Braunes Fettgewebe
Das braune oder plurivakuoläre Fettgewebe ist eine spezielle Form des Fettgewebes, dessen Zellen in der Lage sind, durch die Oxidation von Fettsäuren Wärme zu produzieren (Thermogenese). Dies geschieht in zahlreichen Mitochondrien, die auch für die gelb-bräunliche Färbung des Gewebes verantwortlich sind. Biochemisch wird durch das Membranprotein Thermogenin die Fettsäureoxidation von der Synthese des Energieträgers Adenosintriphosphat (ATP) entkoppelt, so dass die freigesetzte Energie in Wärme umgesetzt wird.
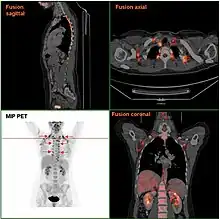
Vorkommen
Braunes Fettgewebe findet sich bei allen neugeborenen Säugetieren außer beim Schwein.[1] Neugeborene sind stärker durch Auskühlung bedroht, da sie auf Grund größerer Körperoberfläche im Vergleich zum Volumen mehr Wärme verlieren, und die Mechanismen der Thermoregulation (z. B. isolierendes weißes Fettgewebe und Kältezittern) noch nicht vollständig ausgebildet sind. Beim menschlichen Säugling findet sich braunes Fettgewebe vor allem an Hals, an den Nieren und zwischen den Schulterblättern.
Besonders Nagetiere besitzen auch im erwachsenen Zustand noch größere Mengen braunen Fettgewebes und können bei Bedarf mittels Katecholaminen weißes in braunes Fettgewebe umwandeln und so Kältephasen gut überstehen. Bei Winterschlaf haltenden Tieren finden sich ebenfalls größere Mengen braunen Fettgewebes, die der schnellen Erwärmung des Tieres in den Aufwachphasen dienen.
In manchen Vögeln treten histologisch ähnliche Gewebe auf, die jedoch kein Thermogenin aufweisen und nicht der Thermogenese dienen.[2] Allerdings können manche Vögel in den Skelettmuskeln über biochemisch ähnliche Mechanismen Wärme erzeugen.[3]
Vorkommen bei erwachsenen Menschen
Für Erwachsene wurde angenommen, dass sie über keine aktiven braunen Fettzellen mehr verfügten. 2009 konnte in einer multinationalen Studie die Aktivität von braunem Fettgewebe bei Erwachsenen durch neue Methoden der funktionellen Bildgebung belegt werden. Hierzu wurde die klare Korrelation zwischen Aufnahme von Glucose in braunes Fettgewebe, gezeigt durch markierte Glucose in der Positronen-Emissions-Tomographie, und dem Body Mass Index dargestellt.[4] Zwischenzeitlich ist klar, dass die Aktivität des braunen Fettgewebes bei Erwachsenen hoch variabel ist und von einer Vielzahl innerer und äußerer Einflüsse abhängt und mittels verschiedener Techniken nachgewiesen werden kann.[5] Während das klassische weiße Fett sich besonders unterhalb der Haut und im Bereich des Bauchs findet, ist das braune Fettgewebe oberhalb der Schlüsselbeine (supraclaviculär) und neben den Wirbeln (paravertebral) der oberen Brust- und unteren Halswirbelsäule lokalisiert.[6] Frauen scheinen einen höheren Anteil aktiven braunen Fettgewebes zu haben.[7]
Entwicklung und Formen
Braunes Fettgewebe
Das braune Fettgewebe ist ein distinkte Zellgruppe, die aus dem Mesoderm stammt und damit einen gemeinsamen Ursprung mit Muskelzellen, Knochenzellen und Bindegewebe hat. Der genaue Ablauf der Differenzierung des Mesoderms zu braunen Fettgewebszellen ist unklar. Braunes Fettgewebe unterscheidet sich in der Entwicklung von weißen Fettgewebe dahingehend, dass es einen gemeinsamen Vorläufer mit der Muskelzelle teilt, was angesichts der metabolischen Ähnlichkeiten Sinn ergibt. Jedoch fanden sich auch braune Fettgewebszellen ohne den gemeinsamen Vorläufer mit der Muskelzelle, weswegen Forscher davon ausgehen, dass die braunen Fettgewebszellen verschiedene Ursprünge haben und sich auch leicht als ausdifferenzierte Zellen unterscheiden, was auch als Adipozyten-Heterogenität bezeichnet wird.[8]
Beiges Fettgewebe
Weiße, subkutan gelegene Fettzellen haben die Möglichkeit sich zu "bräunen", also zu Zellen zu werden, die braunen Fettgewebszellen mit plurivakuolären Fetttröpfchen, erhöhter Mitochondrienzahl und Möglichkeit zu Thermogenese ähneln. Diese braunen Fettzellen im weißen Fettgewebe werden deshalb als beige Fettzellen bezeichnet. Diese beige Zellen unterscheiden sich in ihrem genetischen Profil von weißen und braunen Fettgewebszellen, sind aber den braunen Fettgewebszellen in Funktion deutlich näher.[9] Das sogenannte Bräunen wird über Sympathikus-Aktivität und Noradrenalin eingeleitet und damit mit den gleichen Prozessen wie der Aktivierung von braunen Fettgewebe. Weitere Mechanismen des Bräunens sind
- Myokine, die bei körperlicher Betätigung ausgeschüttet werden.
- Das Schilddrüsenhormon Triiodthyronin, was auch die Wärmeproduktion bei einer Schilddrüsenüberfunktion mit erklärt.
- Der in der Leber produzierte Fibroblast growth factor 21, der Wirkungen auf Lipid- und Glucosemetabolismus hat.
- kardiale natriuretische Peptide.
- Adiponektin, was das weiße Fettgewebe selber herstellen kann.
- Peripheres Serotonin, welches als Gegenspieler wirkt, der das Bräunen verhindert, wohingegen Serotonin im Gehirn eher zu Umwandlung von weißen in beige Fettzellen führt.
Die Adipozyten sind also in die gesamte metabolische und sportliche Situation des Organismus eingebunden. Daneben können auch exogene Stoffe zu einer Umwandlung führen, hierzu gehören Capsaicin, Protoalkaloide der Bitterorange, Fucoxanthin aus manchen Algen und Carotinoide, wobei unklar ist, in welchen Ausmaß diese Stoffe das weiße Fettgewebe beeinflussen können.[10]
Histologie
Die Zellen des braunen Fettgewebes sind allgemein kleiner als die in weißem Fettgewebe und haben viele, kleinere Lipidtropfen. Sie werden daher im Gegensatz zu den univakuolären Zellen des weißen Fettgewebes als plurivakuolär bezeichnet. Zudem zeichnen sie sich durch besonderen Reichtum an Mitochondrien aus, die aufgrund ihres Gehalts an eisenhaltigen Cytochromen auch für die braune Farbe verantwortlich sind. Braunes Fettgewebe ist stark durchblutet, um die produzierte Wärme auch an den Körper weiterleiten zu können.[11]
Biochemie und Regulation
Das in braunem Fettgewebe in der inneren Membran der Mitochondrien vorhandene Protein Thermogenin (auch Uncoupling Protein I genannt) dient als Entkoppler, indem es als Uniporter Protonen über die Membran transportiert. Hierdurch wird der durch β-Oxidation und Atmungskette aufgebaute Protonengradient abgebaut und die darin gespeicherte Energie in Wärme umgesetzt (Thermogenese) und von der Bildung von Adenosintriphosphat (ATP) entkoppelt. Daneben weist braunes Fettgewebe eine besonders hohe Konzentration an Glycerokinase auf, so dass das beim Fettabbau freiwerdende Glycerin phosphoryliert und ebenfalls metabolisch umgesetzt werden kann. In welchen Anteilen die braunen Fettzellen eigene gespeicherte Fette abbauen, Fettsäuren aus dem Blut verwenden oder mittels Glucose Fettsäure de-novo synthetisieren ist noch unklar.[12]
Die genaue Menge der produzierten Wärme muss seitens des Körpers gut reguliert werden und an den Bedarf angepasst werden. Über die drei Faktoren Kälte, Sympathikus-Aktivität und Botenstoffe wird das braune Fettgewebe gesteuert. Zum einen wird die Thermogenese im braunen Fettgewebe über das Hormon Noradrenalin aktiviert, das über einen G-Protein-gekoppelten β-Rezeptor (β3-AR) die Adenylatcyclase aktiviert. Das gebildete intrazelluläre cAMP aktiviert wiederum die Proteinkinase A, die in einem nächsten Schritt über Phosphorylierung von Lipasen den Fettabbau einleitet. Zudem ist braunes Fettgewebe sympathisch innerviert.
Aktivität und Bildung von braunem Fettgewebe wird vom PGC-1alpha (Peroxisome proliferator-activated receptor-gamma coactivator) gesteigert,[13] der wiederum stark nach Kältereiz ausgeschüttet wird.[13] Eine Vielzahl anderer körpereigener und exogen zugeführter (Boten-)Stoffe sind an der Aktivität von braunen Fettgewebe in unterschiedlichen Ausmaß beteiligt.[14] Bei Ratten führt beispielsweise Nahrungsaufnahme zu einem Anstieg der Körpertemperatur bis 1 °C.[15] Braunes Fettgewebe ist aber nicht nur Empfänger, sondern sendet selbst Botenstoffe aus, die als Batokine (vom englischen Brown Adipose Tissue) bezeichnet werden und einen Effekt auf den Gesamtmetabolismus haben.[16]
Die Produktion von braunem Fettgewebe sowie dessen Aktivität kann durch die Einnahme von Statinen reduziert bzw. eingeschränkt werden.[17]
Funktion bei Menschen
Das braune Fettgewebe kann einen relevanten Anteil am gesamten Energieumsatz haben. Mittels FDG-PET konnte bei gesunden Freiwilligen ein relevanter Glucose-Verbrauch nach Kälteexposition nachgewiesen werden.[18] Die Aktivität des braunen Fettgewebes hängt von der Außentemperatur ab: Studien im Sommer zeigen weniger aktives Fettgewebe als im Winter, was auch zu unterschiedlichen Angaben der Häufigkeit von aktivem braunen Fettgewebe bei Erwachsenen führen kann.[19]
Neben der Wärmeproduktion greift das braune Fettgewebe positiv in den Glucose- und Fettmetabolismus ein.[20]
Mit zunehmendem Alter nimmt bei Erwachsenen das braune Fettgewebe in Umfang und Aktivität ab, möglicherweise durch externe Isolation in Form von Kleidung und damit geringerer Notwendigkeit der eigenen Thermogenesis.
Das braune Fettgewebe ist als mögliches Ziel zur Therapie des metabolischen Syndroms und der Adipositas von Interesse. Bereits 1979 erfolgten hierzu erste Überlegungen.[21] Jedoch gab es in der pharmakologischen Entwicklung entsprechender Therapien Rückschläge, weil sich bei Stimulation des braunen Fettgewebes auch Atherosklerose bilden kann, jedoch wird an der Studie, die dies zeigte, kritisiert, dass die Mäuse zu schnell und zu stark einer kalten Umgebung ausgesetzt worden waren, was zu einer starken Stressreaktion, unabhängig der Stimulation des braunen Fettgewebes führte.[22] Erste kleinere Studien in den letzten Jahren mit dem β3-Rezeptoragonist Mirabegron konnten einen positiven Effekt auf das metabolische Profil über Stimulation des braunen Fettgewebes nachweisen.[23]
Literatur
- Georg Löffler, Petro E. Petrides, Peter C. Heinrich: Biochemie und Pathobiochemie. 8. Auflage. Springer, Heidelberg 2006, ISBN 3-540-32680-4.
- Werner A. Müller: Tier- und Humanphysiologie. Springer, Heidelberg 1998, ISBN 3-540-63313-8.
- Ulrich Welsch: Lehrbuch Histologie. Urban & Fischer, München 2002. 3. Auflage unter Mitarbeit von Thomas Deller 2010. S. 122.
Einzelnachweise
- Frida Berg, Ulla Gustafson, Leif Andersson: The Uncoupling Protein 1 Gene (UCP1) Is Disrupted in the Pig Lineage: A Genetic Explanation for Poor Thermoregulation in Piglets. In: PLoS Genetics. 18. August 2006.
- Seppo Saarelaa, Jacqueline S. Keithb, Esa Hohtolaa, Paul Trayhurn: Is the “mammalian” brown fat-specific mitochondrial uncoupling protein present in adipose tissues of birds? In: Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology. Band 100, Nr. 1, 1991, S. 45–49 (englisch).
- Darren A. Talbot, Claude Duchamp, Benjamin Rey, Nicolas Hanuise, Jean Louis Rouanet, Brigitte Sibille, Martin D. Brand: Uncoupling protein and ATP/ADP carrier increase mitochondrial proton conductance after cold adaptation of king penguins. In: The Journal of Physiology. Band 558, Nr. 1, 2004, S. 123–135 (englisch, physoc.org).
- Aaron M. Cypess, Sanaz Lehman, Gethin Williams, Ilan Tal, Dean Rodman, Allison B. Goldfine, Frank C. Kuo, Edwin L. Palmer, Yu-Hua Tseng, Alessandro Doria, Gerald M. Kolodny, C. Ronald Kahn: Identification and Importance of Brown Adipose Tissue in Adult Humans. In: New England Journal of Medicine. Band 360, Nr. 15, 9. April 2009, ISSN 0028-4793, S. 1509–1517, doi:10.1056/NEJMoa0810780, PMID 19357406, PMC 2859951 (freier Volltext) – (nejm.org [abgerufen am 18. Februar 2023]).
- Jing Yang, Haili Zhang, Kadirya Parhat, Hui Xu, Mingshuang Li, Xiangyu Wang, Chongzhao Ran: Molecular Imaging of Brown Adipose Tissue Mass. In: International Journal of Molecular Sciences. Band 22, Nr. 17, 30. August 2021, ISSN 1422-0067, S. 9436, doi:10.3390/ijms22179436, PMID 34502347, PMC 8431742 (freier Volltext) – (mdpi.com [abgerufen am 18. Februar 2023]).
- K. L. Townsend, Y.-H. Tseng: Of mice and men: novel insights regarding constitutive and recruitable brown adipocytes. In: International Journal of Obesity Supplements. Band 5, Suppl 1, August 2015, ISSN 2046-2166, S. S15–20, doi:10.1038/ijosup.2015.5, PMID 27152169, PMC 4850574 (freier Volltext).
- Christina Pfannenberg, Matthias K. Werner, Sabine Ripkens, Irina Stef, Annette Deckert, Maria Schmadl, Matthias Reimold, Hans-Ulrich Häring, Claus D. Claussen, Norbert Stefan: Impact of Age on the Relationships of Brown Adipose Tissue With Sex and Adiposity in Humans. In: Diabetes. Band 59, Nr. 7, 1. Juli 2010, ISSN 0012-1797, S. 1789–1793, doi:10.2337/db10-0004, PMID 20357363, PMC 2889780 (freier Volltext) – (diabetesjournals.org [abgerufen am 18. Februar 2023]).
- Su Myung Jung, Joan Sanchez-Gurmaches, David A. Guertin: Brown Adipose Tissue Development and Metabolism. In: Brown Adipose Tissue. Band 251. Springer International Publishing, Cham 2018, ISBN 978-3-03010512-9, S. 3–36, doi:10.1007/164_2018_168, PMID 30203328, PMC 7330484 (freier Volltext) – (springer.com [abgerufen am 18. Februar 2023]).
- T. Montanari, N. Pošćić, M. Colitti: Factors involved in white-to-brown adipose tissue conversion and in thermogenesis: a review: Factors involved in WAT browning. In: Obesity Reviews. Band 18, Nr. 5, Mai 2017, S. 495–513, doi:10.1111/obr.12520 (wiley.com [abgerufen am 18. Februar 2023]).
- T. Montanari, N. Pošćić, M. Colitti: Factors involved in white-to-brown adipose tissue conversion and in thermogenesis: a review: Factors involved in WAT browning. In: Obesity Reviews. Band 18, Nr. 5, Mai 2017, S. 495–513, doi:10.1111/obr.12520 (wiley.com [abgerufen am 18. Februar 2023]).
- T. Montanari, N. Pošćić, M. Colitti: Factors involved in white-to-brown adipose tissue conversion and in thermogenesis: a review: Factors involved in WAT browning. In: Obesity Reviews. Band 18, Nr. 5, Mai 2017, S. 495–513, doi:10.1111/obr.12520 (wiley.com [abgerufen am 18. Februar 2023]).
- Su Myung Jung, Joan Sanchez-Gurmaches, David A. Guertin: Brown Adipose Tissue Development and Metabolism. In: Brown Adipose Tissue. Band 251. Springer International Publishing, Cham 2018, ISBN 978-3-03010512-9, S. 3–36, doi:10.1007/164_2018_168, PMID 30203328, PMC 7330484 (freier Volltext) – (springer.com [abgerufen am 18. Februar 2023]).
- Huiyun Liang, Walter Ward: PGC-1alpha: a key regulator of energy metabolism. In: Advan. Physiol. Edu. 30, 2006, S. 145–151, doi:10.1152/advan.00052.2006, Volltext (Memento vom 23. November 2010 im Internet Archive) (englisch).
- Rafael C. Gaspar, José R. Pauli, Gerald I. Shulman, Vitor R. Muñoz: An update on brown adipose tissue biology: a discussion of recent findings. In: American Journal of Physiology-Endocrinology and Metabolism. Band 320, Nr. 3, 1. März 2021, ISSN 0193-1849, S. E488–E495, doi:10.1152/ajpendo.00310.2020, PMID 33459179, PMC 7988785 (freier Volltext) – (physiology.org [abgerufen am 18. Februar 2023]).
- Rachel J. Perry, Kun Lyu, Aviva Rabin-Court, Jianying Dong, Xiruo Li, Yunfan Yang, Hua Qing, Andrew Wang, Xiaoyong Yang, Gerald I. Shulman: Leptin mediates postprandial increases in body temperature through hypothalamus–adrenal medulla–adipose tissue crosstalk. In: Journal of Clinical Investigation. Band 130, Nr. 4, 9. März 2020, ISSN 0021-9738, S. 2001–2016, doi:10.1172/JCI134699, PMID 32149734, PMC 7108915 (freier Volltext) – (jci.org [abgerufen am 18. Februar 2023]).
- Francesc Villarroya, Rubén Cereijo, Joan Villarroya, Marta Giralt: Brown adipose tissue as a secretory organ. In: Nature Reviews Endocrinology. Band 13, Nr. 1, Januar 2017, ISSN 1759-5029, S. 26–35, doi:10.1038/nrendo.2016.136 (nature.com [abgerufen am 18. Februar 2023]).
- Inhibition of Mevalonate Pathway Prevents Adipocyte Browning in Mice and Men by Affecting Protein Prenylation. In: Cell Metabolism. 20. Dezember 2018, ISSN 1550-4131, doi:10.1016/j.cmet.2018.11.017 (sciencedirect.com [abgerufen am 23. Dezember 2018]).
- M Matsushita, T Yoneshiro, S Aita, T Kameya, H Sugie, M Saito: Impact of brown adipose tissue on body fatness and glucose metabolism in healthy humans. In: International Journal of Obesity. Band 38, Nr. 6, Juni 2014, ISSN 0307-0565, S. 812–817, doi:10.1038/ijo.2013.206 (nature.com [abgerufen am 18. Februar 2023]).
- Masayuki Saito, Yuko Okamatsu-Ogura, Mami Matsushita, Kumiko Watanabe, Takeshi Yoneshiro, Junko Nio-Kobayashi, Toshihiko Iwanaga, Masao Miyagawa, Toshimitsu Kameya, Kunihiro Nakada, Yuko Kawai, Masayuki Tsujisaki: High Incidence of Metabolically Active Brown Adipose Tissue in Healthy Adult Humans. In: Diabetes. Band 58, Nr. 7, 1. Juli 2009, ISSN 0012-1797, S. 1526–1531, doi:10.2337/db09-0530, PMID 19401428, PMC 2699872 (freier Volltext) – (diabetesjournals.org [abgerufen am 18. Februar 2023]).
- Rajan Singh, Albert Barrios, Golnaz Dirakvand, Shehla Pervin: Human Brown Adipose Tissue and Metabolic Health: Potential for Therapeutic Avenues. In: Cells. Band 10, Nr. 11, 5. November 2021, ISSN 2073-4409, S. 3030, doi:10.3390/cells10113030, PMID 34831253, PMC 8616549 (freier Volltext) – (mdpi.com [abgerufen am 18. Februar 2023]).
- Nancy J. Rothwell, Michael J. Stock: A role for brown adipose tissue in diet-induced thermogenesis. In: Nature. Band 281, Nr. 5726, September 1979, ISSN 0028-0836, S. 31–35, doi:10.1038/281031a0 (nature.com [abgerufen am 18. Februar 2023]).
- Mei Dong, Xiaoyan Yang, Sharon Lim, Ziquan Cao, Jennifer Honek, Huixia Lu, Cheng Zhang, Takahiro Seki, Kayoko Hosaka, Eric Wahlberg, Jianmin Yang, Lei Zhang, Toste Länne, Baocun Sun, Xuri Li, Yizhi Liu, Yun Zhang, Yihai Cao: Cold Exposure Promotes Atherosclerotic Plaque Growth and Instability via UCP1-Dependent Lipolysis. In: Cell Metabolism. Band 18, Nr. 1, Juli 2013, S. 118–129, doi:10.1016/j.cmet.2013.06.003, PMID 23823482, PMC 3701322 (freier Volltext) – (elsevier.com [abgerufen am 18. Februar 2023]).
- Alana E. O’Mara, James W. Johnson, Joyce D. Linderman, Robert J. Brychta, Suzanne McGehee, Laura A. Fletcher, Yael A. Fink, Devika Kapuria, Thomas M. Cassimatis, Nathan Kelsey, Cheryl Cero, Zahraa Abdul Sater, Francesca Piccinini, Alison S. Baskin, Brooks P. Leitner, Hongyi Cai, Corina M. Millo, William Dieckmann, Mary Walter, Norman B. Javitt, Yaron Rotman, Peter J. Walter, Marilyn Ader, Richard N. Bergman, Peter Herscovitch, Kong Y. Chen, Aaron M. Cypess: Chronic mirabegron treatment increases human brown fat, HDL cholesterol, and insulin sensitivity. In: Journal of Clinical Investigation. Band 130, Nr. 5, 23. März 2020, ISSN 0021-9738, S. 2209–2219, doi:10.1172/JCI131126, PMID 31961826, PMC 7190915 (freier Volltext) – (jci.org [abgerufen am 18. Februar 2023]).