Pyrenäen-Desman
Der Pyrenäen-Desman (Galemys pyrenaicus) ist eine Säugetierart aus der Familie der Maulwürfe (Talpidae). Gemeinsam mit dem Russischen Desman bildet er die Gruppe der Desmane (Desmanini), die von allen Maulwürfen am ausgeprägtesten an eine wasserbewohnende Lebensweise angepasst sind. Der gesamte Körperbau wirkt gedrungen. Charakteristische Merkmale finden sich in dem langen, am Ende abgeplatteten Schwanz, der rüsselartigen, sehr beweglichen Schnauze und den Schwimmhäuten zwischen den Zehen der sehr kräftigen Hinterbeine. Abweichend von anderen Maulwürfen sind die Vorderbeine eher schlank. Das Fell hat eine überwiegend dunkle Färbung. Das Verbreitungsgebiet des Pyrenäen-Desmans umfasst die namensgebenden Pyrenäen und den Norden der Iberischen Halbinsel. Als Lebensraum nutzen die Tiere sauerstoffreiche kleinere Flüsse in mittleren bis höheren Gebirgslagen, die eine reichhaltige Wirbellosengemeinschaft bieten und deren Ufer eine vielfältige Gestaltung mit Steinen oder Felsen als Versteckmöglichkeiten aufweisen.
| Pyrenäen-Desman | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Pyrenäen-Desman (Galemys pyrenaicus) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name der Gattung | ||||||||||||
| Galemys | ||||||||||||
| Kaup, 1829 | ||||||||||||
| Wissenschaftlicher Name der Art | ||||||||||||
| Galemys pyrenaicus | ||||||||||||
| (E. Geoffroy, 1811) |
Die Lebensweise ist relativ gut erforscht. Die Tiere sind nachtaktiv, können aber auch in kürzeren Zeitphasen tagsüber auftreten. Jedes Individuum bewegt sich innerhalb eines Aktionsraumes, der sich entlang der Flussufer zieht. In den Aktionsräumen sind mehrere Ruheplätze in Felsspalten und Hohlräumen verteilt. Sowohl in der Dauer der Aktivitätsphasen als auch in der Nutzung der Aktionsräume bestehen gewisse jahreszeitliche Unterschiede. Eine ursprünglich angenommene territoriale und aggressive Verhaltensweise gegenüber Artgenossen hat sich in Untersuchungen seit den 2000er Jahren nicht bestätigt. Der Pyrenäen-Desman ist ein guter Schwimmer, der intensive Tauchgänge unternimmt. Seine Nahrung sucht er überwiegend im Wasser auf dem Grund. Hauptsächlich ernährt er sich von Wirbellosen, selten auch von Wirbeltieren. Zu den bevorzugten Beutetieren gehören Flohkrebse sowie Insekten wie Köcherfliegen und Eintagsfliegen. Die Fortpflanzung erfolgt im Frühjahr, Weibchen können aber mehrmals im Jahr Nachwuchs gebären. Die Tragzeit dauert vermutlich rund 30 Tage.
Die wissenschaftliche Erstbeschreibung des Pyrenäen-Desmans erfolgte im Jahr 1811. Vor allem im Verlauf des 19. Jahrhunderts war er unter der Gattungsbezeichnung Mygale bekannt, die jedoch ebenfalls für eine Artengruppe der Spinnen vergeben ist. Die heute gültige Gattungsbezeichnung Galemys wurde im Jahr 1829 eingeführt. Teilweise werden zwei Unterarten angenommen, die sich aber genetisch nicht bestätigen lassen. Fossilfunde sind seit dem Oberen Miozän beziehungsweise Unteren Pliozän bekannt. In ihrer stammesgeschichtlichen Vergangenheit waren die Vorfahren des Pyrenäen-Desmans über weite Teile Europas verbreitet. Der heutige Bestand gilt als gefährdet, was neben der Zerstückelung der Lebensräume und Störung des Gewässernetzes auch auf aktive Verfolgung durch Fischer zurückzuführen ist.
Merkmale
Habitus


Der Pyrenäen-Desman ist ein großer Vertreter der Maulwürfe, bleibt aber in seinen Ausmaßen hinter dem nahe verwandten Russischen Desman (Desmana moschata) zurück. Er erreicht eine Kopf-Rumpf-Länge von 10,6 bis 12,4 cm. Hinzu kommt ein 11,8 bis 14,8 cm langer Schwanz, der dadurch in etwa die Länge des übrigen Körpers aufweist oder diesen übertrifft. Das Gewicht variiert von 51 bis 79 g. Ein Sexualdimorphismus besteht nicht. Insgesamt wirkt der Pyrenäen-Desman sehr gedrungen, was durch das dichte, wasserabweisende Fell verstärkt wird. Einzelne Anpassungen im Körperbau ergeben sich aus der semi-aquatischen Lebensweise. Der Kopf weist eine konische Form auf und sitzt auf einem, für Maulwürfe typischen kurzen Hals. Die Schnauze ist lang ausgezogen und sehr beweglich, ähnelt dadurch einem Rüssel. Sie wird rund 2 cm lang und endet verbreitert sowie abgeflacht. Oberseits und unterseits verläuft jeweils eine längsgerichtete Rille. Die am Ende sitzenden Nasenlöcher öffnen nach oben. Der Nasenspiegel ist fleischfarben. Ihn bedecken zahlreiche Mikrovibrissen, die rund 100 μm Länge aufweisen. Sie ragen zwischen kleinen buckelartigen Erhebungen hervor, die wiederum rosettenartig angeordnet sind. Es handelt sich hierbei wohl um ein Äquivalent zum Eimerschen Organ der grabenden Maulwürfe.[1][2] Umgeben wird der Nasenspiegel von rund 2 cm langen Vibrissen, die auch am Kinn sprießen. Äußere Ohren sind nicht erkennbar. Die Augen sind sehr klein, aber nicht von einer Hautschicht überzogen. Sie bleiben im Fell verborgen und werden lediglich durch helleres Haar markiert.[3][4][5][6]
Das Fell ist auf dem Rücken dunkelgraubraun bis rötlichbraun, wobei erstere Variante weitgehend in den Populationen im östlichen, letztere im westlichen Teil des Verbreitungsgebietes auftritt. Die Unterseite erscheint durchgehend zumeist schiefergrau und damit heller, die Übergänge an den Seiten verlaufen fließend. Mit zunehmender Alterung des Fells über das Jahr können sich an Rücken und Bauch gelbliche Flecken ausbilden, was durch Farbveränderungen an den Haarspitzen ausgelöst wird.[7] Allgemein ist das Fell dicht, aber gröber als bei anderen Maulwürfen. Es besteht zum Großteil aus Wollhaar, deren Schäfte 10 μm dick und gewellt sind. Sie stehen in kleinen Büscheln. Darüber legen sich Grannenhaare mit lanzettförmiger Gestalt, deren abgeplattetes Ende unter Wasser das Entweichen der Luft aus dem Wollhaar verhindert. Auffallenderweise fehlt an den Haarspitzen die Markröhre. Die Grannenhaare stehen einzeln, ebenso wie die spindelförmigen Leithaare.[8] Der Schwanz ist nur spärlich behaart, am unteren Schwanzende findet sich allerdings ein Büschel aus Borsten. Er beginnt rund und läuft in einer gepressten Spitze aus. Dadurch unterscheidet sich der Pyrenäen-Desman vom Russischen Desman, dessen Schwanz vollständig platt ist. Die an der Schwanzwurzel ausgebildeten Schuppenringe lösen sich zum Schwanzende hin auf. An der Schwanzunterseite sitzt eine Duftdrüse, die ein moschusähnliches Sekret absondert.[3][4][5][6]
Die Vorderfüße sind abweichend von den grabenden Maulwürfen deutlich kleiner als die Hinterfüße. Die Zehen der Füße werden durch Schwimmhäute miteinander verbunden. An den Außenkanten tragen die Füße zusätzlich Borstensäume, ansonsten sind sie nackt und dunkel gefärbt. Jeder Fuß ist fünfstrahlig aufgebaut, wobei die Mittelzehe am längsten ist und die beiden inneren die äußeren übertreffen. Die Zehen tragen spitze, lange Krallen. Der Hinterfuß ist 32,5 bis 36,5 mm lang. Weibchen haben vier Zitzenpaare, von denen eins im Brust-, eins im Bauch- und zwei im Lendenbereich sitzen.[3][4][5][6]
Schädel- und Gebissmerkmale
Der Schädel ist 32,4 bis 35,9 mm lang. Er ähnelt denen der Eurasischen Maulwürfe (Talpa), ist aber am Hirnschädel weitaus eckiger gestaltet und am Rostrum stärker ausgezogen. Die größte Breite liegt im hinteren Schädelabschnitt und übertrifft kaum den Bereich der Jochbogenansätze. Zwischen den Orbita verengt er sich auf 5,9 bis 6,5 mm. Am Mittelkieferknochen treten seitlich warzenartige Buckel auf. Die Gelenke des Hinterhauptsbeines bleiben in der Ansicht von oben unter dem Schädel verborgen. Der Ausgang des Gehörganges an den Paukenblasen ist groß und nach unten gerichtet. Am Gaumen befindet sich ein relativ großes Foramen incisivum. Der Unterkiefer misst 21,6 bis 22,7 mm in der Länge. Seine Unterkante verläuft gerade, der Kronenfortsatz ragt höher auf als im Vergleich bei den Eurasischen Maulwürfen. Das Gebiss besteht aus der vollständigen Anzahl an Zähnen der Höheren Säugetiere, dementsprechend also 44 mit folgender Zahnformel: . Der vordere obere Schneidezahn ist sehr groß und in Seitenansicht keilförmig gestaltet. Getrennt durch ein Diastema folgen dann die beiden hinteren Schneidezähne, die deutlich kleiner sind, allerdings vom Eckzahn überragt werden. Die Zähne des vorderen Gebissabschnittes zuzüglich der ersten drei Prämolaren besitzen jeweils nur eine Spitze, der letzte Prämolar ist dreispitzig. In der unteren Zahnreihe zeigen sich die ersten beiden Schneidezähne verhältnismäßig groß. Der dritte Schneidezahn sowie der Eckzahn und erste Prämolar sind sehr klein. Wie in der oberen Reihe habe alle Zähne bis zum dritten Prämolaren nur eine Spitze. Die obere Zahnreihe wird 16,8 bis 17,3 mm lang, die untere 14,6 bis 15,2 mm.[5]

Genetische Merkmale
Der diploide Chromosomensatz lautet 2n = 42. Er setzt sich aus 8 acrozentrischen und 12 zweiarmigen Autosomenpaaren zusammen. Das X-Chromosom ist groß und submetazentrisch, das Y-Chromosom klein und fleckenförmig. Die Anzahl an Armen der Autosomen (fundamentale Anzahl) beträgt 68.[9][4][5][6]
Verbreitung und Lebensraum
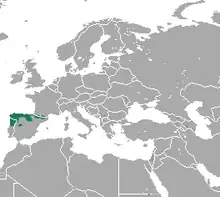
Der Pyrenäen-Desman ist im Norden der Iberischen Halbinsel im südwestlichen Europa verbreitet. Er kommt dort sowohl in den Pyrenäen im Osten als auch im Kantabrischen Gebirge im Westen vor. In den Pyrenäen ist er vorwiegend auf der französischen Seite anzutreffen, während für die spanische Seite nur wenige Fundpunkte existieren. Im Südwesten und Süden erstreckt sich das Vorkommen bis in höhere Gebirgslagen Portugals nördlich des Mondego wie auch in das Iberischen Scheidegebirge im zentraleren Spanien. Zwischen den beiden Landschaftsräumen der Pyrenäen und des Kantabrischen Gebirges besteht eine Besiedlungslücke, die wohl auf die stärkere Industrialisierung und der damit verbundenen Gewässerverschmutzung zurückzuführen ist. Zwei isolierte Populationen finden sich am Zêzere in zentralen Portugal und am Mayor im Baskenland. Die Tiere bewohnen jede Art von Fließgewässer, die sauerstoffreich sind und eine umfangreiche Wirbellosenfauna tragen. Zudem sollten Versteckmöglichkeiten wie Löcher oder Spalten in Ufernähe bestehen. Die generelle Höhenverbreitung reicht von 15 m bis 2700 m über dem Meeresspiegel. In den Pyrenäen sucht der Pyrenäen-Desman schnell fließende Gewässer über der 400 m-Höhenmarke auf. Diese haben häufig eine Durchflussgeschwindigkeit von 2 m/s und eine Sommertemperatur von 1 bis 3 °C. Die meisten zuträglichen Flüsse und Bäche zeichnen sich durch eine hohe Vielfalt an Bodensubstraten mit einem größeren Anteil an Felsen und Gesteinen und einem geringeren an lockeren oder sandigen Böden aus. Gelegentlich dringt er auch in langsam fließende Gewässer wie Kanälen oder in Seen und Sumpfgebiete vor. Die Tiere tolerieren eine gewisse Trübung oder Gewässerverschmutzung, sofern es die präferierte Beute nicht beeinträchtigt. Sie reagieren aber empfindlich auf Störungen des Nahrungsangebotes durch äußere Einflüsse. Insgesamt wird die Habitatwahl zu einem größeren Teil von externen Faktoren wie der generellen Landschaftsgestaltung gesteuert.[10][11][12] Die Populationsdichte kann an Flüssen im Kantabrischen Gebirge Untersuchungen zufolge zwischen 5,0 und 7,3 Individuen je Kilometer Flusslänge betragen, entsprechende Werte für die spanischen Pyrenäen liegen bei 2,8 bis 2,9 Individuen auf einem vergleichbaren Längenabschnitt.[13][14][4][5][6]
Lebensweise
Territorialverhalten

Die Lebensweise des Pyrenäen-Desmans ist in Teilen gut erforscht. Er ist ein guter Schwimmer und an die semi-aquatische Lebensweise angepasst. Hierzu gehören die Schwimmhäute, die kräftigen Hinterbeine, der lange und haarige Schwanze sowie das Fell, das vor zu starker Sonneneinstrahlung im Wasser schützt. Außerdem sind die Nasenlöcher verschließbar.[15] Zum Schwimmen werden nur die Hinterbeine eingesetzt, die Vorderbeine sind vorwärts gerichtet. Dabei vollführen die Hinterbeine abwechselnd kräftige Stöße, die rund 2,5 mal je Sekunde erfolgen. Der am Ende abgeplattete Schwanz dient als Steuerruder.[16] Tauchgänge können bis zu vier Minuten anhalten. Rund 25 % der aktiven Zeit wird am Grund der Gewässer verbracht. Beim Tauchen ist die Nase nach oben gerichtet, so dass die Nasenlöcher nicht direkt der Wasserbewegung ausgesetzt sind. Mit den Krallen der Vorderfüße kann sich der Pyrenäen-Desman an Steinen und Felsen festhalten, wodurch er stärkeren Strömungen gewachsen ist. Sie dienen aber auch zum Erklettern steiler Flusshänge. An Land ist ein Tier eher schwerfällig und zudem kein guter Läufer. Es kann aber gelegentlich auch eigene Tunnel graben. Sobald der Pyrenäen-Desman das Wasser verlässt, befreit er sein Fell von Nässe und beginnt sich zu putzen, was überwiegend mit den Krallen der Hinterfüße erfolgt. Zur Orientierung unter Wasser, aber auch zur Nahrungssuche werden der Geruchs- und Tastsinn eingesetzt. Ersterer ist aber weniger gut entwickelt als bei grabenden Maulwürfen. Die lange und sehr bewegliche Schnauze hat als Tastorgan eine wichtige Funktion und ist dafür mit zahlreichen Vibrissen und einem komplexen Eimerschen Organ ausgestattet, Chemorezeptoren fehlen aber weitgehehend. Der Sehsinn zeigt sich kaum ausgeprägt, allerdings kann der Pyrenäen-Desman zwischen hell und dunkel unterscheiden.[17][18][19][4][5][6]
Der Pyrenäen-Desman ist nachtaktiv. Nach Untersuchungen in der spanischen Provinz Lleida in den östlichen Pyrenäen lässt sich im Frühjahr eine hauptsächliche Aktivitätsphase ausmachen, die überwiegend gegen 23.00 Uhr beginnt und ohne Unterbrechung bis zu acht Stunden anhält. Vor allem im Herbst kommt eine, selten auch zwei zusätzliche Phasen hinzu. Deren durchschnittliche Dauer beträgt fünf Stunden, getrennt durch rund 100 Minuten Ruhe. Die Ruhephase beginnt etwa um 02.00 Uhr. Den Tag über verbringt der Pyrenäen-Desman häufig in einem Unterschlupf. Allerdings können auch dann Aktivitäten auftreten, die im Herbst im Mittel rund 73,7 Minuten anhalten, im Frühjahr aber mit 102 Minuten ausgedehnter sind. Die Gesamtaktivität über den Tag beträgt somit rund neun bis zehn Stunden und ist im Frühjahr ausgiebiger als im Herbst. Der durchschnittliche Anteil der Aktivität an der täglich verfügbaren Zeit liegt bei 36,5 %, dadurch ist der Pyrenäen-Desman insgesamt länger inaktiv als aktiv. Während seiner nächtlichen aktiven Phase legt ein Tier rund 268,5 m zurück, tagsüber verkürzt sich die Distanz auf etwa 141 m.[20][21] Als Unterschlupf nutzen die Tiere Felsüberhänge oder -spalten am Flussufer, deren Eingänge vollständig oder teilweise unter Wasser liegen. Die Ruheplätze selbst befinden sich 30 bis 110 cm vom Ufer entfernt. In der Regel nutzt ein Individuum bis zu sieben derartiger Unterschlüpfe. Sie liegen entlang der Flusslinie rund 248 m auseinander. Einzelne Plätze dienen hierbei als hauptsächliche Rückzugsgebiete, andere werden nur kurzfristig aufgesucht.[20][22] Gelegentlich bezieht der Pyrenäen-Desman auch aufgelassene Baue der Westschermaus. Der Ruheplatz besteht zumeist aus einem 15 cm großen Hohlraum, in dem Blätter, Zweige und Gräser zu einem kugeligen Nest geformt sind. Anhand von Beobachtungen an gefangenen Tieren transportieren diese das benötigte Pflanzenmaterial mit der Schnauze.[16][23][20][4][5][6]
Jedes einzelne Tier nutzt einen Aktionsraum, der sich entlang der Flussufer erstreckt. Nach Untersuchungen an mehreren Flüssen im französischen Département Ariège beanspruchten die Aktionsräume zwischen 184 und 830 m Länge an den Flussläufen. Männchen hatten hier mit rund 429 m Länge größere Aktionsräume als Weibchen mit im Mittel 301 m.[23][20] Ähnliches ließ sich in der spanischen Provinz Lleida feststellen. Dort erstreckten sich die Aktionsräume über 350 bis 2300 m entlang des Flussufers, der Durchschnitt lag bei 523 m. Im Herbst wurden diese intensiver genutzt als im Frühjahr. Um den gesamten Aktionsraum zu erkunden, benötigte ein Individuum rund zwei Tage. Während der kurzen Tagesaktivitäten durchstreifte ein Tier hingegen zwischen 6,2 und 11,8 % der Fläche.[22] Ursprüngliche Annahmen, die auf den Feldstudien in Ariège der 1980er Jahre beruhen, gingen davon aus, dass der Pyrenäen-Desman territorial und aggressiv gegenüber Artgenossen auftritt.[24][23] Dies wird jedoch durch die Untersuchungen in der Provinz Lleida nicht bestätigt. Möglicherweise ist der Pyrenäen-Desman teilweise nomadisch und verlagert seinen Aktionsraum. Es bestehen jedoch gewisse Bindungen an den aktuellen Standort, da Rückkehrexperimente aus bis zu 1,5 km Entfernung erfolgreich verliefen.[19] Innerhalb eines Gebietes teilen sich Individuen des gleichen oder des anderen Geschlechts teilweise ihre Ruheplätze über einen Tag verteilt. Gegenseitige Begegnungen zwischen benachbarten Tieren dauern von zehn Sekunden bis zehn Minuten. Spezielle Verhaltens- oder Raumnutzungsänderungen gegenüber fremden Individuen sind nicht dokumentiert.[22][21] Mit Hilfe von Sekreten der Schwanzdrüse werden Markierungen abgesetzt. Der moschusartige Duft ist allerdings nicht so streng wie beim Russischen Desman. Außerdem nutzt der Pyrenäen-Desman weitgehend die gleichen Latrinen zum Absetzen des Kotes. Lautäußerungen sind nur wenige bekannt. Diese erfolgen als Fiepen in ungewohnten Situationen oder bei der Begegnung mit unbekannten Artgenossen. Ebenso sind kurze Abwehrschreie bei Konfrontationen bekannt. Einzelne Laute erreichen eine Frequenz von 1500 bis 3000 Hz.[16][4][5][6]
Ernährungsweise
Die Nahrung des Pyrenäen-Desmans besteht aus Wirbellosen und Wirbeltieren. Erstere sind mit Flohkrebsen und Insekten vertreten. Unter den Insekten dominieren Köcherfliegen, Steinfliegen, Eintagsfliegen und Taumelkäfer als Bezugsquellen. Häufig sind dabei die Larven von Bedeutung. Bevorzugt werden Tiere, die auf dem Grund leben, seltener solche, die sich in das Bodensediment eingraben. Außerdem meidet der Pyrenäen-Desman hartschalige Lebewesen wie Weichtiere oder rein landbewohnende Tiere.[11][12] Prinzipiell konnte dies bei vergleichenden Untersuchungen an zwei Flüssen im Baskenland, dem Elama und dem Leitzaran, bestätigt werden. Zwischen beiden Lebensräumen ergaben sich starke Übereinstimmungen in der Nutzung der Nahrungsressourcen. Demnach bevorzugt der Pyrenäen-Desman an beiden Fließgewässern zu drei Vierteln Wirbellose, die am Grund leben und sich krabbelnd fortbewegen. Die Laufgeschwindigkeit der Beutetiere, ob langsam oder schnell, spielt dabei eher eine untergeordnete Rolle, wobei aber sesshafte Lebewesen kaum genutzt werden. Fast zwei Drittel der Nahrung besteht wiederum aus Tieren, die zur Schwarmbildung neigen. Die Größe der Beute variiert zu einem ähnlichen Anteil von 8 bis 32 mm, fast ein weiteres Drittel ist kleiner und nur selten größer. Außerdem weist sie zu mehr als 83 % eine gewisse Körperflexibilität auf. In beiden Flussgebieten fängt der Pyrenäen-Desman seine Nahrung überwiegend in schnellfließenden Abschnitten, die mit Steinen oder grobkörnigen Substraten verbunden sind, feinkörnigen Untergrund nutzt er hingegen kaum.[25][4][5][6]
Bei der Analyse von 46 Mageninhalten von Tieren aus der Umgebung von Navarra entfielen rund 47 % auf Eintagsfliegen und 21 % auf Köcherfliegen. Weitere 13 % umfassten Zweiflügler, 9 % Steinfliegen und 2 % Flohkrebse. Bezogen auf die vertilgte Biomasse dominierten Köcherfliegen mit mehr als der Hälfte vor den Zweiflügern und Eintagsfliegen. Vor allem die Köcher- und Eintagsfliegen wurden das ganze Jahr über in hoher Anzahl erbeutet. Stärkere Schwankungen gab es bei den Steinfliegen mit größeren Mengen im Frühjahr, während die Anteile in den anderen Jahreszeiten deutlich zurückgingen. Neben den Wirbellosen wurde in raren Mengen auch Laich von Forellen aufgefunden.[26] Vergleichbare Ergebnisse bezogen auf die Nahrungszusammensetzung und Mengenanteile ermittelten mehrere Analysen von Mageninhalten aus Galicien. Hierbei überwogen im Sommer Köcher- und Eintagsfliegen, im Herbst Flohkrebse, Köcherfliegen und Schnaken nebst Regenwürmern, im Winter wiederum Köcherfliegen.[27][28] Anhand genetischer Untersuchungen von Kotproben aus den französischen Anteilen der Pyrenäen konnten die Beutetiere des Pyrenäen-Desmans genauer bestimmt werden, die sich aus mehr als 100 Gattungen zusammensetzen. Dadurch sind unter den Köcherfliegen Vertreter von Odontocerum und Potamophylax identifiziert. Bei den Eintagsfliegen waren Baetis und Leuctra prominent belegt, bei den Steinfliegen wiederum Perla, verschiedene Angehörige der Gattung Hydropsyche und Protonemura. Insgesamt ergab sich eine sehr reiche Nahrungspalette, die den Pyrenäen-Desman eher als generalistisch charakterisiert.[29][30][31] Neben den Wirbellosen als Hauptbeute enthielten einzelne Nahrungsreste auch Wirbeltiere. Dass diese im Fall von der Forelle oder dem Spanischen Frosch aktiv erbeutet und nachfolgend verzehrt werden, ist auch durch Beobachtungen verifiziert.[32][14][4][5][6]
Seine Beute sucht der Pyrenäen-Desman sowohl am Gewässerboden als auch an der Wasseroberfläche oder an Land. Unter Wasser wird das Substrat am Grund mit den Vorderfüßen oder der langen Nase abgetastet. Kleinere Beute frisst ein Tier sofort, wobei für eine Köcherfliegenlarve etwa 15 bis 20 Kaubewegungen in fünf Sekunden notwendig sind. Größere Beute schleppt der Pyrenäen-Desman an Land. Während der Nahrungsaufnahme steht er auf den Hinterbeinen und bearbeitet die Nahrung mit den Vorderfüßen und der Schnauze. In Gefangenschaft gehaltene Tiere fraßen dargebotene Mäuse mit dem Kopf zuerst. Nicht vertilgte Reste werden verscharrt oder in den Bau verbracht. Verschiedenen Berichten zufolge beträgt die täglich aufgenommene Nahrungsmenge zwischen zwei Drittel und einem Fünftel des eigenen Körpergewichtes.[19][16][15][4][5][6]
Fortpflanzung
Das Fortpflanzungsverhalten des Pyrenäen-Desmans ist nur wenig untersucht. Die Paarung findet wohl vorwiegend im Frühling von Februar bis Juni statt. Bei den Männchen startet die Bildung von Samenzellen bereits im November und dauert bis Mai an. Der Östrus der Weibchen setzt im Januar ein. Trächtige Weibchen werden überwiegend zwischen Februar und Mai beobachtet. Da tragende und säugende weibliche Individuen mehrfach im Jahr beobachtet werden können, ist eine polyöstrische Vermehrung wahrscheinlich. Ein weiterer Indikator hierfür ist auch ein postpartaler Östrus. Weibchen tragen zwischen einem und fünf Embryonen aus, der Durchschnitt beträgt 3,6. Der Nachwuchs wird nach einer Tragzeit von rund dreißig Tagen geboren. Die ersten entwöhnten Jungtiere können bereits im März angetroffen werden. Über die Individualentwicklung ist kaum etwas bekannt, allerdings bringen Jährlinge ihren ersten eigenen Nachwuchs erst im Folgejahr zur Welt.[14] Das maximale Alter wird vermutlich mit fünf Jahren erreicht. Anhand von 83 untersuchten Tieren aus dem nördlichen Spanien verteilen sich die Altersgruppen von einem bis vier Jahre auf jeweils in Viertel (eins bis zwei Jahre) bis ein Fünftel (drei bis vier Jahre). Individuen über fünf Jahre machten hingegen nur rund 6 % aus.[33][34][4][5][6] Weibliche Tiere besitzen Zwitterdrüsen und können als Hermaphroditen auftreten, was auch von einigen Vertretern der Eigentlichen Maulwürfe bekannt ist.[35]
Fressfeinde, Konkurrenten und Parasiten
Es gibt bisher nur wenige Hinweise auf Fressfeinde des Pyrenäen-Desmans. In wenigen Fällen wurde die Erbeutung durch Mauswiesel beobachtet.[4] Genetische Analysen vom Kot des Fischotters verweisen darauf, dass dieser den Pyrenäen-Desman zumindest gelegentlich bejagt.[36] Bezüglich der Lebensraumnutzung und hauptsächlichen Nahrungsressourcen bestehen ökologische Überschneidungen mit der Wasserspitzmaus und der Wasseramsel.[37][12] Die Parasiten des Pyrenäen-Desmans sind bisher nur wenig untersucht. Äußere Schmarotzer wurden mit Milben belegt, darunter Psorergates und Eadiea.[38][39] An inneren Parasiten ließen sich Fadenwürmer wie Paracuaria ebenso Kratzwürmer wie Omphalometra beobachten.[40][41]
Systematik
Innere Systematik der Maulwürfe nach He et al. 2016[42]
|
Der Pyrenäen-Desman ist eine Art aus der Gattung Galemys. Er stellt deren einziges rezentes Mitglied dar, wodurch die Gattung als monotypisch zu betrachten ist. Sowohl die Art als auch die Gattung gehören wiederum zur Familie der Maulwürfe (Talpidae). Zusammen mit dem nahe verwandten Russischen Desman (Desmana) bildet der Pyrenäen-Desman innerhalb der Familie die Tribus der Desmane (Desmanini). Von anderen Maulwürfen unterscheiden sich die Desmane durch ihre semi-aquatische Lebensweise, wodurch sie einige anatomische Besonderheiten aufweisen. Ihr Körperbau ist weniger an eine grabende Aktivität angepasst, was sich unter anderem an dem schlanken Oberarmknochen und den gegenüber den vorderen Füßen größeren hinteren abzeichnet, ebenso wie die unteren Beinabschnitte länger und die Zehen durch Schwimmhäute miteinander verbunden sind oder der Schultergürtel anders konfiguriert ist.[6] Ursprünglich als eigenständige Unterfamilie neben den grabenden Maulwürfen geführt,[43][44] ergaben molekulargenetische Untersuchungen eine nähere Beziehung mit den Eigentlichen Maulwürfen (Talpini) und dem Sternmull (Condylurini). Als enger verwandte Gruppe zu den genannten Triben können die verschiedenen Spitzmulle Ostasiens und Nordamerikas aufgefasst werden. Die Trennung von den anderen Linien der Maulwürfe erfolgte bereits im Oberen Eozän vor rund 37 bis 34 Millionen Jahren. Die Aufspaltung der Desmane in die heutigen Linien setzte aber erst deutlich später, im Verlauf des Mittleren Miozäns vor 13,9 Millionen Jahren ein. Übergeordnet sind alle genannten Maulwurfsgruppen der Unterfamilie der Altweltmaulwürfe (Talpinae) zuzuweisen, innerhalb letzterer stehen zusätzlich noch die Neuweltmaulwürfe (Scalopini). Die Altweltmaulwürfe vereinen sowohl unterirdisch grabende als auch semi-aquatische lebende Tiere aus Eurasien und aus Nordamerika.[45][46][42]
In der Regel werden zwei Unterarten des Pyrenäen-Desmans unterschieden:[3][4][47]
- G. p. pyrenaicus (E. Geoffroy Saint-Hilaire, 1811), Pyrenäen.
- G. p. rufulus (Graells, 1897); Sierra de Guadarrama
Von beiden Vertretern gilt G. p. pyrenaicus als die Nominatform und ist kleiner sowie dunkler gefärbt als G. p. rufulus. Die Unterscheidung ist aber nicht allgemein akzeptiert und ließ sich auch nicht in jedem Fall morphometrisch nachvollziehen.[7][4][48] Genetische Untersuchungen hingegen können zwei parapatrische Gruppen unterscheiden, von denen sich eine (Gruppe A) auf das westliche Kantabrische Gebirge sowie auf das zentrale Spanien und auf Portugal, die andere (Gruppe B) auf das östliche Kantabrische Gebirge, das zentrale Spanien sowie auf die Pyrenäen verteilt. Die Trennung beider Gruppen reicht bis in das Mittelpleistozän vor gut 320.000 Jahren zurück. Zwischen den einzelnen Gruppen besteht kein Austausch mitochdrialer Haplotypen.[46][49][50] Aufgrund dieser Befunde, der die Existenz und die angenommene Verbreitung der postulierten Unterarten nicht bestätigt, wurde der Pyrenäen-Desman im achten Band des Standardwerkes Handbook of the Mammals of the World aus dem Jahr 2018 als monotypisch eingestuft.[6]
Die Gattung Galemys enthält zudem mehrere ausgestorbene Arten:[51][52]
- Galemys kormosi (Schreuder, 1940)
- Galemys semseyi Kormos, 1913
- Galemys sulimskii Rümke, 1985
Die Art Galemys semseyi ist hierbei etwas problematisch. Ursprünglich wurde sie von Theodor Kormos anhand zweier Unterkieferfragmente mit den erhaltenen hinteren Zähnen aus der ungarischen Fundstelle Püspökfürdö eingeführt,[53] ein Exemplar davon ging verloren. Die taxonomisch wichtigen vorderen Zähne sind somit nicht erhalten, morphometrischen Analysen zufolge vermitteln die überlieferten Zähne zwischen den Gattungen Desmana und Galemys.[52] Weiteres von Kormos zu der Art gestelltes Fundmaterial verwies Antje Schreuder im Jahr 1940 zu Desmana kormosi,[54] welches wiederum 45 Jahre später Cornelia G. Rümke zu Galemys ordnete.[51] Eine von Kormos ebenfalls 1913 aufgestellte Art namens Galemys hungaricus, entdeckt an der ungarischen Fundstelle Polgárdi,[53] führte Schreuder 1940 zur Gattung Mygalinia über.[54]
Forschungsgeschichte


Die wissenschaftliche Erstbeschreibung des Pyrenäen-Desmans geht auf Étienne Geoffroy Saint-Hilaire aus dem Jahr 1811 zurück. Geoffroy Saint-Hilaire stand hierfür ein 11 cm langes Exemplar mit 12,5 cm langem Schwanz aus Tarbes im französischen Teil der Pyrenäen zur Verfügung, der Typusregion der Art. Er verglich sein Individuum mit dem Russischen Desman, der bereits mehr als 50 Jahre zuvor von Linnaeus benannt worden war. Als wissenschaftlichen Artnamen gab Geoffroy Saint-Hilaire Mygale pyrenaica an.[55] Die Gattungsbezeichnung Mygale entlieh er aus Georges Cuviers im Jahr 1800 erschienenen ersten Band seiner Leçons d’anatomie comparée, wo sie Cuvier allerdings nur auf einer Übersichtstabelle erwähnte und mit dem Russischen Desman in Beziehung setzte.[56] Ihr liegen die griechischen Wörter μῦς (mys) für „Maus“ und γαλέη (gale) für „Wiesel“ zu Grunde, bedeutet also soviel wie „Mauswiesel“.[57] Bereits vier Jahre vor Cuvier hatte aber Anders Jahan Retzius in seinem Werk Animadversiones in classem mammalium Linneanam den Russischen Desman mit der Gattungsbezeichnung Mygalus in Verbindung gebracht.[58]
Mygale galt lange Zeit als korrekte Gattungsbezeichnung für die Desmane. Sie stand aber in Konflikt mit einer gleichnamigen Gruppe der Spinnen, die im Jahr 1802 durch Pierre André Latreille benannt worden war.[59] Für die Desmane wurden daher auch verschiedene Varianten genutzt. Dazu gehören unter anderem Myogalea, eingeführt von Johann Baptist Fischer im Jahr 1830,[60] oder Myogale, welche wiederum auf Johann Friedrich von Brandt aus dem Jahr 1836 zurückgeht.[61] Vor allem letztere erreichte eine recht weite Verbreitung. Unter anderem befürwortete sie George Edward Dobson in den 1880er Jahren in einer umfangreichen Abhandlung zu den Desmanen im Rahmen seines bedeutenden Werkes zu den Insektenfressern. Seiner Meinung nach war diese Bezeichnung einerseits recht geläufig, andererseits wurde eine Verwechslung mit der gleichfalls Mygale getauften Spinnengattung unterbunden.[62] Im Bezug auf den Russischen Desman sind sowohl Mygale als auch Mygalus und Myogale ungültig, da die durch Johann Anton von Güldenstädt im Jahr 1777 kreierte Bezeichnung Desmana vordatiert. Für den Pyrenäen-Desman führte Johann Jakob Kaup im Jahr 1829 die Bezeichnung Galemys ein.[63] Prinzipiell besitzt Galemys die gleichen Wortbestandteile wie Mygale und bedeutet dementsprechend „Wieselmaus“. Aufgrund der komplexen Vergangenheit von Mygale sprach sich Antje Schreuder im Jahr 1940 für Galemys als korrekte Gattungsbezeichnung für den Pyrenäen-Desman aus.[54] Dies ist seitdem weitgehend akzeptiert.[6]
Die neben der Nominatform einzige, über längere Zeit anerkannte weitere Unterart war mit G. p. rufulus im Jahr 1897 von Mariano de la Paz Graëlls y de la Aguera als Myogalea rufula wissenschaftlich benannt worden. Graëlls hob hierbei die braunere Gesamtfärbung hervor. Als Typusgebiet gan er den Fluss Balsain oberhalb der Venta de los Mosquitos in der Sierra de Guadarrama im nordzentralen Spanien an.[64]
Stammesgeschichte
In ihrer stammesgeschichtlichen Vergangenheit war die Gattung Galemys deutlich weiter verbreitet als es der heutige Pyrenäen-Desman vermuten lässt. Inwiefern Galemys bereits im Oberen Miozän auftrat, ist nicht ganz eindeutig. Funde aus Eichkogel südlich von Wien, die rund 8,7 Millionen Jahre alt sind, werden teilweise der Gattung, aber auch der urtümlicheren Form Archaeodesmana zugewiesen.[65][66] Eindeutig fassen lässt sich Galemys dann im Pliozän. Bedeutend hierbei ist die Fundstelle Węże im südlichen Polen, von wo die Art Galemys sulimskii beschrieben worden ist. Nachgewiesen ist sie über mehrere Schädel- und Kieferfragmente. Es handelt sich hierbei um einen sehr kleinen Vertreter der Desmane. Zeitlich gehören die Fossilfunde in die erste Hälfte des Pliozäns.[51] Die Art wurde nach Andrzej Sulimski benannt, der bereits Ende der 1950er Jahre die Fossilreste von Węże vorgestellt hatte.[67][68]
Nur wenig jünger als die polnischen Funde sind jene von Beremend und Csarnota in Ungarn. Das sehr umfangreiche Material, bestehend aus Schädel- und Gliedmaßenteilen, gehört zu Galemys kormosi. Die Art übertraf den heutigen Pyrenäen-Desman ein wenig an Größe. Der Namenspatron Theodor Kormos veröffentlichte die ersten Funde in den 1930er Jahren.[54] Galemys kormosi erreichte im ausgehenden Pliozän und im Altpleistozän eine recht weite Verbreitung. Vergleichsweise häufig ist die Art im südlichen Europa belegt. Genannt werden kann hier unter anderem die Lokalität Coste San Giacomo im zentralen Italien aus der Frühphase des Altpleistozäns.[69] Auf der Iberischen Halbinsel finden sich zahlreiche Fundpunkte wie etwa in der Umgebung von Granada oder von Baza, beide im Süden Spaniens.[70][51] Von Bedeutung ist auch die Fundregion der Sierra de Atapuerca im Norden des Landes, herausragend wegen ihrer archäologischen und frühmenschlichen Befunde. Galemys kormosi wurde hier an den ältesten Fundstellen wie der Sima del Elefante mehrfach registriert, welche ein Alter von etwa 1,2 bis 1,0 Millionen Jahren aufweist. Jüngere Bereiche wie die Gran Dolina mit Alterswerten um 780.000 Jahren und jünger führen die Art hingegen nicht mehr, es treten jedoch Fossilreste von Galemys auf, die nicht sicher einer bestimmten Art zugeordnet werden können.[71][72][73] Darüber hinaus erreichte die Art auch das Gebiet nördlich der Alpen. Noch in das ausgehende Pliozän gehören die Reste aus dem Tagebau Hambach und dem Tagebau Frechen, die beide zum Rheinischen Braunkohlerevier gehören. Aus ersterem liegen rund 110 Zähne vor, die sich auf drei Arten der Desmane verteilen, darunter auch Galemys kormosi.[74][75] Des Weiteren sind zahlreiche Zähne an der bedeutenden altpleistozänen Fundstelle Tegelen in den Niederlanden aufgedeckt worden.[51][76] Ein vergleichbares Alter zu Tegelen haben Zahnfunde aus dem Meeresarm Oosterschelde, die dort unter Wasser geborgen wurden.[77] Weitere Funde sind aus England bekannt, hier überwiegend aus Norfolk. Sie umfassen Gebissreste und Teile des Bewegungsapparates.[78][79] Aus dem östlichen Mitteleuropa wären die Fossilreste von Rebielice Krolewskie im südlichen Polen zu nennen.[51] Weiter im Osten müssen ursprünglich als zu Galemys gehörig angesprochene Fossilfunde aus der Ukraine mittlerweile in ihrer Stellung als unsicher eingestuft werden, da die aufgefundenen Zähne deutlich größer sind als die der bekannten Vertreter.[80][81]
Ein Unterkieferfragment aus Püspökfürdö in Ungarn mit ebenfalls altpleistozänem Alter wird der Art Galemys semseyi zugeordnet. Sie ist bisher nur von dieser Fundstelle nachgewiesen, einzelne Gebssfragmente aus der gleichen Region könnten ebenfalls dazugehören. Anhand der wenigen bekannten Funde kann auf ein Tier von der Größe des heutigen Pyrenäen-Desmans geschlossen werden.[53][54][51][82] Für das Jungpleistozän sind wiederum aus El Portalón in der Fundregion von Atapuerca einige Funde des heutigen Pyrenäen-Desmans verzeichnet. Das Gebiet befindet sich innerhalb des rezenten Verbreitungsgebietes.[83]
Bedrohung und Schutz
Zu den Bedrohungen des Pyrenäen-Desmans zählen die Verschmutzung der Gewässer in Folge von Landwirtschaft und Industrie sowie die Zerstückelung seines Lebensraums durch Störung und Isolierung des Gewässernetzes etwa bei Errichtung von Kraftwerken oder Talsperren. Die daraus folgende Vereinzelung verschiedener Populationen hat zu einem deutlichen Inzest geführt. Außerdem werden die Tiere teils aktiv durch Fischer verfolgt, die in ihnen Nahrungskonkurrenten sehen. Hinzu kommt die Verdrängung durch den Amerikanischen Nerz, der aus Pelzfarmen ausgebrochen ist oder freigelassen wurde und eine ähnliche ökologische Nische besetzt wie der Pyrenäen-Desman. Insgesamt ist der Lebensraum der Tiere sehr anfällig für anthropogene Einflüsse. Die IUCN hat die Art daher bereits im Jahr 2008 als „gefährdet“ (vulnerable) und Ende 2021 als „stark gefährdet“ (endangered) eingestuft. Der Bestand hat sich laut Angaben der Naturschutzorganisation seit 2011 um die Hälfte verringert. Nach Untersuchungen im nordöstlichen Portugal ging der Bestand hier zwischen 1996 und 2015 um über 63 % zurück.[84] Als wichtige Erhaltungsmaßnahmen werden ein angemessenes Management der Gewässernetzwerke, die Wiederherstellung von Lebensräumen und Kenntnisgewinn über bedrohte Populationen angesehen. Außerdem dient der Pyrenäen-Desman als Aushängeschild für die Förderung der Erhaltung von Flusslandschaften. Forschungen sind vor allem im Bereich der Minderung der schädlichen Einwirkungen durch Wasserkraftwerke und Gewässerwirtschaft notwendig, ebenso muss die Minimalgröße überlebensfähiger Populationen und die genetische Variabilität einzelne Gruppen bestimmt werden. Teile des Verbreitungsgebietes des Pyrenäen-Desmans sind als Nationalparks geschützt, so etwa durch den Nationalpark Pyrenäen in Frankreich und den Nationalpark Picos de Europa in Spanien.[85]
Von der Europäischen Union wird der Pyrenäen-Desman in den Anhängen II und IV der FFH-Richtlinie geführt und gilt somit als streng zu schützende Art von gemeinschaftlichem Interesse, für deren Erhalt besondere Schutzgebiete ausgewiesen werden müssen.[86]
Literatur
- I. I. Barabasch-Nikiforow: Die Desmane. Neue Brehm Bücherei 474, Wittenberg, 1975, S. 1–100
- E.-A. Juckwer: Galemys pyrenaicus (Geoffroy, 1811) – Pyrenäen-Desman. In: J. Niethammer und F. Krapp (Hrsg.): Handbuch der Säugetiere Europas. Wiesbaden, 1990, S. 79–92
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 605–606) ISBN 978-84-16728-08-4
- Ronald M. Nowak: Walker's Mammals of the World. 2 Bände. 6. Auflage. The Johns Hopkins University Press, Baltimore MD u. a. 1999, ISBN 0-8018-5789-9
- Jorge M. Palmeirim und Robert S. Hoffmann: Galemys pyrenaicus. Mammalian Species 207, 1983, S. 1–5
Einzelnachweise
- P. B. Richard: The sensorial world of the Pyrenean desman, Galemys pyrenaicus. Acta Zoologica Fennica 173, 1985, S. 255–258
- Kenneth C. Catania: Epidermal Sensory Organs of Moles, Shrew-Moles, and Desmans: A Study of the Family Talpidae with Comments on the Function and Evolution of Eimer’s Organ. Brain Behavior and Evolution 56, 2000, S. 146–174
- I. I. Barabasch-Nikiforow: Die Desmane. Neue Brehm Bücherei 474, Wittenberg, 1975, S. 1–100
- Jorge M. Palmeirim und Robert S. Hoffmann: Galemys pyrenaicus. Mammalian Species 207, 1983, S. 1–5
- E.-A. Juckwer: Galemys pyrenaicus (Geoffroy, 1811) – Pyrenäen-Desman. In: J. Niethammer und F. Krapp (Hrsg.): Handbuch der Säugetiere Europas. Wiesbaden, 1990, S. 79–92
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 605–606) ISBN 978-84-16728-08-4
- J. Gonzáles-Esteban, E. Castién und J. Gosálbez: Morphological and colour variation in the Pyrenean Desman Galemys pyrenaicus (Goeffroy, 1811). Zeitschrift für Säugetierkunde 64, 1999, S. 1–11
- Walter Poduschka und Bernard Richard: Hair Types in the Fur of the Pyrenean Desman (Galemys pyrenaicus) Geoffroy, 1811 (Insectivora: Talpidae: Desmaninae). Sitzungsberichte der Akademie der Wissenschaften mathematisch-naturwissenschaftliche Klasse 194, 1985, S. 39–44
- В. М. Малыгин und Д. Л. Заславский: Особенности кариотипов Русской (Desmana moschata) и Пиренейской (Galemys pyrenaicus) выхухолей в свази с вопросом о положении етих видов в семействе Talpidae (Mammalia, Insectivora). Зоологический Журнал 65, 1986, S. 1435–1437
- Carlos Nores und Enzesto Gracía Alvarez: Valoración del habitat de Galemys pyrenaicus en España. Council of Europe (Hrsg.): Seminar on the biology and conseration of European desmans and water shrews (Galemys payrenaicus, Desmana moschata, Neomys spp.) Ordesa, Spain, 7-11 June 1995. Strasbourg, 1995, S. 58–61
- Amaiur Esnaola, Vanessa Mata, Aitor Arrizabalaga-Escudero, Jorge González-Esteban, Aitor Larrañaga, Hugo Rebelo, Arturo Elosegi und Joxerra Aihartza: It is the ambience, not the menu. Prey availability does not drive habitat selection by the endangered Pyrenean desman. Aquatic Conservation: Marine and Freshwater Ecosystems 31, 2021, S. 1859–1872, doi:10.1002/aqc.3563
- Alejandro López‑de Sancha, Romero Roig, Pere Aymerich, Anna Vila‑Gispert und Helena Guasch: Trophic competition in a guild of insectivorous semi‑aquatic vertebrates in a Pyrenean headwater stream: diet specialisation in the endangered Galemys pyrenaicus. Mammalian Biology, 2022, doi:10.1007/s42991-022-00265-z
- C. Nores, F. Ojeda, A. Ruano. Villate, J. González, J. M. Cano und E. García: Estimating the population density of Galemys pyrenaicus in four Spanish rivers. Journal of Zoology 246, 1998, S. 454–457
- A. Peyre: Ecologie et Biogeographie du Desman (Galemys pyrenaicus G.) dans les Pyrénées Francaises. Mammalia 20, 1956, S. 405–418
- P. B. Richard und C. Micheau: Le carrefour tracheen dans l'adaptation du desman des Pyrenees (Galemys pyrenaicus) a Ia vie dulcaquicole. Mammalia 39, 1975, S. 467–477
- Günther Niethammer: Beobachtungen am Pyrenäen-Desman, Galemys pyrenaica. Bonner Zoologische Beiträge 21 (3/4), 1970, S. 157–182
- R. Bauchot, C. Buisseret, Y. Leroy und P. B. Richard: L'équipment sensoriel de la trompe du desman des Pyrénées (Galemys pyrenaicus, Insectivora, Talpidae). Mammalia 37, 1973, S. 17–24
- P. B. Richard: Le desman des Pyrénées (Galemys pyrenaicus). Mode de vie. Univers sensoriel. Mammalia 37, 1973, S. 1–16
- P. B. Richard und A. Vallette Viallard: Le desman des Pyrénées (Galemys pyrenaicus): premières notes sur sa biologie. Revue d’Ecologie, Terre et Vie 23, 1969, S. 225–245
- R. David Stone: The activity patterns of the Pyrenean desman (Cufemys pyvenuicus) (Insectivora: Talpidae), as determined under natural conditions. Journal of Zoology 213, 1987, S. 95–106
- Y. Melero, P. Aymerich, G. Santulli und J. Gosàlbez: Activity and space patterns of Pyrenean desman (Galemys pyrenaicus) suggest non-aggressive and non-territorial behaviour. European Journal of Wildlife Research 60, 2014, S. 707–715, doi:10.1007/s10344-014-0838-8
- Yolanda Melero, Pere Aymerich, Juan Jose Luque-Larena und Joaquim Gosàlbez: New insights into social and space use behaviour of the endangered Pyrenean desman (Galemys pyrenaicus). European Journal of Wildlife Research 58, 2012, S. 185–193, doi:10.1007/s10344-011-0561-7
- R. David Stone: The social ecology of the Pyrenean desman (Gdemys pyremicus) (Insectivora: Talpidae), as revealed by radiotelemetry. Journal of Zoology 212, 1987, S. 117–129
- R. David Stone und Martyn L. Gorman: Social organization of the European mole (Talpa europaea) and the Pyrenean desman (Galemyspyrenaicus). Mammal Review 15 (1), 1985, S. 35–42
- Amaiur Esnaola, Aitor Larrañaga, Jorge González-Esteban, Arturo Elosegi und Joxerra Aihartza: Using biological traits to assess diet selection: the case of the Pyrenean desman. Journal of Mammalogy, 2023, doi: 10.1093/jmammal/gyad061
- Enrique Castien und Joaquim Gosálbez: Diet óf Galemys payrenaicus (Geoffroy, 1811) in the North of the Iberian Peninsula. Netherlands Journal of Zoology 45 (3/4), 1995, S. 422–430
- Jesus Santamarina und José Guitan: Quelques données sur le régime alimentaire du desman (Galemys pyrenaicus) dans le nord-ouest de l'Espagne. Mammalia 52 (3), 1988. S. 301–307
- Jesus Santamarina: Feeding ecology of a vertebrate assemblage inhabiting a stream of NW Spain (Riobo; Ulla basin). Hydrobiologia 252, 1993, S. 175–191
- François Gillet, Marie-Laure Tiouchichine, Maxime Galand, Frédéric Blanc, Mélanie Némozc, Stéphane Aulagnier und Johan R. Michaux: A new method to identify the endangered Pyrenean desman (Galemys pyrenaicus) and to study its diet, using next generation sequencing from faeces.Mammalian Biology 80, 2015, S. 505–509, doi:10.1016/j.mambio.2015.08.002
- Marjorie Biffi, François Gillet, Pascal Laffaille, Fanny Colas, Stéphane Aulagnier, Frédéric Blanc, Maxime Galan, Marie-Laure Tiouchichine, Mélanie Némoz, Laëtitia Buisson und Johan R. Michaux: Novel insights into the diet of the Pyrenean desman (Galemys pyrenaicus) using next-generation sequencing molecular analyses. Journal of Mammalogy 98 (5), 2017, S. 1497–1507, doi:10.1093/jmammal/gyx070
- Amaiur Esnaola, Aitor Arrizabalaga-Escudero, Jorge González-Esteban, Arturo Elosegi und Joxerra Aihartza: Determining diet from faeces: Selection of metabarcoding primers for the insectivore Pyrenean desman (Galemys pyrenaicus). PLoS ONE 13 (12), 2018, S. e0208986, doi:10.1371/journal.pone.0208986
- Lorenzo Quaglietta und Pedro Beja: Direct observations of vertebrate killing and consumption by the endangered Pyrenean desman (Galemys pyrenaicus). Mammalia 83 (5), 2019, S. 479–482, doi:10.1515/mammalia-2018-0111
- P. B. Richard: Détermination de l’âge et de la longévité chez le desman des Pyrénées (Galemys pyrenaicus). Revue d’Ecologie, Terre et Vie 2, 1976, S. 181–192
- Jorge González-Esteban, Idoia Villate, Enrique CastiénIsabel rey und Joaquim Gosálbez: Age determination of Galemys pyrenaicus. Acta Theriologica 47 (1), 2002, S. 107–112
- F. David Carmona, Masaharu Motokawa, Masayoshi Tokita, Kimiyuki Tsuchiya, Rafael Jiménez und Marcelo Sánchez-Villagra: The Evolution of Female Mole Ovotestes Evidences High Plasticity of Mammalian Gonad Development. Journal of Experimental Zoology (Mol Dev Evol) 310 B, 2008, S. 259–266
- F. Gillet, A. Mouton, A. André, S. Vanoutryve, F. Blanc, M. Némoz, P. Fournier, C. Fournier-Chambrillon, D. Marc, M. Biffi, P. Lafaille, L. Buisson und J. R. Michaux: Evidence of predation and possible competition among three semi aquatic species, the endangered Pyrenean desman (Galemys pyrenaicus), the aquatic shrew (Neomys fodiens) and the European otter (Lutra lutra) using next-generation sequencing methods from faeces. In: Conference on Conservation Genomics 3 – 6 May 2016. ESF – CIBIO-InBIO – U Porto, Campus de Vairão, Vairão, Portugal, 2016, S. 46
- Marjorie Biffi, Pascal Laffaille und Laëtitia Buisson: Local habitat preferences of a semi-aquatic mammal, the Pyrenean desman Galemys pyrenaicus. Mammalia 84 (1), 2020, S. 50–62
- F. S. Lukoschus: Psorergates desmanae sp. nov., eine neue Krätzmilbe von Galemys pyrenaicus (Psorergatidae: Trombidiformes). Bulletin du Muséum d'histoire Naturelle 40 (1), 1968, S. 125–131
- A. Fain und J. O. Whitaker, Jr.: Two new species of Myobiidae from North American mammals (Acarina). Bulletin et Annales de la Société entomologique de Belgique 111, 1975, S. 57–65
- F. Alvarez, H. Gijón-Botella, P. Quinteiro, J. Rey, F. López-Román und M. L. Sanmartiín: Paracuaria hispanica n. sp. (Nematoda: Acuariidae), a stomach parasite of the pyrenean desman Galemys pyrenaicus Geoffr. (Insectivora: Talpidae), with a redefinition of the genus Paracuaria Rao, 1951. Systematic Parasitology 29, 1994, S. 105–112
- Claude Vaucher: Sur quelques Trématodes parasites de Chiroptères et d'Insectivores. Bulletin de la Société Neuchâteloise des Sciences Naturelles 98, 1975, S. 17–25
- Kai He, Akio Shinohara, Kristofer M. Helgen, Mark S. Springer, Xue-Long Jiang und Kevin L. Campbell: Talpid Mole Phylogeny Unites Shrew Moles and Illuminates Overlooked Cryptic Species Diversity. Molecular Biology and Evolution 34 (1), 2016, S. 78–87
- Leigh van Valen: New Paleocene Insectivores and Insectivore classification. Bulletin of the American Museum of Natural History 135, 1967, S. 217–284
- J. Howard Hutchison: Notes on type specimens of European Miocene Talpidae and a tentative Classification of Old World Tertiary Talpidae (Insectivora: Mammalia). Geobios 7 (3), 1974, S. 211–256
- María Teresa Cabria, Jonathan Rubines, Benjamín Gómez-Moliner und Rafael Zardoya: On the phylogenetic position of a rare Iberian endemic mammal, the Pyrenean desman (Galemys pyrenaicus). Gene 375, 2006, S. 1–13, doi:10.1016/j.gene.2006.01.038
- Javier Igea, Pere Aymerich, Angel Fernández-González, Jorge González-Esteban, Asunción Gómez, Rocío Alonso, Joaquim Gosálbez und Jose Castresana: Phylogeography and postglacial expansion of the endangered semi-aquatic mammal Galemys pyrenaicus. BMC Evolutionary Biology 13, 2013, S. 115, doi:10.1186/1471-2148-13-115
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. A taxonomic and geographic Reference. 2 Bände. 3. Auflage. Johns Hopkins University Press, Baltimore MD 2005, ISBN 0-8018-8221-4 ()
- María José López-Fuster, Rosa García-Perea, Rosa Fernández-Salvador, Julio Gisbert und Jacint Ventura: Craniometric variability of the Iberian desman, Galemys pyrenaicus (Mammalia: Erinaceomorpha: Talpidae). Folia Zoologica 55 (1), 2006, S. 29–42
- Marina Querejeta, Jorge González-Esteban, Asunción Gómez, Angel Fernández-González, Pere Aymerich, Joaquim Gosálbez, Lídia Escoda, Javier Igea und Jose Castresana: Genomic diversity and geographical structure of the Pyrenean desman. Conservation Genetics 17, 2016, S. 1333–1344, doi:10.1007/s10592-016-0865-y
- Lídia Escoda und Jose Castresana: The genome of the Pyrenean desman and the effects of bottlenecks and inbreeding on the genomic landscape of an endangered species. Evolutionary Applications 14, 2021, S. 1898–1913, doi:10.1111/eva.13249
- Cornelia G. Rümke: A review of fossil and recent Desmaninae (Talpidae, Insectivora). Utrecht Micropaleontological Bulletins, Special Publication 4, 1985, S. 1–241
- Raef Minwer-Barakat, Antonio García-Alix, Elvira Martín-Suárez und Matthijs Freudenthal: Early Pliocene Desmaninae (Mammalia, Talpidae) from Southern Spain and the Origin of the Genus Desmana. Journal of Vertebrate Paleontology 40 (5), 2020, S. e1835936, doi:10.1080/02724634.2020.1835936
- Theodor Kormos: Trois nouvelles espèces fossiles des Desmans en Hongrie. Annales Musei Nationalis Hungarici 11, 1913, S. 135–145
- Antje Schreuder: A revision of the fossil water-moles (Desmaninae). Archives Néerlandais de Zoologie 4, 1940, S. 201–233
- Etienne Geoffroy Saint-Hilaire: Memoire sur les especes des genres Musaraigne et Mygale. Annales du Muséum d'histoire naturelle 17, 1811, S. 169–194 ()
- Georges Cuvier: Leçons d’anatomie comparée. Tome 1. Paris, 1800, S. 1–521 ()
- T. S. Palmer: Index Generum Mammalium: A List of the Genera and Families of Mammals. North American Fauna 23, 1904, S. 437 ()
- Anders Jahan Retzius: Animadversiones in classem mammalium Linneanam. Lund, 1796, S. 1–22 (S. 22) ()
- Pierre André Latreille: Histoire naturelle, générale et particulière des Crustacés et des Insectes. Tome Troisième. Paris, 1802, S. 1–467 (S. 48–59) ()
- Johann Baptist Fischer: Addenda, emendanda et index ad Synopsis Mammalium. Stuttgart, 1830, S. 1–752 (S. 250) ()
- Johann Friedrich von Brandt: Bemerkungen über den inneren Bau des Wuychuchol (Myogale moschaqta), im Vergleich zu dem des Maulwurfs und der Spitzmaus (Sorex araneus). Archiv für Naturgeschichte 2 (1), 1836, S. 176–182 ()
- George Edward Dobson: A Monograph of the Insectivora, systematic and anatomical. Part II. London, 1883, S. 126–172 ()
- Johann Jakob Kaup: Skizzirte Entwickelungs-Geschichte und Natürliches System der Europäischen Thierwelt. Darmstadt und Leipzig, 1829, S. 1–203 (S. 119) ()
- Mariano de la Paz Graells: Fauna mastodológica Ibérica. Memorias de la Real Academia de Ciencias Exactas Fisicas y Naturalesde Madrid 17, 1897, S. 1–806 (S. 460–461) ()
- Gernot Rabeder: Die Wirbeltierfauna aus dem Alt-Pliozän (O-Pannon) vom Eichkogel bei Mödling (NÖ.) I. Allgemeines — II. Insectivora. Annalen des Naturhistorischen Museums in Wien 74, 1970, S. 589–595
- Reinhard Ziegler: Insectivores (Lipotyphla) and bats (Chiroptera) from the Late Miocene of Austria. Annalen des Naturhistorischen Museums Wien 107A, 2006, S. 93–196
- Andrzej Sulimski: Pliocene insectivores from Węże. Study on the Tertiary bone breccia fauna from Węże near Dzialoszyn in Poland. Part XI. Acta Palaeontologica Polonica 4 (2), 1959, S. 119–173
- Andrzej Sulimski: Supplementary studies on the insectivores from Węże 1 (Poland). Study on the Tertiary bone breccia fauna from Węże near Dzialoszyn in Poland. Part xVII. Acta Palaeontologica Polonica 7 (3/4), 1962, S. 441–498
- Fabio Bona, Luca Bellucci und Raffaele Sardella: The Gelasian (Late Villanyan-MN17) diversified micromammal assemblage with Mimomys pliocaenicus from Coste San Giacomo (Anagni basin, central Italy), taxonomy and comparison with selected European sites. Hystrix, the Italian Journal of Mammalogy 26, 2015, S. 141–151, doi:10.4404/hystrix-26.2-11441
- Antonio García-Alix, Raef Minwer-Barakat, Elvira Martín Suárez und Matthijs Freudenthal: Small mammals from the early Pleistocene of the Granada Basin, southern Spain. Quaternary Research 72, 2009, S. 265–274, doi:10.1016/j.yqres.2009.06.004
- Raquel López Antoñanzas und Gloria Cuenca-Bescós: The Gran Dolina site (Lower to Middle Pleistocene, Atapuerca, Burgos, Spain): new palaeoenvironmental data based on the distribution of small mammals. Palaeogeography, Palaeoclimatology, Palaeoecology 186, 2002, S. 311–334
- Gloria Cuenca-Bescós, Juan Rofes, Juan Manuel López-García, Hugues-Alexandre Blain, Raquel Rabal-Garcés, Victor Sauqué, Juan Luis Arsuaga, José María Bermúdez de Castro und Eudald Carbonell: The small mammals of Sima del Elefante (Atapuerca, Spain) and the first entrance of Homo in Western Europe. Quaternary International 295, 2013, S. 28–35, doi:10.1016/j.quaint.2011.12.012
- Gloria Cuenca-Bescós, Hugues-Alexandre Blain, Juan Rofes, Iván Lozano-Fernández, Juan Manuel López-García, Mathieu Duval, Julia Galán und Carmen Núñez-Lahuerta: Comparing two different Early Pleistocene microfaunal sequences from the caves of Atapuerca, Sima del Elefante and Gran Dolina (Spain): Biochronological implications and significance of the Jaramillo subchron. Quaternary International 389 (2), 2015, S. 148–158, doi:10.1016/j.quaint.2014.12.059
- Roland Kersting, Thomas Mörs: Water moles (Desmaninae, Lipotyphla) from the Pliocene Reuver Clay of north-west Germany. In: Lutz C. Maul und Ralf Dietrich Kahlke (Hrsg.): 18th International Senckenberg Conference; VI International Palaeontological Colloquium in Weimar. Late Neogene and Quaternary biodiversity and evolution: Regional developments and interregional correlations. Weimar (Germany). 25th-30th April, 2004. Terra Nostra Schriften der Alfred-Wegener-Stiftung, 2004, S. 135–136
- Jelle W. F. Reumer: Insectivora (Mammalia: Soricidae, Talpidae) from the Late Pliocene of Frechen, Germany. Deinsea 2, 1995, S. 9–15
- Lars W. van den Hoek Ostende und John de Vos: A century of research on the classical locality of Tegelen (province of Limburg, The Netherlands). Courier Forschungsinstitut Senckenberg 256, 2006, S. 291–304
- Jelle W. F. Reumer, Joop C. van Veen, Albert J. van der Meulen, Leen W. Hordijk und John de Vos: The first find of small mammals (Desmaninae, Arvicolidae) from the Early Pleistocene Oosterschelde fauna in The Netherlands. Deinsea 4, 1998, S. 41–45
- David L. Harrison, J. J. Bates und J. D. Clayden: On the Occurrence of Galemys kormosi (Schreuder, 1940) (Insectívora: Desmaninae) in the British Lower Pleistocene. Acta Theriologica 33 (26), 1988, S. 369–378
- David L. Harrison und Simon A. Parfitt: Fossil remains of shrews (Soricomorpha: Soricidae) and desmans (Talpidae: Desmaninae) from Norfolk, England, with biostratigraphic implications for the Plio-Pleistocene boundary in East Anglia. Acta zoologica cracoviensia 52A (1–2), 2009, S. 61–79, doi:10.3409/azc.52a_1-2.61-79
- Barbara Rzebik-Kowalska, Leonid I. Rekovets: Recapitulation of data on Ukrainian fossil insectivore mammals (Eulipotyphla, Insectivora, Mammalia). Acta zoologica cracoviensia 58 (2), 2015, S. 137–171, doi:10.3409/azc.58_2.137
- Barbara Rzebik-Kowalska, Leonid I. Rekovets: New data on Eulipotyphla (Insectivora, Mammalia) from the Late Miocene to the Middle Pleistocene of Ukraine. Palaeontologia Electronica 19 (1), 2016, S. 9A ()
- Barbara Rzebik-Kowalska: Insectivores (Mammalia) from the Early and earlv Middle Pleistocene of Betfia in Romania. II. Erninaceidae Bonaparte, 1838 and Talpidae Gray, 1825. Acta zoologica cracoviensia 43 (1/2), 200, S. 55–77
- Juan Manuel López-García, Hugues-Alexandre Blain, Gloria Cuenca-Bescós, María Blanca Ruiz-Zapata, Miriam Dorado-Valiñoc, Maria José Gil-García, Ana Valdeolmillos, Ana Isabel Ortega, Jose Miguel Carretero, Juan Luis Arsuaga, Jose María Bermúdez de Castro und Eudald Carbonell: Palaeoenvironmental and palaeoclimatic reconstruction of the Latest Pleistocene of El Portalón Site, Sierra de Atapuerca, northwestern Spain. Palaeogeography, Palaeoclimatology, Palaeoecology 292, 2010, S. 453–464, doi:10.1016/j.palaeo.2010.04.006
- L. Quaglietta, J. Paupério, F. M. S. Martins, P. C. Alves und P. Beja: Recent range contractions in the globally threatened Pyrenean desman highlight the importance of stream headwater refugia. Animal Conservation 21 (6), 2018, S. 515–525, doi:10.1111/acv.12422
- L. Quaglietta: Galemys pyrenaicus (amended version of 2021 assessment). The IUCN Red List of Threatened Species 2022. e.T8826A214429993 (); zuletzt aufgerufen am 11. November 2022
- EU: Amtsblatt der Europäischen Gemeinschaften Nr. L 206 /7 II: Richtlinie 92/43/EWG des Rates. 1992 ().
Weblinks
- Galemys pyrenaicus in der Roten Liste gefährdeter Arten der IUCN 2021. Eingestellt von: L. Quaglietta, 2022. Abgerufen am 11. November 2022.