Haptolina
Haptolina ist eine Gattung von Haptophyten, die zur Familie der Prymnesiaceae gehört.[1] Die Gattung wurde von Chrysochromulina ausgegliedert.[2]
| Haptolina | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
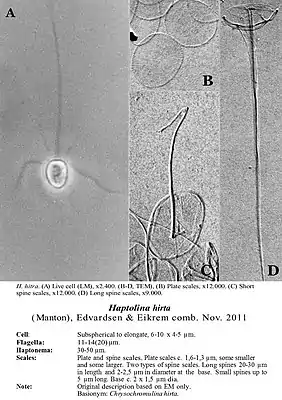
H. hirta von der Ostsee, Schuppen und Dornen | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Haptolina | ||||||||||||
| Edvardsen & Eikrem, 2011 |

Die Gattung hat eine kosmopolitische Verbreitung.[1] Einige Arten können Algenblüten (mit-)
Arten
Die Systematik der Gattung Haptolina ist wie folgt:[4][1][5][6]
Gattung Haptolina Edvardsen & Eikrem, 2011
- Haptolina brevifila (Parke & Manton) Edvardsen & Eikrem, 2011[7] – Typus; früher Chrysochromulina brevifilum
- Haptolina ericina (Parke & Manton) Edvardsen & Eikrem, 2011[8] – früher C. ericina
- Haptolina fragaria (Eikrem & Edvardsen) Eikrem & Edvardsen, 2011[9] – früher C. fragaria
- Haptolina herdlensis (B.Leadbeater) Edvardsen & Eikrem, 2011[10] – früher C. herdlensis
- Haptolina hirta (Manton) Edvardsen & Eikrem, 2011[11] – früher C. hirta
- „Haptolina sp. RCC2300“ – früher „Chrysochromulina sp. RCC2300“ (NCBI)[12]
Algenblüten
Kattegat-Algenblüte 1992: Von April bis Mai 1992 gab es im südlichen Kattegat eine große Blüte, die aus vielen Phytoplanktonarten bestand, wobei über 90 % der Biomasse Chrysochromulina-Arten waren (viele aber inzwischen der Gattung Haptolina zugeordnet).[3] Die am häufigsten vorkommenden Arten in der Blüte waren:[3]
- Haptolina hirta (früher Chrysochromulina hirta),
- H. ericina (früher C. ericina),
- H. brevifila (früher C. brevifilum),
- Chrysochromulina spinifera, sowie
- eine unbeschriebene Art (Ø. Moestrup).
H. hirta, H. ericina und C. spinifera zeichnen sich durch relativ kleine Zellen mit langen Dornen aus, die den Gesamtorganismen einen Durchmesser von 25–76 μm verleihen, was für die vorhandenen Ciliaten (als potentielle Fressfeinde) zu groß ist, um sie zu verschlingen; wahrscheinlich ein Grund für den „Erfolg“ der Blüte war.[3]
Es gab zwar keine Beweise, die diese Blüte oder die vorhandenen Arten direkt mit der Produktion von Toxinen (wie bei der C. polylepis-Blüte 1988) in Verbindung brachten.[13][3] Es wurden jedoch etwa 50 Tonnen toter Regenbogenforellen (Oncorhynchus mykiss) in Verbindung mit der Blüte gebracht.[3]
Viren
Es wurde über mehrere Virenspezies-Kandidaten berichtet, insbesondere über:
- Chrysochromulina-ericina-Virus 01B (CeV-01B)[14] alias Haptolina-ericina-Virus 01B (HeV-01B), Spezies Tethysvirus raunefjordenense, Ordnung Imitervirales[15][16][14]
- „Haptolina ericina virus RF02“ (HeV-RF02) – mit Stand Ende April 2023 noch nicht vom International Committee on Taxonomy of Viruses (ICTV) offiziell bestätigtes Mitglied derselben Familie.
CeV-01B/HeV-01B
CeV-01B (HeV-01B) wurde erstmals 1998 aus Küstengewässern Norwegens isoliert. Es handelt sich um ein ikosaedrisches DNA-Doppelstrangvirus mit einer Genomgröße von 474 kbp (Kilobasenpaaren).[14]
HeV-RF02
HeV-RF02 wurde isoliert auf dem Stamm Haptolina ericina UIO028 aus dem Raunefjord (Provinz Vestland, Norwegen). Torill Vik Johannessen und Kollegen berichteten 2015 über die Entdeckung dreier neuer lytischer Doppelstrang-DNA-Viren aus dem Raunefjord, die der Familie Mimiviridae nahezustehen schienen oder zu ihr gehörten. Neben „Haptolina ericina virus RF02“ (HeV-RF02) waren die beiden anderen Kandidaten Prymnesium kappa virus RF01 (PkV-RF01, ICTV-bestätigt in der Spezies Biavirus raunefjordenense)[17][18][19][15][16] und „Prymnesium kappa virus RF02“ (PkV-RF02). Jedes dieser neuen Haptophytenviren stellte zum Zeitpunkt der Entdeckung die gängigen Vorstellungen von Algenviren in Bezug auf Wirtsspektrum, phylogenetische Zugehörigkeit und Größe in Frage. HeV-RF02 war (mit einem Kapsiddurchmesser von ca. 310 nm) das bis dato größte bekannte Algenvirus. Die Ergebnisse zeigten eine große Variation unter Viren, die eng verwandte Mikroalgen infizieren, und stellen damit die gängige Vorstellung in Frage, dass Algenviren einen engen Wirtsbereich haben der die Phylogenie ihre Wirtszugehörigkeit widerspiegelt.[20] HeV-RF02 und PkV-RF02 stehen CeV-01B/HeV-01B, Phaeocystis globosa virus 12 (PgV-12T, Spezies Tethysvirus hollandense) und „Phaeocystis pouchetii virus 01“ (PpV-01) nahe, alle in der Familie Mesomimiviridae (alias OLPG) der Ordnung Imitervirales.[15][16][20]
Einzelnachweise
- Haptolina Edvardsen & Eikrem, 2011. In: www.gbif.org. Abgerufen am 25. April 2021 (englisch).
- B. Edvardsen, W. Eikrem, J. Throndsen, A. G. Sáez, I. Probert, L. K. Medlin: Ribosomal DNA phylogenies and a morphological revision provide the basis for a revised taxonomy of the Prymnesiales (Haptophyta). European Journal of Phycology, Band 46, Nr. 3, 2011, S. 202–228
- Per Juel Hansen, Torkel Gissel Nielsen, Hanne Kaas: Distribution and Growth of Protists and Mesozooplankton during a Bloom of Chrysochromulina Spp. (Prymnesiophyceae, Prymnesiales). In: Phycologia. 34. Jahrgang, Nr. 5, 1995, S. 409–416, doi:10.2216/i0031-8884-34-5-409.1. Epub 22. Apr 2019. Freier Volltext (ResearchGate)
- AlgaeBase: AlgaeBase: Haptolina Edvardsen & Eikrem, 2011
- NCBI: Haptolina Edvardsen & Eikrem (genus); graphisch: Haptolina, auf: Lifemap, NCBI Version.
- WoRMS: Haptolina Edvardsen & Eikrem, 2011
- Nordic Microalgae: Chrysochromulina brevifilum, auf: Nordic Microalgae and aquatic protozoa, Swedish Meteorological and Hydrological Institute (SMHI)
- Nordic Microalgae: Chrysochromulina ericina, auf: Nordic Microalgae and aquatic protozoa, Swedish Meteorological and Hydrological Institute (SMHI)
- Nordic Microalgae: Chrysochromulina fragaria, auf: Nordic Microalgae and aquatic protozoa, Swedish Meteorological and Hydrological Institute (SMHI)
- Nordic Microalgae: Chrysochromulina herdlensis, auf: Nordic Microalgae and aquatic protozoa, Swedish Meteorological and Hydrological Institute (SMHI)
- Nordic Microalgae: Chrysochromulina hirta, auf: Nordic Microalgae and aquatic protozoa, Swedish Meteorological and Hydrological Institute (SMHI)
- Sergio Balzano, Dominique Marie, Priscillia Gourvil, Daniel Vaulot: Composition of the summer photosynthetic pico and nanoplankton communities in the Beaufort Sea assessed by T-RFLP and sequences of the 18S rRNA gene from flow cytometry sorted samples, in: ISME J, Band 6, Nr. 8, 2012, S. 1480–1498, PMID 22278671, PMC 3400408 (freier Volltext)
- Torkel Nielsen, Thomas Kiørboe, Peter Bjørnsen: Effects of a Chrysochromulina polylepis subsurface bloom on the planktonic community. In: Marine Ecology Progress Series. 62. Jahrgang, Nr. 1/2, 1990, S. 21–35, doi:10.3354/meps062021, bibcode:1990MEPS...62...21N (englisch, int-res.com [PDF]).
- Lucie Gallot-Lavallée, António Pagarete, Matthieu Legendre, Sebastien Santini, Ruth-Anne Sandaa, Heinz Himmelbauer, Hiroyuki Ogata, Gunnar Bratbak, Jean-Michel Claverie: The 474-Kilobase-Pair Complete Genome Sequence of CeV-01B, a Virus Infecting Haptolina (Chrysochromulina) ericina (Prymnesiophyceae). In: Genome Announcements. 3. Jahrgang, Nr. 6, 3. Dezember 2015, doi:10.1128/genomeA.01413-15, PMID 26634761, PMC 4669402 (freier Volltext) – (englisch).
- ICTV: Master Species Lists § ICTV Master Species List 2022 MSL38 v1 (xlsx), 8. April 2023.
- Frank O. Aylward, Jônatas S. Abrahão, Corina P. D. Brussaard C, Matthias G. Fischer, Mohammad Moniruzzaman, Hiroyuki Ogata, Curtis A. Suttle: Create 3 new families, 3 subfamilies, 13 genera, and 20 new species within the order Imitervirales (phylum Nucleocytoviricota) and rename two existing species (zip:docx). Vorschlag 2022.004F an das ICTV vom Oktober 2021.
-
Romain Blanc-Mathieu, Håkon Dahle, Antje Hofgaard, David Brandt, Hiroki Ban, Jörn Kalinowski, Hiroyuki Ogata, Ruth-Anne Sandaa: A Persistent Giant Algal Virus, with a Unique Morphology, Encodes an Unprecedented Number of Genes Involved in Energy Metabolism, in: ASM Journals: Journal of Virology, Band 95, Nr. 8, 25. März 2021, doi:10.1128/JVI.02446-20 .
PrePrint: bioRxiv, doi:10.1101/2020.07.30.228163, CSH, 13. Januar 2021 - NCBI: Prymnesium kappa virus (species) – unclassified Phycodnaviridae
- Lucie Gallot-Lavallee, Guillaume Blanc, Jean-Michel Claverie: Comparative genomics of Chrysochromulina Ericina Virus (CeV) and other microalgae-infecting large DNA viruses highlight their intricate evolutionary relationship with the established Mimiviridae family (PDF) in: J. Virol., 26 April 2017, doi:10.1128/JVI.00230-17
- Torill Vik Johannessen, Gunnar Bratbak, Aud Larsen, Hiroyuki Ogatac, Elianne S. Egged, Bente Edvardsen, Wenche Eikremd, Ruth-Anne Sandaa: Characterisation of three novel giant viruses reveals huge diversity among viruses infecting Prymnesiales (Haptophyta), in: Virology, Band 476, Februar 2015, S. 180–188, doi:10.1016/j.virol.2014.12.014, PMID 25546253, insbes. Fig. 4 und Fig. S2
{kind=link}