Gorgosaurus
Gorgosaurus (griech.: „ungestüme Echse“, von γοργός gorgos „ungestüm, schrecklich“ und σαῦρος sauros „Echse“) war ein theropoder Dinosaurier aus der Familie der Tyrannosauridae, der vor etwa 76 bis 72 Millionen Jahren in der Oberkreide (spätes Campanium) im westlichen Nordamerika lebte. Fossile Überreste wurden in der kanadischen Provinz Alberta und möglicherweise im US-amerikanischen Bundesstaat Montana entdeckt. Zurzeit wird von dieser Gattung lediglich eine Art anerkannt, die Typusart Gorgosaurus libratus.
| Gorgosaurus | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Skelettrekonstruktion von Gorgosaurus | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Oberkreide (spätes Campanium)[1] | ||||||||||||
| 76,4 bis 72 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Gorgosaurus | ||||||||||||
| Lambe, 1914 | ||||||||||||
| Art | ||||||||||||
|
Wie die meisten bekannten Tyrannosauriden war auch Gorgosaurus ein bipeder Fleischfresser, der als adultes Tier mehr als eine Tonne wog und dutzende von großen, scharfen Zähnen im Kiefer trug, während die zweifingrigen Arme im Verhältnis klein waren. Gorgosaurus war nah mit dem sehr ähnlichen Albertosaurus verwandt; beide Gattungen werden nur anhand von geringfügigen Unterschieden an den Zähnen und Schädelknochen getrennt. Einige Experten halten Gorgosaurus libratus für eine Albertosaurus-Art – nach dieser Ansicht wäre Gorgosaurus ein juveniles Synonym dieser Gattung.
Gorgosaurus lebte in einem üppigen Überschwemmungsgebiet entlang des Western Interior Seaway, einem Meerarm, der in der Oberen Kreide Nordamerika in zwei Hälften teilte. Als ein Spitzenprädator stand Gorgosaurus an der Spitze der Nahrungskette und jagte vermutlich die häufigen Ceratopsiden und Hadrosauriden. In einigen Gebieten lebte Gorgosaurus zusammen mit Daspletosaurus, einem anderen Tyrannosauriden. Obwohl diese Tiere etwa gleich groß waren, gibt es Hinweise darauf, dass beide Gattungen unterschiedliche ökologische Nischen besetzten. Gorgosaurus ist der am häufigsten gefundene Tyrannosauride – die zahlreichen Fossilien erlauben es Wissenschaftlern, Rückschlüsse auf die Biologie von Tyrannosauriden zu ziehen, zum Beispiel auf die Individualentwicklung (Ontogenese).
Beschreibung
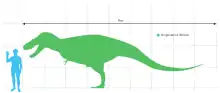
Gorgosaurus erreichte etwa die Größe von Albertosaurus und Daspletosaurus. Ausgewachsene Tiere wurden zwischen acht und neun Meter lang[2][3] und wogen vermutlich mehr als 2,4 Tonnen.[4] Der größte gefundene Schädel misst 99 Zentimeter Länge und ist damit nur geringfügig kleiner als der von Daspletosaurus, einem Tyrannosauriden der zur gleichen Zeit im gleichen Gebiet lebte.[2] Wie bei anderen Tyrannosauriden war der Schädel im Verhältnis zum Körper groß, wobei Kammern in den Schädelknochen und große Schädelöffnungen (Schädelfenster) sein Gewicht reduzierten. Albertosaurus und Gorgosaurus haben im Verhältnis längere und flachere Schädel als Daspletosaurus und andere Tyrannosauriden. Das Schnauzenende war stumpf und das paarige Nasenbein (Nasale) sowie das paarige Scheitelbein (Parietale) war, entlang der Mittellinie des Schädels verschmolzen, wie bei allen anderen Vertretern der Familie. Die Augenhöhle (Orbitalfenster) war rund und unterschied sich damit von den ovalen bis schlüsselloch-artigen Formen bei anderen Tyrannosauriden. Ähnlich wie bei Albertosaurus und Daspletosaurus erstreckt sich vor jedem Auge ein hoher Kamm aus dem Tränenbein (Lacrimale).[3] Gorgosaurus wird von Albertosaurus anhand von Unterschieden an den Knochen abgegrenzt, die das Gehirn umgeben.[5]
Die Zähne von Gorgosaurus waren typisch für alle bekannten Tyrannosauriden. Die acht Zähne des Zwischenkieferbeins (Prämaxillare) am vorderen Schnauzenende waren im Vergleich zu den restlichen Zähnen klein, dicht gepackt und im Querschnitt „D“-förmig. Die restlichen Zähne waren im Querschnitt oval, und nicht klingenförmig wie bei den meisten anderen Theropoden.[3] Neben den acht Zähnen des Zwischenkieferbeins hatte Gorgosaurus 26 bis 30 Zähne im Oberkiefer und 30 bis 34 Zähne im Unterkiefer. Gorgosaurus hat damit etwa so viele Zähne wie Albertosaurus und Daspletosaurus, aber weniger als Tarbosaurus oder Tyrannosaurus.[6]
Wie bei allen Tyrannosauriden ist der Bauplan von Gorgosaurus durch einen großen Kopf gekennzeichnet, der auf einen „S“-förmigen Hals sitzt. Im Kontrast zum großen Kopf waren die Arme sehr klein. Die Hände hatten lediglich zwei Finger und Klauen, obwohl ein drittes Glied in der Mittelhand bei einigen Funden nachgewiesen wurde – ein Rudiment des dritten Fingers, wie ihn andere Theropoden besaßen. Die Hinterbeine der Tyrannosauriden waren im Verhältnis zum Körper länger als bei anderen Theropoden.[3] Der längste von Gorgosaurus bekannte Oberschenkelknochen (Femur) misst 105 Zentimeter. Bei verschiedenen kleineren Exemplaren war jedoch das Schienbein (Tibia) länger als das Femur – ein Verhältnis, wie man es häufig bei schnell laufenden Tieren findet.[2] Die beiden Knochen waren bei den größten gefundenen Skeletten etwa gleich lang.[7]
Nach der Erstbeschreibung von Yutyrannus 2012, einer großen und sehr nah verwandten Art, die vollständig befiedert war, ist man sich sicher, dass auch Gorgosaurus in einen weichen Flaum aus Daunenfedern gekleidet war.
Systematik

Innerhalb der Tyrannosauridae wird Gorgosaurus zu der Unterfamilie Albertosaurinae gezählt – zusammen mit dem nahverwandten, geologisch etwas jüngeren Albertosaurus.[6] Diese beiden Gattungen sind die einzigen definitiven Vertreter der Albertosaurinae, die bisher beschrieben wurden – obwohl möglicherweise weitere, noch unbeschriebene Gattungen existieren.[5] Alle anderen Tyrannosauriden werden in der zweiten Unterfamilie klassifiziert, der Tyrannosaurinae. Verglichen mit den Tyrannosaurinen zeichnen sich Albertosaurinen durch einen schlankeren Körperbau mit proportional kleineren, flacheren Schädeln und längeren Unterbein- und Fußknochen aus.[6][8]
Die deutlichen Ähnlichkeiten zwischen Gorgosaurus libratus und Albertosaurus sarcophagus führten dazu, dass viele Experten vorschlugen, beide Gattungen zu vereinen – so wurde Gorgosaurus manchmal als ein juveniles Synonym von Albertosaurus erkannt. Albertosaurus wurde vor Gorgosaurus benannt und wäre damit der gültige Name, falls es sich um ein und dieselbe Gattung handelt. Die Paläontologen William Diller Matthew und Barnum Brown bezweifelten die Unterscheidung zweier Gattungen schon im Jahr 1922.[9] Gorgosaurus libratus wurde von Dale Russell (1970) formell dem Albertosaurus (als Albertosaurus libratus) neu zugeordnet, und viele Autoren folgten dieser Annahme.[10][11] Wäre Gorgosaurus tatsächlich eine Unterart von Albertosaurus, würde das die geografische und chronologische Verbreitung von Letzterem stark erweitern. Andere Experten unterscheiden zwischen den beiden Gattungen.[3] So merkt der kanadische Paläontologe Philip J. Currie an, dass es so viele anatomische Unterschiede zwischen Albertosaurus und Gorgosaurus gibt wie zwischen Daspletosaurus und Tyrannosaurus, welche fast immer als selbstständige Gattungen aufgeführt werden. Weiter bemerkt er, dass noch unbeschriebene Tyrannosauriden aus Alaska, New Mexico und anderen Fundorten in Nordamerika zur Klärung der Situation beitragen könnten.[5]
Entdeckungsgeschichte und Benennung

Gorgosaurus libratus wurde erstmals von Lawrence Lambe im Jahr 1914 beschrieben.[12][13] Der Name ist von den griechischen Wörtern γοργός gorgos („ungestüm“, „schrecklich“) und σαῦρος sauros („Echse“) abgeleitet.[14] Das Artepitheth libratus ist das Partizip des lateinischen Verbs librare, was „balancieren“ bedeutet.[13]
Der Holotyp von Gorgosaurus libratus (NMC 2120) ist ein nahezu vollständiges, mit einem Schädel verbundenes Skelett, das im Jahr 1913 von Charles Sternberg entdeckt wurde. Dieses Skelett war der erste Fund eines Tyrannosauriden, bei dem die Hand vollständig erhalten geblieben ist.[12] Der Fund wurde in der Dinosaur-Park-Formation von Alberta gemacht und befindet sich heute im Canadian Museum of Nature in Ottawa.[5] Sammler des American Museum of Natural History in New York City haben entlang des Red Deer Rivers in Alberta zur gleichen Zeit hunderte spektakuläre Dinosaurierfunde entdeckt, einschließlich vier vollständige Schädel von Gorgosaurus libratus, von denen drei mit Skeletten verbunden waren. Matthew und Brown beschrieben vier dieser Funde im Jahr 1923.[7]
Des Weiteren beschrieben Matthew und Brown ein fünftes Skelett (AMNH 5664), das Sternberg im Jahr 1917 gesammelt und dann an das American Museum of Natural History verkauft hatte. Es ähnelte anderen Gorgosaurus-Skeletten, hatte aber einen flacheren, leichteren Schädel und längere Bein-Proportionen. Auch waren viele Näthe zwischen den Knochen nicht verschmolzen. Zwar bemerkten Matthew und Brown, dass diese Merkmale für juvenile Tiere charakteristisch sind, trotzdem beschrieben sie anhand dieses Skeletts die neue Art Gorgosaurus sternbergi.[7] Heutige Paläontologen sehen dieses Skelett jedenfalls als ein juveniles Exemplar von Gorgosaurus libratus.[3][5] Dutzende weitere Funde wurden aus der Dinosaur-Park-Formation ausgegraben und sind heute in verschiedenen Museen in den Vereinigten Staaten und Kanada ausgestellt.[2][5] Gorgosaurus libratus ist damit der in der Fossilüberlieferung am häufigsten auftauchende Tyrannosauride. Die Funde decken nahezu sämtliche Altersstufen ab.[3][15]
Im Jahr 1856 beschrieb Joseph Leidy zwei Tyrannosauriden-Zähne des Zwischenkieferbeins (Prämaxillare) aus Montana. Obwohl es keine Hinweise auf das Aussehen des Tieres gab, waren die Zähne groß und robust, sodass Leidy ihnen den Namen Deinodon gab.[16] Matthew und Brown schrieben im Jahr 1922, dass diese Zähne von denen des Gorgosaurus nicht unterscheidbar sind – da zusätzliches Skelettmaterial jedoch nicht vorhanden war, vermieden es die Forscher, Deinodon als ein Synonym mit Gorgosaurus zu klassifizieren.[9] Obwohl die Deinodon-Zähne den Zähnen von Gorgosaurus sehr ähnlich sehen, ist die Form von Tyrannosauriden-Zähnen sehr einheitlich, weshalb man nicht mit Sicherheit sagen kann, zu welcher Gattung sie gehörten. Heute gilt Deinodon als Nomen dubium (zweifelhafter Name).[2][15] Verschiedene weitere Tyrannosauriden-Skelette aus der Judith River-Formation von Montana gehörten eventuell zu Gorgosaurus, wobei es jedoch unklar ist, ob diese Funde zu Gorgosaurus libratus oder zu einer neuen Art gehörten.[5] Ein Exemplar aus Montana (TCMI 2001.89.1), das im Children’s Museum of Indianapolis ausgestellt ist, zeigt ernste Verletzungen und Krankheiten: So wurden verheilte Bein-, Rippen- und Wirbelfrakturen entdeckt. Es wurde auch festgestellt, dass das Tier an Osteomyelitis litt, einer infektiösen Entzündung des Knochenmarks, die im Unterkiefer zu permanentem Zahnausfall führte. Ein möglicher Hirntumor war ebenfalls vorhanden.[17][18]
Neu zugeordnete Arten
Verschiedene Arten wurden im 20. Jahrhundert fälschlicherweise dem Gorgosaurus zugeschrieben. Aus jüngeren Gesteinsschichten (spätes Maastrichtium) der Hell-Creek-Formation aus Montana stammt ein vollständiger Schädel eines kleinen Tyrannosauriden (CMNH 7541), den Charles Whitney Gilmore im Jahr 1946 als Gorgosaurus lancensis beschrieb.[19] Diese Spezies wurde im Jahr 1988 von Bob Bakker und anderen in Nanotyrannus umbenannt.[20] Heute glauben die meisten Paläontologen, es handle sich bei Nanotyrannus um ein juveniles Exemplar des Tyrannosaurus rex.[3][15] Evgeny Maleev beschrieb im Jahr 1955 zwei kleine Tyrannosauriden-Skelette (PIN 553-1 and PIN 552-2) aus der Nemegt-Formation der Mongolei als Gorgosaurus lancinator und Gorgosaurus novojilovi.[21] Zwar benannte Kenneth Carpenter 1992 das kleinere Skelett in Maleevosaurus novojilovi um,[22] beide Skelette werden gelten heute jedoch als juvenile Exemplare von Tarbosaurus bataar.[3][15][23]
Paläobiologie
Koexistenz mit Daspletosaurus

In der Dinosaur-Park-Formation lebte Gorgosaurus zusammen mit Daspletosaurus, einer selteneren Tyrannosaurinen-Gattung. Dies ist eines der wenigen Beispiele einer Koexistenz zweier Tyrannosauriden. In modernen Raubtier-Gilden besetzen ähnlich große Raubtiere verschiedene ökologische Nischen, durch die der Konkurrenzkampf begrenzt wird. Inwieweit dies bei den Tyrannosauriern der Dinosaur-Park-Formation der Fall war, ist nicht hinreichend geklärt.[24] Im Jahr 1970 vermutete Dale Russell, der häufigere Gorgosaurus könnte die flinken Hadrosaurier gejagt haben, während der schwerer gebaute Daspletosaurus die selteneren und schwieriger zu jagenden Ceratopsier und Ankylosaurier bevorzugt haben könnte.[2] Jedenfalls sind in einem Daspletosaurus-Skelett (OTM 200) aus der zur selben Zeit abgelagerten Two-Medicine-Formation von Montana die verdauten Überreste eines juvenilen Hadrosauriers in der Bauchhöhle erhalten geblieben.[25]
Anders als bei einigen anderen Dinosauriergruppen war weder Gorgosaurus noch Daspletosaurus in bestimmten Höhenlagen häufiger als der jeweils andere.[24] Trotzdem scheint Gorgosaurus häufiger in den nördlicheren Formationen wie der Dinosaur-Park-Formation vorzukommen, während Daspletosaurus im Süden häufiger ist. Das gleiche Muster zeigt sich bei anderen Dinosauriergruppen; so sind chasmosaurine Ceratopsier und hadrosaurine Hadrosaurier in der Two-Medicine-Formation von Montana und im südwestlichen Nordamerika während des Campaniums häufiger, während die Centrosaurinae und die Lambeosaurinae die nördlicheren Breiten dominierten. Holtz vermutet aufgrund dieses Musters, dass Hadrosaurinen, Chasmosaurinen und Tyrannosaurinen ähnliche Lebensräume bevorzugten. Am Ende des späten Maastrichtiums waren Tyrannosaurinen wie Tyrannosaurus rex, Hadrosaurinen wie Edmontosaurus und Chasmosaurinen wie Triceratops im westlichen Nordamerika weit verbreitet, während Albertosaurinen und Centrosaurinen ausstarben und Lambeosaurinen sehr selten wurden.[3]
Ontogenese und Populationsbiologie

Forscher um Gregory Erickson haben mithilfe der Knochenhistologie bei verschiedenen Tyrannosauriden-Funden berechnet, wie alt die Tiere waren, als sie starben. Dies lässt Rückschlüsse auf die Individualentwicklung (Ontogenese) zu. Wie alle Tyrannosauriden zeigte auch Gorgosaurus ein sehr schnelles, etwa vier Jahre währendes Wachstum, das nach einer sehr langen Juvenilphase eintrat. Während dieser Wachstumsphase erreichte Gorgosaurus maximale Wachstumsrate von 110 Kilogramm pro Jahr. Das ist langsamer als bei Tyrannosaurinen wie Daspletosaurus und Tyrannosaurus, aber vergleichbar mit Albertosaurus. Tyrannosauriden teilten ihren Lebensraum nur mit Theropoden, die deutlich kleiner waren; es fehlen jedoch Prädatoren, die in ihrer Größe zwischen Tyrannosauriden und kleinen Theropoden standen. Da Gorgosaurus und andere Tyrannosauriden etwa die Hälfte ihres Lebens im Juvenilstadium verbrachten, vermuten einige Forscher, dass diese Nische von juvenilen Tyrannosauriden besetzt wurde.[26][3] Belege für diese Vermutung lieferte der Fund eines jungen Gorgosaurus: In seinem Magen fanden sich die Überreste zweier Exemplare der Gattung Citipes. Diese Pflanzenfresser dürften zu klein gewesen sein, um für ausgewachsene Gorgosaurier relevante Beutetiere dargestellt zu haben, wurden von juvenilen Tieren aber ganz offensichtlich bejagt. Das Forschungsteam, das die Überreste des juvenilen Gorgosaurus analysiert hat, schätzt, dass Gorgosaurier erst ab einem Körpergewicht von ca. 600 Kilogramm (welches sie etwa im Alter von elf Jahren erreicht haben dürften) damit begannen, größeren Beutetieren nachzustellen.[27]
Paläoökologie
Alle bekannten Skelette von Gorgosaurus libratus stammen aus der Dinosaur-Park-Formation[5][28], die durch ihre immense Dichte an Dinosaurierfossilien berühmt ist und vor ca. 76 bis 72 Millionen Jahren ablagerte. Zu dieser Zeit war Nordamerika durch einen Meerarm, dem Western Interior Seaway, in zwei Hälften geteilt, während sich im Westen die Rocky Mountains im Zuge der laramischen Gebirgsbildung zu erheben begannen. Große Flüsse ergossen sich von den aufsteigenden Rocky Mountains in den Western Interior Seaway im Osten, wobei sie Sedimente in Überschwemmungsebenen entlang der Küste ablagerten, die unter anderem die heutige Dinosaur-Park-Formation bilden.[29][30] Das Klima war subtropisch und wies periodische Trockenzeiten auf, was unter großen Dinosaurierherden zu Massensterben führte, wie sich an den zahlreichen Bonebeds der Dinosaur-Park-Formation erkennen lässt.[31] Die Vegetation wurde von Koniferen, Farnen, Baumfarnen und Bedecktsamern gebildet. Die Dinosaurierfauna bestand aus großen Herden von Ceratposiern und Hadrosauriden; andere Herbivoren waren Ornithomimosaurier, Therizinosaurier, Pachycephalosaurier, kleine Ornithopoden sowie Ankylosaurier. Kleine Fleischfresser wie Oviraptorosaurier, Troodontiden und Dromeosauriden jagten kleinere Beute als die großen Tyrannosauriden wie Daspletosaurus und Gorgosaurus.[32][2][3]
Weblinks
- Gorgosaurus auf Palaeos.com (englisch)
Einzelnachweise
- Gregory S. Paul: The Princeton Field Guide To Dinosaurs. Princeton University Press, Princeton NJ 2010, ISBN 978-0-691-13720-9, S. 105, Online (Memento vom 13. Juli 2015 im Internet Archive).
- Dale A. Russell: Tyrannosaurs from the late cretaceous of Western Canada (= Publications in Palaeontology. Bd. 1, ISSN 0068-8029). National Museum of Natural Sciences (Canada), Ottawa 1970, Digitalisat.
- Thomas R. Holtz: Tyrannosauroidea. In: David B. Weishampel, Peter Dodson, Halszka Osmólska (Hrsg.): The Dinosauria. 2nd edition. University of California Press, Berkeley CA u. a. 2004, ISBN 0-520-24209-2, S. 111–136.
- Frank Seebacher: A new method to calculate allometric length-mass relationships of dinosaurs. In: Journal of Vertebrate Paleontology. Bd. 21, Nr. 1, 2001, ISSN 0272-4634, S. 51–60, doi:10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2.
- Philip J. Currie: Cranial anatomy of tyrannosaurid dinosaurs from the Late Cretaceous of Alberta, Canada. In: Acta Palaeontologica Polonica. Bd. 48, Nr. 2, 2003, ISSN 0567-7920, S. 191–226, (PDF; 1,8 MB).
- Philip J. Currie, Jørn H. Hurum, Karol Sabath: Skull structure and evolution in tyrannosaurid dinosaurs. In: Acta Palaeontologica Polonica. Bd. 48, Nr. 2, 2003, S. 227–234, (PDF; 137 kB).
- William D. Matthew, Barnum Brown: Preliminary notices of skeletons and skulls of Deinodontidæ from the Cretaceous of Alberta. In: American Museum Novitates. Nr. 89, 1923, S. 1–9, (PDF; 4,2 MB).
- Philip J. Currie: Allometric growth in tyrannosaurids (Dinosauria: Theropoda) from the Upper Cretaceous of North America and Asia. In: Canadian Journal of Earth Sciences. Bd. 40, Nr. 4, 2003, ISSN 0008-4077, S. 651–665, doi:10.1139/e02-083.
- William D. Matthew, Barnum Brown: The family Deinodontidae, with notice of a new genus from the Cretaceous of Alberta. In: Bulletin of the American Museum of Natural History. Bd. 46, Nr. 6, 1922, ISSN 0003-0090, S. 367–385, (PDF; 1,8 MB).
- Thomas D. Carr, Thomas E. Williamson, David R. Schwimmer: A new genus and species of tyrannosauroid from the Late Cretaceous (middle Campanian) Demopolis Formation of Alabama. In: Journal of Vertebrate Paleontology. Bd. 25, Nr. 1, 2005, S. 119–143, doi:10.1671/0272-4634(2005)025[0119:ANGASO]2.0.CO;2.
- Gregory S. Paul: Predatory Dinosaurs of the World. Simon & Schuster, New York NY u. a. 1988, ISBN 0-671-61946-2.
- Lawrence M. Lambe: On the fore-limb of a carnivorous dinosaur from the Belly River Formation of Alberta, and a new genus of Ceratopsia from the same horizon, with remarks on the integument of some Cretaceous herbivorous dinosaurs. In: Ottawa Naturalist. Bd. 27, Nr. 10, 1914, S. 129–135, Digitalisat.
- Lawrence M. Lambe: On a new genus and species of carnivorous dinosaur from the Belly River Formation of Alberta, with a description of Stephanosaurus marginatus from the same horizon. In: Ottawa Naturalist. Bd. 28, Nr. 1, 1914, S. 13–20, Digitalisat.
- Henry George Liddell, Robert Scott: A Lexicon abridged from Liddell and Scott's Greek-English Lexicon. 24th edition, carefully revised throughout. Ginn, Boston 1891 (Reprinted edition. Clarendon Press, Oxford 2010, ISBN 978-0-19-910207-5).
- Thomas D. Carr: Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Coelurosauria). In: Journal of Vertebrate Paleontology. Bd. 19, Nr. 3, 1999, S. 497–520, doi:10.1080/02724634.1999.10011161.
- Joseph Leidy: Notice of remains of extinct reptiles and fishes, discovered by Dr. F.V. Hayden in the badlands of the Judith River, Nebraska Territory. In: Proceedings of the Academy of Natural Sciences of Philadelphia. Bd. 8, 1956, ISSN 0097-3157, S. 72–73, Digitalisat.
- John Pickrell: First dinosaur brain tumor found, experts suggest. In: National Geographic News. 24. November 2003, abgerufen am 24. Juli 2014 (englisch).
- Meet the Gorgosaur. In: The Children's Museum of Indianapolis. Archiviert vom am 25. Februar 2014; abgerufen am 24. Juli 2014.
- Charles W. Gilmore: A new carnivorous dinosaur from the Lance Formation of Montana (= Smithsonian Miscellaneous Collections. Bd. 106, Nr. 13, ISSN 0096-8749 = Smithsonian Institution. Publication. 3857). Smithsonian Institution, Washington DC 1946, Digitalisat.
- Robert T. Bakker, Michael Williams, Philip J. Currie: Nanotyrannus, a new genus of pygmy tyrannosaur, from the latest Cretaceous of Montana. In: Hunteria. Bd. 1, Nr. 5, 1988, ZDB-ID 1251702-1, S. 1–30.
- Евгений А. Малеев: Новый хищный динозавр из верхнего мела Монголии. In: Доклады Академии наук СССР. Bd. 104, Nr. 5, 1955, ISSN 0002-3264, S. 779–783 (In englischer Sprache: New carnivorous dinosaurs from the Upper Cretaceous of Mongolia. online (PDF; 12,41 kB)).
- Ken Carpenter: Tyrannosaurids (Dinosauria) of Asia and North America. In: Niall J. Mateer, Chen Pei-ji (Hrsg.): Aspects of Nonmarine Cretaceous Geology. China Ocean Press, Beijing 1992, ISBN 7-5027-1463-4, S. 250–268.
- Анатолий К. Рождественский: Возрастная изменчивость и некоторые вопросы систематики динозавров Азии. In: Палеонтологический Журнал. Nr. 3, 1965, ISSN 0031-031X, S. 95–109.
- James O. Farlow, Eric R. Pianka: Body size overlap, habitat partitioning and living space requirements of terrestrial vertebrate predators: implications for the paleoecology of large theropod dinosaurs. In: Historical Biology. Bd. 16, Nr. 1, 2002, ISSN 0891-2963, S. 21–40, doi:10.1080/0891296031000154687.
- David J. Varricchio: Gut contents from a Cretaceous tyrannosaurid: implications for theropod dinosaur digestive tracts. In: Journal of Paleontology. Bd. 75, Nr. 2, 2001, ISSN 0022-3360, S. 401–406, doi:10.1666/0022-3360(2001)075<0401:GCFACT>2.0.CO;2.
- Gregory M. Erickson, Peter J. Makovicky, Philip J. Currie, Mark A. Norell, Scott A. Yerby, Christopher A. Brochu: Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs. In: Nature. Bd. 430, Nr. 7001, 2004, S. 772–775, doi:10.1038/nature02699.
- François Therrien, Darla K. Zelenitsky, Kohei Tanaka, Jared T. Voris, Gregory M. Erickson, Philip J. Currie, Christopher L. Debuhr, Yoshitsugu Kobayashi: Exceptionally preserved stomach contents of a young tyrannosaurid reveal an ontogenetic dietary shift in an iconic extinct predator. In: Science Advances. Band 9, Nr. 49, 2023, doi:10.1126/sciadv.adi0505.
- David A. Eberth, Anthony P. Hamblin: Tectonic, stratigraphic, and sedimentologic significance of a regional discontinuity in the upper Judith River Group (Belly River wedge) of southern Alberta, Saskatchewan, and northern Montana. In: Canadian Journal of Earth Sciences. Bd. 30, Nr. 1, 1993, S. 174–200, doi:10.1139/e93-016.
- Joseph M. English, Stephen T. Johnston: The Laramide Orogeny: what were the driving forces? In: International Geology Review. Bd. 46, Nr. 9, 2004, ISSN 0020-6814, S. 833–838, doi:10.2747/0020-6814.46.9.833.
- David A. Eberth: Judith River Wedge. In: Philip J. Currie, Kevin Padian (Hrsg.): Encyclopedia of Dinosaurs. Academic Press, San Diego CA u. a. 1997, ISBN 0-12-226810-5, S. 199–204.
- Dennis R. Braman, Eva B. Koppelhus: Campanian palynomorphs. In: Phillip J. Currie, Eva B. Koppelhus (Hrsg.): Dinosaur Provincial Park. A Spectacular Ancient Ecosystem Revealed. Indiana University Press, Bloomington IN u. a. 2005, ISBN 0-253-34595-2, 101–130.
- James O. Farlow: Speculations about the diet and foraging behavior of large carnivorous dinosaurs. In: American Midland Naturalist. Bd. 95, Nr. 1, 1976, ISSN 0003-0031, S. 186–191, doi:10.2307/2424244.