Desmane
Die Desmane (Desmanini), auch Bisamspitzmäuse genannt, sind eine Gattungsgruppe (Tribus) der Familie der Maulwürfe (Talpidae). Sie umfasst mit dem Russischen Desman und dem Pyrenäen-Desman zwei rezente Arten. Die Tiere sind von allen Maulwürfen am besten ans Wasserleben angepasst. Anzeichen dafür finden sich unter anderem in dem gedrungenen Körperbau mit langer Schnauze und verschließbaren Nasenlöchern, dem langen und teilweise oder vollständig abgeplatteten Schwanz, den gegenüber den Vorderfüßen sehr großen Hinterfüßen und den Schwimmhäuten zwischen den Zehen. Das dichte Fell hat eine überwiegend dunkle Färbung. Das Verbreitungsgebiet der Desmane ist zweigeteilt mit einem Bereich im östlichen Europa und einem weiteren in den Pyrenäen sowie im Norden der Iberischen Halbinsel. Die Lebensräume bestehen aus fließenden oder stehenden Gewässern mit reicher Wirbellosenfauna und dichter Ufervegetation. Die Desmane sind meist nachtaktiv und verbringen ihre Zeit schwimmend im Wasser, wo sie auch ihre Nahrung suchen. Diese umfasst Wirbellose, mitunter auch Wirbeltiere und zum Teil auch Pflanzen. Zum Rückzug nutzen die Tiere selbst gegrabene Gänge oder natürliche Höhlen. Abweichend von anderen Maulwürfen sind die Desmane eher sozial und nicht aggressiv. Die Fortpflanzung kann ganzjährig erfolgen, konzentriert sich aber häufig auf das Frühjahr. Wissenschaftlich bekannt sind die Desmane seit wenigstens dem beginnenden 17. Jahrhundert. In der forschungsgeschichtlichen Vergangenheit waren sie weitgehend unter der Gattungsbezeichnung Mygale bekannt. Der heute gebräuchliche wissenschaftliche Name Desmanini für die Gattungsgruppe wurde im Jahr 1912 geprägt. Fossile Nachweise der Desmane reichen bis in das Oligozän zurück. Sie fanden sich, abweichend von der heutigen Verbreitung, über einen größere Bereich Eurasiens verteilt sowie in Nordamerika. Die Funde lassen sich mehreren ausgestorbenen Gattungen zuweisen. Der Bestand der beiden heutigen Arten gilt als gefährdet, was neben der Gewässerverschmutzung auch in der illegalen Jagd und in der Konkurrenz mit eingeschleppten Tierarten begründet ist.
| Desmane | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Russischer Desman (Desmana moschata) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Desmanini | ||||||||||||
| Thomas, 1912 |
Merkmale
Habitus

.jpg.webp)
Die beiden Arten der Desmane gehören zu den größten Vertretern der Maulwürfe. Der Pyrenäen-Desman (Galemys pyrenaicus) ist mit einer Kopf-Rumpf-Länge von 10,6 bis 12,4 cm, einer Schwanzlänge von 11,8 bis 14,8 cm und einem Gewicht von 51 bis 79 g die kleinere Form. Der Russische Desman (Desmana moschata) repräsentiert mit entsprechenden Werten von 18,5 bis 24,0 cm, 18,0 bis 21,0 cm und 370 bis 440 g die größere. Das äußere Erscheinungsbild der Desmane wirkt aufgrund des gedrungenen Körpers sehr kompakt. Der Kopf ist konisch geformt. Er sitzt auf einem für Maulwürfe charakteristisch sehr kurzen Hals. Die Nase ist ausgesprochen lang und überaus beweglich, wodurch sie rüsselartig wirkt. Die Augen sind klein und die Ohren bleiben im Fell verborgen. Der lange Schwanz ist beim Russischen Desman vollständig, beim Pyrenäen-Desman nur an der Spitze seitlich abgeplattet. Zwischen den Zehen sind insbesondere an den Hinterbeinen Schwimmhäute ausgeprägt. Außerdem übertreffen die Hinterfüße die Vorderfüße deutlich an Größe, was untypisch für Maulwürfe ist. Entsprechend zu diesen tragen die jeweils fünf Strahlen der Vorder- und Hinterfüße aber kräftige Krallen. Das Fell weist an der Oberseite einen dunklen zumeist grau- bis rotbraunen Farbton auf, die Unterseite ist häufig heller. Es hat eine dichte Textur bestehend aus der Unterwolle und dem Deckhaar. Die Fußkanten sind zusätzlich mit einem Saum an borstenartigen Haaren versehen. An der Schnauze und am Kinn kommen zudem zahlreiche Vibrissen vor. Der Schwanz ist eher spärlich behaart und teilweise von Schuppenringen bedeckt, die besonders deutlich am Schwanzansatz ausgebildet sind. An der Schwanzunterseite verfügen die Desmane über eine Duftdrüse, aus der sie ein moschusähnliches Sekret absondern.[1][2][3]
Schädel- und Gebissmerkmale
Der Schädel der Desmane ist konisch geformt mit einem lang ausgestreckten Rostrum. Er besitzt in Seitenansicht eine kantige Form mit einer flach verlaufenden Stirnlinie und einem nahezu senkrecht stehenden Hinterhauptsbein. An diesem sind die Gelenkflächen für den Halswirbelansatz nur wenig hervorgewölbt. Auf dem Scheitelbein ist ein schwacher Scheitelkamm ausgebildet. Die seitlich am Schädel verlaufenden Jochbögen haben eine schlanke Gestalt. Das Foramen infraorbitale ist relativ groß. An der Schädelunterseite zeigt sich am Gaumen das Foramen incisivum vergleichsweise groß. Der Gaumen selbst reicht bis kurz hinter den letzten Mahlzahn. Die Paukenblasen sind gut entwickelt. Die Gehörknöchelchen des Mittelohrs besitzen keine auffälligen Vergrößerungen, was vor allem für den Hammer zutrifft und möglicherweise mit der Wahrnehmung höherfrequenter Töne in Verbindung steht.[4][5][6] Der Unterkiefer ist ähnlich dem Schädel langgestreckt. Der Kronenfortsatz ragt steil auf und erreicht fast die Höhe des Scheitelkamms. Dagegen ist der Gelenkfortsatz eher kurz, das Unterkiefergelenk kragt seitlich weit aus, was sich auch an der Glenoidgrube an der Schädelunterseite widerspiegelt.[7][1]
| Gattung | Zahnformel | Zahnanzahl |
|---|---|---|
| Desmana | oder | 44 |
| Galemys | 44 |


Das Gebiss der Desmane besteht aus 44 Zähnen, was der ursprünglichen Zahnanzahl bei den Höheren Säugetiere entspricht. Für beide Arten käme demnach eine Zahnformel in Frage, die mit je drei Schneidezähnen, einem Eckzahn, vier Prämolaren und drei Molaren je Kieferbogen dem ursprünglichen Typus entspricht.[8] Während dies für den Pyrenäen-Desman bisher nicht angezweifelt wird, besteht für den Russischen Desman auch eine andere Interpretation. Bei ihm läuft die Knochennaht zwischen dem Mittelkieferknochen und dem Oberkiefer entlang des zweiten oberen Zahns. Da bei den Säugetieren die Schneidezähne aber alleinig im Mittelkieferknochen stehen, wären hier nur die ersten beiden als solche anzusprechen. Alternativ hat sich dann beim Russischen Desman ein zusätzlicher fünfter Prämolar ausgebildet.[9] Der vorderste obere Schneidezahn ist jeweils sehr groß, von dreieckigem Querschnitt und spitz endend. Ihm folgen mehrere einspitzige kleine Zähne. Der letzte Prämolar ist größer und weist drei Spitzen auf. Auf den Kauoberflächen der Molaren, von denen der erste den größten repräsentiert, zeichnet sich das für Maulwürfe typische dilambdodonte Muster ab, bei dem die Haupthöcker zusammen eine W-artige Struktur bilden. In der unteren Zahnreihe formen alle Zähne eine geschlossene Reihe. Die vorderen stehen schräg nach vorn gerichtet. Der zweite Schneidezahn ragt hier am höchsten auf, dem wieder mehrere einspitzige kleine Zähne folgen. Die Molaren verfügen dann wieder über mehrere Spitzen. Vergleichbar den Eigentlichen Maulwürfen (Talpini) findet bei den Desmanen kein Wechsel vom Milch- zum Dauergebiss statt.[7][1][3]
Skelettmerkmale
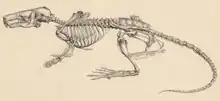
Im Skelettbau ergeben sich bei den Desmanen deutliche Abweichungen zu anderen, zumeist grabenden Maulwürfen. Die Wirbelsäule besteht aus 58 bis 59 Einzelelementen, verteilt auf 7 Hals-, 13 Brust-, 6 Lenden-, 5 bis 6 Kreuzbein- und 27 Schwanzwirbel. Vor allem die Halswirbel sind stark abgeplattet und bewirken so den kurzen Hals. Der Trend setzt sich bei den Brustwirbeln fort, erst nach hinten zu werden sie größer und höher. Die größten Wirbel finden sich im Lendenbereich. Sie unterstützen eine hohe Beweglichkeit des Körperabschnittes, was der besseren Schwimmfähigkeit zugutekommt. An den vorderen Schwanzwirbeln sind Chevronknochen ausgebildet. Der Schultergürtel ist bei den Desmanen etwas anders konfiguriert, da er unterhalb des letzten Halswirbels liegt, während er bei den grabenden Maulwürfen eine Position nahe dem Schädel einnimmt. Das Schlüsselbein zeigt sich generell lang und schlank und nicht so deutlich gestaucht und nahezu kubisch wie bei den grabenden Maulwürfen. Ebenfalls abweichend von diesen ist am schmalen Schulterblatt zudem noch ein Metacromion (ein Fortsatz am Acromion) ausgebildet. Der Oberarmknochen ist bei den Desmanen schlank und nicht so wuchtig und extrem verbreitert wie bei den grabenden Maulwürfen. Die maximale Breite erreicht bei ersteren nur rund ein Achtel der Länge, bei letzteren ist es ein Drittel bis fast die Hälfte. Er artikuliert wie bei den meisten anderen Maulwürfen jedoch an der Schulter gemeinsam mit dem Schulterblatt und dem Schlüsselbein.[10][11] Der Oberschenkelknochen wiederum ist kurz, flach und kräftig. Die unteren Extremitätenabschnitte sind bei den Desmanen stark verlängert. Ersichtlich wird dies etwa am Schienbein, das fast doppelt so lang ist wie der Oberschenkelknochen. Die Hand- und Fußknochen sind ebenfalls gestreckt und abgeplattet. Der bei den grabenden Maulwürfen an der Hand ausgebildete Präpollex („Vordaumen“) beziehungsweise das Os falciforme, ein Sesambein, welches seitlich die Handfläche verbreitert und so das Graben unterstützt, ist beim Pyrenäen-Desman extrem klein, beim Russischen Desman gar nicht vorhanden.[12][13][11][1][2][3]
Moschusdrüse
Als besondere Bildung der Weichteilanatomie kommt bei den Desmanen die Moschusdrüse vor, die am vorderen Drittel der Schwanzes ausgebildet ist. Es handelt sich um eine komplexe Struktur, die aus zwei Reihen sechseckiger Säckchen besteht. Insgesamt können bis zu 40 derartiger Säckchen bei einem Individuen vorkommen. Jedes einzelne davon ist bis zu 13 mm lang und 6 mm breit. Die Säckchen sind jeweils in Fasergewebe eingebettet, das sie voneinander trennt. Die Wandung der im Inneren hohlen Säckchen setzt sich wiederum aus kleineren Mikro-Säckchen von bis zu 3 mm Größe zusammen, von denen bis zu fünf einen gemeinsamen Kanal in das Hohlrauminnere formen. Die jeweiligen Ausgänge der größeren Säckchen führen zur Schwanzunterseite und liegen hinter den Schuppenringen. Wahrscheinlich stellt die Moschusdrüse eine funktional umgestaltete Talgdrüse dar.[7][1]
Genetische Merkmale
| Gattung | Chromosomensatz | Fundamentale Anzahl | X-Chromosom | Y-Chromosom |
|---|---|---|---|---|
| Desmana | 2n = 32 | FNa = 64 | submeta-subtelozentrisch | metazentrisch |
| Galemys | 2n = 42 | FNa = 68 | submetazentrisch | fleckenförmig |
Der diploiden Chromosomensatz beträgt beim Pyrenäen-Desman 2n = 42 und beim Russischen Desman 2n = 32. Trotz dieses großen Unterschiedes lautet die fundamentale Anzahl, also jene der einzelnen Chromosomenarme je Autosomenpaar bei ersterem 68, bei letzterem 64. Diese geringere Abweichung wird dadurch hervorgerufen, dass beim Pyrenäen-Desman im Vergleich zu zahlreichen anderen Maulwürfen ein sehr hoher Anteil an acrozentrischen Chromosomen vorkommt. Der Chromosomensatz wird bei diesem daher als urtümlich eingeschätzt. Derjenige des Russischen Desman entspricht stärker dem der meisten anderen Maulwürfe und entstand wohl durch die Kombination mehrerer acrozentrischer Chromosomen miteinander bei seinen Vorfahren.[14]
Verbreitung und Lebensraum
Die beiden Arten der Desmane haben deutlich voneinander getrennte Verbreitungsgebiete. Der Pyrenäen-Desman lebt im südwestlichen Europa in den namengebenden Pyrenäen beidseitig der französisch-spanischen Grenze und im Norden der Iberischen Halbinsel. Der Russische Desman bewohnt das östliche Europa und ist im Südwesten Russlands sowie im Nordosten der Ukraine heimisch. Mit dem äußersten Nordwesten von Kasachstan erreicht er noch zentralasiatisches Gebiet. Allgemein leben Desmane nahe bei Gewässern, wobei der Pyrenäen-Desman eher schnell fließende Bäche in stärker gebirgigen Regionen bevorzugt, während der Russische Desman breitere und langsam fließende Flüsse sowie Seen und Teiche in Überschwemmungsebenen nutzt. Für beide Vertreter sind eine reichhaltige Vegetation, Unterschlupfmöglichkeiten an den Uferhängen und ein diverses Angebot an Wirbellosen Voraussetzung.[1][2][3]
Lebensweise

Die Desmane sind stärker als die meisten Maulwürfe an eine semi-aquatische Lebensweise angepasst. Die dafür notwendigen anatomischen Voraussetzungen zeichnen sich durch den kompakten Körperbau mit langem, teilweise oder vollständig abgeplattetem Schwanz, den gegenüber den Vorderbeinen kräftigeren Hinterbeinen, den Schwimmhäuten zwischen den Zehen und den verschließbaren Nasenlöchern ab. Beim Schwimmen werden ausschließlich die Hinterbeine eingesetzt, die Vorderbeine sind nach vorn gerichtet. Schwimmbewegungen mit den Hinterbeinen erfolgen alternierend, der Schwanz dient als Steuerruder.[15][16] In der Regel halten sich die Desmane in ihrer aktiven Phase fast ausschließlich im Wasser auf. Das Ufer wird nur selten erklommen, wofür dann die kräftigen Krallen der Vorderbeine Einsatz finden. Zur Orientierung werden Geräusche sowie Geruchs- und Tastreize genutzt. Das moschusartige Drüsensekret hat hierbei vor allem bei der Geruchsorientierung eine größere Bedeutung. Beim Russischen Desman ist außerdem belegt, dass die Tiere unter Wasser Luftblasen ausstoßen, an die Geruchsstoffe anhaften. Beim erneuten Einatmen der Blasen werden diese aufgenommen.[17] Ein vergleichbares Prinzip wurde beim Sternmull (Condylura) beobachtet.[1][2][3]
Die Tiere nutzen Aktionsräume entlang der Uferlinie und bewohnen Baue, deren Eingänge stets unter Wasser liegen. Diese gewähren Zutritt zu einem unterirdischen Gangsystem, das oft selbst angelegt, teilweise aber auch von anderen grabenden Tieren übernommen wird oder aus natürlichen Höhlen besteht. Sie enthalten mit Pflanzenmaterial ausgestattete Nestkammern oberhalb der Wasserlinie. Die vorwiegenden Aktivitäten finden nachts statt mit einer Bevorzugung der Dämmerungsphasen. Die Desmane sind anders als die meisten Maulwürfe eher soziale Tiere. Verschiedene Individuen nutzen mitunter die gleichen Gangsysteme. Beim Russischen Desman formen sich in der Fortpflanzungsphase Familiengruppen, bestehend aus den Elterntieren und dem Nachwuchs. Diese lösen sich auf, nachdem der Nachwuchs selbständig geworden ist. Für den Pyrenäen-Desman ist das momentan nicht so eindeutig, doch haben sich ursprüngliche Annahmen eines aggressiv-territorialen Verhaltens in jüngeren Studien nicht bestätigt.[18][19][1][2][3]
Die Nahrung der Desmane setzt sich aus Wirbellosen und zum Teil aus Wirbeltieren zusammen. Beim Russischen Desman spielen auch Pflanzen eine größere Rolle. Unter den Wirbellosen dominieren Flohkrebse und Insekten wie Köcher- und Eintagsfliegen. Das Nahrungsrepertoire wird beim Russischen Desman durch Weichtiere ergänzt, die der Pyrenäen-Desman hingegen eher verschmäht. Bei den Wirbeltieren sind vor allem Fischen und Amphibien von größerer Bedeutung. Das pflanzliche Nahrungsspektrum ist beim Russischen Desman recht vielgestaltig und umfasst Wasserpflanzen und ufernah wachsende Pflanzen. Die tierische Beute wird von den Desmanen überwiegend im Wasser gefangen, häufig bei Tauchgängen am Grund des Gewässers. In der Regel verzehren sie nur kleinere Beute direkt im Wasser, größere schleppen sie meist an Land.[1][2][3]
Die Paarung erfolgt vermutlich überwiegend im Frühjahr. Da aber tragende und säugende Weibchen zu einem gewissen Prozentsatz ganzjährig beobachtet werden können, sind die Tiere vermutlich polyöstrisch. Die Tragzeit beträgt vermutlich 30 bis 45&nbp;Tage, die Aufzucht währt etwa genauso lang. Neugeborene sind nackt sowie blind und haben geschlossene Ohren. Die Fortpflanzung der Desmane ist allgemein nur wenig untersucht.[1][2][3] Zumindest für den Pyrenäen-Desman ist belegt, dass Weibchen Zwitterdrüsen besitzen und somit als Hermaphroditen auftreten können.[20]
Systematik
Innere Systematik der Maulwürfe nach He et al. 2016[21]
|
Die Desmane sind eine Tribus aus der Familie der Maulwürfe (Talpidae) und der Ordnung der Insektenfresser (Eulipotyphla). Sie vereint Tiere, die abweichend von anderen Maulwürfen an eine semi-aquatische Lebensweise angepasst sind. Es handelt sich um vergleichsweise große Vertreter der Familie, die aufgrund ihrer, an das Wasser gebundenen Lebensweise einzelne anatomische Besonderheiten aufweisen wie etwa der lange Schwanz oder die zwischen den Zehen bestehenden Schwimmhäute. Ursprünglich stufte man die Desmane als eine eigenständige Unterfamilie innerhalb der Maulwürfe ein. Jedoch ergaben molekulargenetische Studien, dass die Desmane eine engere Verbindung zu den Eigentlichen Maulwürfen (Talpini) und zum Sternmull (Condylurini) aufweisen. Als nächstverwandte Klade dieser gemeinsamen Verwandtschaftsgruppe stellten sich die verschiedenen Spitzmullle Nordamerikas und Asiens heraus. Die einzelnen Linien trennten sich bereits vor 37 bis 34 Millionen Jahren im Oberen Eozän voneinander ab. Die Aufspaltung der Desmane in die beiden heutigen Linien vollzog sich aber erst im Mittleren Miozän vor etwa 13,9 Millionen Jahren. Alle genannten Maulwurfsgruppen können der Unterfamilie der Altweltmaulwürfe (Talpinae) zugeordnet werden. Diesen gegenüber stehen die Spitzmausmaulwürfe (Uropsilinae) als relativ ursprünglich Gruppe, deren genetischer Ursprung bis zu 47 Millionen Jahre in das Mittlere Eozän zurückreicht.[22][21]
Innerhalb der Desmane werden zwei rezente Gattungen mit ebenso vielen Arten unterschieden. Die Gruppe gliedert sich demnach folgendermaßen:[3]
- Tribus: Desmane (Desmanini Thomas, 1912)
- Russischer Desman (Desmana Güldenstädt, 1877); Osteuropa (südwestliches Russland, nordöstliche Ukraine, nordwestliches Kasachstan)
- Pyrenäen-Desman (Galemys Kaup, 1829); Südwesteuropa (südwestliches Frankreich, nördliches und nordöstliches Spanien, nördliches Portugal)
Neben den beiden rezenten Vertretern sind noch mehrere Fossilformen bekannt:
- Archaeodesmana Topachevski & Pashkov, 1983
- Asthenoscapter Hutchison, 1974
- Gerhardstorchia Dahlmann & Doğan, 2011
- Lemoynea Bown, 1980
- Magnatalpa Oberg & Samuels, 2022
- Mygalea Schreuder, 1940
- Mygalinia Schreuder, 1940
- Mygatalpa Schreuder, 1940
Ursprünglich wurde auch Desmanella zu den Desmanen gezählt. Hier erwies sich aber aufgrund einer anderen Gebissstruktur und eines funktionalen Milchgebisses, dass die Gattung eher zu den Spitzmausmaulwürfen zu stellen ist. Des Weiteren wird die Zuweisung von Asthenoscapter und Mygatalpa zu den Desmanen von einigen Autoren ebenfalls kritisch gesehen und auch hier eine Stellung innerhalb der Spitzmausmaulwürfe bevorzugt.[23] Magnatalpa erhielt in der Erstbeschreibung der Gattung nur eine fragliche Position innerhalb der Desmane.[24] Von Miklós Kretzoi wurde im Jahr 1953 die Gattung Desmagale aus Kisláng in Ungarn eingeführt, basierend auf einem Unterkieferfragment. Der altpleistozäne Fund ist aber nach Cornelia G. Rümke identisch mit Desmana, weswegen sie erstere Form im Jahr 1985 mit letzterer gleichsetzte.[25] Rümke selbst etablierte im gleichen Jahr die Gattung Dibolia mit mehreren Arten, die im Oberen Miozän und Pliozän verbreitet waren. Der Name ist aber schon seit 1829 für eine Gattung der Blattkäfer vergeben. Barbara Rzebik-Kowalska benannte die Form daher 1994 in Ruemkelia um.[26] Allerdings synonymisierte nur ein Jahr später Rainer Hutterer die Gattung aufgrund zu weniger definierender Unterschiede mit Archaeodesmana.[27] Des Weiteren bildet Gerhardstorchia einen Ersatznamen für Storchia, etabliert im Jahr 2001 durch Thomas Dahlmann, welcher aber bereits durch eine Artengruppe der Milben präokkupiert ist.[28]
Forschungsgeschichte
Schriftliche Überlieferungen über Desmane sind wenigstens seit dem Beginn des 17. Jahrhunderts bekannt, wie unter anderen die Abhandlungen von Charles de l’Écluse über eine Mus aquaticus aus dem Jahr 1605 zeigen.[29] Gut 153 Jahre später führte Carl von Linné in seinem wegweisenden Werk zur biologischen Nomenklatur Systema Naturae den Russischen Desman offiziell wissenschaftlich ein. Er ordnete die Art zwar innerhalb der Biber ein,[30] was jedoch im Jahr 1777 durch Johann Anton Güldenstädt mit der Etablierung des Gattungsnamens Desmana korrigiert wurde.[31] Der Pyrenäen-Desman erhielt im Jahr 1811 durch Étienne Geoffroy Saint-Hilaire seine wissenschaftliche Anerkennung. Hierbei bevorzugte er die Gattungsbezeichnung Mygale,[32] welche wiederum auf Georges Cuvier zurückgeht und von diesem im Jahr 1800 im Zusammenhang mit dem Russischen Desman erwähnt worden war.[33] Der heute gültige Gattungsname Galemys für den Pyrenäen-Desman wurde wiederum von Johann Jakob Kaup im Jahr 1829 geprägt.[34] Dadurch waren bereits in der ersten Hälfte des 19. Jahrhunderts alle relevanten Namen für die Desmane vergeben worden. Während sich die Bezeichnung Desmana aus dem Schwedischen ableitet und mit dem mittellateinischen Wort bisamum für den Geruchsstoff Bisam verwandt ist, sich als auf das markante Sekret bezieht, setzen sich Mygale und Galemys jeweils aus den griechischen Wörtern μῦς (mys) für „Maus“ und γαλέη (gale) für „Wiesel“ zusammen, bedeuten also „Mauswiesel“ beziehungsweise „Wieselmaus“ und verweisen so auf das Erscheinungsbild der Tiere.[35]

Auch wenn Desmana und Galemys bereits sehr früh eingeführt worden waren, fanden sie im Verlauf des 19. und beginnenden 20. Jahrhundert kaum Verwendung. Vielmehr war es die Bezeichnung Mygale, die überwiegend als Gattung für die Desmane in Benutzung war. Als problematisch hierbei erwies sich, dass nur zwei Jahre nach Cuviers Erwähnung von Mygale eine Spinnengattung mit der gleichen Bezeichnung belegt worden war. Aufgrund dessen kamen verschiedene Alternativen wie Myogale oder Myogalea in Gebrauch. Erstere erwies sich als besonders durchsetzungsfähig und wurde vielfach angewendet, so unter anderem durch George Edward Dobson in seiner umfangreichen Abhandlung über Insektenfresser aus dem Jahr 1883.[7] Eine erste übergeordnete taxonomische Gruppe für die Desmane basierend auf Mygale als Gattungsnamen kreierte John Edward Gray im Jahr 1821, die aber aufgrund ihrer falschen Schreibweise als „Myaladae“ nicht anerkannt ist. Gray fasste die Gruppe deutlich weiter und integrierte neben dem Sternmull und dem Ostamerikanischen Maulwurf auch die Goldmulle hierin, welche mit den Maulwürfen jedoch nicht näher verwandt sind.[36] Weitere Ableitungen wurden in den nachfolgenden Jahren genutzt. So etablierte etwa Charles Lucien Jules Laurent Bonaparte im Jahr 1837 die „Myogalina“, die er den Spitzmäusen zuschlug.[37] Auguste Pomel fasste die Desmane im Jahr 1848 in den „Mygalina“ zusammen,[38] während George Jackson Mivart analog zu Bonaparte die Bezeichnung „Myogalina“ verwendete,[39][40] Dobson 1883 hingegen „Myogalae“ favorisierte.[7] Mivart galt eine Zeit lang als Autor der Tribus der Desmane.[41]
Insgesamt erwiesen sich alle Namen als nicht tragfähig. Mygale, von allen ursprünglichen Autoren auf den Russischen Desman bezogen, stellt im Sinne der Prioritätsregel der zoologischen Nomenklatur lediglich ein Synonym für Desmana dar, da letzteres zuvor benannt worden war. Für den Pyrenäen-Desman existiert der Alternativname Galemys. Antje Schreuder schlug daher im Jahr 1940 vor, für die rezenten Formen ausschließlich diese beiden Gattungen zu verwenden.[42] Bereits vor Schreuders Bekundung hatte Oldfield Thomas im Jahr 1912 unter Zugrundelegung der Gattung Desmana die übergeordnete taxonomische Gruppe der Desmaninae eingeführt. Er tätigte dies im Rahmen einer kurzen Abhandlung über die Gliederung der Maulwürfe, die er darin in fünf Unterfamilien unterteilte, darunter die Desmane.[43] Thomas gilt daher als anerkannter Taxonautor für die Tribus. Seine Einschätzung der Desmane als Unterfamilie hielt sich einige Zeit,[44][41] sie wurden dann aber spätestens in den 2000er Jahren aufgrund genetischer Befunde auf den Rang einer Tribus heruntergestuft.[45][21]
Stammesgeschichte

Frühe Vorfahren der Desmane sind bereits seit dem ausgehenden Paläogen belegt. Zu den ältesten bekannten Gattungen gehört Mygatalpa, die im Oberen Oligozän nachweisbar ist. Erstmals im Jahr 1844 von Auguste Pomel als „Desman arvernien“ anhand eines Unterkiefers aus Chauffours vorgestellt, benannte er die Form vier Jahre später in „Mygale arverniensis“ um und fügte noch einen Oberarmknochen bei.[46][38] Fast einhundert Jahre später etablierte Antje Schreuder die Bezeichnung Mygatalpa für die Form, sah aber keinen unmittelbaren Zusammenhang mit den Desmanen.[42] Die Diskussion um die tatsächliche Zugehörigkeit von Mygatalpa zu den Desmanen hält bis heute an, da einige Autoren sie näher bei den Spitzmausmaulwürfen einordnen. Sie basiert aber zum Teil auf falsch zugewiesenem Fundmaterial.[47] Insgesamt sind Fossilreste dieses kleinen Maulwurfs eher selten. Für das Untere Miozän wird noch ein äußerst schlanker Oberarmknochen aus Tomerdingen bei Ulm genannt.[48][49] Eindeutiger verhält es sich mit Mygalea. Diese im Jahr 1940 von Antje Schreuder eingeführte Form basiert auf einzelnen Unterkiefern und Gliedmaßenresten aus Sansan im Süden Frankreichs, die bereits in den 1840er und 1850er Jahren von Auguste Pomel und Édouard Armand Lartet (als Mygale) veröffentlicht worden waren.[38][50][42] Fossilfunde liegen darüber hinaus von einigen weiteren Lokalitäten in Europa vor. An der Fundstelle Merkur-Nord bei Tuchořice im nördlichen Tschechien bildet Mygalea mit mehreren hundert Einzelfunden den häufigsten Maulwurfsvertreter. Das Ensemble bestehend aus Kieferresten und Oberarmknochen datiert in den Beginn des Unteren Miozäns.[51] Nahezu gleichalt ist das ebenfalls vergleichsweise umfangreiche Material von verschiedenen Fundpunkten aus der Umgebung von Ulm, das mehrere Unterkiefer einschließt.[48] Bereits in das beginnende Mittlere Miozän vor rund 15 Millionen Jahren gehört der bemerkenswerte Fund eines disartikulierten Skeletts aus Viehhausen bei Regensburg. Es besteht aus einem fragmentierten Schädel, den vorderen Extremitäten und einigen Wirbeln, wobei vor allem die Beinknochen äußerst schlank sind.[11] Zu erwähnen sind noch die Funde aus den Spaltenfüllungen von Petersbuch in der Fränkischen Alb, von wo mehrere Unterkiefer stammen. Ihr Alter entspricht dem ausgehenden Mittleren Miozän.[52][49]
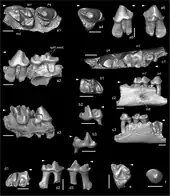
Noch im Übergang vom Unteren zum Mittleren Miozän erreichten Desmane auch Areale sehr weit östlich in Eurasien, wovon Zahn- und Kieferreste von der Fundstelle Tagay am Westufer der Insel Olchon im Baikalsee zeugen.[53] Im westlichen Eurasien hingegen trat im Mittleren Miozän parallel zu Mygalea die Gattung Asthenoscapter in Erscheinung. Beschrieben wurde die Form im Jahr 1974 von John Howard Hutchison anhand von rund 250 Fossilresten aus einer Spaltenfüllung bei La Grive-Saint-Alban im östlichen Frankreich. Das Material entfällt zur Hälfte auf Zahn- beziehungsweise Gebissfragmente und auf Oberarmknochen. Weitere Nachweise der Gattungen liegen aus Sansan und aus der Umgebung von Ulm vor. Die überlieferten Reste lassen einen kleinen Maulwurfsertreter rekonstruieren, der mit schlanken Vorderbeinen ausgestattet war und in dessen unteren Gebiss die Schneidezähne nahezu die gleiche Größe aufwiesen. Ähnlich wie bei Mygatalpa wird aber die genaue systematische Stellung von Asthenoscapter diskutiert.[47][48] Eine weitere, im Mittleren Miozän belegte Maulwurfsform ist Gerhardstorchia, benannt zu Ehren von Gerhard Storch.[28] Sie wurde im Jahr 2001 über Funde aus Wölfersheim in Hessen definiert und zeichnet sich durch eine schlanke Bezahnung aus. Zusätzliches Material aus dieser Zeit stammt aus Devínska Nová Ves in der Slowakei, von wo ein Unterkiefer dokumentiert wurde. Mit Dorn-Dürkheim in Rheinland-Pfalz sowie mit verschiedenen Fundpunkten in Niederösterreich, etwa Eichkogel oder Götzendorf an der Leitha, können auch einige obermiozäne Lokalitäten genannt werden, wobei letztere mit 11 Millionen Jahren noch einmal knapp 2 Millionen Jahre älter ist als erstere. Mit Studienka kann auch ein recht umfangreicher slowakischer Fundkomplex benannt werden. In der Regel handelt es sich um isolierte Zähne oder einzelne Kieferfragmente.[54][49][55][56]
Im Oberen Miozän und im Übergang zum Pliozän kommt noch Mygalinia hinzu, ein sehr kleiner Vertreter der Desmane, der sogar die Ausmaße des Pyrenäen-Desmans unterbot. Er wurde erstmals im Jahr 1913 von Theodor Kormos an der ungarischen Fundstelle Polgárdi nordöstlich des Balaton dokumentiert, damals von ihm aber noch der Gattung Galemys zugeordnet.[57] Erst gut ein Viertel Jahrhundert später überführte Antje Schreuder die Funde zur neuen Gattung Mygalinia.[42] Das Ausgangsmaterial besteht aus einzelnen Unterkiefern und einigen Gliedmaßenknochen. Später wurde die Gattung noch von mehreren spätmiozänen Lokalitäten wie Soblay, Mollon oder Lobrieu in Frankreich berichtet,[58] ebenso wie von Werchnjaja Kriniza aus der Ukraine. Insgesamt ist das Fundmaterial aber zu spärlich – aus Werchnjaja Kriniza liegt beispielsweise nur ein einziger Zahn vor –, um genauere Aussagen treffen zu können.[59][60] Den wohl am weitesten verbreiteten und häufigsten Vertreter des Oberen Miozäns und des folgenden Pliozäns stellt aber Archaeodesmana dar, von dem über ein halbes Dutzend Arten beschrieben wurden. Fundnachweise reichen vom südwestlichen über das zentrale bis in das südöstliche und östliche Europa sowie in das westliche Asien. Vergleichsweise alte Fossilreste aus dem Oberen Miozän in Form von Einzelzähnen sind unter anderem aus den bereits erwähnten Lokalitäten von Dorn-Dürkheim, Eichkogel und Götzendorf an der Leitha geborgen worden.[49][55] Als nahezu gleichalt erwiesen sich ein recht umfangreicher Fundkomplex mit mehr als dreißig Zähnen und einzelnen Beinknochen aus Šalgovce in der Slowakei,[56] darüber hinaus auch ein Oberkieferfragment aus Polgárdi in Ungarn und einige wenige Zähne aus Werchnjaja Kriniza und Michailovka in der Ukraine.[59][60] Mehrere ähnlich alte Fundkomplexe stammen von der Iberischen Halbinsel, so beispielsweise Masada del Valle und Aljezar in der ostspanischen Provinz Teruel. Im darauffolgenden Pliozän erlangt Archaeodesmana vor allem auf der Iberischen Halbinsel eine recht weite Verbreitung. Genannt werden können etwa Purcal bei Granada und Tollo de Chiclana bei Guadix.[61][62][63] Während es sich auch hier weithgehend um Zähne und Kieferfragmente handelt, stammt ein nahezu vollständiger Schädel aus Sète im südlichen Frankreich. Im zentralen Europa reiht sich hier Gundersheim in Rheinhessen ein.[64] Weiteres Material wurde aus Griechenland berichtet, neben Ptolemais außerdem aus Maramena, beide im Norden des Landes gelegen. Letztere Fundstelle barg Zähne und Skelettteile.[65] Im Pliozän ist Archaeodesmana dann auch aus der heutigen Türkei belegt, stellvertretend sei hier Dinar-Akçaköy im westlichen Landesteil genannt. Allgemein handelt es sich bei Archaeodesmana um einen kleinen Maulwurf, dessen oberer vorderer Schneidezahn zwei Spitzen aufwies. Die Form steht an den jeweiligen Fundplätzen mit limnisch-fluviatilen Umweltverhältnissen in Verbindung und gilt als Ausgangspunkt der Entwicklung von sowohl Desmana als auch Gelemys. Die beiden heutigen Gattungen sind im Verlauf des Pliozäns erstmals fossil fassbar und nachfolgend recht weit über Europa verbreitet.[25][63]

Außerhalb des eurasischen Raumes wurden Desmane zusätzlich aus Nordamerika beschrieben. Der älteste Vertreter ist als Lemoynea benannt worden, wobei die Lemoyne Quarry bei der gleichnamigen Ortschaft im Keith County des US-Bundesstaates Nebraska Pate für die Bezeichnung stand. Die Funde, die der Ash-Hollow-Formation angehören und aus einzelnen Unterkieferfragmenten bestehen, zeichnen ein Tier, das etwas größer als Mygalinia wurde und sich durch einen besonders großen dritten unteren Schneidezahn auszeichnete. Wie für die Ash-Hollow-Formation allgemein, wird den Überresten ein spätmiozänes Alter zugesprochen.[66] Etwa jünger mit einer Stellung im Unteren Pliozän ist dann Magnatalpa. Die Form wurde an der Gray fossil site im US-Bundesstaat Tennessee nachgewiesen, wo die Tiere in einer wasserreichen Karstlandschaft mit vielfältiger Fauna lebten. Entsprechend dem Namen handelt es sich um eine große Form, die alle heutigen Maulwürfe Nordamerikas an Größe übertraf und die Dimensionen des Russischen Desmans erreichte. Es liegen allerdings lediglich einzelne isolierte Zähne und ein zahnloser Unterkiefer vor.[24]
Bedrohung und Schutz
Die beiden Arten der Desmane werden von der IUCN in ihrem jeweiligen Bestand als „stark gefährdet“ (endangered) eingestuft. Zu den Hauptbedrohungen zählen die Gewässerverschmutzung, die Zerstörung des Lebensraumes einschließlich der Störung der Gewässernetze sowie die illegale Jagd. Dies betrifft insbesondere den Russischen Desman, der in der Vergangenheit auch wegen des Desmanfells verfolgt wurde. Außerdem besteht eine starke Konkurrenz durch Neozoen wie dem Amerikanischen Nerz, der Nutria und der Bisamratte. Sowohl der Pyrenäen-Desman als auch der Russische Desman sind heute geschützt. Mit ersterem wird zudem als Aushängeschild für die Förderung der Erhaltung von Flusslandschaften geworben. Die Tiere sind in ihrem jeweiligen Verbreitungsgebiet in Schutzzonen präsent.[67][68]
Literatur
- I. I. Barabasch-Nikiforow: Die Desmane. Neue Brehm Bücherei 474, Wittenberg, 1975, S. 1–100
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 605–607) ISBN 978-84-16728-08-4
- Ronald M. Nowak: Walker’s Mammals of the World. 2 Bände. 6. Auflage. The Johns Hopkins University Press, Baltimore MD u. a. 1999, ISBN 0-8018-5789-9
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. A taxonomic and geographic Reference. 2 Bände. 3. Auflage. The Johns Hopkins University Press, Baltimore MD 2005, ISBN 0-8018-8221-4
Einzelnachweise
- I. I. Barabasch-Nikiforow: Die Desmane. Neue Brehm Bücherei 474, Wittenberg, 1975, S. 1–100
- E.-A. Juckwer: Galemys pyrenaicus (Geoffroy, 1811) – Pyrenäen-Desman. In: J. Niethammer und F. Krapp (Hrsg.): Handbuch der Säugetiere Europas. Wiesbaden, 1990, S. 79–92
- Boris Kryštufek und Masaharu Motokawa: Talpidae (Moles, Desmans, Star-nosed Moles and Shrew Moles). In: Don E. Wilson und Russell A. Mittermeier (Hrsg.): Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths, Colugos. Lynx Edicions, Barcelona 2018, S. 552–620 (S. 605–607) ISBN 978-84-16728-08-4
- S. U. Stroganov: Morphological characters of the auditory ossicles of recent Talpidae. Journal of Mammalogy 26 (4), 1945, S. 12–42
- Matthew J. Mason: Evolution of the Middle Ear Apparatus in Talpid Moles. Journal of Morphology 267, 2006, S. 678–695
- Daisuke Koyabu, Misato Hosojima und Hideki Endo: Into the dark: patterns of middle ear adaptations in subterranean eulipotyphlan mammals. Royal Society Open Science 4, 2017, S. 170608, doi:10.1098/rsos.170608
- George Edward Dobson: A Monograph of the Insectivora, systematic and anatomical. Part II. London, 1883, S. 126–172 ()
- Alan C. Ziegler: Dental homologies and possible relationships of recent Talpidae. Journal of Mammalogy 52 (1), 1971, S. 50–68
- Shin-ichiro Kawada, Kazuhiro Koyasu, Elena I. Zholnerovskaya und Sen-ichi Oda: A discussion of the dental formula of Desmana moschata in relation to the premaxillary suture. Mammal Study 27, 2002, S. 107–111
- B. Campbell: The shoulder anatomy of the moles. A study in phylogeny and adaptation. American Journal of Anatomy 64, 1939, S. 1–39
- Achim H. Schwermann und Richard S. Thompson: Extraordinarily preserved talpids (Mammalia, Lipotyphla) and the evolution of fossoriality. Journal of Vertebrate Paleontology 35 (4), 2015, S. e934828, doi:10.1080/02724634.2014.934828
- Marcelo R. Sánchez-Villagra und Peter R. Menke: The mole’s thumb – evolution of the hand skeleton in talpids (Mammalia). Zoology 108, 2005, S. 3–12
- Marcelo R. Sánchez-Villagra, Inés Horovitz und Masaharu Motokawa: A comprehensive morphological analysis of talpid moles (Mammalia) phylogenetic relationships. Cladistics 22, 2006, S. 59–88
- В. М. Малыгин und Д. Л. Заславский: Особенности кариотипов Русской (Desmana moschata) и Пиренейской (Galemys pyrenaicus) выхухолей в свази с вопросом о положении етих видов в семействе Talpidae (Mammalia, Insectivora). Зоологический Журнал 65, 1986, S. 1435–1437
- Günther Niethammer: Beobachtungen am Pyrenäen-Desman, Galemys pyrenaica. Bonner Zoologische Beiträge 21 (3/4), 1970, S. 157–182
- Yu. F. Ivlev, A. N. Kuznetsov und M. V. Rutovskaya: Preliminary Data on the Swimming Kinematics of the Russian Desman (Desmana moschata L.). Doklady Biological Sciences 431, 2010, S. 144–148
- Yu. F. Ivlev, M. V. Rutovskaya und O. S. Luchkina: The Use of Olfaction by the Russian Desman (Desmana moschata L.) during Underwater Swimming. Doklady Biological Sciences 452, 2013, S. 280–283
- Y. Melero, P. Aymerich, G. Santulli und J. Gosàlbez: Activity and space patterns of Pyrenean desman (Galemys pyrenaicus) suggest non-aggressive and non-territorial behaviour. European Journal of Wildlife Research 60, 2014, S. 707–715, doi:10.1007/s10344-014-0838-8
- Yolanda Melero, Pere Aymerich, Juan Jose Luque-Larena und Joaquim Gosàlbez: New insights into social and space use behaviour of the endangered Pyrenean desman (Galemys pyrenaicus). European Journal of Wildlife Research 58, 2012, S. 185–193, doi:10.1007/s10344-011-0561-7
- F. David Carmona, Masaharu Motokawa, Masayoshi Tokita, Kimiyuki Tsuchiya, Rafael Jiménez und Marcelo Sánchez-Villagra: The Evolution of Female Mole Ovotestes Evidences High Plasticity of Mammalian Gonad Development. Journal of Experimental Zoology (Mol Dev Evol) 310 B, 2008, S. 259–266
- Kai He, Akio Shinohara, Kristofer M. Helgen, Mark S. Springer, Xue-Long Jiang und Kevin L. Campbell: Talpid Mole Phylogeny Unites Shrew Moles and Illuminates Overlooked Cryptic Species Diversity. Molecular Biology and Evolution 34 (1), 2016, S. 78–87
- María Teresa Cabria, Jonathan Rubines, Benjamín Gómez-Moliner und Rafael Zardoya: On the phylogenetic position of a rare Iberian endemic mammal, the Pyrenean desman (Galemys pyrenaicus). Gene 375, 2006, S. 1–13, doi:10.1016/j.gene.2006.01.038
- Lars W. van den Hoek Ostende: Insectivore faunas from the Lower Miocene of Anatolia – Part 5: Talpidae. Scripta Geologica 122, 2001, S. 1–35
- Danielle E. Oberg und Joshua X. Samuels: Fossil moles from the Gray fossil site (Tennessee): Implications for diversification and evolution of North American Talpidae. Palaeontologia Electronica 25 (3), 2022, S. a33, doi:10.26879/1150
- Cornelia G. Rümke: A review of fossil and recent Desmaninae (Talpidae, Insectivora). Utrecht Micropaleontological Bulletins, Special Publication 4, 1985, S. 1–241
- Barbara Rzebik-Kowalska und Jerzy Pawłowski: Ruemkelia (Mammalia, Insectivora, Talpidae) nom. nov. for Dibolia Rümke, 1985 (nec Latreille, 1829). Acta Zoologica Cracoviensia 37 (1), 1994, S. 75–76
- Rainer Hutterer: Archaeodesmana Topachevsky & Pashkov, the correct name for Dibolia Rümke, a genus of fossil water moles (Mammalia: Talpidae). Bonner Zoologische Beiträge 45 (3/4), 1995, S. 171–172
- Thomas Dahlmann und Salih Doğan: Gerhardstorchia nomen novum: a new name for Storchia Dahlmann 2001 (Mammalia: Lipotyphla: Talpidae: Desmaninae). Paläontologische Zeitschrift 85, 2011, S. 91
- Charles de l’Écluse: Exoticorum libri decem: quibus animalium, plantarum, aromatum, aliorumque peregrin. fructuum historiae describuntur / item Pt. Bellonii Observationes, eodem C.C. interprete. Leiden 1605, S. 1–378 (S. 375) ()
- Carl von Linné: Systema naturae. 10. Auflage, 1758, Band 1, S. 59 ()
- Johann Anton Güldenstädt: Beschreibung des Desmans. Beschäftigungen der Berlinischen Gesellschaft Naturforschender Freunde 3, 1777, S. 107–137 ()
- Etienne Geoffroy Saint-Hilaire: Memoire sur les especes des genres Musaraigne et Mygale. Annales du Muséum d’histoire naturelle 17, 1811, S. 169–194 ()
- Georges Cuvier: Leçons d’anatomie comparée. Tome 1. Paris, 1800, S. 1–521 ()
- Johann Jakob Kaup: Skizzirte Entwickelungs-Geschichte und Natürliches System der Europäischen Thierwelt. Darmstadt und Leipzig, 1829, S. 1–203 (S. 119) ()
- T. S. Palmer: Index Generum Mammalium: A List of the Genera and Families of Mammals. North American Fauna 23, 1904, S. 288 und 437 ()
- John Edward Gray: On the natural arrangement of vertebrose animals. London Medical Repository 15, 1821, S. 297–310 (300) ()
- Charles Lucien Jules Laurent Bonaparte: New systematic arrangement of vertebrated animals. Transactions of the Linnean Society 18, 1841, S. 247–304 ()
- Auguste Pomel: Étude sur les carnassiers insectivores. Bibliothèque universelle de Genève 9, 1848, S. 159–167 () und S. 244–251 ()
- St. George Mivart: On Hemicentetes, a new Genus of Insectivora, with some additional remarks on the osteology of that order. Proceedings of the Zoological Society of London, 1871, S. 58–79 ()
- Theodore Gill: Synopsis of insectivorous mammals. Bulletin of the United States Geological and Geographical Survey of the Territories 1, 1875, S. 91–120 ()
- Leigh van Valen: New Paleocene Insectivores and Insectivore classification. Bulletin of the American Museum of Natural History 135, 1967, S. 217–284
- Antje Schreuder: A revision of the fossil water-moles (Desmaninae). Archives Néerlandais de Zoologie 4, 1940, S. 201–233
- Oldfield Thomas: On the collection of mammals from the Tsin-ling mountains, Central China, presented by Mr. G. Fenwick Owen to the National Museum. Annals and Magazine of Natural History 8 (10), 1912, S. 395–403 ()
- George Gaylord Simpson: The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History 85, 1945, S. 1–350 (S. 52–53)
- Don E. Wilson und DeeAnn M. Reeder (Hrsg.): Mammal Species of the World. A taxonomic and geographic Reference. 2 Bände. 3. Auflage. Johns Hopkins University Press, Baltimore MD 2005, ISBN 0-8018-8221-4 ()
- Auguste Pomel: Description géologique et paléontologique des collines de la Tour de Boulade et du Puy de Teiller (Puy de Dôme). Bulletin de la Société géologique de France 2 (1), 1844, S. 579–596 ()
- J. Howard Hutchison: Notes on type specimens of European Miocene Talpidae and a tentative Classification of Old World Tertiary Talpidae (Insectivora: Mammalia). Geobios 7 (3), 1974, S. 211–256
- Reinhard Ziegler: Talpidae (Insectivora, Mammalia) aus dem Oberoligozän und Untermiozän Süddeutschlands. Stuttgarter Beiträge zur Paläontologie Serie B 167, 1990, S. 1–80
- Reinhard Ziegler: Miocene Insectivores from Austria and Germany – An Overview. Beiträge zur Paläontologie 30, 2006, S. 481–494
- Édouard Armand Lartet: Notice sur la colline de Sansan. Paris, 1851, S. 1–46 ()
- Lars W. van den Hoek Ostende und Oldrich Fejfar: Erinaceidae and Talpidae (Erinaceomorpha, Soricomor-pha, Mammalia) from the Lower Miocene of Merkur-Nord (Czech Republic, MN 3). Beiträge zur Paläontologie 30, 2006, S. 175–203
- Reinhard Ziegler: Moles (Talpidae) from the late Middle Miocene of South Germany. Acta Palaeontologica Polonica 48 (4), 2003, S. 617–648
- Leonid L. Voyta, Thomas Mörs und Margarita A. Erbajeva: Erinaceomorpha and Soricomorpha (Mammalia) of the Miocene Tagay fauna (Olkhon Island, Lake Baikal, Eastern Siberia): A preliminary report. Palaeobiodiversity and Palaeoenvironments 102, 2022, S. 897–914, doi:10.1007/s12549-022-00557-9
- Martin Sabol: Middle Miocene assemblage of insectivores from Bonanza site near Devínska Nová Ves (Slovakia). Geologica Carpathica 56 (5), 2005, S. 433—445
- Reinhard Ziegler: Insectivores (Lipotyphla) and bats (Chiroptera) from the Late Miocene of Austria. Annalen des Naturhistorischen Museums zu Wien 107 A, 2006, S. 93–196
- Florentin Cailleux, Lars W. van den Hoek Ostende und Peter Joniak: The Late Miocene Talpidae (Eulipotyphla, Mammalia) from the Pannonian Region, Slovakia. Journal of Paleontology 98 (1), 2024, S. 128–151, doi:10.1017/jpa.2023.95
- Theodor Kormos: Trois nouvelles espèces fossiles des Desmans en Hongrie. Annales Musei Nationalis Hungarici 11, 1913, S. 135–145
- Claude Guérin und Pierre Mein: Les principaux gisements de mammifères miocènes et pliocènes du domaine rhodanien. Documents du Laboratoire de Géologie de la Faculté des Sciences de Lyon 1, 1971, S. 131–170
- Barbara Rzebik-Kowalska und Leonid I. Rekovets: Recapitulation of data on Ukrainian fossil insectivore mammals (Eulipotyphla, Insectivora, Mammalia). Acta zoologica cracoviensia 58 (2), 2015, S. 137–171, doi:10.3409/azc.58_2.137
- Barbara Rzebik-Kowalska und Leonid I. Rekovets: New data on Eulipotyphla (Insectivora, Mammalia) from the Late Miocene to the Middle Pleistocene of Ukraine. Palaeontologia Electronica 19 (1), 2016, S. 9A ()
- Elvira Martín Suárez, Nicolás Bendala und Matthijs Freudenthal: Archaeodesmana baetica, sp. nov . (Mammalia, Insectivora, T alpidae) from the Mio–Pliocene transition of the Granada Basin, southern Spain. Journal of Vertebrate Paleontology 21 (3), 2001, S. 547–554
- Raef Minwer-Barakat, Antonio García-Alix und Matthijs Freudenthal: Desmaninae (Talpidae, Mammalia) from the Pliocene of Tollo de Chiclana (Guadix Basin, Southern Spain) Considerations on the phylogeny of the genus Archaeodesmana.Geobios 41, 2008, S. 381–398, doi:10.1016/j.geobios.2007.08.001
- Raef Minwer-Barakat, Antonio García-Alix, Elvira Martín-Suárez und Matthijs Freudenthal: Early Pliocene Desmaninae (Mammalia, Talpidae) from Southern Spain and the Origin of the Genus Desmana. Journal of Vertebrate Paleontology 40 (5), 2020, S. e1835936, doi:10.1080/02724634.2020.1835936
- Thomas Dahlmann und Gerhard Storch: Eine pliozäne (ober-ruscinische) Kleinsäugerfauna aus Gundersheim, Rheinhessen. 2. Insektenfresser: Mammalia, Lipotyphla Senckenbergiana Lethaea 76 (1/2), 1996, S. 181–191
- Constantin S. Doukas, Lars W. van den Hoek Ostende, Constantin Theocharopoulos und Jelle W. F. Reumer: Insectivora (Erinaceidae, Talpidae, Soricidae, Mammalia). In: N. Schmidt-Kittler (Hrsg.): The vertebrate locality Maramena (Macedonia, Greece) at the Turolian–Ruscinian boundary. Münchner Geowissenschaftliche Abhandlungen A28, 1995, S. 43–64
- Thomas M. Bown: The Fossil Insectivora of Lemoyne Quarry (Ash Hollow Formation, Hemphillian), Keith County, Nebraska. Transactions of the Nebraska Academy of Sciences and Affiliated Societies 284, 1980, S. 99–122
- R. Kennerley und S. T. Turvey: Desmana moschata. The IUCN Red List of Threatened Species 2016. e.T6506A22321477 (); zuletzt aufgerufen am 17. Dezember 2022
- L. Quaglietta: Galemys pyrenaicus (amended version of 2021 assessment). The IUCN Red List of Threatened Species 2022. e.T8826A214429993 (); zuletzt aufgerufen am 17. Dezember 2022
{kind=link}
Weblinks
- Desmana moschata in der Roten Liste gefährdeter Arten der IUCN 2016. Eingestellt von: R. Kennerley & S. T. Turvey, 2016. Abgerufen am 17. Dezember 2022.
- Galemys pyrenaicus in der Roten Liste gefährdeter Arten der IUCN 2021. Eingestellt von: L. Quaglietta, 2022. Abgerufen am 17. Dezember 2022.
- BBC: Pyrenean desman: On the trail of Europe’s oddest beast, Filmaufnahmen eines Pyrenäen-Desmans