Hacrobia
Die Kryptomonaden-Haptophyten-Gruppe (englisch cryptomonads-haptophytes assemblage) ist eine vorgeschlagene monophyletische Gruppierung (Klade) von einzelligen Eukaryoten, die nicht zur SAR-Supergruppe gehören.[1] Mehrere alternative Namen wurden für diese Gruppe verwendet, darunter
- Hacrobia, (abgeleitet von „ha-“, für Haptophyta, „-cr-“, für Cryptomonada, und „-bia“ als allgemeiner Suffix, Plural von „-bion“, der sich auf Lebewesen im Allgemeinen bezieht anstelle der speziellen „-zoa“ und „-phyta“ für Tiere bzw. Pflanzen[2][3])[4]
- CCTH (für Cryptophyta, Centrohelida, Telonemia, Haptophyta);[5] sowie
- Eukaryomonadae.[6]
| Hacrobia | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|

Gephyrocapsa oceanica (Prymnesiophyceae). | ||||||||||
| Systematik | ||||||||||
| ||||||||||
| Wissenschaftlicher Name | ||||||||||
| Hacrobia | ||||||||||
| Okamoto et al., 2009 |
Mit Stand Februar 2012 war noch unklar, ob diese Gruppe tatsächlich monophyletisch ist, die Ergebnisse phylogenetischer Studien zeigten sich bis dato „oft von der Auswahl der Taxa und des Gendatensatzes abhängig“[5] und zwei Studien aus diesem Jahr kamen zu gegenteiligen Ergebnissen.[5][7] Die Hacrobia werden gegenwärtig (Mitte Januar 2022) von der Global Biodiversity Information Facility (GBIF) und vom World Register of Marine Species (Worms) als gültiges Taxon gelistet.[8][9]
Forschungsgeschichte



.png.webp)

_(1910)_(17950796051)-9%252B10%252B11.jpg.webp)

Heterokonta, Haptophyta und Cryptomonada (alias Cryptophyta) wurden manchmal in einer Gruppe namens Chromista zusammengefasst (Csurös et al. 2008).[10] Andere Studien zeigten deuteten aber darauf hin, dass die Cryptophyta und Haptophyta vergleichsweise eng miteinander verwandt sind, die Heterokonta aber nicht mit dazu gehören (Rice et al. 2006 und Oren et al. 2010),[11][12] sodass diese Klade manchmal einfach als „Cryptophyta+Haptophyta“ bezeichnet wurde (Reeb et al. 2009).[13] In einem weiteren Artikel aus dem Jahr 2009 schlugen Burki et al. vor, dass die Telonemia (alias Telonemea) und die Centrohelida (d. h. Heliozoen mit einem Zentroplast und exzentrisch gelegenem Zellkern) in die Gruppe mit diesen Cryptophyta und Haptophyten einbezogen werden sollten.[14] Die Picobiliphyta (alias Picozoa) könnten ebenfalls dieser gemeinsamen Gruppe angehören, aber es fehlen genug Anhaltspunkte für eine sichere taxonomische Klassifizierung.[4] Damit wären dann die wesentlichen Mitglieder der Hacrobia zusammengefasst.
Mehrere neuere Studien haben allerdings ein abweichendes Bild entworfen, in dem die Haptophyta und die Cryptophyta keine monophyletische Gruppe bilden.[15] Die Haptophyta könnten danach eine Schwestergruppe der SAR-Supergruppe darstellen und die Cryptophyta zu den Archaeplastida (Pflanzen im weiteren Sinne: Glaucophyta, Rotalgen, sowie Chloroplastida alias Viridiplantae: Grünalgen und Landpflanzen). Mit Stand Februar 2012 ist nach wie vor unklar, ob die Hacrobia eine monophyletische Gruppe bilden.[5]
Burki et al. schlugen 2012 eine Gliederung vor, in der die Centrohelida mit den Haptophyta verwandt sind und gemeinsam die Klade der Haptista[16] bilden (was alleine mit den Hacrobia verträglich wäre, s. u.). Die Cryptista (mit den Cryptophyta) erscheinen hier aber als verwandt mit den Archaeplastida. Zusammen erscheinen hier Haptista+SAR als Schwestergruppe von Cryptista+Archaeplastida.[17]
Phylogenie
Die folgende Phylogenie basiert (wo nicht anders angegeben) auf Studien von Ruggiero et al. (2015) und Silar (2016). Die Hauptzweige der Hacrobia sind danach die Cryptista (mit den Cryptomonada und Telonemea), die Haptista[16] (mit den Centrohelida/Centroheliozoa und den Haptophyta/Haptomonda),[18][19] sowie die Alveidia (mit der einzigen Gattung Ancoracysta):[20][21]
| Hacrobia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Mitglieder
Die Hacrobia bestehen – neben den Alveidia – aus zwei Hauptgruppen, den Haptophyta (Haptomonada) und den Cryptophyta (Cryptomonada). Beide sind Algen, die Chloroplasten mit den Chlorophyllen a und c besitzen. Weil sie von vier Membranen umgeben sind und aufgrund der Art der Chlorophylle nimmt man an, dass diese Chloroplasten aus einer sekundären Endosymbiose mit einer Rotalge stammen (siehe komplexe Chloroplasten). Darüber hinaus gibt es mehrere Gruppen von heterotrophen Organismen, die früher gemeinsam als Heliozoa (Sonnentierchen) klassifiziert wurden, die aber eine erwiesenermaßen nicht-monophyletischen Gruppe darstellen. Die hier als Endohelia bzw. Endohelea Cavalier-Smith 2012 bezeichnete Gruppe Endohelia umfasst zwei kleinere Gruppen, Microhelida und Heliomonadida, und ist zu unterscheiden von Endohelia Milne Edwards & Haime, 1849, einer veralteten Bezeichnung für die Filigrankorallen-Gattung Crypthelia Milne Edwards & Haime, 1849.[22]
Anmerkungen zu den einzelnen Gruppen:[23]
Haptista
Die Linie der Haptista fasst die Haptophyta-Algen und die Centrohelida-Heliozoen (Centroheliden) zusammen. Sie zeichnen sich aus durch mikrotubulär gestützte fadenförmige Anhängsel, die der Nahrungsaufnahme dienen: das Haptonema bei ersteren und die Axopoden – axiale Pseudopodien – bei letzteren. Bei beiden Gruppen ist der Zellkörper oft mit komplexen mineralisierten Schuppen bedeckt (kieselhaltig bei Centroheliden, meist kalkhaltig, aber manchmal auch kieselhaltig bei den Haptophyten).
- Haptomonada (Haptophyta): Eine Gruppe von hauptsächlich mariner einzelliger Algen, einige Mitglieder sind jedoch Mixotrophe oder Heterotrophe. Sie zeichnen sich aus durch zwei eukaryotische Geißeln, einem Haptonema (einem oberflächlich gesehen einer Geißel ähnlichen Anhängsel, das sich von diesen aber durch Mikrotubulus-Bündel und die Verwendung unterscheidet), und nackten oder mit Kalkplatten bedeckten Zellen.[24]
- Rappemonada (Rappephyceae): Eine Gruppe von Meeres- und Süßwasser-Protisten, die wahrscheinlich mit den obigen Haptomonaden verwandt ist. Die Zellen enthalten Chloroplasten mit Chlorophyll a. Sie sind nur aus Umweltproben bekannt, schlecht charakterisiert und noch keine Art oder Gattung wurde beschrieben.[25][26][27]
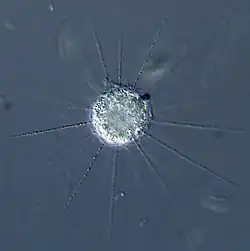
- Centroheliozoa (Centrohelida, Centroplasthelida):[28] Eine wichtige Teilgruppe der Sonnentierchen (Heliozoen, nicht-taxonomisch), charakterisiert als annähernd kugelförmige Einzeller, die strahlenförmig von axialen Pseudopodien (Scheinfüßchen), hier auch Axopodien genannt, umgeben sind, und die dazu dienen, Nahrung zu fangen. Charakteristisch für die Centroheliden ist eine in der Mitte gelegene dreiteilige Granula, genannt Zentroplast und ein exzentrisch gelegener Zellkern.
Die Axopodien werden von Mikrotubuli in einer dreieckig-hexagonalen Anordnung getragen, die vom Zentroplast ausgeht. Die Gruppe umfasst sowohl freie als auch sessile Formen in Süßwasser- oder Meereslebensräumen.
Cryptista
Die Linie der Cryptista zeichnet sich durch Anhängsel in Form eines Tripletts von röhrenförmigen Fasern an der vorderen Geißel aus. Sie umfasst die folgenden Gruppen:
- Cryptophyta (Cryptomonada): Eine Gruppe einzelliger Algen, die in Meeres- und Binnengewässern leben, wobei einige Mitglieder mixotroph oder heterotroph sind. Die meisten besitzen eine starre Schale, eine ventrale Einstülpung mit zwei Geißeln und mehrere Reihen von Ejektosomen, die eine Verteidigungsfunktion haben.
- Telonemia (Telonemea): Eine kleine Gruppe von Organismen, die sowohl im Meer als auch im Süßwasser vorkommen. Sie sind heterotroph und ernähren sich von einer breiten Palette von Bakterien sowie von Piko- und Nano-Phytoplankton.
- Palpitia: Diese Gruppe besteht aus einer einzigen biflagellaten marinen heterotrophen Spezies Palpitomonas bilix[29][30][31] mit zwei nahezu gleich großen Geißeln. Ultrastrukturelle Analysen ergaben, dass ihr Geißelapparat dem von Grünalgen (Charophyceae) ähnelt, während die Mastigonema einige Merkmale der Cryptophyta und Telonemia aufweisen.
- Picomonadea (Picozoa): Eine vor noch nicht langer Zeit entdeckte Gruppe mariner heterotropher Protisten, bestehend aus der einzigen Gattung Picomonas,[32] und nur etwa 3 µm groß.
- Microhelida: Diese Teilgruppe der Sonnentierchen (Heliozoen, nicht-taxonomisch) besteht ebenfalls nur aus einer einzigen Spezies, Microheliella maris,[33] sehr kleine Meeresorganismen, die sich von Bakterien ernähren. Sie haben einen großen Zentroplast im Zentrum der Zelle und sind von Axopodien umgeben, die dem Nahrungserwerb dienen. Die Axoneme der Axopodien sind in einer Triade angeordnet, was sie von anderen Gruppen von Heliozoen wie den Centrohelida unterscheidet.
- Heliomonadida[34] (Dimorphida[35]): Eine kleine Teilgruppe von Sonnentierchen (Heliozoen, nicht-taxonomisch), die sich von den anderen dadurch auszeichnen, dass sie während ihres gesamten Lebenszyklus Geißeln besitzen (zwei oder vier). Die typischerweise quadratisch angeordneten Mikrotubuli-Bündel gehen von einem Körper nahe der Basis der Geißeln aus und tragen zahlreiche Axopodien, die von der Zelloberfläche abstehen.
Einzelnachweise
- M. Sakaguchi, K. Takishita, T. Matsumoto, T. Hashimoto, Y. Inagaki: Tracing back EFL gene evolution in the cryptomonads-haptophytes assemblage: separate origins of EFL genes in haptophytes, photosynthetic cryptomonads, and goniomonads. In: Gene. 441. Jahrgang, Nr. 1–2, Juli 2009, S. 126–131, doi:10.1016/j.gene.2008.05.010, PMID 18585873.
- Eugene M. McCarthy: Suffix Dictionary – B-C §-bia, Medical Suffixes, B-C. Auf macroevolution.net
- Wiktionary: βίος
- N. Okamoto, C. Chantangsi, A. Horák, B. Leander, P. Keeling, J. E. Stajich: Molecular Phylogeny and Description of the Novel Katablepharid Roombia truncata gen. et sp. nov., and Establishment of the Hacrobia Taxon nov. In: PLOS ONE. 4. Jahrgang, Nr. 9, 2009, S. e7080, doi:10.1371/journal.pone.0007080, PMID 19759916, PMC 2741603 (freier Volltext), bibcode:2009PLoSO...4.7080O.
- Sen Zhao, Fabien Burki, Jon Bråte, Patrick J. Keeling, Dag Klaveness, Kamran Shalchian-Tabriz: Collodictyon—An Ancient Lineage in the Tree of Eukaryotes. In: Molecular Biology and Evolution. 29. Jahrgang, Nr. 6, 2012, S. 1557–1568, doi:10.1093/molbev/mss001, PMID 22319147, PMC 3351787 (freier Volltext).
- Jack R. Holt: DESCRIPTION OF THE KINGDOM EUKARYOMONADAE. 19. Februar 2013, abgerufen am 15. Januar 2022. Systematic Biology — Diversity of Life
- F. Burki, N. Okamoto, J. F. Pombert, P. J. Keeling: The evolutionary history of haptophytes and cryptophytes: phylogenomic evidence for separate origins. In: Proc. Biol. Sci. 279. Jahrgang, Nr. 1736, 2012, S. 2246–2254, doi:10.1098/rspb.2011.2301, PMID 22298847, PMC 3321700 (freier Volltext).
- GBIF: Acanthocystis Carter, 1863 – zeigt in der Taxonomie dieser Gattung „Hacrobia Okamoto, Chantangsi, Horåk, Leander & Keeling, 2009“.
- WoRMS: Hacrobia
- M. Csurös, I. B. Rogozin, E. V. Koonin: Extremely intron-rich genes in the alveolate ancestors inferred with a flexible maximum-likelihood approach. In: Mol. Biol. Evol. 25. Jahrgang, Nr. 5, Mai 2008, S. 903–911, doi:10.1093/molbev/msn039, PMID 18296415.
- D. W. Rice, J. D. Palmer: An exceptional horizontal gene transfer in plastids: gene replacement by a distant bacterial paralog and evidence that haptophyte and cryptophyte plastids are sisters. In: BMC Biol. 4. Jahrgang, 2006, S. 31, doi:10.1186/1741-7007-4-31, PMID 16956407, PMC 1570145 (freier Volltext).
- Aharon Oren, R. Thane Papke: Molecular Phylogeny of Microorganisms. Horizon Scientific Press, 2010, ISBN 978-1-904455-67-7, S. 190– (google.com [abgerufen am 21. Januar 2011]).
- Valérie C. Reeb, Michael T. Peglar, Hwan Su Yoon et al.: Interrelationships of chromalveolates within a broadly sampled tree of photosynthetic protists. In: Mol. Phylogenet. Evol. 53. Jahrgang, Nr. 1, Mai 2009, S. 202–211, doi:10.1016/j.ympev.2009.04.012, PMID 19398025.
- Fabien Burki, Yuji Inagaki, Jon Bråte, John M Archibald, Patrick J Keeling, Thomas Cavalier-Smith, Miako Sakaguchi, Tetsuo Hashimoto, Ales Horak, Surendra Kumar, Dag Klaveness, Kjetill S Jakobsen, Jan Pawlowski, Kamran Shalchian-Tabrizi: Large-scale phylogenomic analyses reveal that two enigmatic protist lineages, Telonemia and Centroheliozoa, are related to photosynthetic chromalveolates. In: Genome Biology and Evolution. 1. Jahrgang, 2009, S. 231–238, doi:10.1093/gbe/evp022, PMID 20333193, PMC 2817417 (freier Volltext).
- Denis Baurain, Henner Brinkmann, Jörn Petersen, Naiara Rodríguez-Ezpeleta, Alexandra Stechmann, Vincent Demoulin, Andrew J. Roger, Gertraud Burger, B. Franz Lang, Hervé Philippe: Phylogenomic Evidence for Separate Acquisition of Plastids in Cryptophytes, Haptophytes, and Stramenopiles. In: Molecular Biology and Evolution. 27. Jahrgang, Nr. 7, 2010, S. 1698–1709, doi:10.1093/molbev/msq059, PMID 20194427.
- NCBI: Haptista, Details: Haptista Cavalier-Smith, 2003 (clade); graphisch: Haptista, auf: Lifemap NCBI Version.
- Fabien Burki, Maia Kaplan, Denis V. Tikhonenkov, Vasily Zlatogursky, Bui Quang Minh, Liudmila V. Radaykina, Alexey Smirnov, Alexander P. Mylnikov, Patrick J. Keeling: Untangling the early diversification of eukaryotes: a phylogenomic study of the evolutionary origins of Centrohelida, Haptophyta and Cryptista. In: Proc Biol Sci. 283. Jahrgang, Nr. 1823, 27. Januar 2016, S. 20152802, doi:10.1098/rspb.2015.2802, PMID 26817772, PMC 4795036 (freier Volltext).
- Philippe Silar: Protistes Eucaryotes: Origine, Evolution et Biologie des Microbes Eucaryotes. In: HAL Archives-ouvertes. 2016, S. 1–462 (archives-ouvertes.fr).
- Michael A. Ruggiero, Dennis P. Gordon, Thomas M. Orrell, Nicolas Bailly, Thierry Bourgoin, Richard C. Brusca, Thomas Cavalier-Smith, Michael D. Guiry, Paul M. Kirk: Higher Level Classification of All Living Organisms. In: PLOS ONE. 10. Jahrgang, Nr. 4, 29. April 2015, S. e0119248, doi:10.1371/journal.pone.0119248, PMID 25923521, PMC 4418965 (freier Volltext), bibcode:2015PLoSO..1019248R.
- Thomas Cavalier-Smith, E. E. Chao, R. Lewis: Multigene phylogeny and cell evolution of chromist infrakingdom Rhizaria: contrasting cell organisation of sister phyla Cercozoa and Retaria. In: Protoplasma. 255. Jahrgang, Nr. 5, 17. April 2018, S. 1517–1574, doi:10.1007/s00709-018-1241-1, PMID 29666938, PMC 6133090 (freier Volltext).
- NCBI: Ancoracysta, Details: Ancoracysta Janoukovec, Tikhonenkov, Burki, Howe, Rohwer, Mylnikov & Keeling 2017 (genus)
- WoRMS: Endohelia Milne Edwards & Haime, 1849
- Thomas Cavalier-Smith, Ema E. Chao, Rhodri Lewis: Multiple origins of Heliozoa from flagellate ancestors: New cryptist subphylum Corbihelia, superclass Corbistoma, and monophyly of Haptista, Cryptista, Hacrobia and Chromista. In: Molecular Phylogenetics and Evolution, Band 93, Dezember 2015, S. 331–362. doi:10.1016/j.ympev.2015.07.004.
- NCBI: Haptophyta, Details: Haptophyta Hibberd, 1976 (phylum); graphisch: Haptophyta, auf: Lifemap NCBI Version
- AlgaeBase: Class Rappephyceae M.Kawachi, R.Kamikawa & T.Nakayama 20219
- New class of marine phytoplankton named for UH professor, University of Hawaiʻi News. 22. April 2021. ProQuest 2516733806.
- Masanobu Kawachi, Takuro Nakayama, Motoki Kayama, Ian Probert, Isao Inouye, Ryoma Kamikawa et al.: Rappemonads are haptophyte phytoplankton. In: Current Biology, Band 31, 7. Juni 2021, S. 2395–2403; doi:10.1016/j.cub.2021.03.012.
- NCBI: Centroplasthelida, Details: Centroplasthelida Febvre-Chevalier & Febvre 1984 (class), heterotypic synonyms: Centroheliozoa, Centrohelida Kuehn 1926, centrohelids; graphisch: Centroplasthelida (centrohelids), auf: Lifemap NCBI Version
- WoRMS: Palpitia (Subphylum)
- WoRMS: Palpitomonas A. Yabuki & K.-I. Ishida in Yabuki, Inagaki & Ishida, 2010
- NCBI: Palpitomonas, Details: Palpitomonas Yabuki, Inagaki & Ishida 2010 (genus) — korrigiert
- NCBI: Picomonas, Details: Picomonas Seenivasan, Sausen, Medlin & Melkonian, 2013 (genus)
- NCBI: Microheliella, Details: Microheliella Cavalier-Smith & Chao 20120 (genus)
- David Bass, Ema E.-Y. Chao, Sergey Nikolaev, Akinori Yabuki, Ken-Ichiro Ishida, Cédric Berney, Ursula Pakzad, Claudia Wylezich, Thomas Cavalier-Smith: Phylogeny of novel naked Filose and Reticulose Cercozoa: Granofilosea cl. n. and Proteomyxidea revised. In: Protist. 160. Jahrgang, Nr. 1, Februar 2009, S. 75–109, doi:10.1016/j.protis.2008.07.002, PMID 18952499.
- Sergey I. Nikolaev, Cédric Berney, José F. Fahrni, Ignacio Bolivar, Stephane Polet, Alexander P. Mylnikov, Vladimir V. Aleshin, Nikolai B. Petrov, Jan Pawlowski: The twilight of Heliozoa and rise of Rhizaria, an emerging supergroup of amoeboid eukaryotes. In: Proc. Natl. Acad. Sci. U.S.A. 101. Jahrgang, Nr. 21, Mai 2004, S. 8066–8071, doi:10.1073/pnas.0308602101, PMID 15148395, PMC 419558 (freier Volltext).