Thermoregulation
Unter Thermoregulation versteht man in der Biologie die mehr oder weniger große Unabhängigkeit der Körpertemperatur eines Organismus von der Außenwelt. Es lassen sich Thermokonforme (= poikilotherme, also wechselwarme Tiere) und Thermoregulatoren (= homoiotherme, also gleichwarme Tiere) unterscheiden.
Die Bezeichnungen Warmblüter und Kaltblüter werden inzwischen in diesem Zusammenhang nicht mehr verwendet und nur noch in Bezug auf Pferderassen angewandt, bei denen sich die Begriffe aber nicht auf die Körpertemperatur, sondern auf das Temperament beziehen.
Begriffserläuterung
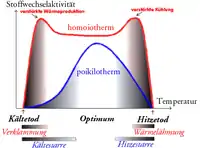
Es gibt zwei Möglichkeiten, eine Einteilung der Regulationstypen vorzunehmen. Die erste Einteilung beruht darauf, wie sich die Körpertemperatur im Vergleich zur Außentemperatur ändert:
- Als homoiotherm oder homöotherm bezeichnet man Tiere, die ihre Körpertemperatur auf einem konstant hohen, gleichwarmen Niveau halten und in engen Grenzen regulieren können. Hierzu gehören fast alle Säugetiere und alle Vögel.
- Als poikilotherm (vom griechischen: wechselnd) werden alle anderen Tiere bezeichnet, deren Körpertemperatur der Außentemperatur mehr oder weniger passiv folgt, also je nach Umgebungstemperatur wechselt. Hierzu gehören die niederen Wirbeltiere, nämlich die Fische, Amphibien und Reptilien, sowie die wirbellosen Tiere.
- Heterotherm sind Tiere, die ihre Körpertemperatur zwar um einige Grad, aber nur kurzfristig und auf bestimmte Körperabschnitte beschränkt variieren können, nicht jedoch in engen Grenzen. Sie werden noch in temporär Heterotherme und regional Heterotherme unterteilt. Der Ameisenigel (Echidna) und das Schnabeltier (beides Kloakentiere) sind temporär heterotherm, das heißt, ihre Körpertemperatur variiert zeitlich sehr stark. Zeitliche Variationen der Körpertemperatur findet man auch bei einigen wüstenbewohnenden Säugetieren und dem Asiatischen Elefanten[1] und auch bei vielen Insekten und bei der Python-Schlange. Bienen und Hummeln können durch Muskelzittern ihren Thorax auf Betriebstemperatur bringen. Sie besitzen einen Mechanismus (Gegenstromprinzip), der verhindert, dass die erzeugte Wärme ins Abdomen abfließt. Damit wären sie ein Beispiel für regional heterotherme Tiere.
Die zweite, aber problematische Einteilung fragt nach der Herkunft der Körperwärme, wobei
- endotherme Tiere ihre Wärme selbst produzieren und
- ektotherme Tiere ihre Körperwärme fast vollständig aus der Umgebung erhalten.

Diese Einteilung ist insofern sinnvoll, als es beispielsweise im Lebensraum eines poikilothermen Tiefseefischs so gut wie keine Temperaturschwankungen gibt. Daher bleibt auch seine Körpertemperatur innerhalb sehr enger Grenzen konstant, also sozusagen homoiotherm (gleichwarm). Manche poikilothermen Hochseefische, wie beispielsweise die Thunfische, können während der Bewegung durch ihre Muskelarbeit eine um 10–15 K höhere Temperatur als die Wassertemperatur haben, besitzen also nicht die gleiche Körpertemperatur wie ihre Umgebung. Im Gegensatz dazu verfällt ein Kolibri als homoiothermes Tier nachts in einen Torpor und hat eine 10–20 K niedrigere Temperatur als am Tag.
Allerdings sind endotherm und exotherm klar definierte Begriffe in der physikalischen Chemie und „ektotherm“ ist geradezu dazu prädestiniert, mit „exotherm“ verwechselt zu werden. Außerdem produzieren natürlich auch wechselwarme Tiere bei ihren Lebensprozessen selbst Wärme, allerdings verfügen sie nicht über die gleichen Mechanismen wie gleichwarme Tiere, um das ausreichend und konstant zu tun.
Trotz der geschilderten Probleme werden in der Biologie die Begriffe homoiotherm und endotherm beziehungsweise poikilotherm und ektotherm häufig gemengt und synonym verwendet. Der Grund hierfür dürfte in der großen Vielfalt der thermoregulatorischen Leistungen von Tieren liegen. Die Begriffe homoiotherm und poikilotherm beschreiben diese nur annäherungsweise. Die Zuordnung einer Tiergruppe ist insofern nicht einfach, da man häufig auf fließende Übergänge stößt.
| Thermokonformer | Thermoregulator | ||
|---|---|---|---|
| poikilotherm | homoiotherm | ||
| ektotherm | endotherm | ||
| (kaltblütig) | (warmblütig) | ||
| heterotherm | |||
Im Folgenden werden nur die Begriffe homoiotherm und poikilotherm verwendet.
Körperkern und Körperschale
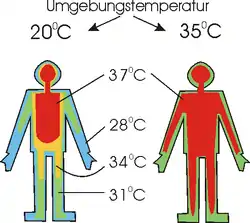
Wenn man allgemein von Körpertemperatur spricht, so nimmt man nur dann keine unzulässige Verallgemeinerung vor, wenn man von der Temperatur im Innern des Körpers spricht. Die Körpertemperatur gibt es nämlich nicht. Misst man an verschiedenen Stellen, ergeben sich unterschiedliche Werte. Misst man zu unterschiedlichen Zeiten, ergeben sich selbst für den Kern unterschiedliche Werte (siehe auch Chronobiologie).
Die Temperatur verschiedener Körperstellen, die nicht zum Kern gehören, hängt von der Umgebungstemperatur und der Muskelaktivität ab. Konstant gehalten wird die Temperatur bei Homoiothermen nur im Inneren des Körpers, weshalb sich ihre Körpertemperatur am genauesten über die rektale Temperaturmessung ermitteln lässt. Dem Körperkern steht die Körperschale entgegen.
Im Körperkern liegen die Organe mit hohem Energieumsatz (Herz, Leber, Niere und Gehirn), welche die Orte der Wärmebildung darstellen. Ihre Masse macht beim Menschen nur 8 % der Körpermasse aus, ihr Anteil am Energieumsatz eines Ruhenden beträgt aber mehr als 70 %. Haut und Muskulatur bilden dagegen 52 % der Körpermasse, liefern aber in Ruhe nur 18 % der gesamten Wärme. Bei Bewegung entsteht allerdings mehr Wärme in der Körperschale; dann übersteigt deren Anteil den des Kerns bei weitem.
Als Isothermen bezeichnet man Linien mit gleicher Temperatur. Die Körperschale ist also kein fest umrissenes Gebiet, sondern von der Umgebungstemperatur abhängig. Für die Verschiebung der Isothermen ist die wechselnde Durchblutung der einzelnen Körperpartien verantwortlich. Beim Menschen ist beispielsweise die Durchblutung der Finger sehr variabel, sie kann um den Faktor 600 schwanken. Werden die Finger bei einer tiefen Umgebungstemperatur nur schwach durchblutet, ist die Temperaturdifferenz zwischen ihnen und ihrer Umgebung nicht mehr so groß, und sie verlieren weniger Wärme. Wechselnde Durchblutung der Körperschale ist eine wichtige temperaturregulatorische Maßnahme, die von allen Arten eingesetzt wird und zwar in beide Richtungen – gegen Unterkühlung und Überhitzung. Auch die Körpertemperatur eines Schlittenhundes zeigt nicht überall die gleichen Werte. Erreicht werden die Unterschiede durch allerlei Maßnahmen: Durchblutungsveränderung, Gegenstromprinzip, Fettablagerungen, isolierendes Fell.
Sinn der Thermoregulation
Bei poikilothermen Tieren ist die Metabolismusrate ebenso temperaturabhängig wie die Reaktionsgeschwindigkeit in biochemischen Systemen (Van-’t-Hoff’sche Regel), während sie bei homoiothermen Tieren mit steigender Außentemperatur bis zu einem kritischen Punkt steigt (ab diesem Punkt wird sie unabhängig). Sinkt die Außentemperatur unter einen kritischen Punkt, muss ein homoiothermes Tier seinen Stoffwechsel steigern. Die Aufrechterhaltung einer konstanten Körpertemperatur ist aber aufwendig: Homoiotherme brauchen für die gleiche „Arbeitseinheit“ mehr Nahrung als Poikilotherme.
Dem erhöhten Aufwand zur Aufrechterhaltung einer konstanten Körpertemperatur steht jedoch ein Nutzen gegenüber, denn enzymatische Stoffwechselreaktionen sind stark temperaturabhängig. Während poikilotherme Tierarten zum Leben bestimmte Außentemperaturbereiche benötigen, sind homoiothermen Tiere wie Säuger und Vögel durch ihre Fähigkeit zur Temperaturregulation weit weniger davon abhängig und in der Lage, auch in gemäßigteren Breiten oder sogar subpolaren und polaren Zonen leben und Stoffwechselreaktionen ablaufen lassen zu können. Sie benötigen zwar eine größere Nahrungsmenge als poikilotherme Tiere, dafür stehen ihnen aber mehr Möglichkeiten zur Verfügung – ihre ökologische Potenz ist größer.
Die Körpertemperatur eines Homoiothermen ist als Regelkreislauf zu verstehen. Das heißt, es existiert ein Sollwert, der mit einem Istwert ständig verglichen wird und bei dem Stellglieder existieren, die den Istwert an den Sollwert angleichen, falls dieser abweicht.
Da die optimale Temperatur zur Produktion von Spermien beim Menschen drei oder vier Grad niedriger liegt als diejenige des Körperkerns, ist auch für den menschlichen Hodensack das Prinzip der Thermoregulation sowie dasjenige der Gegenstromkühlung kennzeichnend. Die entsprechende Wärmeregulierung wird durch die Tunica dartos in Zusammenarbeit mit dem Musculus cremaster gesteuert, der den bei höherer Temperatur vergleichsweise tief hängenden Hoden nur bei Kälte an den Körper heranzieht. Gleichzeitig wird bei Kälte die Haut am Hoden durch die Tunica dartos stark kontrahiert (runzliges Aussehen), was die Oberfläche verkleinert und die Wärmeabstrahlung reduziert.
Regulation des Temperatursollwerts bei homoiothermen Säugern
Nicht eine einzelne Region des Nervensystems fungiert als alleiniges Thermoregulationszentrum, sondern hierarchisch aufeinander aufgebaute Strukturen: Schon das Rückenmark und der Hirnstamm können grob Änderungen in der Körpertemperatur wahrnehmen und Regulationen veranlassen. Dies zeigt sich, wenn Nervenbahnen von weiter oben unterbrochen sind und dennoch eine grobe Temperaturregelung stattfindet. Jedoch erst wenn diese tiefer gelegenen Bereiche des zentralen Nervensystems mit der Regio praeoptica des Hypothalamus verbunden sind, wird die Thermoregulation präzise, insbesondere auch bei Änderungen der Umgebungstemperatur und bei körperlicher Anstrengung. Dort wird die Ist-Körperkerntemperatur präziser direkt wahrgenommen, ferner laufen dort afferente Signale z. B. von Wärme- und Kälterezeptoren aus der Haut des ganzen Körpers zusammen.

Die Temperaturinformationen aus der Peripherie werden mit den zentralen Temperaturinformationen verglichen und integriert; es resultiert eine von hier letztlich gesteuerte passende thermoregulatorische Antwort in Richtung Wärmeverlust (periphere Gefäßerweiterung und Hauterwärmung, Schwitzen, beim Hund hecheln etc.) oder in Richtung Wärmeproduktion und -einsparung (periphere Gefäßverengung und Hautkälte, Kältezittern etc.). Normale pyrogeninduzierte Fieberreaktionen sind ebenfalls nur mit einer intakten Regio praeoptica des Hypothalamus möglich.[2]
In der Regio praeoptica des Hypothalamus finden sich verschiedene Neurone: Ca. 30 % sind wärmesensitiv (das heißt, sie feuern schneller, wenn die lokale Wärme steigt), über 60 % reagieren nicht auf Temperaturänderungen und weniger als 5 % sind kältesensitiv. Es wurde vermutet, dass der Temperatursollwert durch einen Vergleich der Neuronenaktivität der temperaturinsensitiven Neurone mit den wärmesensitiven Neuronen entstehe. Insbesondere die Aktivität der kältesensitiven Neurone ist stark abhängig von excitatorischen und inhibitorischem Input benachbarter Neurone, während die wärmesensitiven Neurone vor allem Input aus der Peripherie bekommen.[2]
Stellglieder bei Homoiothermen

Zu den regel-theoretischen Fachbegriffen siehe Regelkreis#Biologische Regelkreise.
Gegen Unterkühlung
Eine Unterkühlung und im Extremfall Erfrierung wird neben der Lufttemperatur vor allem durch Faktoren wie den Niederschlag (Wirkung von Nässe) und den Wind (siehe Windchill) bedingt.
Durchblutung
Eine Durchblutungsänderung der Haut bedeutet für den Körper keinen nennenswerten Energieaufwand. Die Vasomotorik bewirkt eine Verengung (Vasokonstriktion) oder Erweiterung (Vasodilatation) der Blutgefäße. Die Steuerung kann auf unterschiedliche Arten erfolgen. Ohne Beteiligung des Nervensystems erfolgt die Reaktion der glatten Muskulatur auf örtliche Erwärmung oder Abkühlung. Durch Kontraktion oder Entspannung der Muskeln wird die örtliche Durchblutung verändert. Unter Beteiligung eines Rückenmarkreflexes können Durchblutungsreaktionen ausgelöst werden. Diese nennt man nervös. Körperweite Reaktionen bleiben hier ebenso aus wie bei der lokalen Reaktion der glatten Muskulatur. Eine weitere Möglichkeit ist die Umstellung des Kreislaufs als Folge von Befehlen des Hypothalamus. Dieser steht mit Kälte- und Wärmerezeptoren der Haut und mit bluttemperaturregulierenden Zentren in Kontakt.
Gefäßerweiterung bei Hitze – Steuerung durch die Außentemperatur:
- Bei Hitze werden aus Plasmaglobulinen durch eine in den Schweißdrüsen gebildete Protease Kinine, vor allem Bradykinin, abgespalten. Eine erhöhte Kinin-Konzentration führt dazu, dass die Muskeln der Blutgefäße der Haut weniger stark auf die gefäßverengende Wirkung des Sympathikus reagieren; sie erschlaffen und die Blutgefäße werden erweitert.
- Axonreflex-Hypothese: Die Nervenfortsätze der Hitzerezeptoren der Haut, die über den Spinalnerv ins Rückenmark ziehen, haben Abzweigungen, sogenannte Kollaterale, die direkt, also ohne zwischengeschaltete Synapsen, mit den Muskeln der Blutgefäße verbunden sind und bewirken, dass bei Hitze diese Muskeln erschlaffen.
Kältezittern
Wie bei der normalen Muskelarbeit entsteht auch beim Zittern Wärme. Beim Zittern wird sie zur Wärmeproduktion verwendet. Auch Poikilotherme wie beispielsweise Bienen, Wespen, Hummeln oder Nachtfalter können sich durch Muskelzittern aufwärmen. Allerdings ist die Netto-Ausbeute gering, falls der Körper schlecht isoliert ist: Da die Muskulatur mehr durchblutet werden muss, um arbeiten zu können, geht bei Zittern viel Wärme verloren. Die Kerntemperatur steigt erst, wenn die Muskeln warm geworden sind, siehe auch Afterdrop. Für Schiffbrüchige im Wasser gilt: Möglichst nicht bewegen.
Braunes Fettgewebe – zitterfreie Wärmeproduktion
Die Zellen des braunen Fettgewebes sind in der Lage, durch Oxidation von Fettsäuren Wärme zu erzeugen (Thermogenese). Dazu entkoppelt das in den Mitochondrien vorhandene Membranprotein Thermogenin die Fettsäureoxidation von der Synthese des Energieträgers Adenosintriphosphat (ATP), so dass die freigesetzte Energie in Wärme umgesetzt wird. Braunes Fettgewebe findet sich insbesondere bei neugeborenen Säugetieren.
Akklimatisation
Bei der Akklimatisation finden physiologische Änderungen statt, wenn sich eine langfristige Veränderung des Milieus einstellt. Dies geschieht durch eine enzymatische Reaktion, eine Änderung der Molekularstruktur oder andere Faktoren, die das Verhalten beeinflussen.
Winterschlaf und Winterruhe
Um eines von beiden tun zu können, ist ein System zur Temperaturregulation nötig. Daher kommen Winterschlaf und Winterruhe nur bei Homoiothermen vor – und zwar fast ausschließlich bei Säugetieren. Bei den Vögeln gibt es ebenfalls noch ein paar Arten, die Winterschlaf halten. Manche Vogelarten zeigen einen kurz andauernden Zustand, der dem Winterschlaf in vielen Zügen gleicht. Dieser Zustand wird Torpor genannt und tritt beispielsweise bei den Kolibris auf. Er hilft ihnen, die Nacht zu überstehen.
Bei Poikilothermen nennt man den Zustand, in dem sie sich im Winter in höheren Breiten befinden, Kälte- oder Winterstarre. Diese ist auf die sinkende Umgebungstemperatur zurückzuführen, die ihre Körpertemperatur und damit ihre Stoffwechselrate ebenfalls sinken lässt. Manche Poikilothermen haben die Fähigkeit entwickelt, auch Temperaturen unter dem Gefrierpunkt zu überstehen. Der Mehlkäfer kann beispielsweise −17 °C überstehen. Arten aus tropischen Gebieten können das nicht. So stirbt ein Alligator schon bei Temperaturen knapp über 0 °C.
Winterschlaf

Entgegen der landläufigen Meinung halten Bären im Winter keinen Winterschlaf. Echte Winterschläfer finden sich nur in fünf Ordnungen: bei den Insektenfressern (Igel), bei einigen Nagetieren (z. B. Hamster, Murmeltier), bei einem Primaten (Fettschwanzmaki), bei den Fledertieren (Fledermäusen) und den Schildkröten.
Beim Winterschlaf sinkt die Atem- und Herzschlagfrequenz, der Gesamtenergieumsatz wird reduziert und das Tier verfällt in einen stärkeren Starrezustand als im Schlaf. Es wird nicht geträumt. Man weiß, dass Hamster und Ziesel ab und zu aufwachen, um – es hört sich vielleicht merkwürdig an – zu schlafen. Winterschlaf hat nämlich rein physiologisch nichts mit dem normalen Schlaf zu tun.
Welche Faktoren zu einem Winterschlaf führen, ist noch unklar. Eventuell ist ein gewisser Prozentanteil Fett nötig und/oder die Abnahme der Tageslänge, Umgebungstemperatur und anderes. Mit dem Phänomen des Timing beschäftigt sich die Chronobiologie. Ein Winterschläfer muss Maßnahmen gegen das Erfrieren treffen. Entweder er wacht ab und zu auf und läuft umher oder er hält seine Temperatur ständig auf einem Wert kurz über dem Gefrierpunkt. So oder so – auch während des Winterschlafes muss die Körpertemperatur kontrolliert werden. Daher kann man davon ausgehen, dass der Sollwert herabgesetzt ist und nur bei Unterschreitung Maßnahmen eingeleitet werden.
Winterruhe
Bei der Winterruhe wird der Stoffwechsel bei weitem nicht so drastisch reduziert wie beim Winterschlaf. Bei Bären etwa sinkt zwar die Herzfrequenz, aber sie erreicht jeden Tag etwa für 30 Minuten einen höheren Wert. Außerdem bringen viele Arten während der Winterruhe ihre Jungen zur Welt, was auf jeden Fall ein geregeltes endokrines System erfordert. Winterruher bewegen sich, während sie ruhen. Dennoch kann eine Winterruhe lange dauern. Die Tiere zehren in dieser Zeit von ihrem weißen Fettgewebe oder betreiben Vorratshaltung.
Wanderung
Bei Tierwanderungen, wozu auch der Vogelzug zählt, handelt es sich um Verhalten. Aber auch Verhalten kann zur Temperaturregulation eingesetzt werden. Die meisten Tierwanderungen geschehen allerdings nicht wegen der schlechten Witterung, sondern wegen der damit verbundenen Nahrungsmittelknappheit. Beispielsweise ist es in manchen Gebieten in New South Wales in Australien im (dortigen) Winter recht kalt – nachts etliche Grad unter Null. Da zu dieser Zeit aber heimische Bäume und Sträucher blühen, verweilt der Honeyeater (Honigfresser), eine Vogelart, dort so lange, bis es keine Nahrung mehr gibt, und zieht dann erst weiter.
Haare und Fell

Ein Fell besteht aus Haaren und ist neben den Milchdrüsen ein Schlüsselmerkmal der Säugetiere. Haare sind den Reptilienschuppen oder den Vogelfedern homologe Bildungen der Haut.
Für alle nicht in den Tropen lebenden Arten ist der Haarwechsel im Allgemeinen eine temperaturregulatorische Maßnahme: Viele Tiere legen sich in unterschiedlichen Jahreszeiten ein Fell mit anderen wärmedämmenden Eigenschaften als das vorige zu. Ein Winterfell hat in der Regel längere und auch dichter stehende Haare als ein Sommerfell und kann in der kalten Jahreszeit eine dickere ruhende Luftschicht festhalten als im Sommer. Durch Aufrichten der Haare mit Hilfe von Haarmuskeln kann die ruhende Luftschicht noch vergrößert werden. Außerdem sorgen die im Haarmark eingeschlossenen Luftmassen für eine thermische Isolation. Im Wasser lebende Arten nutzen den Effekt, dass Luft ein wesentlich schlechterer Wärmeleiter ist als Wasser. Viele Arten machen ihre Haare wasserabstoßend, indem sie sie mit einem öligen Drüsensekret einreiben. Tropische Arten wechseln oft in unauffälliger Weise einzelne Haare.
Gegen Überhitzung
Die Spitzenwerte der oberen kritischen Temperatur liegen bei allen Arten relativ dicht beieinander, da starke Überhitzung des Körpers zu Schäden am zentralen Nervensystem führt. Bei der unteren kritischen Temperatur lassen sich dagegen große Unterschiede feststellen.
Beim Menschen liegt die Schwitzgrenze knapp oberhalb von 30 °C. Steigt die Umgebungstemperatur weiter, so kann der Organismus dies so lange ertragen, wie die Maßnahmen gegen Überhitzung ausreichen. Ab einer bestimmten Temperatur ist dies nicht mehr möglich und die Überhitzungsgrenze ist erreicht. Sie ist von der Luftfeuchtigkeit abhängig und wird für den Menschen durch den Hitzeindex quantitativ beschrieben. Bei 30 % relativer Luftfeuchtigkeit kann ein Mensch 50 °C über Stunden ertragen. Steigt die Luftfeuchtigkeit auf 70 %, liegt die erträgliche Höchsttemperatur nur noch bei 40 °C.
Wird der Körper überhitzt, kommt es zum Hitzschlag, sowohl beim Menschen als auch bei Tieren. Dabei sind die Blutgefäße maximal erweitert; Die Menge des Blutes reicht nicht mehr, um sie zu füllen.
Verdunstungskälte

Schwitzen ist eine Maßnahme, die fast ausschließlich Primaten zur Verfügung steht. Durch das Verdunsten des Schweißes auf der Haut entsteht Verdunstungskälte; diese Art der Wärmeabgabe wird als Evaporation bezeichnet. Nachteil des Schwitzens ist der hohe Wasserverlust. Auch Pferde schwitzen, allerdings anders als Primaten (ekkrine Drüsen) aus apokrinen Drüsen deren Sekret anders zusammengesetzt ist. Insbesondere enthält Pferdeschweiß das Protein BPIFA4P (Latherin), das einen Schaum auf dem Fell bildet.[3]
Auch andere Tiere nutzen Verdunstungskälte. Speichel oder Nasendrüsensekret (siehe beispielsweise Rete mirabile). Flughunde und Riesenkängurus speicheln mit der Zunge über den Körper. Flughunde fächeln dann mit den Flügeln, wodurch die Verdunstung beschleunigt wird. Der asiatische Elefant kann mit dem Rüssel Speichel aus den Backentaschen holen und ihn über den Körper verteilen. Der afrikanische Elefant, der diese Möglichkeit nicht hat, besitzt dafür große Ohren.
Beim Hecheln des Hundes handelt es sich um Verdunstung von Nasendrüsensekret. Es wird Luft durch die Nase eingesogen und durch den Mund wieder abgeatmet. Dieses Verhalten ist auch bei Katzen, Schafen und Antilopen zu beobachten. Hecheln entzieht dem Körper im Gegensatz zum Schwitzen kein Salz. Allerdings besteht die Gefahr der Alkalose (der pH-Wert des Blutes steigt, da zu viel Kohlendioxid abgeatmet wird). Die Schleimhaut in den Nasenmuscheln und der Mundhöhle ist von einem dichten Netz von Arterien und Venen durchzogen, und durch ihre vielen Faltungen besitzt sie eine riesige Oberfläche. Beim Hund ist sie beispielsweise größer als die Körperoberfläche.
Thermische Fenster

Thermische Fenster sind Bereiche mit nur dünnem Fellbewuchs. An diesen Stellen ist die Isolation weniger gut. So hat der Hund beispielsweise thermische Fenster zwischen den Vorderbeinen, am Brustkorb und in der Lendengegend. Je nach Körperhaltung sind diese Fenster geöffnet oder geschlossen.
Auch Robben besitzen diese thermischen Fenster. Solange sie sich im Wasser befinden, droht ihnen die Gefahr der Überhitzung kaum. Dann müssen sie sich eher gegen Unterkühlung schützen. Eine isolierende Fettschicht ist bei allen im Wasser lebenden, homoiothermen Arten zu finden. Suchen sie das Land auf, wie Robben dies zur Brunftzeit tun, müssen sie ihre thermischen Fenster öffnen. Bei ihnen geschieht das durch unterschiedliche lokale Hautdurchblutung.
Verhalten

Verhalten kann ebenfalls gegen Überhitzung helfen. Beispielsweise können Tiere Schattenzonen aufsuchen, nachts aktiv sein oder einen „Sonnenschirm“ mit sich führen, wie es das Afrikanische Borstenhörnchen (Xerus inauris) tut, das seinen Schattenspender in Form seines buschigen Schwanzes immer dabei hat.
Beim sogenannten Splooting legen sich einige Säugetierarten bei großer Hitze flach auf den Boden, um so Körperwärme an den Boden abgeben zu können.[4][5]
Die tropischen Seidenspinnen (Gattung Nephila) richten sich bei starker Sonneneinstrahlung so zur Sonne aus, dass nur der schmale Hinterleib direkt bestrahlt wird. Bei stärker ansteigender Körpertemperatur ziehen sie sich in den Schatten eines selbstgewebten Sonnenschutzes in der Mitte ihres Radnetzes zurück. Bei morgendlicher Kälte der Tropen richtet sich die Spinne im 90-Grad-Winkel zur Sonne aus, so dass sie ihre Körpertemperatur um bis zu 7 K erhöhen kann. Viele Wüstentiere wie die Schwarzkäfer graben sich tagsüber im Sand ein, um sich so vor der Sonnenstrahlung zu schützen.
Stellgliederprinzipien
Das Gegenstromprinzip

Das Gegenstromprinzip ist allgemein ein Verfahren zum Wärme- oder Stoffaustausch zwischen zwei Flüssigkeiten oder Gasen. Hier im Besonderen wird das Gegenstromprinzip zur Energieersparnis eingesetzt. (Man vergleiche auch das Gegenstromprinzip in der Niere.) Für den Rücktransport des Blutes ins Innere des Körpers stehen zwei Wege zur Verfügung. Der erste Weg führt über die an der Oberfläche liegenden Hautvenen, der zweite über die tief neben den Arterien liegenden Venen.
In kalter Umgebung fließt nur sehr wenig Blut durch die oberflächlichen Gefäße; das geringe Volumen genügt aber, um Stoffwechselprozesse zu ermöglichen. Das meiste Blut fließt durch die tiefen Venen und nimmt dabei Wärme von den Arterien auf, so dass es schon vorgewärmt ins Körperinnere gelangt. Es muss also nicht so viel Energie aufgewandt werden, um es auf die Kerntemperatur zu erwärmen. Gleichzeitig wird das arterielle Blut infolge des Temperaturgefälles abgekühlt und erreicht das Körperende schon weitgehend auf Außentemperaturniveau. Das System arbeitet demnach höchst effektiv: Eine Möwe, die versuchsweise für zwei Stunden in Eiswasser gestellt wurde, verlor nur 1,5 Prozent ihrer metabolischen Wärmeproduktion über die Füße.[6] In der Grafik sieht man die vom Körper kommende Arterie mit warmem Blut auf dem Weg in den Vogelfuß in kalter Umgebung.
Bei warmer Umgebung nimmt das Blut den Weg über die oberflächlichen Hautvenen und gibt zusätzlich Wärme ab. Dieses Prinzip ermöglicht Walen und Robben in kühler Umgebung Wärmeverluste zu verringern. Bei ihnen ist eine in die Peripherie führende Arterie völlig von mehreren Venen umgeben. Es ermöglicht ihnen beispielsweise, auf dem Eis zu liegen, ohne einzuschmelzen. Schlittenhunde können in ihrem Körper sehr verschiedene Temperaturen haben. Da die Pfoten kühl sind, wird kaum Wärme über sie abgegeben.
Wundernetze – Retia mirabilia
Wundernetze (Retia mirabilia) sind eine besondere, im Rahmen der Thermoregulation vor allem bei Katzen und Paarhufern vorkommende Form des Gegenstromprinzips. Hier wird das Gegenstromprinzip nicht, wie meist, als Maßnahme gegen Unterkühlung, sondern als Kühlsystem in der Kopfregion eingesetzt. Beuteltieren, Primaten, Nagetieren, Hasenartigen und Unpaarhufern fehlt ein solches Rete mirabile.
Ein Beispiel: Wird der in der Mojave-Wüste lebende Eselhase an einem 41 °C heißen Tag nur 10 Minuten lang gejagt, steigt seine Körpertemperatur schnell von 41 auf 43 °C, knapp unterhalb der tödlichen 44 °C. Das Gehirn, ein gegen Überhitzung besonders empfindliches Organ, wird als erstes geschädigt. Ist der Jäger des Hasen ein Hund, so bleibt dessen Gehirn kühl, da ein Hund über ein Wundernetz verfügt. Bei ihm verästelt sich die Halsarterie und liegt eingebettet im Sinus cavernosus – einem Sammelbecken für venöses Blut. In diesem kühlt das arterielle Blut ab, bevor es ins Gehirn weiter fließt. Die Temperaturdifferenz kann bis zu 3 °C betragen.
Das Blut im Sinus cavernosus ist verhältnismäßig kühler, da es aus dem Nasen-Mundbereich des Tieres kommt und dort in den reich durchbluteten Nasenmuscheln durch Verdunstung gekühlt wurde (Hecheln). Bei Ruhe wird weniger gehechelt also auch weniger gekühlt als bei Anstrengung. Arten ohne diesen Mechanismus der Blutkühlung müssen ihren ganzen Körper auf einer Temperatur halten, die dem Gehirn zuträglich ist. Geschieht die Kühlung durch Schwitzen, bedeutet das einen großen Wasserverlust.
Körpergröße eines Homoiothermen
Kleine Tiere haben verhältnismäßig große Oberflächen, daher verlieren sie viel Wärme (Oberfläche-zu-Volumen-Verhältnis).[7] Entsprechend viel Energie müssen sie auch wieder zuführen. So vertilgt eine Spitzmaus beispielsweise täglich ihr eigenes Körpergewicht in Form von Insekten und Würmern. Sie muss fast ständig auf der Nahrungssuche unterwegs sein, um dies zu bewältigen. Kolibris können nur tagsüber Nahrung sammeln. Sie müssen, um die Nacht zu überstehen, in einen Torpor verfallen, bei dem sie ihre Körpertemperatur drastisch senken. Der Größe von Homoiothermen nach unten sind also Grenzen gesetzt.
Eine Kugel hat bei gleichem Volumen von allen geometrischen Körpern die kleinste Oberfläche. Im Vergleich der Kugelgeraden mit der Maus-Elefant-Gerade kommt man auf einen Wert von 1,12 Gramm als minimale Körpermasse. Allerdings gilt dieser Wert für einen perfekt runden Körper. Die Schweinsnasenfledermaus als kleinstes Säugetier und die Bienenelfe (oft als Hummelkolibri bezeichnet) als kleinster Vogel weichen mit 1,5 Gramm beziehungsweise 1,6 Gramm nur geringfügig von dem Idealwert ab.
Poikilotherme Tiere
Bei den meisten poikilothermen Tieren ist die Wärmeabgabe im Verhältnis zur Wärmeproduktion so groß, dass sie praktisch die Temperatur der Umgebung annehmen. Dies gilt insbesondere für im Wasser lebende Tiere: Da Wasser weniger als ein Volumenprozent Sauerstoff enthält (Luft: 21 Volumenprozent) müssen die Wassertiere eine sehr große Menge des umgebenden Mediums pro Zeitspanne über ihre respiratorischen Austauschflächen (Kiemen, Haut) strömen lassen, um ihren Sauerstoffbedarf aus dem Wasser zu decken. Dadurch wirken die respiratorischen Flächen zugleich als Wärmeübertrager mit der Umgebung und führen bis zu 60 % der durch Stoffwechseltätigkeit gebildeten Wärme des Körpers ab.
Bei poikilothermen Landtieren konnte nachgewiesen werden, dass sich die Betriebstemperatur artspezifisch unterscheidet. Ihre Muskel-ATPase zeigt je nach Habitat der Tiere höhere oder niedrigere Aktivitätsoptima. Bietet man den Tieren (beispielsweise verschiedenen Arten von Wüsteneidechsen) im Labor einen Temperaturgradienten, suchen sie artspezifisch ihre Vorzugstemperatur auf. Man kann bei poikilothermen Tieren also häufig von verhaltensgesteuerter Thermoregulation sprechen.
Bei Insekten kommt es durch den intensiven Stoffwechsel in den Flugmuskeln während des Fliegens zu einem Wärmestau, der bei großen Insekten nicht schnell genug abgeführt werden kann. Die Thorax-Temperatur kann während des Fluges bis auf 45 °C ansteigen. Die Betriebstemperatur zeigt bei vielen dieser größeren Insekten (Schmetterlingen, Hautflüglern, Heuschrecken, Käfern) einen relativ hohen Wert (25–40 °C). Daher können sie erst dann starten, wenn diese Temperatur erreicht ist. Sie müssen ihren Flug unterbrechen, wenn der artspezifische Temperaturbereich über- oder unterschritten wird. In der Ruhephase entspricht ihre Körpertemperatur in der Regel derjenigen der Umgebung.

Dies bedeutet aber, dass sie vor Flugbeginn ihre Muskeln aufwärmen müssen. Das geschieht durch bestimmte Verhaltensweisen. Entweder sie nehmen eine günstige Haltung zur Sonne ein und wärmen sich durch die Absorption von Wärmestrahlen auf oder sie erzeugen ihre Betriebstemperatur selbst durch Muskelzittern, falls ihnen diese Möglichkeit zur Verfügung steht. Eine Anzahl von Großinsekten kann durch ihre antagonistisch arbeitenden Flugmuskeln, die sie synchron aktivieren, Wärme erzeugen. So beispielsweise auch der nebenstehende Maikäfer. Man spricht dann auch von physiologischer Thermoregulation.
Hier werden Ansätze einer autonomen Regulation der Körpertemperatur deutlich.
Reptilien zeigen neben Verhaltensweisen ebenfalls Ansätze zur autonomen Wärmeregulation. So können sie durch Wärmehecheln die respiratorische Verdunstung erhöhen oder durch Veränderung der Hautdurchblutung den Wärmeaustausch mit der Umgebung beeinflussen.
Poikilotherme in kalter Umgebung
Solange die meisten marinen wirbellosen Tiere unter Wasser existieren, bleiben sie über dem Gefrierpunkt ihrer Körperflüssigkeiten. Einige können aber Temperaturen unter dem Gefrierpunkt überleben. Normalerweise ist die Bildung von Eiskristallen in Zellen tödlich, weil sie bei ihrem Wachstum die Gewebe zerreißen und zerstören. Einige Tiere (beispielsweise Käfer) besitzen in ihrer extrazellulären Flüssigkeit Substanzen, welche die Kristallbildung beschleunigen. Daher gefriert diese Flüssigkeit, die die Zellen von außen umspült, schneller. Durch das Gefrieren nimmt die Flüssigkeit ab und wird konzentrierter, was wiederum Wasser aus den Zellen herauszieht und den intrazellulären Gefrierpunkt erniedrigt. Senkt sich die Temperatur weiter ab, setzt sich auch dieser Vorgang fort. Da Eiskristalle in der extrazellulären Flüssigkeit keinen Schaden anrichten können, kann beispielsweise eine Süßwasserlarve der Mücke Chironomus mehrfaches Einfrieren bei −32 °C überleben. Trotzdem enthalten ihre Zellen noch freies Wasser. Ein Überleben des Tieres bei der gesamten Gefrierung des Zellwassers ist nicht bekannt.
Einige Tiere können sich superkühlen, wobei Flüssigkeiten unter den Gefrierpunkt abgekühlt werden können und trotzdem nicht gefrieren. Es dienen beispielsweise Glykoproteine als Frostschutzmittel. Obwohl dieses Glycoprotein isoliert und seine chemische Struktur aufgeklärt wurde, ist der Mechanismus, durch den die Kristallbildung verzögert wird, nicht bekannt (Stand 1990).
Soziale Thermoregulation
Poikilotherme Tiere sorgen für eine konstante Nesttemperatur, wofür ein isoliertes Nest nötig ist.
- Die Rote Waldameise nutzt die Sonne als Wärmequelle. Spezialisierte Wärmeüberträgerinnen heizen ihren Körper in der Sonne auf. In den kühleren Brutkammern geben sie die gespeicherte Wärme wieder ab. Wird das Nest durch Sonneneinstrahlung zu heiß, werden Gänge auf der Schattenseite geöffnet, so dass ein kühlender Luftzug entsteht.
- Die Termite Macrocystis bellicosus unterhält Pilzkolonien, die sie auf einem Nahrungsbrei zieht. Dieser wird durch Bakterien fermentiert, wobei Wärme frei wird.
- Bei staatenbildenden Bienen, Wespen, Hummeln und Hornissen wird Wärme durch den Stoffwechsel der Individuen produziert, siehe auch Westliche Honigbiene#Thermoregulation der Honigbiene
- Eine Hummelkönigin kann ihre Brut an kühlen Tagen und Nächten durch ständiges Muskelzittern warm halten.
- Termiten in heißen Regionen erreichen ein gleichmäßiges Klima im Stock durch einen komplizierten Stockaufbau (beispielsweise Ausrichtung in Nord-Süd-Richtung bei Kompasstermiten und/oder Nutzung von Kamineffekten im Stock).
Weitere Beispiele zur Thermoregulation bei Tieren
Thermometerhuhn
Das Thermometerhuhn (Leipoa ocellata) gehört zu den Großfußhühnern (Megapodiidae) und lebt in trockenen Buschgebieten im südlichen Australien. Diese Tiere brüten ihre Eier auf eine besondere Art und Weise aus. Sie schichten große Haufen Blätter und anderes kompostierbares Material zu bis zu 150 cm hohen Haufen auf und legen ihre Eier darin ab. Indem sie ihren langen Schnabel in die verrottende und damit Wärme erzeugende Masse stecken, prüfen sie die Temperatur im Innern und nehmen, je nach Bedarf, Laub weg oder schichten neues dazu. Diese Arbeit wird von den Männchen ausgeführt. Sie halten die Temperatur bei 33 °C.
Schwertfisch
Der Schwertfisch Xiphias gladius weist während der Jagd eine im Vergleich zur Umgebung bis zu 15 °C wärmere Netzhaut auf. Die Erwärmung auf 19 bis 28 °C erfolgt durch einen Muskel hinter dem Auge. Dadurch können die Nervenzellen der Netzhaut schneller arbeiten, und der Schwertfisch kann seine Beute siebenmal schneller ausmachen als die Beute ihren Fressfeind.
Siehe auch
Literatur
- H. Precht, J. Christophersen, H. Hensel: Temperatur und Leben. Springer Verlag, 1955.
Weblinks
- Wiener Zeitung: Diagnose durch Körperwärme (Regulationsthermographie) (Memento vom 1. November 2005 im Internet Archive)
- Archiv Link: staff.uni-marburg.de (Memento vom 11. Juni 2007 im Internet Archive) HM Sommer: Thermoregulation
Einzelnachweise
- Nicole M. Wiesenböck et al.: Taking the heat: thermoregulation in Asian elephants under different climatic conditions. In: J. Comp. Physiol. B 182 (2012), S. 311–319. doi:10.1007/s00360-011-0609-8.
- J. A. Boulant: Role of the preoptic-anterior hypothalamus in thermoregulation and fever. In: Clin Infect Dis 31, 2000, S. 157–161, PMID 11113018.
- Rhona E. McDonald, Rachel Fleming, John G. Beeley, DL Bovell, Jian R. Lu, Xiubo Zhao, Alan Cooper, Malcolm W. Kennedy: Latherin: A Surfactant Protein of Horse Sweat and Saliva. In: PLOS ONE. 2009, Band 4, Nummer 5, S. e5726 doi:10.1371/journal.pone.0005726.
- Christian Weber: Warum Eichhörnchen flach liegen. In: sueddeutsche.de. 6. Juli 2023, abgerufen am 30. Juli 2023.
- Warum New Yorks Eichhörnchen aktuell „splooten“. In: Focus Online. 13. August 2022, abgerufen am 30. Juli 2023.
- Wieso erfrieren Vögeln im Winter nicht die Füße? Spektrum.de vom 21. September 2007
- Hans Joachim Schlichting, Bernd Rodewald: Von großen und kleinen Tieren. Praxis der Naturwissenschaften - Physik 37/5, 2 (1988)