Hardy-Weinberg-Gleichgewicht
Das Hardy-Weinberg-Gleichgewicht (HWG) (nach dem Mathematiker G. H. Hardy und dem Arzt und Vererbungsforscher Wilhelm Weinberg) ist ein Begriff der Populationsgenetik.
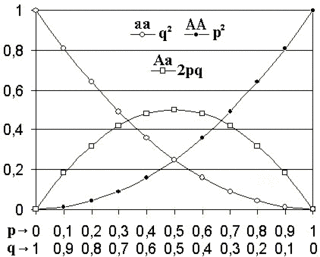
Zur Berechnung dieses mathematischen Modells geht man von einer in der Realität nicht vorzufindenden idealen Population aus. Dies bedeutet, dass keine Evolution stattfindet, da keine Evolutionsfaktoren greifen, die den Genpool verändern könnten. Für diesen Fall ergibt sich für jede beliebige Genotypverteilung der Elterngeneration eine nur von den Allelfrequenzen abhängige Genotypverteilung der ersten Tochtergeneration, die sich in den folgenden Generationen nicht mehr ändert. Mathematisch handelt es sich bei diesem sogenannten Gleichgewicht um einen Fixpunkt der durch den Vererbungsmechanismus definierten Funktion.
Das Hardy-Weinberg-Gleichgewicht wird trotz seines modellhaften Charakters zum Ableiten von populationsgenetischen Gesichtspunkten vom Modell auf die Realität verwendet. Insbesondere bei relativ großen Populationen lässt sich dieses Modell realistisch anwenden. Ferner findet die Regel Anwendung zur Berechnung des Anteils von heterozygoten Individuen (hier im Beispiel: Aa) bei dominant-rezessiven Erbgängen, da heterozygote Organismen von homozygot dominanten (hier: AA) phänotypisch nicht zu unterscheiden sind, weil sich das dominante Allel durchsetzt.
Geschichte
Der Beitrag G. H. Hardys

Die Mendelschen Gesetze wurden im Jahre 1900 wiederentdeckt, allerdings wurden sie noch einige Jahre bezweifelt, da man noch keine Aussage fand, wie daraus eine stabile Nachfolgegeneration entstehen kann. Udny Yule argumentierte 1902 gegen die Anwendung[1], da er glaubte, die dominanten Allele müssten sich mit der Zeit in der Population verbreiten. Der US-Amerikaner William Ernest Castle zeigte 1903, dass ohne Selektion die genotypischen Häufigkeiten stabil blieben.[2] Karl Pearson, heute bekannt für seine Beiträge zur Statistik, fand 1903 einen Gleichgewichtspunkt bei p = q = 0,5. Der britische Genetiker Reginald Punnett, der Yules Gegendarstellung nicht widerlegen konnte, befragte seinen Cricket-Spielpartner Godfrey Harold Hardy, einen reinen Mathematiker, der die angewandte Mathematik eigentlich verachtete. Im Jahre 1908 veröffentlichte Hardy einen Beitrag, in dem er das „sehr einfache“ Problem (seine Worte) mit den Begriffen der Biologen erläuterte.
“Suppose that Aa is a pair of Mendelian characters, A being dominant, and that in any given generation the number of pure dominants (AA), heterozygotes (Aa), and pure recessives (aa) are as p:2q:r. Finally, suppose that the numbers are fairly large, so that mating may be regarded as random, that the sexes are evenly distributed among the three varieties, and that all are equally fertile. A little mathematics of the multiplication-table type is enough to show that in the next generation the numbers will be as (p+q)2:2(p+q)(q+r):(q+r)2, or as p1:2q1:r1, say.
The interesting question is — in what circumstances will this distribution be the same as that in the generation before? It is easy to see that the condition for this is q2 = pr. And since q12 = p1r1, whatever the values of p, q, and r may be, the distribution will in any case continue unchanged after the second generation.”
„Nehmen wir an, dass Aa ein Paar von Mendelschen Charakteren sei, A sei dabei dominant, und dass in einer gegebenen Generation die Zahl der reinerbig Dominanten (AA), Heterozygoten (Aa) und reinen Rezessiven (aa) sich wie p:2q:r verhält. Nehmen wir schließlich an, dass die Zahlen hinreichend groß seien, so dass die Paarungen als zufällig angesehen werden können, dass das Geschlechterverhältnis zwischen den 3 Varianten gleichverteilt sei und dass alle gleichermaßen fertil seien. Wenig Mathematik vom Multiplikations-Tabellen-Typ reicht aus, um zu zeigen, dass für die Zahlen in der nächsten Generation gilt: (p+q)2:2(p+q)(q+r):(q+r)2, oder p1:2q1:r1.
Die interessierende Frage ist nun: unter welchen Umständen bleibt die Verteilung dieselbe, wie in der Vorgängergeneration? Es ist einfach zu sehen, dass die Bedingung dafür q2 = pr lautet. Und da q12 = p1r1, unabhängig, welche Werte p, q, und r annehmen, folgt, dass die Verteilung in jedem Fall nach der zweiten Generation dieselbe bleiben wird.“
Damit war dieses Prinzip in der englischsprachigen Welt als „Hardys Gesetz“ bekannt.
Der Beitrag Wilhelm Weinbergs
Ebenfalls im Jahr 1908 hielt der deutsche Arzt und Vererbungsforscher Wilhelm Weinberg einen wissenschaftlichen Vortrag in Stuttgart mit dem Titel „Über den Nachweis der Vererbung beim Menschen“. Darin führte er aus:
„Ganz anders ist das Verhältnis, wenn man die MENDELsche Vererbung unter dem Einfluß der Panmixie betrachtet. Ich gehe dabei von der allgemeinen Voraussetzung aus, daß ursprünglich je m männliche und weibliche reine Vertreter des Typus A und ebenso je n reine Vertreter des Typus B vorhanden seien. Wenn sich diese wahllos kreuzen, so erhält man unter symbolischer Anwendung des binomischen Lehrsatzes als Zusammensetzung der Tochtergeneration:
oder wenn ist
- .
Kreuzt man nun die männlichen und weiblichen Glieder der 1. Generation wahllos untereinander, so erhält man folgende Häufigkeit der verschiedenen Kreuzungskombinationen:
oder die relative Häufigkeit beträgt für
und die Zusammensetzung der zweiten Tochtergeneration ist wieder
- .
Wir erhalten also unter dem Einfluss der Panmixie für jede Generation dieselbe Verteilung der reinen Typen und der Bastarde und damit die Möglichkeit, für jede Generation zu berechnen, wie sich bei den Eltern, Geschwistern und Kindern der verschiedenen Typen und Bastarde bei Panmixie und mendelscher Vererbung die Vertretung dieser Typen stellt.“


















Weinbergs Arbeit blieb im angelsächsischen Raum ganz unbekannt, bis der deutsche Emigrant Curt Stern 1943 auf Weinbergs Arbeiten aufmerksam machte.[7] Seither trägt das populationsgenetische Gesetz den Namen beider Männer. Selten wird auch Castles Name mit hinzugenommen, der das Prinzip früh erkannte, jedoch war dessen Formulierung nicht identisch.
Kennzeichen einer idealen Population
- Sehr große Individuenzahl: Der zufällige Verlust eines Individuums oder Gendrift verändert praktisch nicht die Häufigkeit der Allele, was bei einer kleinen Population relativ große Auswirkungen hätte.
- Panmixie: Alle Paarungen, auch von Trägern verschiedener Genotypen, sind gleich wahrscheinlich und gleich erfolgreich.
- Es gibt keine Selektion, somit also weder Selektionsvorteile noch -nachteile für die Träger bestimmter Gene (Genotyp), die sich phänotypisch auswirken.
- Es finden keine Mutationen statt.
- Es finden keine Zu- oder Abwanderungen (Migration) statt, die die Allelfrequenz verändern.
Die ideale Population ist ein theoretisches Konstrukt, da in der Realität mindestens eine der Bedingungen, welche alle Evolutionsfaktoren sind, nicht erfüllt wird. Evolution findet also stets dann statt, wenn die obigen Voraussetzungen nicht gelten.
Berechnungsformel für 2 Allele
In dem Fall, in dem nur zwei verschiedene Allele P und Q mit den relativen Häufigkeiten ("Allelfrequenzen") p und q existieren, lautet die Formel für das Hardy-Weinberg-Gleichgewicht:

Dabei sind:
- : Allelfrequenz von Allel
- : Allelfrequenz von Allel




Die Schreibweise p2 + 2pq + q2 = 1 ist im biologischen Kontext nützlich. Es gilt nämlich im Hardy-Weinberg-Gleichgewicht:
- : Frequenz der Homozygoten mit Merkmal P
- : Frequenz der Homozygoten mit Merkmal Q
- : Frequenz der Heterozygoten (Merkmale P und Q)



Da es sich bei den Homozygoten- und Heterozygoten-Frequenzen in der Regel um experimentell bestimmbare Größen handelt, kann man daraus die entsprechenden Allelfrequenzen errechnen. Umgekehrt kann man bei Kenntnis einer Allelfrequenz auch die Zahl der zu erwartenden Heterozygoten und Homozygoten berechnen.
1. Beispiel: Phenylketonurie
Die Phenylketonurie ist eine Stoffwechselkrankheit mit autosomal-rezessivem Erbgang. In Deutschland (ca. 80 Millionen Einwohner) gibt es grob geschätzt ungefähr 8.000 Betroffene. Damit ergibt sich für die Homozygotenfrequenz :

Mit

und

folgt

Für die Frequenz der Heterozygoten gilt damit:

Umgerechnet auf die Gesamtbevölkerung ergibt sich für die absolute Zahl der Heterozygoten:

D. h. fast 1,6 Millionen Personen (ca. 2 % der Bevölkerung, etwa eine unter 50 Personen) sind in Deutschland heterozygot für das krankheitsauslösende Phenylketonurie-Allel.
Bei einem sehr kleinen Wert von p kann man in erster Näherung sagen, dass q ≈ 1 und damit gilt näherungsweise für die Heterozygotenfrequenz 2pq ≈ 2p. Im obigen Beispiel ergibt diese Abschätzung den Wert 1,6 Millionen.
2. Beispiel: Chorea Huntington
Die Chorea Huntington ist eine autosomal-dominant vererbte neurologische Erkrankung. Sowohl die Heterozygoten als auch die Homozygoten sind klinisch erkrankt. Die Inzidenz der Erkrankung wird mit 5:100.000 angegeben. Die Erkrankten setzen sich aus den für das krankheitsauslösende Allel p Homozygoten (p2) und die Heterozygoten (2pq) zusammen. Somit sind die Gesunden homozygote Träger des rezessiven Merkmals. Es gilt:

Daraus erhält man q

Und damit p

Die Homozygotenfrequenz (des dominanten Merkmals) ist damit

Das entspräche einer Person auf etwa 1,6 Milliarden. Mit anderen Worten sind mit großer Wahrscheinlichkeit alle in Deutschland an Chorea Huntington erkrankten Personen heterozygot für das krankheitsauslösende Allel. Auch hier gilt in erster Näherung 2pq ≈ 2p.
Verallgemeinerung für mehr als 2 Allele
Die Hardy-Weinberg-Formel lässt sich einfach für den Fall von mehr als 2 Allelen verallgemeinern. Im Folgenden ist der Fall von 3 verschiedenen Allelen P, Q, R mit Allelfrequenzen (p q r) ausgeführt. Dann gilt:

Dabei sind:
- : Frequenz der bezüglich Merkmal P Homozygoten
- : Frequenz der bezüglich Merkmal Q Homozygoten
- : Frequenz der bezüglich Merkmal R Homozygoten
- : Frequenz der bezüglich der Merkmale P und Q Heterozygoten
- : Frequenz der bezüglich der Merkmale Q und R Heterozygoten
- : Frequenz der bezüglich der Merkmale P und R Heterozygoten



Verallgemeinert auf n Allele A1 … An mit den relativen Häufigkeiten p1, …, pn gilt dann im Hardy-Weinberg-Gleichgewicht:

mit den jeweiligen Homozygotenfrequenzen von Merkmal Ai:

und den Heterozygotenfrequenzen (Merkmale Ai und Aj):
- .

Beispiel: AB0-Blutgruppensystem (3 Allele)


Die Allele für die Blutgruppen A und B sind kodominant, während das Allel für die Blutgruppe 0 rezessiv ist. Wenn die Häufigkeiten der Gene für A, B und 0 im Genpool a, b und o sind (mit a+b+o=1), gilt für die Häufigkeit der Blutgruppen (Phänotypen):
- : Frequenz der Personen mit Blutgruppe A
- : Frequenz der Personen mit Blutgruppe B
- : Frequenz der Personen mit Blutgruppe AB
- : Frequenz der Personen mit Blutgruppe 0




Die in Deutschland beobachteten Blutgruppenhäufigkeiten sind: Blutgruppe A 43 %, Blutgruppe 0 41 %, Blutgruppe B 11 % und Blutgruppe AB 5 %.[8]
Daraus ergeben sich die folgenden Beziehungen (Zahlenwerte gerundet):
1. Für das Allel :


2. Für das Allel :





3. Für das Allel :





Die aus beobachteten Daten berechneten Werte für die Allelfrequenzen entsprechen einem Hardy-Weinberg-Gleichgewicht:

Verallgemeinerung für einen polyploiden Chromosomensatz
Die oben aufgeführten Beispielrechnungen beziehen sich auf einen diploiden Chromosomensatz, wie er z. B. beim Menschen vorliegt, bei dem Allele immer doppelt vorhanden sind (jedes Chromosom ist immer zweimal vorhanden, eine Ausnahme bilden nur die Geschlechtschromosomen). Viele Organismen weisen jedoch polyploide Chromosomensätze auf, in denen ein Allel mehr als zweimal vorhanden ist. Die Hardy-Weinberg-Formel lässt sich auch auf solche Fälle verallgemeinern.
Zwei Allele in einem polyploiden Chromosomensatz

wobei x der Ploidiegrad ist (diploid: x=2; triploid: x=3; tetraploid: x=4; etc.), und
- : Allelfrequenz von Allel
- : Allelfrequenz von Allel
Für einen triploiden Chromosomensatz ergibt sich:

| Genotyp | Frequenz |
|---|---|








Für einen tetraploiden Chromosomensatz ergibt sich:

| Genotyp | Frequenz |
|---|---|










Mehr als zwei Allele in einem polyploiden Chromosomensatz
Im allgemeinsten Fall ergibt sich für einen polyploiden Chromosomensatz mit Ploidiegrad x und n verschiedenen Allelen:

bzw.

mit dem „Multiindices“ k = (k1,k2,…,kn) und pk = p1k1p2k2⋯pnkn.
Weblinks
- Arbeitsmaterial und Berechnungen zur Populationsgenetik (Memento vom 27. September 2007 im Internet Archive) (PDF-Datei; 401 kB)
- Simulation zum Gesetz von Hardy-Weinberg
Einzelnachweise
- George Udny Yule: Mendel's laws and their probable relation to intra-racial heredity. In: New Phytologist. Band 1, Nr. 9, 1902, S. 193–207, doi:10.1111/j.1469-8137.1902.tb06590.x.
- William E. Castle: The Laws of Heredity of Galton and Mendel, and Some Laws Governing Race Improvement by Selection. In: Proceedings of the American Academy of Arts and Sciences. Band 39, Nr. 8, 1903, S. 223–242, doi:10.2307/20021870.
- Godfrey H. Hardy: Mendelian proportions in a mixed population. In: Science. Band 28, Nr. 706, 1908, S. 49–50, doi:10.1126/science.28.706.49.
- Sic in der Niederschrift, hier müsste eigentlich links × stehen.
- Sic in der Niederschrift, hier müsste eigentlich rechts stehen.
- Wilhelm Weinberg: Über den Nachweis der Vererbung beim Menschen. In: Jahreshefte des Vereins für Vaterländische Naturkunde in Württemberg. Band 64, 1908, ISSN 0368-4717, S. 369–382, Digitalisat.
- Curt Stern: The Hardy–Weinberg law. In: Science. Band 97, Nr. 2510, 1943, S. 137–138, doi:10.1126/science.97.2510.137. PMID 17788516.
- Blutgruppen & ihre Verteilung. Blutspendedienst des bayrischen Roten Kreuzes, abgerufen am 9. Mai 2016.


