Kollagene
Kollagene (Vorstufe Tropokollagene; internationalisierte Schreibweise Collagene; Betonung auf der zweitletzten Silbe) sind eine Gruppe nur bei vielzelligen Tieren (einschließlich Menschen) vorkommender Strukturproteine (ein Faserbündel bildendes „Eiweiß“) hauptsächlich des Bindegewebes (genauer: der extrazellulären Matrix). Kollagene finden sich unter anderem in den weißen, unelastischen Fasern von Sehnen, Bändern, Knochen und Knorpeln. Auch Schichten der Haut (Unterhaut) bestehen aus Kollagenen.
| Kollagen | ||
|---|---|---|
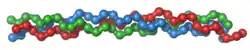
| ||
| Kollagen-Tripelhelix | ||
| Masse/Länge Primärstruktur | 3 × 200–1000 Aminosäuren | |
| Sekundär- bis Quartärstruktur | Tripelhelix | |
| Präkursor | Prokollagen-Polypeptidketten und Prokollagen | |
| Isoformen | 28 beim Menschen | |
| Arzneistoffangaben | ||
| ATC-Code | B02BC07, G04BX11, D11AX57 | |
| Wirkstoffklasse | Faserprotein | |
| Vorkommen | ||
| Übergeordnetes Taxon | Gewebetiere | |
Hintergrund



Im menschlichen Körper ist Kollagen mit über 30 % Anteil an der Gesamtmasse aller Proteine das am häufigsten vorkommende Eiweiß. Es ist ein wesentlicher organischer Bestandteil des Bindegewebes (Knochen, Zähne, Knorpel, Sehnen, Bänder) und der Haut. Seinen Namen erhielt das Kollagen (aus dem Griechischen: Leim erzeugend) ursprünglich aufgrund seiner früheren Nutzung als Knochenleim im Holzhandwerk. Es ist der Hauptgrundstoff für die Herstellung von Gelatine.
Kollagen besteht aus einzelnen langen Kollagenmolekülen (Proteinketten bzw. korrekterweise Aminosäureketten), die eine linksgängige Helix (ähnlich der Polyproline-II-Helix) ausbilden. Jeweils drei dieser Helices sind in einer rechtsgängigen Superhelix arrangiert, dem eigentlichen Kollagen. Diese Tripelhelix wird durch Wasserstoffbrücken zwischen den einzelnen Strängen stabilisiert.
Auffallend an der Primärstruktur (Aminosäuresequenz) des Kollagens ist, dass jede dritte Aminosäure Glycin ist. Ein in der Proteinfamilie der Kollagene häufig wiederholtes Sequenzmotiv ist Glycin-Prolin-Hydroxyprolin.
Kollagenfasern besitzen eine enorme Zugfestigkeit und sind kaum dehnbar. Die dichte Wicklung ist hierbei ausschlaggebend für die hohe Zugfestigkeit von Kollagenfasern.
Kollagene spielen in der Biomineralisation der Wirbeltiere eine entscheidende Rolle.[1]
Im allgemeinen Sprachgebrauch wird Kollagen Typ I gleichgesetzt mit „Kollagen“. Kollagen Typ I ist zwar mengenmäßig im Säugetier das bedeutendste Kollagen und durch seine Verwendung als Gelatine auch das Bekannteste, es existieren jedoch weitere Kollagentypen, die sich strukturell wesentlich vom Kollagen Typ I unterscheiden und andere wichtige biologische Funktionen wahrnehmen. Gelatine ist die denaturierte Form von fibrillärem Kollagen Typ I, II und/oder III und wird meist aus Schlachtabfällen gewonnen. Hierbei ist zu beachten, dass Kollagen Typ II vornehmlich im Knorpel vorkommt, Gemische von Kollagen Typ I und III stammen aus Sehnen, Bändern und der Haut.
Vorkommen
Funktionale Gene kollagener Strukturproteine finden sich bei allen Stämmen der vielzelligen Tiere, bei Schwämmen,[2] Nesseltieren[3][4] bis zu Säugetieren, hauptsächlich in deren extrazellulären Matrix und im Bindegewebe. Kollagene treten bei anderen Organismen wie Pilzen, Pflanzen oder Einzellern nicht auf.
Aufbau
Kollagenmolekül
Die Polypeptidketten des Kollagens werden einzeln durch die Ribosomen des rauen Endoplasmatischen Retikulums synthetisiert. Als Kollagenmolekül oder Tropokollagen werden nur tripelhelikale Moleküle der extrazellulären Matrix (EZM) bezeichnet. Sie haben gemeinsam, dass sie aus drei Polypeptidketten aufgebaut sind. Diese liegen jeweils in linksgängigen Kollagen-Helices (α-Ketten, nicht zu verwechseln mit den rechtsgängigen α-Helices) vor und sind gemeinsam in Form der charakteristischen rechtshändigen Tripelhelix umeinander gewunden. Jede einzelne Kollagen-Helix kann in Abhängigkeit vom Kollagentyp aus 600 bis 3000 Aminosäuren zusammengesetzt sein und ist mit großen Domänen ausgestattet, die aus sich wiederholenden (repetitiven) G-X-Y-Sequenzen aufgebaut sind.
Somit befindet sich an jeder dritten Position ein Glycin (G)-Rest. Glycin als die kleinste Aminosäure passt ideal in die Tripelhelix mit ihren sehr engen Windungen. Die Aminosäure Prolin ist sehr häufig an Position X zu finden. Prolin fungiert hier aufgrund seiner starren Ringstruktur als „Ecke“ in der Polypeptidkette und unterstützt die Ausbildung von engen Windungen innerhalb der Tripelhelix. 4-Hydroxyprolin ist überwiegend an Position Y lokalisiert und stabilisiert die Tripelhelix über Wasserstoffbrücken zwischen benachbarten Polypeptidketten. Durch die Verwendung von Glycin, Prolin und Hydroxyprolin wird die Rotation der Polypeptidkette begrenzt und den engen Raumbedingungen innerhalb der Tripelhelix Rechnung getragen.
| Strukturebene | molekularer Bereich |
|---|---|
| Primärstruktur (= Sequenz) | Polypeptidketten mit repetitiven G-X-Y-Sequenzen |
| Sekundärstruktur | linksgängige Kollagen-Helices (α-Ketten) |
| Tertiärstruktur | rechtsgängige Tripelhelix aus 3 Polypeptidketten (Tropokollagen Ø1,5 nm) |
| Quartärstruktur | Mikrofibrillen Ø20–40 nm, Fibrillen Ø300–500 nm, Fasern Ø4–12 μm |
Das Vorkommen von Hydroxylysin neben Hydroxyprolin ist ebenfalls charakteristisch für Kollagen. Hydroxylysin bildet die Voraussetzung für die Ausbildung kovalenter Quervernetzungen, womit die einzelnen Tripelhelices innerhalb der Kollagenfibrillen räumlich fixiert werden können.
Kollagenfibrille

In den Fibrillen sind benachbarte Kollagenmoleküle nicht bündig angeordnet, sondern um 67 nm, d. h. um etwa ein Fünftel ihrer Länge, gegeneinander versetzt. Diese Anordnung hat zur Folge, dass auf elektronenmikroskopischen Aufnahmen von Metall-kontrastierten Kollagenfibrillen eine Querstreifung zu sehen ist. Es entsteht ein charakteristisches Bänderungsmuster, das sich alle 67 nm (234 Aminosäuren) wiederholt und als D-Periode bezeichnet wird. Dadurch werden die α-Ketten in vier homologe Bereiche D1–D4 unterteilt. Die in einer D-Einheit auftretenden Banden werden mit a–e bezeichnet. Die Kollagen-Fibrillen sind geordnete Polymere, die im ausgereiften Gewebe viele Mikrometer lang werden können. Sie sind oft zu größeren, kabelartigen Bündeln, den Kollagenfasern, zusammengefasst. Bei Sehnen betragen die Kollagen Typ I Fibrillendurchmesser 50–500 nm, in der Haut 40–100 nm und in der Kornea (Hornhaut des Auges) 25 nm. Die Fibrillogenese des Kollagens wird oftmals durch kleine leucinreiche Proteoglykane reguliert, so dass in den entsprechenden Geweben Fibrillen mit definiertem Durchmesser und definierter Anordnung entstehen können.
Strukturaufklärung
Das heutige Bild der Kollagen-Tripelhelix und die räumliche Einordnung der Aminosäurereste und ihrer Wasserstoffbrücken untereinander geht maßgeblich auf die Röntgen-kristallographischen Arbeiten der indischen Wissenschaftler G. N. Ramachandran und Gopinath Kartha zurück (1954).
Wesentliche Aufklärung (die gesamte Primärstruktur des Typ I Kollagens sowie die Makrostrukturen der Typen IV und VI) leistete das ehemalige Max-Planck-Institut für Eiweiß- und Lederforschung in München (1956 zur Aufklärung des Bindegewebes durch Sponsoring der Lederindustrie gegründet) ab 1966 unter der Leitung von Klaus Kühn (nach Institutsverlegung am Max-Planck-Institut für Biochemie in Martinsried).[5]
Biosynthese
Kollagen wird hauptsächlich in Fibroblasten, Chondroblasten, Osteoblasten und Odontoblasten hergestellt, jedoch wird Kollagen auch in vielen anderen Zelltypen synthetisiert. Am besten untersucht ist die Biosynthese des Kollagens in Fibroblasten. Fibroblasten besitzen ein ausgedehntes raues endoplasmatisches Retikulum und einen gut entwickelten Golgi-Apparat, um an der Sekretion der Kollagene in die extrazelluläre Matrix teilnehmen zu können. Fibroblasten stellen Kollagen de novo her und sezernieren es in die extrazelluläre Matrix. Außerdem sind Fibroblasten in der Lage, Kollagen mithilfe von Enzymen, den sogenannten Kollagenasen, abzubauen.[6]
1. Transkription: Bislang sind 42 Gene bekannt, die sich an der Biosynthese von Kollagen beteiligen und für 28 verschiedene Kollagentypen codieren.[7] Jedes Gen codiert für eine bestimmte mRNA-Sequenz und der Genname enthält typischerweise COL als Präfix. Die Synthese wird durch die Aktivierung von bestimmten Genen gestartet, die für die Bildung von bestimmten α-Peptiden zuständig sind. Hauptsächlich sind dies die α-Peptide α1, α2 oder α3.
2. Translation: Sobald die mRNA vom Zellkern in das Cytoplasma gelangt, verbindet sich die mRNA zur Translation mit den ribosomalen Untereinheiten zur Bildung von Präpro-α-Ketten (auch Präpropeptid genannt). Diese Präpro-α-Ketten enthalten am N-Terminus eine Signalsequenz. Diese Signalsequenz wird durch ein Signalerkennungspartikel am rauen endoplasmatischen Retikulum (ER) erkannt, sodass die Kette in das Lumen des rauen ER gelangen kann.[8]
3. Präpro-α-Kette zum Prokollagen: Drei Modifikationen der Präpro-α-Kette sind zur Bildung einer α-Kette nötig. Danach folgt die Tripelhelixbildung dreier α-Ketten zum Prokollagen.
- Abspaltung der Signalsequenz: Die Signalsequenz am N-Terminus der Präpro-α-Kette wird durch Signalpeptidasen abgespalten und es entsteht eine Pro-α-Kette, die mit N- und C-terminalen Propeptiden versehen ist.[9]
- Hydroxylierung: Im Endoplasmatischen Retikulum werden an einzelne Prolin- und Lysin-Reste entstehender oder bereits entstandener Polypeptidketten OH-Gruppen angehängt (Hydroxylierung). Ascorbinsäure (Vitamin C) ist ein wichtiger Cofaktor bei der Hydroxylierung der Aminosäuren Prolin zu Hydroxyprolin durch das Enzym Prolyl-4-Hydroxylase[10] (EC 1.14.11.2) und Lysin zu Hydroxylysin durch das Enzym Lysylhydroxylase[11] (EC 1.14.11.4). Hydroxyprolin kommt die Funktion zu, über Wasserstoffbrücken zwischen benachbarten Kollagen-Polypeptidketten die Tripelhelix innerhalb eines Kollagenmoleküls zu festigen. Hydroxylysin dient der Verankerung von kovalenten Quervernetzungen zwischen Kollagenmolekülen. Bei fehlender Hydroxylierung werden nur schadhafte Kollagenmoleküle gebildet, die ihrer Funktion als Strukturprotein nicht nachkommen können. Hierbei ist anzumerken, dass nahezu alle Symptome der Ascorbinsäure-Mangelerkrankung Skorbut auf die fehlerhafte Biosynthese des Kollagens zurückzuführen sind.
- Glycosylierung: Schließlich erfolgt meist eine Glycosylierung mancher Lysinreste durch die Prokollagen-Galactosyltransferase (EC 2.4.1.50) oder die Prokollagen-Glucosyltransferase (EC 2.4.1.66) oder weitere Glycosylierungsenzyme. Eine Kollagen-α-Kette wurde synthetisiert.[12]
- Tripelhelixbildung: Durch Ausbildung von Disulfidbindungen zwischen den C-terminalen Propeptiden mithilfe einer Proteindisulfid-Isomerase (EC 5.3.4.1) wird die Tripelhelixbildung eingeleitet. Drei α-Ketten formieren dabei über Wasserstoffbrücken ein dreisträngiges Helixmolekül, das Prokollagen.
4. Prokollagen-Sekretion und Transport zum Golgi-Apparat: Aufgrund der Größe von Prokollagen-Molekülen (ca. 300 nm) passen sie nicht in die normalen COPII-Vesikel (50–90 nm) des endoplasmatischen Retikulums hinein. Um den Transport zu ermöglichen, wird eine Kopie des Proteins Ubiquitin durch das Enzym CUL3–KLHL12 an SEC31 des Proteinkomplexes SEC13–SEC31 gebunden, sodass die Größe der Transportvesikel modifiziert werden kann. Außerdem beteiligt sich das Transmembranprotein TANGO1 an der Koordination der Prokollagen-Sekretion. Nachdem das Prokollagen in den modifizierten Transportvesikeln verpackt worden ist, ist der Transport zum Golgi-Apparat möglich.[13]
5. Modifikation im Golgi-Apparat: Die letzte posttranslationale Modifikation am Prokollagen erfolgt im Golgi-Apparat durch das Anhängen und die Modifikation von Oligosacchariden. Welche Enzyme für die Modifizierung von N-terminusgebundenen Oligosacchariden verantwortlich sind, hängt vom Ort der posttranslationalen Modifikation im Golgi-Apparat ab. Im Folgenden sind die Enzyme für die jeweiligen Bereiche des Golgi-Apparats tabelliert:[14]
| Bereich des Golgi-Apparats | Enzym |
|---|---|
| cis | α-Mannosidase I |
| medial | N-Acetylglucosaminyl-Transferase I |
| medial | α-Mannosidase II |
| medial | N-Acetylglucosaminyl-Transferase II |
| medial | Fucosyltransferase |
| trans | Galactosyltransferase |
| trans | Sialyltransferase |
6. Transport vom Golgi-Apparat zur Plasmamembran und anschließende Exozytose in die extrazelluläre Matrix: Die sekretorischen Vesikel, die sich an der trans-Seite des trans-Golgi-Netzwerks abschnüren, bezeichnet man als Golgi to plasma membrane carrier (GPC). In den GPC wird das Prokollagen zur Plasmamembran, genauer gesagt zu den Vorwölbungen der Plasmamembran, sogenannte Fibripositoren, transportiert. Die tripelhelikalen Kollagenmoleküle werden aus der Zelle entlassen. Die Abgabe der Moleküle in den extrazellulären Raum erfolgt durch Exozytose mit der Basis der Fibripositoren, woran die Glycosylbestandteile beteiligt zu sein scheinen.[15]
7. Bildung des Tropokollagens: Unmittelbar nach der Abgabe aus der Zelle werden die Propeptide mit Hilfe von Prokollagen-Peptidasen abgespalten.[16] Dabei ist das Enzym Prokollagen-N-Endopeptidase (EC 3.4.24.14) bei der Abspaltung aminoterminaler Sequenzen erforderlich, während das Enzym Prokollagen-C-Endopeptidase (EC 3.4.24.19) carboxyterminale Prokollagen-Sequenzen abspaltet. Es kommt zur Bildung des Tropokollagens.
8. Fibrillogenese: Nach Abspaltung der Prokollagenpeptide lagern sich einzelne Tropokollagen-Moleküle zu Kollagenfibrillen zusammen (Fibrillogenese). Andere Moleküle können sich an die Fibrillen anlagern und somit den Fibrillendurchmesser anpassen. Dazu gehören die sogenannten small leucine-rich repeat proteoglycans (SLRPs), zu denen beispielsweise Decorin, Fibromodulin und Lumican gehören.[17][18]
.svg.png.webp)
9. Quervernetzung: Nachdem sich einzelne tripelhelikale Tropokollagenmoleküle um ein Fünftel ihrer Länge versetzt aneinander gelagert haben, erfolgen kovalente Quervernetzungen über erst umzuwandelnde nahestehende Hydroxylysinreste, womit die räumliche Anordnung dauerhaft fixiert wird. Die (intrazellulär durch die Lysylhydroxylase entstandenen) Hydroxylysinreste werden durch die Lysyloxidase (EC 1.4.3.13) zu Allysin oxidiert. Die beiden benachbarten Allysinreste gehen eine Aldolkondensation ein, womit diese Nachbarschaft durch eine dauerhafte Quervernetzung fixiert ist. Es kommt zur Bildung von quervernetzten Kollagenfibrillen.
10. Bildung von Kollagenfasern: Viele derartig kovalent stabilisierte Kollagenfibrillen bilden schließlich Kollagenfasern, welche die Grundstruktur der extrazellulären Matrix aller Gewebetiere darstellen.
Kollagentypen
Die Kollagene werden in mehrere Untergruppen unterteilt. 28 verschiedene Kollagentypen sind bekannt (Typ I bis XXVIII) sowie mindestens zehn weitere Proteine mit kollagenähnlichen Domänen. In der folgenden Zusammenstellung sind die bislang bekannten Mitglieder der Kollagenfamilie aufgeführt.
| Typ | Beschreibung | Gen(e) | Krankheiten |
| I | Bei Säugetieren ist Kollagen Typ I, ein fibrilläres Kollagen, der häufigste Kollagentyp und kommt in Haut, Sehnen, Faszien, Knochen, Gefäßen, inneren Organen und im Dentin vor. | COL1A1, COL1A2 | Osteogenesis imperfecta Typ I–IV, Ehlers-Danlos-Syndrom (klassisch, Arthrochalasie), Infantile kortikale Hyperostose |
| II | fibrillär; Strukturprotein des hyalinen und elastischen Knorpels; 50 % aller Proteine des Knorpels bestehen aus Kollagen Typ II; Bestandteil des Glaskörpers | COL2A1 | Hypochondrogenesie, Achondrogenesie Typ II, Stickler-Syndrom, Kongenitale Spondyloepiphysäre Dysplasie, Spondyloepimetaphysäre Dysplasie Typ Strudwick, Kniest-Dysplasie |
| III | fibrillär; Bestandteil des Granulationsgewebes und der retikulären Fasern; ebenfalls Bestandteil der Gefäßwände, inneren Organe, Haut, Uterus und Hornhaut | COL3A1 | Akrogerie, Ehlers-Danlos-Syndrom (vaskulär) |
| IV | Bestandteil der Basallamina und der Augenlinse; dient ebenfalls als Teil des Filtrationssystems in Kapillaren und der Glomeruli des Nephrons | COL4A1, COL4A2, COL4A3, COL4A4, COL4A5, COL4A6 | Alport-Syndrom, Goodpasture-Syndrom, Nephropathie vom Typ der dünnen Basalmembran |
| V | fibrillär; Bestandteil des Interstitiums, Plazentagewebe und der dermoepidermalen Junktionszone; meistens mit Zwischengewebe, die Kollagen Typ I beinhalten, assoziiert | COL5A1, COL5A2, COL5A3 | Ehlers-Danlos-Syndrom (klassisch, hypermobil) |
| VI | Organisation verschiedener Komponenten in der extrazellulären Matrix; Aufrechterhaltung der Integrität der verschiedenen Geweben; meistens mit Gewebe, die Kollagen Typ I beinhalten, assoziiert | COL6A1, COL6A2, COL6A3, COL6A5 | Kongenitale Muskeldystrophie Typ Ullrich, Bethlem-Myopathie, Atopisches Ekzem[19] |
| VII | bildet Ankerfibrillen in der dermoepidermalen Junktionszone | COL7A1 | Epidermolysis bullosa dystrophica, Bart-Syndrom |
| VIII | kurzkettig; integraler Bestandteil der subendothelialen Schicht von Bindegewebszellen in Blutgefäßen und der Descemet-Membran der Hornhaut; Migration und Proliferation von glatten Muskelzellen in der Tunica media der Blutgefäße | COL8A1, COL8A2 | Hintere-polymorphe-Hornhautdystrophie, Fuchs-Endotheldystrophie 1 |
| IX | FACIT1; Bestandteil des hyalinen Knorpels und des Glaskörpers; meistens mit Gewebe, die Kollagen Typ II und XI beinhalten, assoziiert | COL9A1, COL9A2, COL9A3 | Multiple epiphysäre Dysplasie (Typ 2,3 und 6) |
| X | kurzkettig; Bestandteil von Chondrozyten in der hypertrophen Zone | COL10A1 | Metaphysäre Chondrodysplasie Typ Schmid, Spondylometaphysäre Dysplasie Typ Sutcliffe |
| XI | fibrillär; Bestandteil des Knorpels | COL11A1, COL11A2 | Stickler-Syndrom Typ 2, Marshall-Syndrom, Weissenbacher-Zweymüller-Phänotyp, Oto-spondylo-megaepiphysäre Dysplasie |
| XII | FACIT, Interaktion mit Kollagenfibrillen Typ I, Decorin und Glykosaminoglykanen | COL12A1 | Bethlem-Myopathie 2, Kongenitale Muskeldystrophie Typ Ullrich 2 |
| XIII | MACIT2; interagiert mit Integrin α1β1, Fibronectin und anderen Bestandteilen der Basalmembran wie Nidogen-2 und Perlecan; ist in der Zell-Matrix-Verbindung und in der Zelladhäsion involviert; Verknüpfung der Muskelfaser mit der Basalmembran | COL13A1 | Kongenitales myasthenes Syndrom Typ 19 |
| XIV | FACIT; auch als Undulin bekannt; spielt eine Rolle bei der adhäsiven Integration von Kollagenfaszikeln; ist während der embryonalen Entwicklung in der Basalmembran präsent | COL14A1 | – |
| XV | Multiplexin; ist in der Lage, die Basalmembran mit dem lockeren Bindegewebe zu verbinden; stabilisiert Mikrogefäße und Muskelzellen im Herz und im Skelettmuskel; kann die Angiogenese inhibieren | COL15A1 | – |
| XVI | FACIT; in der Zelladhäsion involviert und induziert Integrin-vermittelte zelluläre Reaktionen, wie die Proliferation; fördert die Lebensdauer von intestinalen subepithelialen Myofibroblasten (engl. intestinal subepithelial myofibroblasts, ISEMF) in der Darmwand | COL16A1 | – |
| XVII | MACIT; spielt eine wichtige Rolle in der Integrität von Hemidesmosomen und in der Besfestigung von Keratinozyten an die darunterliegende Basalmembran; fördert die Invasion von extravillösen Trophoblasten während der Entwicklung der Plazenta | COL17A1 | Bullöses Pemphigoid, Epidermolysis bullosa junctionalis |
| XVIII | Multiplexin; Aufrechterhaltung der Netzhautstruktur und Schließung des Neuralrohrs; ein Spaltprodukt des Kollagen XVIII ist das Endostatin mit einer Molekülmasse von 20 kDa | COL18A1 | Knobloch-Syndrom Typ 1 |
| XIX | FACIT; befindet sich meistens in der vaskulären, neuronalen, mesenchymalen und in der epithelialen Basalmembran; Querbrücke zwischen Fibrillen und anderen extrazellulären Matrixmolekülen | COL19A1 | – |
| XX | FACIT; befindet sich meistens in der Epithelschicht der Hornhaut, der embryonalen Haut, dem Schwertfortsatz und in der Sehne | COL20A1 | – |
| XXI | FACIT; ist eine extrazelluläre Matrixkomponente der Blutgefäßwand, das von glatten Muskulaturzellen sekretiert wird | COL21A1 | – |
| XXII | FACIT; beteiligt sich an der Zelladhäsion von Liganden in mehrschichtigen Epithelien und in Fibroblasten | COL22A1 | – |
| XXIII | MACIT; befindet sich in der Epidermis und in anderen Epithelien wie in der Zunge, Darm, Lunge, Gehirn, Niere und Hornhaut; interagiert mit Integrin α2β1 | COL23A1 | – |
| XXIV | hauptsächlich im Knochengewebe exprimiert | COL24A1 | – |
| XXV | MACIT; fügt amyloide Fibrillen mit Protease-resistenten Aggregaten zusammen und kann Heparin binden | COL25A1 | – |
| XXVI | wird oftmals mit der Bildung von Nasenpolypen in der Nasenhöhle assoziiert | COL26A1 | – |
| XXVII | befindet sich in der Basalmembran der Keratinozyten in Hemidesmosomen Typ I, das im Wesentlichen als Adhäsionsmolekül und Oberflächenrezeptor auch in mehrschichtigen Epithelien fungiert | COL27A1 | – |
| XXVIII | befindet sich hauptsächlich im Ischiasnerv und an der Basalmembran einiger Schwann-Zellen; integraler Bestandteil des Ranvier-Schnürrings und umgibt nicht-myelinisierte Gliazellen | COL28A1 | – |
Aufbau des Kollagens Typ I
Die drei Kollagen-Polypeptidketten sind im Falle von Kollagen Typ I die α-Ketten, [α1(I)2α2(I)], die sich zu einer Tripelhelix umeinander winden. Das Gen der α1-Kette von Kollagen Typ I besteht aus 50 Exons, von denen über die Hälfte eine Länge von 54 Basenpaaren (bp) oder das zwei- bis dreifache dieser Länge besitzen. Sie codieren für die Sequenz (G-X-Y)6 oder ein Vielfaches davon.
Nutzung
Ernährung und Futterstoffe
In Deutschland werden jährlich etwa 32.000 t Gelatine in Speisequalität hergestellt, die europäische Gesamtproduktion beträgt 120.000 t (70 % Schweineschwarten, 18 % Knochen, 10 % Rinderspalt, 2 % sonstige).[20] Verwendet werden in Deutschland etwa 90.000 t, wobei zwei Drittel auf den Ernährungsbereich und von dem Rest etwa die Hälfte auf den Futtermittelbereich entfallen.[21]
Daneben wird Kollagen für die Herstellung von Kunstdärmen, die als Wursthülle dienen, benutzt.[22]
Pharma

Etwa 15.000 t werden in der chemischen und pharmazeutischen Industrie verarbeitet. Dabei stellen Umhüllungen von Tabletten und Vitaminpräparaten (Hart- und Weichkapseln) sowie Gelatinezäpfchen die Haupteinsatzbereiche in der Pharmaindustrie dar. Außerdem wird Gelatine für blutstillende Schwämmchen sowie als Blutplasma-Ersatz eingesetzt.
Kollagen wird außerdem in der regenerativen Medizin als Nährmedium in der Gewebezüchtung benutzt. Damit können beispielsweise Hautersatzmaterialien für schwere Verbrennungen hergestellt werden.[22]
Kosmetik
Kollagen findet seit vielen Jahren auch in der Kosmetik Anwendung und soll dort hauptsächlich zur Minderung von Hautalterung, dem Anti-Aging, dienen.
Eine investigative Reportage des Guardian konnte 2023 zeigen, dass Produzenten weltweit Kollagen von Kühen verkaufen, die auf Farmen gehalten werden, welche für die Abholzung des Regenwaldes verantwortlich sind.[23]
Kollagen-Cremes gehören zu den Bestsellern auf dem Kosmetikmarkt. Allerdings kann das enthaltene Kollagen nicht in die Haut eindringen, da seine Fasern zu lang sind. Studien konnten bisher nicht zeigen, dass kürzere Fasern (Peptide) in die Haut eindringen können. Neben Cremes werden auch Supplemente zur oralen Einnahme verkauft. Bislang wurden die meisten, wenn nicht alle, Studien zu Kollagen von Industrien finanziert, welche durch den Verkauf von Kollagenprodukten Vorteile hätte. Daher kann kein unabhängiges Urteil über die Effektivität von Kollagen getroffen werden.[24]
Zur Behandlung von Falten oder Runzeln wird Kollagen als Faltenunterspritzung gelegentlich in die Haut injiziert.
Technik
In der analogen Fotografie stellt Gelatine die Basis für die fotoempfindlichen Schichten auf dem Film und dem Fotopapier. Auch moderne Druckerpapiere zum Ausdrucken von Farbbildern sind mit Gelatine beschichtet.[21]
Leder
Vernetzte Kollagenfasern bilden die Struktur von Leder und geben ihm seine Zugfestigkeit. Mit Hilfe von Gerbstoffen werden bestimmte Eigenschaften wie Flexibilität und Resistenz gegen Zersetzung durch Mikroorganismen erreicht.
Gewinnung
Kollagen wird vor allem in Form der Gelatine genutzt, die aus Rinderspalt, Schweineschwarten sowie Knochen von Rindern und Schweinen gewonnen wird. Nach dieser Herstellungsmethode kann das Kollagen denaturiert vorliegen und verunreinigt sein. Diese Form kann somit ungeeignet für manche Anwendungen sein.[25] Rein synthetisches Kollagen kann beispielsweise über biotechnologische Prozesse mittels genetisch modifizierten Mikroorganismen hergestellt werden.[26]
Siehe auch

Literatur
- Shirley Ayad, Ray P. Boot-Hanford, Martin J. Humphries, Karl E. Kadler, C. Adrian Shuttleworth: The Extracellular Matrix FactsBook. 2nd edition. Academic Press (Harcourt Brace & Company), San Diego CA u. a. 1998, ISBN 0-12-068911-1, S. 43 ff.
- Jürgen Brinckmann, Holger Notbohm, Peter K. Müller (Hrsg.): Collagen. Primer in Structure, Processing and Assembly (= Topics in Current Chemistry. Bd. 247). Springer, Berlin u. a. 2005, ISBN 3-540-23272-9.
Weblinks
Einzelnachweise
- Hermann Ehrlich: Chitin and collagen as universal and alternative templates in biomineralization. In: International Geology Review. Band 52, Nr. 7-8, 30. April 2010, S. 661–699, doi:10.1080/00206811003679521 (englisch).
- Aaron L. Fidler u. a.: A unique covalent bond in basement membrane is a primordial innovation for tissue evolution. In: Proceedings of the National Academy of Sciences. Band 111, Nr. 1, 2014, S. 331–336. doi:10.1073/pnas.1318499111.
- Jason W. Holland u. a.: A novel minicollagen gene links cnidarians and myxozoans. In: Proceedings of the Royal Society B: Biological Sciences. Band 278, Nr. 1705, 2011, S. 546–553. doi:10.1098/rspb.2010.1301.
- Richard P. Tucker, Josephine C. Adams: Adhesion networks of cnidarians: A postgenomic view. In: Kwang W. Jeon (Hrsg.): International Review of Cell and Molecular Biology. Academic Press. Band 308, Kapitel 8, 7. Januar 2014, S. 323–377. doi:10.1016/B978-0-12-800097-7.00008-7.
- Klaus Kühn: Struktur und Biochemie des Kollagens. In: Chemie in unserer Zeit. Band 8, 1974, S. 97–103. doi:10.1002/ciuz.19740080402.
- Gregory D. Cramer, Susan A. Darby: Basic and Clinical Anatomy of the Spine, Spinal Cord. Elsevier Health Sciences, 2005, ISBN 0-323-07142-2, S. 580 (eingeschränkte Vorschau in der Google-Buchsuche).
- Gerhard Meisenberg, William H. Simmons: Principles of Medical Biochemistry E-Book. Elsevier Health Sciences, 2011, ISBN 978-0-323-08107-8, S. 212.
- Hao Wang, Branko Stefanovic: Role of LARP6 and Nonmuscle Myosin in Partitioning of Collagen mRNAs to the ER Membrane. In: PLoS One. 9(10) 2014, PMID 25271881, doi:10.1371/journal.pone.0108870.
- U. Lindert, M. Gnoli, M. Maioli, M.F. Bedeschi, L. Sangiorgi, M. Rohrbach, C. Giunta: Insight into the Pathology of a COL1A1 Signal Peptide Heterozygous Mutation Leading to Severe Osteogenesis Imperfecta. In: Calcified Tissue International. 102(3) März 2018, S. 373–379, doi:10.1007/s00223-017-0359-z.
- J. Myllyharju: Prolyl 4-hydroxylases, the key enzymes of collagen biosynthesis. In: Matrix Biology. Band 22, Nummer 1, März 2003, S. 15–24, PMID 12714038 (Review).
- Sakari Kellokumpu, Raija Sormunen, Jari Heikkinen, Raili Myllylä: Lysyl Hydroxylase, a Collagen Processing Enzyme, Exemplifies a Novel Class of Luminallyloriented Peripheral Membrane Proteins in the Endoplasmic Reticulum. In: The Journal of Biological Chemistry. 269(148) 2. Dezember 1994, S. 30524–30529, (PDF).
- Richard Harwood, Michael E. Grant, David S. Jackson: Studies on the glycosylation of hydroxylysine residues during collagen biosynthesis and the subcellular localization of collagen galactosyltransferase and collagen glucosyltransferase in tendon and cartilage cells. In: Biochemical Journal. 152, 1975, S. 291, doi:10.1042/bj1520291.
- David J. Stephens: Collagen secretion explained. In: Nature. 482, 2012, S. 474, doi:10.1038/482474a.
- R. Kornfeld, S. Kornfeld: Assembly of Asparagine-Linked Oligosaccharides. In: Annual Review of Biochemistry. 54, 1985, S. 631, doi:10.1146/annurev.bi.54.070185.003215.
- E. G. Canty: Procollagen trafficking, processing and fibrillogenesis. In: Journal of Cell Science. 118, 2005, S. 1341, doi:10.1242/jcs.01731.
- Charles M. Lapière, Albert Lenaers, Leonard D. Kohn: Procollagen peptidase: An enzyme excising the coordination peptides of procollagen. In: Proc Natl Acad Sci U S A. Band 68, Nr. 12, Dezember 1971, S. 3054–3058, PMC 389589 (freier Volltext).
- A. Oldberg, P. Antonsson, K. Lindblom, D. Heinegård: A collagen-binding 59-kd protein (fibromodulin) is structurally related to the small interstitial proteoglycans PG-S1 and PG-S2 (decorin). In: The EMBO Journal. Band 8, Nummer 9, September 1989, S. 2601–2604, PMID 2531085, PMC 401265 (freier Volltext).
- J. A. Rada, P. K. Cornuet, J. R. Hassell: Regulation of corneal collagen fibrillogenesis in vitro by corneal proteoglycan (lumican and decorin) core proteins. In: Experimental eye research. Band 56, Nummer 6, Juni 1993, S. 635–648, doi:10.1006/exer.1993.1081, PMID 8595806.
- Cilla Söderhäll, Ingo Marenholz, Tamara Kerscher, Franz Rüschendorf, Jorge Esparza-Gordillo, Margitta Worm, Christoph Gruber, Gabriele Mayr, Mario Albrecht, Klaus Rohde, Herbert Schulz, Ulrich Wahn, Norbert Hubner, Young-Ae Lee: Variants in a Novel Epidermal Collagen Gene (COL29A1) Are Associated with Atopic Dermatitis. In: PLOS Biology. Band 5, Nr. 9, 2007, S. e242, doi:10.1371/journal.pbio.0050242, PMID 17850181.
- K. Rappold: Gelatine – Ein natürliches Nahrungsmittel. bmi aktuell 1/2004, Hrsg. Informationszentrale für Backmittel und Backgrundstoffe zur Herstellung von Brot und Feinen Backwaren e. V.
- Gelatine. Gelatine Manufacturers of Europe, abgerufen am 23. Mai 2013.
- Oliver Türk: Stoffliche Nutzung nachwachsender Rohstoffe. 1. Auflage. Springer Vieweg, Wiesbaden 2014, ISBN 978-3-8348-1763-1, S. 111–115.
- Andrew Wasley, Elisângela Mendonça, Fábio Zuker: Global craze for collagen linked to Brazilian deforestation. In: The Guardian. 6. März 2023 (theguardian.com [abgerufen am 6. März 2023]).
- Harvard T.H. Chan School of Public Medicine: Collagen. 26. Mai 2021, abgerufen am 6. März 2023 (amerikanisches Englisch).
- David Olsen, Chunlin Yang, Michael Bodo, Robert Chang, Scott Leigh, Julio Baez, David Carmichael, Maritta Perälä, Eija-Riitta Hämäläinen, Marko Jarvinen, James Polarek: Recombinant collagen and gelatin for drug delivery. In: Advanced Drug Delivery Reviews (= Collagen in drug delivery and tissue engineering). Band 55, Nr. 12, 28. November 2003, ISSN 0169-409X, S. 1547–1567, doi:10.1016/j.addr.2003.08.008 (sciencedirect.com [abgerufen am 1. April 2023]).
- Julio Báez, David Olsen, James W. Polarek: Recombinant microbial systems for the production of human collagen and gelatin. In: Applied Microbiology and Biotechnology. Band 69, Nr. 3, 1. Dezember 2005, ISSN 1432-0614, S. 245–252, doi:10.1007/s00253-005-0180-x.
- PDB Community Focus: Julian Voss-Andreae, Protein Sculptor. In: Protein Data Bank Newsletter. Nr. 32, 2007 (wwpdb.org (Memento vom 22. Mai 2023 im Internet Archive) [PDF; 2,0 MB]).
- Barbara Ward: ‘Unraveling Collagen’ structure to be installed in Orange Memorial Park Sculpture Garden. In: Expert Rev. Proteomics. Band 3, Nr. 2, April 2006, S. 174, doi:10.1586/14789450.3.2.169.