environmental change through the Middle Cenozoic of Patagonia. Cambridge University Press, 2010, S. 106–129
- Martín Ricardo Ciancio, Alfredo A. Carlini, Kenneth E. Campbell und Gustavo J. Scillato-Yané: New Palaeogene cingulates (Mammalia, Xenarthra) from Santa Rosa, Perú and their importance in the context of South American faunas Journal of Systematik Palaeontology 11 (5–6), 2013, S. 727–741 doi:10.1080/14772019.2012.704949
- Laureano Raúl González und Gustavo Juan Scillato-Yané: Una nueva especie de Stegotherium Ameghino (Xenarthra, Dasypodidae, Stegotheriini) del Mioceno de la provincia de Santa Cruz (Argentina). Ameghiniana 45 (4), 2008, S. 641–648
- Sergio F. Vizcaíno, Juan C. Fernicola und M. Susana Bargo: Paleobiology of Santacrucian glyptodonts and armadillos (Xenarthra, Cingulata). In: Sergio F. Vizcaíno, Richard F. Kay und M. Susana Bargo (Hrsg.): Early Miocene paleobiology in Patagonia: High-latitude paleocommunities of the Santa Cruz Formation. Cambridge University Press, New York, 2012, S. 194–215
- Alfredo A. Carlini, Sergio F. Vizcaíno und Gustavo J. Scillato-Yané: Armored Xenarthrans: A unique taxonomic and ecologic assemblage. In: Richard F. Kay, Richard H. Madden, Richard L. Cifelli und John J. Flynn (Hrsg.): Vertebrate Paleontology in the Neotropics. The Miocene Fauna of La Venta, Colombia. Smithsonian Institution Press, Washington, 1997, S. 213–226
- Daniel Barasoain, Laureano González-Ruiz, Alfredo Zurita und Carlos Villarroel: Oldest new Dasypodini (Xenarthra, Cingulata) provides new trails about armadillos evolutionary history. In: Historical Biology: An International Journal of Paleobiology, 2021, doi:10.1080/08912963.2021.1917566
- M. C. Castro: Sistemática y evolución de los armadillos Dasypodini (Xenarthra, Cingulata, Dasypodidae). Revista del Museo de La Plata, Sección Paleontología 15 (73), 2015, S. 1–50
- Vanessa Gregis Pitana und Ana Maria Ribeiro: Novos materiais de Propraopus Ameghino, 1881 (Mammalia, Xenarthra, Cingulata) do Pleistoceno final, Rio Grande do Sul, Brasil. Gaea 3 (2), 2007, S. 60–67
- Analía Francia, Alfredo E. Zurita und Alfredo A. Carlini: How Marine Isotope Stage 3 (MIS3) is reflected in northern Mesopotamia faunal assemblage of Argentina: The Xenarthra Cingulata case. Quaternary International 377, 2015, S. 126–139, doi:10.1016/j.quaint.2015.03.012
- Frédéric Delsuc, Mariella Superina, Marie-Ka Tilak, Emmanuel J. P. Douzery und Alexandre Hassanin: Molecular phylogenetics unveils the ancient evolutionary origins of the enigmatic fairy armadillos. Molecular Phylogenetics and Evolution 62, 2012, S. 673–680
- Marcos D. Ercoli, Alicia Álvarez, Carla Santamans, Sonia A. González Patagua, Juan Pablo Villalba Ulberich und Ornela E.Constantini: Los Alisos, a new fossiliferous locality for Guanaco Formation (late Miocene) in Jujuy (Argentina), and a first approach of its paleoecological and biochronology implications. Journal of South American Earth Sciences 93, 2019, S. 203–213
- Gustavo J. Scillato-Yané, Flávio Góis, Alfredo E. Zurita, Alfredo A. Carlini, Laureano R. González Ruiz, Cecilia M. Krmpotic, Cristian Oliva und Martín Zamorano: Los ingulata (Mammalia, Xenarthra) del Conglomerado osífero (Mioceno Tardío) de la Formación Ituzaingó de Entre Ríos, Argentina. In: Diego. Brandoni und J. I. Noriega (Hrsg.): El Neógeno de la Mesopotamia argentina. Asociación Paleontológica Argentina, Publicación Especial 14, 2013, S. 118–134
- Gerardo Carbot-Chanona: The First Record of Dasypus (Xenarthra: Cingulata: Dasipodidae) in the Late Pleistoceneof México. Current Research in the Pleistocene 27, 2010, S. 164–166
- H. Gregory McDonald: Paleoecology of extinct Xenarthans and the Great American Biotic Interchange. Bulletin of the Florida Museum of Natural History 45 (4), 2005, S. 313–333
- Édison V. Oliviera und Jamil C. Pereira: Intertropical Cingulates (Mammalia, Xenarthra) from the Quaternary of southern Brazil: Systematics and paleobiogeographical aspects. Revista Brasileira de Paleontologia 12 (3), 2009, S. 167–178
- Esteban Soibelzon, L. S. Avilla und M. Castro: The cingulates (Mammalia: Xenarthra) from the late Quaternary of northern Brazil: Fossil records, paleoclimates and displacements in America. Quaternary International 377, 2015, S. 118–125, doi:10.1016/j.quaint.2015.02.052
- Paulo V. Oliveira, Ana Maria Ribeiro, Édison V. Oliveira und Maria Somália S. Viana: The Dasypodidae (Mammalia, Xenarthra) from the Urso Fóssil Cave (Quaternary), Parque Nacional de Ubajara, State of Ceará, Brazil: paleoecological and taxonomic aspects. Anais da Academia Brasileira de Ciências 86 (1), 2014, S. 147–158
- Beth Shapiro, Russell W. Graham und Brandon Leets: A revised evolutionary history of armadillos (Dasypus) in North America based on ancient mitochondrial DNA. Boreas 2014, doi:10.1111/bor.12094
- Karen McBee und Robert J. Baker: Dasypus novemcinctus. Mammalian Species 162, 1982, S. 1–9