Geschichte der Katecholaminforschung
Zu den Katecholaminen (bzw. Catecholaminen) oder Brenzcatechinaminen gehören die körpereigenen Stoffe Dopamin, Noradrenalin (Norepinephrin) und Adrenalin (Epinephrin) sowie zahlreiche künstlich hergestellte Substanzen, unter denen Isoprenalin hervorzuheben ist. Ihre Erforschung bildet ein wichtiges Kapitel der Geschichte der Physiologie, Biochemie und Pharmakologie. Adrenalin war das erste aus einer endokrinen Drüse extrahierte, das erste rein dargestellte und das erste in seiner Struktur und Biosynthese geklärte Hormon, vor der Prägung des Wortes „Hormon“.[1] Adrenalin und Noradrenalin waren neben Acetylcholin die erstentdeckten Neurotransmitter und die ersten Botenstoffe, die in intrazellulären Vesikeln gespeichert gefunden wurden. Der β-Adrenozeptor war der erste G-Protein-gekoppelte Hormon- und Neurotransmitterrezeptor, dessen Gen kloniert wurde.
Die zielgerichtete Katecholaminforschung begann mit der Herstellung eines pharmakologisch wirksamen Extraktes aus Nebennieren durch George Oliver und Edward Albert Schäfer in den Jahren 1893 und 1894. Doch gab es frühere Hinweise.
Adrenalin
Frühe Hinweise auf einen Stoff im Nebennierenmark
In das bekannteste Buch des 19. Jahrhunderts über Asthma nahm Henry Hyde Salter (1823–1871) ein Kapitel „Behandlung mit Stimulantien“ auf.[2] Starker Kaffee half danach bei Asthmaanfällen. Noch besser wirkte starke psychische Erregung. „Die Heilung des Asthmas durch heftige Erregung erfolgt schneller und vollständiger als durch jedes andere Mittel. … Sie geschieht augenblicklich, der schlimmste Paroxysmus löst sich im nu.“ Salter belegte das mit Krankengeschichten. Zweifellos war eine Ausschüttung von Adrenalin aus den Nebennieren der therapeutische Mechanismus.
Salter wusste von diesem Mechanismus nichts. Zur gleichen Zeit bemerkte der französische Arzt Alfred Vulpian, dass das Nebennierenmark eine einzigartige Eigenschaft hatte: Es färbte sich beim Kontakt mit Eisen(III)-chlorid grün, anders als die Nebennierenrinde oder jedes andere Organ. Das Nebennierenmark enthielt also „eine besondere, bisher unbekannte Substanz“. Vulpian erkannte auch, dass die Substanz in den Blutkreislauf gelangte, denn das Blut in den Nebennierenvenen gab die Eisen(III)-Reaktion.[3]

Zu dem klinischen und chemischen Hinweis kam ein physiologischer. In Versuchen des deutschen Pharmakologe Carl Jacobj unterdrückte elektrische Reizung der Nebennieren die Peristaltik des Darms.[4] Man hat dies als den „ersten indirekten Nachweis der Funktion des Nebennierenmarks als einer endokrinen Drüse“ bezeichnet.[5] Jedoch dachte Jacobj nicht an ein chemisches Signal aus den Nebennieren, also ein Hormon, sondern an eine Nervenverbindung von den Nebennieren zum Darm, „Hemmungsbahnen für die Darmbewegung“.
Oliver und Schäfer 1893/94
Die Entdeckungsgeschichte hat 1938 und 1948 Henry Hallett Dale erzählt.[6][7] Danach injizierte der praktizierende Arzt Oliver seinem Sohn Extrakte verschiedener Organe subkutan und maß dabei mit einem von ihm selbst konstruierten Apparat den Durchmesser der Arteria radialis. Ein Nebennierenextrakt verengerte die Arterie. Oliver bat darauf den Physiologen Schäfer am University College London um eine Prüfung der Blutdruckwirkung im Tierversuch. Schäfer war skeptisch, fand dann aber, dass bei einem narkotisierten Hund nach intravenöser Injektion das Quecksilber im Manometer „rasant auf eine ungeahnte Höhe stieg. So wurde das aktive Prinzip der Nebenniere entdeckt, das später als Inhaltsstoff ausschließlich des Nebennierenmarks erkannt und noch später rein, kristallin dargestellt und ‚Epinephrin‘ oder ‚Adrenalin‘ genannt wurde.“
Oliver experimentierte im Sinne der zeitgenössischen Vorstellung einer „Organtherapie“, nach der Organe wirkkräftige Substanzen enthielten, deren therapeutischen Nutzen es herauszufinden galt.[8] Er schritt denn auch gleich weiter zur Extraktion der Hypophyse, die zur Entdeckung – wieder zusammen mit Schäfer – des blutdrucksteigernden antidiuretischen Hormons oder Vasopressins führte.[9] Dale kannte die Entdeckungsgeschichte des Adrenalins aus der Tradition am University College London, wo er selbst und vor ihm Schäfer gearbeitet hatte. Sein Bericht von subkutanen Injektionen Olivers widerspricht Mitteilungen von Oliver und Schäfer selbst, nach denen Oliver die Extrakte oral verabreichte.[10][11] Wirksamkeit von Adrenalin nach oraler Gabe ist sehr unwahrscheinlich.[12] Olivers Nachkommen wussten später nichts mehr von Experimenten an dem Sohn.[13] Teile der Überlieferung sind wohl Legende.

Jedenfalls verursachten Oliver und Schäfers Berichte im Journal of Physiology 1894 eine Sensation. Oliver versuchte seine Extrakte gleich auch therapeutisch, und zwar recht willkürlich, von der Addisonschen Krankheit („Addison’s disease“) über arterielle Hypotonie („loss of vasomotor tone“), Diabetes mellitus und Diabetes insipidus bis zum endokrinen Exophthalmus („exophthalmic goitre“).[10] 1903 begann die Anwendung des – inzwischen gereinigten – Adrenalins beim Asthma.[14] Die New Yorker Ärzte versuchten es nicht wegen der bronchospasmolytischen Wirkung, die erst später entdeckt wurde,[15][16] sondern wegen der Vasokonstriktion, von der man sich eine Abschwellung der Bronchialschleimhaut erhoffte. Im selben Jahr begann der Zusatz zu Lösungen von Lokalanästhetika. 1905 zeigte der Leipziger Chirurg Heinrich Braun, dass Adrenalin deren Wirkung am Injektionsort verlängerte, während es zugleich die „systemischen“ Wirkungen, also Wirkungen außerhalb der Injektionsstelle, abschwächte (siehe Geschichte der zahnärztlichen Lokalanästhesie).[17]
Unabhängige weitere Entdecker
Ein Jahr nach Oliver und Schäfer berichteten Władysław Szymonowicz (1869–1939) und Napoleon Cybulski von der Jagiellonen-Universität in Kraków über ähnliche Tierexperimente.[18] Auch das Blut der Nebennierenvenen enthielt die blutdrucksteigernde Substanz. Diese trat also in den Kreislauf ein, wie bereits Vulpian aus der Eisen(III)-Reaktion gefolgert hatte.
Ein weiteres Jahr später, also 1896, referierte der US-amerikanische Arzt William Bates über zweijährige Erfahrungen mit der Einträufelung von Nebennierenextrakten in den Bindehautsack des Auges. Die Bindehaut wurde innerhalb weniger Minuten weiß, „whitened in a few minutes“, durch Vasokonstriktion, wie er richtig feststellte.[19] Er hat im Gegensatz zu Oliver und Schäfer sowie Szymonowicz und Cybulski kaum Eingang in die Forschungsliteratur gefunden.[20] Selbst ein augenheilkundlicher Übersichtsartikel von 1905 erwähnt ihn nur beiläufig.[21]
Isolierung, Strukturaufklärung und Synthese
John Jacob Abel in Baltimore reinigte Nebennierenextrakte 1899 unvollständig zum „Epinephrin“,[22] Otto von Fürth in Straßburg 1900 zum „Suprarenin“.[23] Die Reindarstellung gelang 1901 dem Japaner Jokochi Takamine, der in New York ein eigenes Laboratorium eingerichtet hatte.[24] Er ließ sein Produkt, das er „Adrenalin“ nannte,[25] patentieren und von der Firma Parke, Davis & Co., heute aufgegangen in Pfizer Inc., als „Adrenalin“ auf den Markt bringen.[26] 1903 wurde Adrenalin als optisch aktiv und linksdrehend erkannt. 1905 synthetisierten Friedrich Stolz bei den Farbwerken Hoechst AG und Henry Drysdale Dakin an der University of Leeds das Racemat. 1906 klärte Ernst Joseph Friedmann (1877–1957) in Straßburg die Struktur.[27] 1908 erkannte Arthur Robertson Cushney (1866–1926) an der University of Michigan, dass rechtsdrehendes Adrenalin pharmakologisch fast unwirksam war, und zog den bemerkenswerten Schluss, die „receptive substance affected by adrenaline“ unterscheide zwischen den optischen Isomeren und sei daher selbst optisch aktiv.[28] Insgesamt erhielt Adrenalin im Laufe der Zeit 32 Namen, von denen die Wissenschaft „Adrenalin“ – vorzugsweise im Vereinigten Königreich – und „Epinephrin“ – vorzugsweise in den Vereinigten Staaten – benutzt.[29]
Entdeckung der Neurotransmitter-Funktion
Ein neues Kapitel begann mit der Feststellung Max Lewandowskys in Berlin 1899, dass Nebennierenextrakt auf die glatte Muskulatur der Augen von Katzen – darunter die Nickhaut und den Musculus dilatator pupillae – ebenso wirkte wie eine Reizung der sympathischen Nerven.[30] John Newport Langley und unter seiner Leitung Thomas Renton Elliott in Cambridge fügten weitere Parallelen hinzu. In einem Vortrag vor der Physiological Society in London am 21. Mai 1904 äußerte Elliott die visionäre Hypothese, Adrenalin wirke auf einen Bestandteil der Muskelzellen an deren Kontaktstelle mit den sympathischen Nervenfasern. Aufgabe dieses Bestandteils sei es, den Nervenimpuls zu empfangen und in eine Antwort der Muskelzellen zu übersetzen. „Adrenalin might then be the chemical stimulant liberated on each occasion when the impulse arrives at the periphery – Adrenalin könnte das chemische Stimulans sein, das jedes Mal freigesetzt wird, wenn ein Nervenimpuls in der Peripherie ankommt.“[31] Die Publikation war die „Geburtsurkunde“ der chemischen Neurotransmission, der chemischen synaptischen Informationsübertragung.[32] Elliott hat sich nie wieder so eindeutig geäußert. Er war wohl enttäuscht von dem Mangel an Resonanz bei den tonangebenden Physiologen, darunter Langley, und zog sich 1906 aus der physiologischen Forschung zurück.
- Die „Geburtsurkunde“ der chemischen Neurotransmission


Der Durchbruch der chemischen synaptischen Übertragung kam 1921 mit Otto Loewis Experimenten an Frosch- und Krötenherzen in Graz und seinem Aufsatz „Über humorale Übertragbarkeit der Herznervenwirkung“.[33] Vagusstoff übertrug die Wirkung des Nervus vagus, Acceleransstoff die Wirkung des Sympathikus auf das Herz.[34] 1926 identifizierte Loewi den Vagusstoff mit Acetylcholin, und 1936 schrieb er: „So stehe ich nicht mehr an, den Sympathicusstoff mit Adrenalin für identisch zu halten.“[35]
Er hatte Glück. Bei Amphibien ist Adrenalin in der Tat der Haupt-Transmitter des (postganglionären) Sympathikus. Die Frage nach dem Transmitter bei Säugern dagegen führte zu Schwierigkeiten. In einer ausführlichen Struktur-Wirkungsanalyse wiesen Dale und der Chemiker George Barger 1910 darauf hin, dass sich nach Elliotts Hypothese die Wirkungen von Adrenalin und sympathischer Nervenreizung genauer decken müssten, als sie es täten. „Amino-ethanol-catechol“, das ist Noradrenalin, ahme den Sympathikus genauer nach.[36] In den 1930er Jahren brachten die Physiologen Walter Bradford Cannon und Arturo Rosenblueth an der Harvard Medical School die Idee zweier Transmitter auf, Sympathin E und Sympathin I. Noradrenalin wurde immer wieder in Erwägung gezogen. Die Frage blieb bis nach dem Zweiten Weltkrieg unbeantwortet. Inzwischen schuf Dale eine Terminologie, die seither das Denken der Neurowissenschaftler prägte: Man solle Nervenzellen nach ihrem Transmitter nennen, also „adrenerg“, wenn der Transmitter „eine Substanz wie Adrenalin – some substance like adrenaline“, und „cholinerg“, wenn er „eine Substanz wie Acetylcholin – some substance like acetylcholine“ wäre.[37] Inzwischen begannen sich auch Biosynthese und Abbau der Katecholamine zu klären.
1936, im Jahr, in dem Loewi sich für Adrenalin als sympathischen Transmitter (bei Amphibien) entschied, erhielten er und Dale den Nobelpreis für Physiologie oder Medizin „für ihre Entdeckungen bei der chemischen Übertragung der Nervenimpulse“.
Noradrenalin
Dank Holtz und Blaschko war bekannt, dass Tiere Noradrenalin synthetisierten. Als Transmitter musste es aber in sympathisch innervierten Geweben gespeichert und nicht nur kurzlebiges Zwischenprodukt sein. Der Nachweis gelang Ulf von Euler, der schon die Prostaglandine entdeckt und Substanz P (mit John Henry Gaddum) mitentdeckt hatte, am Karolinska-Institut in Stockholm. Seine erste Mitteilung ging am 16. April 1946 bei der Zeitschrift Nature ein.[38] Nach vielen weiteren Bioassays und chemischen Tests folgerte er, sympathisch innervierte Gewebe, in kleineren Mengen auch das Gehirn, nicht aber die nervenfreie Placenta, enthielten Noradrenalin und dies sei das „Sympathin“ von Cannon und Rosenblueth, „der physiologische Transmitter der Wirkung adrenerger Nerven bei Säugern“.[39] Abgabe von Noradrenalin ins venöse Blut der Milz von Katzen bei Sympathikusreizung – nachgewiesen an Gaddums Pharmakologischem Institut in Edinburgh – unterstützte zwei Jahre später die Folgerung.[40] Bei Amphibien bestätigte von Euler die Transmitterfunktion des Adrenalins.
Der Krieg verhinderte, dass Holtz und seine Kollegen in Rostock als Mitentdecker des Noradrenalins als eines Neurotransmitters in die Geschichte der Katecholaminforschung eingingen. Sie suchten nach Katecholaminen im menschlichen Harn und fanden blutdrucksteigernde Stoffe, „Urosympathin“, die sie als Gemisch von Dopamin, Noradrenalin und Adrenalin identifizierten. Dopamin sei nur Vorläufer von Noradrenalin und Adrenalin. Dagegen: „Arterenol [Noradrenalin] und Adrenalin werden im Bereich sympathischer Nervenendigungen frei, wenn diese in Erregung geraten.“[41] Das Manuskript ging am 8. Oktober 1944 beim Springer-Verlag in Leipzig ein. Am 15. Oktober wurde die Druckerei des Verlags in Braunschweig durch Bomben zerstört. Die Veröffentlichung verzögerte sich bis Band 204, 1947, von Naunyn-Schmiedebergs Archiv für experimentelle Pathologie und Pharmakologie. Holz pflegte den Aufsatz später als „Holtz et al. 1944/47“ oder „Holtz, Credner und Kroneberg 1944/47“ zu zitieren.
Rückblickend schrieb Dale 1953, er habe 1910 mit Barger eigentlich sehen müssen, dass Noradrenalin der Haupt-Transmitter, Elliotts Theorie also im Prinzip richtig und nur in diesem Detail falsch war. Es sei kein Ruhmesblatt, der Wahrheit so nah gekommen zu sein und dann angehalten zu haben.[42]

Marthe Vogt, die Deutschland 1935 verlassen hatte, tat 1954 in Gaddums Institut den Schritt vom peripheren Sympathikus zum Zentralnervensystem.[43] Es enthielt hauptsächlich Noradrenalin, daneben etwas Adrenalin; Vogt fasste die beiden als „Sympathin“ zusammen. Waren sie nur Transmitter der sympathischen Nerven der Blutgefäße des ZNS oder spielten sie in den Nervenzellen selbst eine Rolle? Die verschiedenen Areale des ZNS enthielten sehr verschiedene Konzentrationen an „Sympathin“, und die Verteilung ließ sich nicht mit einer unterschiedlichen Dichte der Blutgefäße erklären. Zudem blieben die Konzentrationen nach Zerstörung der sympathischen Nerven gleich. „Es liegt also nah, dem Sympathin im Gehirn eine Transmitterfunktion ähnlich dem Sympathin in den postganglionären sympathischen Nerven zuzuschreiben.“[44] Sie hatte recht. Mit der 1962 von Nils-Åke Hillarp (1916–1965) und Bengt Falck (* 1927) in Schweden entwickelten Formaldehyd-Fluoreszenzmethode und mit Immunhistochemie-Methoden sind später die Noradrenalin-, Adrenalin- und Dopamin-Bahnen im Zentralnervensystem direkt mikroskopisch sichtbar gemacht worden.
Dopamin
Die Noradrenalingeschichte wiederholte sich. Wie Noradrenalin auf dem Biosynthesepfad zum Adrenalin liegt Dopamin auf dem Pfad zum Noradrenalin (und damit Adrenalin). War es, anders als Peter Holtz vermutet hatte, doch ein eigenständiger Botenstoff? Arvid Carlsson am Pharmakologischen Institut der Universität Lund und seine studentischen Mitarbeiter Åke Bertler and Evald Rosengren fanden 1958/59 nach dem Muster von Marthe Vogt Dopamin im Gehirn ungleichmäßig verteilt, und zwar anders als Noradrenalin. Das sprach für eine Funktion über die eines Intermediärprodukts hinaus. Die höchste Konzentration enthielt das Corpus striatum, wo es nur Spuren von Noradrenalin gab. Carlssons Gruppe hatte zuvor gefunden, dass das Alkaloid Reserpin, das bei Menschen und Versuchstieren ein Parkinson-Syndrom auslöste, die Dopaminspeicher (sowie die Noradrenalin- und Serotoninspeicher) im Gehirn entleerte. So brachte die Gruppe das Corpus striatum, den Reserpin-Parkinsonismus, den menschlichen Morbus Parkinson und Dopamin zum ersten Mal in einen Zusammenhang.[45][46] Ein Jahr später sah Oleh Hornykiewicz, den Hermann Blaschko zum Dopamin geführt hatte, als er im Pharmakologischen Institut der Universität Wien mit Extrakten des Corpus striatum eine Farbreaktion durchführte, den Dopaminmangel beim menschlichen Morbus Parkinson „mit seinen eigenen bloßen Augen: Statt der Rosafärbung durch die verhältnismäßig hohen Dopaminkonzentrationen der Kontrollproben zeigten die Reagenzgläser mit den Proben von Parkinsonkranken kaum einen Hauch von Rosa.“[47][48]
1970 waren von Euler und Axelrod zwei der drei Gewinner des Nobelpreises für Physiologie oder Medizin „für ihre Entdeckungen zu den chemischen Transmittern in Nervenendigungen und zum Mechanismus ihrer Speicherung, Freisetzung und Inaktivierung“, und im Jahr 2000 war Carlsson einer von drei Gewinnern, die den Preis „für ihre Entdeckungen zur Signalübersetzung im Nervensystem“ erhielten.
Biosynthese und Abbau
Der Jude Hermann Blaschko, der Deutschland 1933 verlassen hatte, schrieb 1987 in Oxford:[49] „Unsere heutige Kenntnis der Biosynthese der Katecholamine begann 1939 mit einer Veröffentlichung von Peter Holtz und seinen Kollegen. Sie beschrieben ein Enzym in der Niere von Meerschweinchen, das sie DOPA-Decarboxylase nannten, weil es die Entstehung von Dopamin und Kohlendioxid aus der Aminosäure L-DOPA katalysierte.“ Die Publikation von Holtz und Mitarbeitern stammte aus dem Pharmakologischen Institut der Universität Rostock.[50] Blaschkos eigene Forschung in den 1930er Jahren – er war damals in Cambridge – stand der von Peter Holtz nah. Noch im selben Jahr 1939 sagten sowohl er als auch Holtz den kompletten Biosyntheseweg Tyrosin → L-DOPA → Dopamin → Noradrenalin → Adrenalin vorher.[51][52] Den letzten Schritt, die Methylierung von Noradrenalin zu Adrenalin, wies Edith Bülbring, die ebenfalls vor dem Nationalsozialismus nach England geflohen war, 1949 in Oxford nach,[53] und das verantwortliche Enzym Phenylethanolamin-N-Methyltransferase reinigte Julius Axelrod 1962 in Bethesda.[54] Die beiden noch fehlenden Biosynthese-Enzyme, Tyrosinhydroxylase und Dopamin β-Hydroxylase, wurden ebenfalls um 1960 charakterisiert.
Schon 1937, vor der Formulierung der Biosynthese, hatte Blaschko eine Abbaumöglichkeit erkannt: Eine 1928 gefundene „Tyraminoxidase“[55] oxidierte auch Dopamin, Noradrenalin und Adrenalin.[56] Sie wurde dann Monoaminoxidase genannt. Doch schrieb Blaschko 1956, die Oxidation scheine ihm zu langsam, es müsse andere Inaktivierungsvorgänge geben. „Hier besteht eine Lücke in unserem Wissen.“[57] Innerhalb eines Jahres verkleinerte Axelrod die Lücke durch die Entdeckung der Catechol-O-Methyltransferase.[58] Damit waren auch die Abbauenzyme vollständig. Zur Schließung der Lücke, zum wirklichen Verständnis des Schicksals freigesetzter Katecholamine, bedurfte es aber der Entdeckung der Rolle von Membranen.
Die Rolle von Membranen
Membranen haben für Katecholamine eine doppelte Bedeutung: Sie müssen Membranen durchqueren und müssen an Membranen ihre chemische Botschaft weitergeben.
Membranpassage
Die Katecholamine werden intrazellulär synthetisiert und in intrazellulären membranumhüllten Vesikeln gespeichert. Das wurde zuerst, 1953, dank Blaschko und Arnold Welch (1908–2003) in Oxford sowie Hillarp und seiner Gruppe in Lund für das Nebennierenmark[59][60] bekannt und später – hier handelt es sich um synaptische Vesikel – für sympathische Nerven[61] und die Katecholamin-Neurone des Gehirns.[62] Zusätzlich enthielten die Vesikel Adenosintriphosphat, in den sympathischen Nerven der Milz von Rindern nach Hans-Joachim Schümann und Horst Grobecker (1934–2019) im Peter Holtzschen Pharmakologischen Institut der Universität Frankfurt am Main in einem molaren Noradrenalin:ATP-Verhältnis von 5,2:1.[63] Blaschko und Welch fragten: „Was geschieht in den Zellen, wenn Nervenimpulse ankommen?“ Die Antwort „Exozytose“ zogen sie nicht in Betracht. Sie zu geben und ihr Anerkennung zu verschaffen, bedurfte es der Analogie der „quantalen Freisetzung“ von Acetylcholin in der motorischen Endplatte, die Bernard Katz entdeckt hatte, der dritte Gewinner des 1970er Nobelpreises für Physiologe oder Medizin; des Nachweises der gemeinsamen Freisetzung von Katecholaminen und anderen Vesikelinhaltsstoffen wie ATP; und der eindeutigen elektronenmikroskopischen Bilder des Verschmelzens von Vesikeln mit der Zellmembran.[64]
Acetylcholin wird nach seiner Freisetzung extrazellulär abgebaut, denn die Acetylcholinesterase wendet ihr aktives Zentrum dem Extrazellularraum zu. Anders die Katecholamine. Wie die Syntheseenzyme befinden sich auch Monoaminoxidase und Catechol-O-Methyltransferase intrazellulär. Nicht Abbau, sondern Aufnahme in Zellen ist der primäre Inaktivierungsschritt. Ab 1959 erkannte man ihn. Axelrod und seine Mitarbeiter injizierten Katzen 3H-Adrenalin und 3H-Noradrenalin von hoher spezifischer Radioaktivität intravenös. Ein Teil wurde zu 3H-Metanephrin und 3H-Normetanephrin O-methyliert; ein anderer Teil aber wurde in Zellen aufgenommen und dort unmetabolisiert gespeichert. Zum selben Ergebnis kam auf anderem Wege Erich Muscholl in Mainz, der bei Marthe Vogt in Edinburgh gearbeitet hatte. Er wollte wissen, wie Cocain Organe für Katecholamine sensibilisierte – eine von Loewi und Alfred Fröhlich 1910 in Wien entdeckte Fundamentalwirkung des Cocains.[65] Ratten nahmen intravenös infundiertes Noradrenalin unverändert in Herz und Milz auf; Cocain verhinderte die Aufnahme „und erhöhte so die Menge an Noradrenalin, die mit den Adrenozeptoren reagieren konnte“. Es waren hauptsächlich sympathische Nervenzellen, die 3H-Noradrenalin aufnahmen, denn Zerstörung des Sympathikus verminderte die Aufnahme. Zudem wurde aufgenommenes 3H-Noradrenalin, wie Axelrod und Georg Hertting elegant zeigten, bei Sympathikusreizung wieder freigesetzt.[66] Jedoch fand wenige Jahre später Leslie Iversen in Cambridge, dass auch andere Zellen Katecholamine aufnahmen. Er nannte die Aufnahme in noradrenerge Neurone, die durch Cocain blockiert wurde, „uptake1“ und Aufnahme in andere Zellen, die durch Cocain nicht blockiert wurde, „uptake2“. Mit der Aufnahme in die Speichervesikel, die durch Reserpin blockiert wurde, gab es also drei Membranpassage-Mechanismen für Katecholamine. Iversens Buch 1967 „The Uptake and Storage of Noradrenaline in Sympathetic Nerves“ hatte großen Erfolg, Zeichen der Faszination der Forscher durch die Membrantransporter und ihre reiche Pharmakologie.[67] „Metabolisierende Systeme“ nannte der Würzburger Pharmakologe Ullrich Trendelenburg in den 1980er Jahren biologische Konstruktionen, bei denen zur Inaktivierung extrazellulärer Stoffe ein Membrantransporter und ein intrazelluläres Enzym Hand in Hand arbeiten.[68]
Mit Hilfe der molekularen Genetik sind seit 1990 die Transporter bis zu ihren Genen und ihrer Proteinstruktur verfolgt worden. Sie umfassen den Zellmembrantransporter NAT oder NET, den klassischen uptake1, und den analogen Dopamintransporter DAT; den „extraneuronalen Monoamintransporter“ EMT, auch „organic cation transporter 3“ genannt, aus der Gruppe der SLC-Transporter, Iversens uptake2; und den vesikulären Monoamintransporter VMAT mit zwei Isoformen, VMAT1 und VMAT2.
Rezeptoren

Forschungen über die Katecholamine und Forschungen über ihre Rezeptoren waren miteinander verwoben. 1904 wurde Dale Leiter der Wellcome Physiological Research Laboratories in London und begann mit der Untersuchung von Mutterkornextrakten. 1906 publizierte er „On some physiological actions of ergot“.[69] Der Aufsatz ist weniger wichtig wegen der Wirkung der Extrakte allein als wegen ihrer Wechselwirkung mit Adrenalin: Sie verkehrten dessen normale blutdrucksteigernde Wirkung in eine Blutdrucksenkung und die normale Kontraktionsauslösung am Uterus von frühschwangeren Katzen in eine Erschlaffung: die berühmte Adrenalinumkehr. Die Blutdruck- und Uteruswirkungen von Hypophysenextrakten blieben dagegen unverändert, ebenso die Herzwirkungen des Adrenalins und die Wirkungen einer Reizung parasympathischer Nerven. Dale sah klar die Spezifität der „paralysierenden“ Wirkung des Mutterkorns auf „die myoneuralen Kontaktstellen (myoneural junctions) des sympathischen oder thorako-lumbalen Teils des vegetativen Nervensystems“ – in heutiger Terminologie die „Adrenozeptoren“. Er sah auch klar die Spezifität für diejenigen myoneuralen Kontaktstellen, die bei glatten Muskeln Kontraktion – und nicht Erschlaffung – vermittelten. Aber er ging nicht weiter. Er sah keine Beziehung zwischen den glattmuskelrelaxierenden und den herzstimulierenden Wirkorten der Katecholamine.
In diesem Nebel blieben die Katecholaminrezeptoren mehr als vierzig Jahre. Es wurden neue Antagonisten gefunden, so in der Schweiz das Tolazolin[70] und in den USA das Phenoxybenzamin,[71] aber sie blockierten wie die Mutterkornalkaloide nur die glattmuskelstimulierenden Rezeptoren. Wichtiger war die Synthese neuer Agonisten bei Boehringer Ingelheim. Unter ihnen ragt das Isoprenalin, N-Propylnoradrenalin, hervor. Es wurde zusammen mit Adrenalin und weiteren N-substituierten Noradrenalinderivaten von Richard Rössler und Heribert Konzett am Pharmakologischen Institut der Universität Wien untersucht, vor allem auf Bronchospasmolyse. Die beiden Pharmakologen bedienten sich des selbst entwickelten, später so genannten Konzett-Rössler-Tests.[72] Zuerst injizierten sie den narkotisierten Versuchstieren zur Auslösung eines Bronchialkrampfs Pilocarpin, anschließend das Katecholamin. „Vergleicht man nun alle untersuchten Amine unter dem Gesichtspunkt der broncholytischen Wirkungsstärke, so ergibt sich eine Reihe, die vom höchstwirksamen Isopropyladrenalin <= Isoprenalin> über Äthyladrenalin zu den ungefähr gleich wirksamen Körpern Adrenalin, Propyladrenalin, Butyladrenalin und schließlich zu dem nur schwach wirksamen Isobutyladrenalin führt.“[73] Daneben wirkte Isoprenalin stark positiv inotrop und chronotrop. Boehringer führte es 1940 als Asthmamittel ein. Nach dem Krieg kam es auch bei Deutschlands ehemaligen Feinden in Gebrauch und erhielt im Lauf der Zeit ungefähr 50 Markennamen. Über den therapeutischen Nutzen hinaus war es einer der Agonisten, mit dessen Hilfe Raymond Ahlquist das Rätsel der myoneuralen Kontaktstellen, der „myoneural junctions“, löste. „Dadurch verbreitete sich die Reputation dieser Substanz weltweit, und sie wurde ein Werkzeug vieler Forschungen in Pharmakologie und Therapie.“[74] Allerdings ist es bei Überdosierung durch die Herzwirkung zu zahlreichen Todesfällen gekommen, angeblich allein im Vereinigten Königreich etwa dreitausend.[75]
Ahlquist leitete die Pharmakologische Abteilung der University of Georgia, heute Georgia Health Sciences University. Er sah 1948, was Dale 1906 entgangen war. „Man hat zwei Klassen von Adrenozeptoren unterschieden, solche zur Erregung und solche zur Hemmung der Effektorzellen. Die hier beschriebenen Experimente zeigen, dass es zwar zwei Arten von Adrenozeptoren gibt, diese aber nicht einfach als erregend oder hemmend differenziert werden können, weil jede Art Rezeptor sowohl Erregung als auch Hemmung vermitteln kann, je nachdem wo sie sich befindet.“ Ahlquist bestimmte die Wirkung von sechs Agonisten, darunter Adrenalin, Noradrenalin, α-Methylnoradrenalin und Isoprenalin, auf diverse Organe. Er fand, dass die sechs an den Organen zwei und nur zwei Wirkstärkenreihenfolgen (orders of potency) besaßen. Zum Beispiel wirkten sie in der Reihenfolge „Adrenalin > Noradrenalin > α-Methylnoradrenalin > Isoprenalin“ vasokonstriktorisch, aber in der Reihenfolge „Isoprenalin > Adrenalin > α-Methylnoradrenalin > Noradrenalin“ am Herzen positiv ino- und chronotrop. Die Rezeptoren mit der ersten Wirkstärkenreihenfolge (zum Beispiel für Vasokonstriktion) nannte er α-Adrenozeptoren („alpha adrenotropic receptor“), die Rezeptoren mit der zweiten Wirkstärkenreihenfolge (zum Beispiel für Herzstimulation, aber auch für Bronchodilatation) nannte er β-Adrenozeptoren („beta adrenotropic receptor“). „Dies Konzept zweier fundamentaler Rezeptortypen steht direkt dem Konzept zweier Mediatorsubstanzen (Sympathin E und Sympathin I) entgegen, das Cannon und Rosenblueth vorgelegt haben und das heute vielfach als ‚Gesetz‘ der Physiologie bezeichnet wird. Es gibt nur ein adrenerges Neuro-Hormon oder Sympathin, und dies Sympathin ist identisch mit Adrenalin.“[76]
Der Nebel um die Adrenozeptoren war weggeblasen. Doch wurde Ahlquists Manuskript vom eigentlich zuständigen Journal of Pharmacology and Experimental Therapeutics abgelehnt und ersten beim zweiten Versuch vom American Journal of Physiology akzeptiert. Vielleicht trug die harsche Kritik an Cannon und Rosenblueth dazu bei.
Im Rückblick ist festzustellen, dass Ahlquist zwar bei dem Postulat „ein Transmitter – zwei Rezeptoren“ recht hatte, bei der Identifizierung des Transmitters mit Adrenalin aber unrecht. Er unterließ es auch, aus der Selektivität der damals bekannten Antagonisten für die α-Adrenozeptoren ein Zusatzargument zu machen. Der Rückblick nötigt sogar zu noch weitergehender Relativierung: Man weiß heute, dass zur Kontraktion vieler glatter Muskeln bei Sympathikusreizung, darunter zur neurogenen Vasokonstriktion, das mit dem Noradrenalin zusammen gespeicherte Adenosintriphosphat als Kotransmitter beiträgt. Es wirkt auf die von den Adrenozeptoren sehr verschiedenen auf P2X-Purinozeptoren.[77]
Die α,β-Terminologie setzte sich zunächst langsam, seit zwei Publikationen des Jahres 1958 schnell durch. In der ersten Publikation, aus den Laboratorien von Eli Lilly and Company in Indianapolis, blockierte Dichlorisoprenalin selektiv einige glattmuskelrelaxierende Wirkungen von Adrenalin und Isoprenalin.[78] In der zweiten blockierte es die Herzwirkungen der beiden Substanzen.[79] In der ersten, die Ahlquist nicht erwähnt, blockierte Dichlorisoprenalin „gewisse hemmende Adrenozeptoren“; in der zweiten aber untermauerten die Ergebnisse „das Postulat Ahlquists (1948), dass die hemmenden Adrenozeptoren und die positiv chronotropen und inotropen Herzrezeptoren funktionell identisch, nämlich beide vom beta-Typ sind. … Wir schlagen vor, diese Terminologie auszudehnen, die Antagonisten nämlich nach dem Rezeptor zu benennen, zu dem sie die höchste Affinität besitzen, als entweder α- oder β-Adrenozeptor-Antagonisten (either alpha or beta adrenergic blocking drugs).“
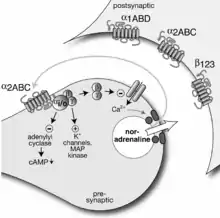
Dichlorisoprenalin war der erste „Betablocker“ (mit einer gewissen intrinsischen Aktivität). Pronethalol folgte 1962 und Propranolol 1964,[80] beide von James Black und seinen Mitarbeitern bei Imperial Chemical Industries Pharmaceuticals in England erfunden. 1967 wurden die β-Adrenozeptoren in β1 und β2 untergliedert,[81] und in den späten 1970er Jahren kündigte sich ein dritter β-Typ an, vor allem in Fettzellen.[82]
Die Untergliederung der α-Adrenozeptoren begann 1971 mit der Entdeckung einer Selbstregelung der Noradrenalinfreisetzung über α-Rezeptoren an den noradrenergen präsynaptischen Endigungen, sogenannte präsynaptische α-Autorezeptoren. Ihre Existenz war anfangs heftig umstritten, ist aber jetzt zum Beispiel durch den Nachweis ihrer Messenger-RNA in noradrenergen Nervenzellen gesichert.[83][84][85] Sie wichen von den lange bekannten α-Rezeptoren der Effektorzellen ab und wurden Prototyp der α2-Rezeptoren, während die glattmuskelerregenden Rezeptoren α1 wurden.[86]

Schon bevor Dopamin als dritter Katecholamin-Transmitter erkannt wurde, vermutete Blaschko, es besitze eigene Rezeptoren. Er erinnerte an eine 1942er Rostocker Arbeit von Holtz und seiner Gruppe: Dopamin senkte in kleinen Dosen bei Meerschweinchen und Kaninchen den Blutdruck, während Adrenalin ihn stets steigerte.[88] Holtz interpretierte irrig, aber Blaschko hatte „keinen Zweifel, dass seine Beobachtungen von größter historischer Bedeutung sind als erster Hinweis auf eine Wirkung des Dopamins, die sich charakterstisch und spezifisch von den Wirkungen der beiden anderen Catecholamine unterschied“.[89] Ein Nachuntersucher schlug 1964 „spezifische Vasodilatations-vermittelnde Dopaminrezeptoren“ vor,[90] und gleichzeitig mehrten sich andere Belege für separate, von den α- und β-Adrenozeptoren verschiedene Dopaminrezeptoren.
1986 klonierten sechzehn Wissenschaftler, darunter Robert Lefkowitz und Brian Kobilka von der Duke University in Durham, North Carolina, das erste Gen eines Catcholaminrezeptors, des β2-Adrenozeptors aus der Lunge von Hamstern.[91] Heute sind die Gene aller Säuger-Katecholaminrezeptoren kloniert, für die neun Adrenozeptoren α1A, α1B, α1D, α2A, α2B, α2C, β1, β2 und β3 sowie die fünf Dopaminrezeptoren D1, D2, D3, D4 und D5. Man beginnt, ihre Feinstruktur in Agonist-freiem und Agonist-aktiviertem Zustand zu verstehen.[92]
Earl Wilbur Sutherland erhielt den 1971er Nobelpreis für Physiologie oder Medizin „für seine Entdeckungen zu den Wirkmechanismen von Hormonen“, insbesondere die Entdeckung des cyclischen Adenosinmonophosphats als second messenger der Katecholamine an β-Adrenozeptoren und des Glucagons an Glucagonrezeptoren. Von hier schritt die Forschung zu den G-Protein-gekoppelten Rezeptoren. 1988 war James Black einer von drei Gewinnern des Preises „für ihre Entdeckungen wichtiger Prinzipien der Arzneibehandlung“, bei Black des Prinzips der Blockade von β-Adrenozeptoren und Histamin-H2-Rezeptoren. 2012 teilten sich Robert Lefkowitz und Brian Kobilka den Nobelpreis für Chemie „für ihre Untersuchungen G-Protein-gekoppelter Rezeptoren“.
Übersichtsartikel zur Katecholaminforschung
- Paul Trendelenburg: Adrenalin und adrenalinverwandte Substanzen. In: A. Heffter (Hrsg.): Handbuch der experimentellen Pharmakologie. Zweiter Band, 2. Hälfte. Berlin, Julius Springer 1924, S. 1130–1293.
- H. Blaschko: Catecholamines 1922–1971. In: H. Blaschko und E. Muscholl (Hrsg.): Catcholamines. Handbuch der experimentellen Pharmakologie XXXIII. Berlin, Springer-Verlag, 1972, S. 1–15. ISBN 0-387-05517-7.
- Herman Blaschko: A half–century of research on catecholamine biosynthesis. In: Journal of Applied Cardiology 1987; 2:171–183.
- Zénon M. Bacq: Chemical transmission of nerve impulses. In: M. J. Parnham und J. Bruinvels (Hrsg.): Discoveries in Pharmacology. Volume 1: Psycho- and Neuropharmacology, Amsterdam, Elsevier, 1983, S. 49–103. ISBN 0-444-80493-5.
- M. R. Bennett: One hundred years of adrenaline: the discovery of autoreceptors. In: Clinical Autonomic Research 9, 1999, S. 145–159. doi:10.1007/BF02281628
- Josef Donnerer und Fred Lembeck: Adrenaline, noradrenaline and dopamine: the catecholamines. In: The Chemical Languages of the Nervous System. Basel, Karger, 2006, S. 150–160.
Einzelnachweise
- John Henderson: Ernest Starling and ‚Hormones’: an historical commentary. In: Journal of Endocrinology. Band 184, 2005, S. 5–10, doi:10.1677/joe.1.06000.
- Henry Hyde Salter: On Asthma: its pathology and therapy. Philadelphia, Blanchard 1864.
- A. Vulpian: Note sur quelques réactions propres à la substance des capsules surrénales. In: Comptes rendus de l'Académie des Sciences. 43, 1856, S. 663–665.
- C. Jacobj: Beiträge zur physiologischen und pharmakologischen Kenntniss der Darmbewegungen mit besonderer Berücksichtigung der Beziehung der Nebenniere zu denselben. In: Archiv für experimentelle Pathologie und Pharmakologie. Band 29, 1892, S. 171–211, doi:10.1007/BF01966116.
- Stephen W. Carmichael: The history of the adrenal medulla. In: Reviews in Neurosciences. Band 2, 1989, S. 83–99, doi:10.1515/REVNEURO.1989.2.2.83.
- H. Dale: Natural chemical stimulators. In: Edinburgh Medical Journal. Band 45, 1938, S. 461–480.
- Henry Dale: Accident and opportunism in medical research. In: British Medical Journal. 1, 1916, S. 73–73, doi:10.1136/bmj.2.4574.451
- Merriley Borell: Organotherapy, British physiology, and discovery of the internal secretions. In: Journal of the History of Biology 9, 1976, S. 235–286.
- G. Oliver und E. A. Schäfer: On the physiological action of extracts of pituitary body and certain other glandular organs. In: Journal of Physiology 18, 1895, S. 277–279. PMC 1514634 (freier Volltext)
- George Oliver: On the therapeutic employment of the suprarenal glands. In: British Medical Journal. 1895, S. 653–655, doi:10.1136/bmj.2.1811.635 (Teil 2).
- E. A. Schäfer: On the present condition of our knowledge of the function of the suprarenal capsules. In: British Medical Journal. 1908, S. 1277–1281, doi:10.1136/bmj.1.2474.1277 (Teil 1).
- K. Starke: Pharmakologie noradrenerger und adrenerger Systeme – Pharmakotherapie des Asthma bronchiale – Doping. In: K. Aktories, U. Förstermann, F. Hofmann und K. Starke: Allgemeine und spezielle Pharmakologie und Toxikologie. 10. Auflage, Elsevier, München 2009, S. 161–199. ISBN 978-3-437-42522-6.
- H. Barcroft und J. F. Talbot: Oliver and Schäfer's discovery of the cardiovascular action of suprarenal extract. In: Postgraduate Medical Journal 44, 1968, S. 6–8. doi:10.1136/pgmj.44.507.6
- Jesse G. M. Bullowa und David M. Kaplan: On the hypodermatic use of adrenalin chloride in the treatment of asthmatic attacks. In: New York Medical Journal and Medical Record: a Weekly Review of Medicine. Band 83, 1903, S. 787–790.
- R. H. Kahn: Zur Physiologie der Trachea, In Archiv für Anatomie und Physiologie, Archiv für Physiologie 1907, S. 398–426.
- Hans Januschke und Leo Pollak: Zur Pharmakologie der Bronchialmuskulatur. In: Archiv für experimentelle Pathologie und Pharmakologie 66, 1911, S. 205–220. doi:10.1007/BF01841068.
- H. Braun: Ueber den Einfluss der Vitalität der Gewebe auf die örtlichen und allgemeinen Giftwirkungen localanästhesirender Mittel und über die Bedeutung des Adrenalins für die Localanästhesie. In: Archiv für klinische Chirurgie 69, 1903, S. 541–591.
- Ladislaus Szymonowicz: Die Function der Nebenniere. In: Archiv für die gesamte Physiologie des Menschen und der Tiere. Band 64, 1895, S. 97–164, doi:10.1007/BF01661663.
- W. H. Bates: The use of extract of suprarenal capsule in the eye. In: New York MedicalJournal. 1896, S. 647–650.
- Carl G. A. Persson: Astute observers discover anti-asthma drugs. In: Pharmacology & Toxicology 77, 1995, Supplement 3, S. 7–15. doi:10.1111/j.1600-0773.1995.tb01934.x
- Erich Spengler: Kritisches Sammelreferat über die Verwendung einiger neuerer Arzneimittel in der Augenheilkunde. In: Ophthalmologica 13, 1905, S. 33–47. doi:10.1159/000290295
- John J. Abel: Ueber den blutdrucksteigernden Bestandtheil der Nebenniere, das Epinephrin. In: Zeitschrift für physiologische Chemie 28, 1899–1900, S. 318–361
- Otto v. Fürth: Zur Kenntniss der brenzcatechinähnlichen Substanz der Nebennieren. In: Zeitschrift für physiologische Chemie 29, 1900, S. 105–123.
- Jokichi Takamine: Adrenalin the active principle of the suprarenal glands and its mode of preparation. In: The American Journal of Pharmacy 73, 1901, S. 523–535.
- N. Ph. Tendeloo, Allgemeine Pathologie, 9. März 2013, Springer-Verlag, S. 654 ISBN 978-3-642-92320-3. Abgerufen am 15. September 2015.
- Vgl. auch E. Mayer: Clinical experience with adrenalin. In: Philadelphia. M. J. Band 7, 1901, S. 819 ff.
- E. Friedmann. Die Konstitution des Adrenalins. In: Beiträge zur chemischen Physiologie und Pathologie 8, 1906, S. 95–120.
- Arthur R. Cushney: The action of optical isomers. In: The Journal of Physiology 37, 1908, S. 130–138. PMC 1533541 (freier Volltext)
- E. M. Tansey: What's in a name? Henry Dale and adrenaline, 1906. In: Medical History 39, 1995, S: 459–476. PMC 1037030 (freier Volltext)
- M. Lewandowsky: Ueber die Wirkung des Nebennierenextractes auf die glatten Muskeln, im Besonderen des Auges. In: Archiv für Anatomie und Physiologie, Archiv für Physiologie 1899, S. 360–366.
- T. R. Elliott: On the action of adrenalin. In: The Journal of Physiology 31, 1904, S. XX–XXI. PMC 1465436 (freier Volltext)
- L. Stjärne, P. Hedqvist, H. Lagercrantz und Å. Wennmalm (Hrsg.): Chemical Neurotransmission 75 Years. London, Academic Press, 1981, S. XIII.
- O. Loewi: Über humorale Übertragbarkeit der Herznervenwirkung. I. Mitteilung. In: Pflügers Archiv für die gesamte Physiologie des Menschen und der Tiere. 189, 1921, S. 239–242. doi:10.1007/BF01738910
- O. Loewi: Über humorale Übertragbarkeit der Herznervenwirkung. II. Mitteilung. In: Pflügers Archiv für die gesamte Physiologie des Menschen und der Tiere 193, 1922, S. 201–213. doi:10.1007/BF02331588
- O. Loewi: Quantitative und qualitative Untersuchungen über den Sympathicusstoff. In: Pflügers Archiv für die gesamte Physiologie des Menschen und der Tiere 237, 1936, S. 504–514. doi:10.1007/BF01753035
- G. Barger und H. H. Dale: Chemical structure and sympathomimetic action of amines. In: The Journal of Physiology 1910, 41, S. 19–59. PMC 1513032 (freier Volltext)
- H. H. Dale: Nomenclature of fibres in the autonomic system and their effects. In: The Journal of Physiology 80, 1934, S. 10P–11P. PMC 1394004 (freier Volltext)
- U. S. v. Euler: A sympathomimetic pressor substance in animal organ extracts. In: Nature 156, 1945, S. 18–19. doi:10.1038/156018b0
- U. S. v. Euler: A specific sympathomimetic ergone in adrenergic nerve fibres (sympathin) and its relations to adrenaline and nor-adrenaline. In: Acta Physiologica Scandinavica 12, 1946 S. 73–97.
- W. S. Peart: The nature of splenic sympathin. In: The Journal of Physiology 1948, 108, S. 491–501. PMC 1392468 (freier Volltext)
- Peter Holtz, Kartl Credner und Günther Kroneberg: Über das sympathicomimetische pressorische Prinzip des Harns („Urosympathin“).In: Naunyn-Schmiedebergs Archiv für experimentelle Pathologie und Pharmakologie 204, 1947 S. 228–243. doi:10.1007/BF00738347
- Henry Hallett Dale: Adventures in Pharmacology. With Excursions into Autopharmacology. Pergamon Press, London 1953, S. 98.
- Bangen, Hans: Geschichte der medikamentösen Therapie der Schizophrenie. Berlin 1992, ISBN 3-927408-82-4 S. 90–94 Neuroleptika und psychiatrische Theorienbildung.
- Marthe Vogt: The concentration of sympathin in different parts of the central nervous system under normal conditions and after the administration of drugs. In: The Journal of Physiology 123, 1954, S. 451–481. PMC 1366219 (freier Volltext)
- Å. Bertler und E. Rosengren: Occurrence and distribution of dopamine in brain and other tissues. In: Experientia 15, 1959, S. 10–11.
- Arvid Carlsson: The occurence, distribution and physiological role of catcholamines in the nervous system. In: Pharmacological Reviews 11, 1959, S. 490–493.
- H. Ehringer und O. Hornykiewicz: Verteilung von Noradrenalin und Dopamin (3-Hydroxytyramin) im Gehirn des Menschen und ihr Verhalten bei Erkrankungen des extrapyramidalen Systems. In: Klinische Wochenschrift 38, 1960, S. 1236–1239. doi:10.1007/BF01485901
- Oleh Hornykiewicz: From dopamine to Parkinson’s disease: a personal research record. In: Fred Samson und George Adelman (Hrsg.): The Neurosciences: Paths of Discovery II. Basel, Birkhäuser, 1992, S. 125–147.
- Blaschko 1987.
- Peter Holtz, Rudolf Heise und Käthe Lüdtke: Fermentativer Abbau von l-Dioxyphenylalanin (Dopa) durch Niere. In: Naunyn-Schmiedebergs Archiv für experimentelle Pathologie und Pharmakologie 191, 1939, S. 87–118. doi:10.1007/BF01994628
- Hermann Blaschko: The specific action of l-dopa decarboxylase. In: The Journal of Physiology 1939, 96, S. 50 P–51 P. PMC 1393737 (freier Volltext)
- P. Holtz: Dopadecarboxylase. In: Die Naturwissenschaften 27, 1939, S. 724–725.
- Edith Bülbring: The methylation of noradrenaline by minced suprarenal tissue. In: British Journal of Pharmacology 4, 1949, S. 234–244.
- Julius Axelrod: Purification and properties of phenylethanolamine-N-methyl transferase. In: The Journal of Biological Chemistry 237, 1962, S: 1657–1660.
- Mary Lilias Christian Hare: Tyramine oxidase. I. A new enzyme system in liver. In: Biochemical Journal 22, 1928, S. 968–979. (PDF; 1,2 MB)
- Hermann Blaschko, Derek Richter und Hans Schlossmann: The oxidation of adrenaline and other amines. In: Biochemical Journal 31, 1937, S. 2187–2196. (PDF; 1,1 MB)
- Blaschko 1972.
- Julius Axelrod: O-Methylation of epinephrine and other catechols. In: Science 126, 1962, S. 400–401.
- H. Blaschko und A. D. Welch: Localization of adrenaline in cytoplasmic particles of the bovine adrenal medulla. In: Naunyn-Schmiedebergs Archiv für experimentelle Pathologie und Pharmakologie 219, 1953, S: 17–22. doi:10.1007/BF00246245
- Nils-Åke Hillarp, Sten Lagerstedt und Bodil Nilson: The isolation of a granular fraction from the suprarenal medulla, containing the sympathomimetic catechol amines. In: Acta physiologica scandinavica 29, 1954, S: 251–263.
- U. S. von Euler und N. Å. Hillarp: Evidence for the presence of noradrenaline in submicroscopic structures of adrenergic axons. In: Nature 177, 1956, S. 44–45. doi:10.1038/177044b0
- E. de Robertis, Amanda Pellegrino de Iraldi, Georgina Rodríguez de Lores Arnaiz und Luis M. Zieher: Synaptic vesicles from the rat hypothalamus. Isolation and norepinephrine content. In: Life Sciences 4, 1965, S. 193–201. doi:10.1016/0024-3205(65)90119-0
- H. J. Schümann und H. Grobecker: Über den Noradrenalin- und ATP-Gehalt sympathischer Nerven. In: Naunyn-Schmiedebergs Archiv für experimentelle Pathologie und Pharmakologie 233, 1958, S. 296–300. doi:10.1007/BF00245643
- Å. Thureson-Klein: Exocytosis from large and small dense cored vesicles in noradrenergic nerve terminals. In: Neuroscience 10, 1983, S. 245–252. doi:10.1016/0306-4522(83)90132-X
- A. Fröhlich und O. Loewi: Über eine Steigerung der Adrenalinempfindlichkeit durch Cocaïn. In: Archiv für experimentelle Pathologie und Pharmakologie 62, 1910, S. 159–169. doi:10.1007/BF01840652
- G. Hertting und J. Axelrod: Fate of tritiated noradrenaline at the sympathetic nerve-endings. In: Nature 192, 1961, S. 172–173. doi:10.1038/192172a0
- Leslie L. Iversen: The Uptake and Storage of Noradrenaline in Sympathetic Nerves. University Press, Cambridge 1967.
- U. Trendelenburg: The metabolizing systems involved in the inactivation of catecholamines. In: Naunyn-Schmiedeberg's Archives of Pharmacology 332, 1986, S. 201–207. doi:10.1007/BF00504854
- H. H. Dale: On some physiological actions of ergot. In: The Journal of Physiology 34, 1906, S. 163–206.PMC 1465771 (freier Volltext)
- Max Hartmann und Hans Isler: Chemische Konstitution und pharmakologische Wirksamkeit von in 2-Stellung substituierten Imidazolinen. In: Naunyn-Schmiedebergs Archiv für experimentelle Pathologie und Pharmakologie 192, 1939, S. 141–154. doi:10.1007/BF01924807
- Mark Nickerson: The pharmacology of adrenergic blockade. In: Pharmacological Reviews 1, 1949, S. 27–101.
- Heribert Konzett und Richard Rössler: Versuchsanordnung zu Untersuchungen an der Bronchialmuskulatur. In: Naunyn-Schmiedebergs Archiv für experimentelle Pathologie und Pharmakologie 195, 1940, S. 71–74. doi:10.1007/BF01861842
- Heribert Konzett: Neue broncholytisch hochwirksame Körper der Adrenalinreihe. In: Naunyn-Schmiedebergs Archiv für experimentelle Pathologie und Pharmakologie 197, 1941, S. 27–40. doi:10.1007/BF01936304
- H. Konzett: On the discovery of isoprenaline. In: Trends in Pharmacological Sciences 2, 1981, S. 47–49. doi:10.1016/0165-6147(81)90259-5
- Walter Sneader: Drug Discovery: The Evolution of Modern Medicines. John Wiley & Sons, Chichester, 1985, S. 103. ISBN 0-471-90471-6.
- Raymond P. Ahlquist: A study of the adrenotropic receptors. In: American Journal of Physiology 153, 1948, S. 586–600.
- Ivar von Kügelgen und Klaus Starke: Noradrenalin-ATP co-transmission in the sympathetic nervous system. In: Trends in Pharmacological Sciences 12, 1991, S. 319–324. doi:10.1016/0165-6147(91)90587-I
- C. E. Powell und I. H. Slater: Blocking of inhibitory adrenergic receptors by a dichloro analog of isoproterenol. In: Journal of Pharmacology and Experimental Therapeutics 122, 1958, S. 480–488.
- Neil C. Moran und Marjorie E. Perkins: Adrenergic blockade of the mammalian heart by a dichloro analogue of isoproterenol. In: Journal of Pharmacology and Experimental Therapeutics 124, 1958, S. 223–237.
- J. W. Black, A. F. Crowther, R. G. Shanks und A. C. Dornhorst: A new adrenergic beta-receptor antagonist. In: The Lancet 283, 1964, S. 1080–1081. doi:10.1016/S0140-6736(64)91275-9.
- A. M. Lands, A. Arnold, J. P. McAuliff, F. P. Luduena und T. G. Brown: Differentiation of receptor systems activated by sympathomimetic amines. In: Nature 214, 1967, S. 597–598. doi:10.1038/214597a0.
- Johan Zaagsma und Stefan R. Nahorski: Is the adipocyte β-adrenoceptor a prototype of the recently cloned β3-adrenoceptor? In: Trends in Pharmacological Sciences 11, 1990, S. 3–7. doi:10.1016/0165-6147(90)90032-4.
- Anthony P. Nicholas, Vincent Pieribone und Tomas Hökfelt: Distributions of mRNAs for alpha-2 adrenergic receptor subtypes in rat brain: an in situ hybridization study. In: Journal of Comparative Neurology 328, 1993, S. 575–594. doi:10.1002/cne.903280409.
- Klaus Starke: Presynaptic autoreceptors in the third decade: focus on α2-adrenoceptors. In: Journal of Neurochemistry 78, 2001, S. 685–693. doi:10.1046/j.1471-4159.2001.00484.x
- Ralf Gilsbach und Lutz Hein: Presynaptic metabotropic receptors for acetylcholine and adrenaline/noradrenaline. In: Thomas C. Südhoff und Klaus Starke (Hrsg.): Pharmacology of Neurotransmitter Release. Handbook of Experimental Pharmacology 184. Springer, Berlin 2008, S. 261–288. ISBN 978-3-540-74804-5.
- Salomón Z. Langer: Presynaptic regulation of catecholamine release. In: Biochemical Pharmacology 23, 1974, S. 1793–1800. doi:10.1016/0006-2952(74)90187-7
- Robert Frederiksson, Malin C. Lagerström, Lars-Gustav Lundin und Helgi B. Schiöth: The G-protein-coupled receptors in the human genome form five main families. Phylogenetic analysis, paralogon groups, and fingerprints. In: Molecular Pharmacology 63, 2003, S. 1256–1272.
- Peter Holtz, Karl Credner und Wolfgang Koepp: Die enzymatische Entstehung von Oxytyramin im Organismus und die physiologische Bedeutung der Dopadecarboxylase. In: Naunyn-Schmiedebergs Archiv für experimentelle Pathologie und Pharmakologie 200, 1942, S. 356–388. doi:10.1007/BF01860725
- Blaschko 1987.
- John Nelson Eble: A proposed mechanism for the depressor effect of dopamine in the anesthetized dog. In: Journal of Pharmacology and Experimental Therapeutics 145, 1964, S. 64–70.
- Richard A. F. Dixon, Brian K. Kobilka, David J. Strader und 13 andere Autoren; Cloning of the gene and cDNA for mammalian β-adrenergic receptor and homology with rhodopsin. In: Nature 321, 1986, S. 75–79. doi:10.1038/321075a0
- Daniel M. Rosenbaum, Cheng Zhang, Joseph A. Lyons und 15 andere Autoren: Structure and function of an irreversible agonist-β2 adrenoceptor complex. In: Nature 469, 2011, S. 236–240. doi:10.1038/nature09665