Schädeltypen der Amnioten
Beim Schädel der Amniota lassen sich anhand der Anzahl und Lage bestimmter Schädelfenster im Deckknochen des Schädeldachs drei Basistypen unterscheiden: anapsider, synapsider und diapsider Schädel. Diese drei Typen besaßen in der Vergangenheit eine relativ hohe Bedeutung für die innere Systematik der Amnioten (bzw. der Reptilien im traditionellen Sinn) und sind namensgebend für die Großgruppen Anapsida, Synapsida und Diapsida. Heute ist die Bedeutung der Schädelöffnungen für die Systematik etwas geringer, da neuere Untersuchungen ergeben haben, dass gleiche oder ähnliche Schädelkonfigurationen bei verschiedenen Amniotengroßgruppen auftreten.
Anapsider Schädel


Der anapside Schädel ist durch das Fehlen von Schädelfenstern im Schädeldach gekennzeichnet. Dieser Typus repräsentiert vermutlich den ursprünglichen Zustand des Schädels der Amnioten und wurde von deren nicht-amniotischen Vorfahren ererbt.
Taxa mit anapsidem Schädel sind in beiden Hauptlinien der Reptilien, Parareptilien und Eureptilien, vertreten. Mit Ausnahme der Schildkröten handelt es sich dabei ausschließlich um sehr frühe, lange ausgestorbene Formen wie zum Beispiel die Captorhiniden (Eureptilia) oder die Pareiasaurier (Parareptilia).
Die Schildkröten sind die einzige rezente Reptiliengruppe mit anapsidem Schädelbau. Jedoch stellt deren anapsider Schädel wahrscheinlich nicht den ursprünglichen Zustand dar, sondern ist durch sekundären Verschluss vormals vorhandener Schädelfenster zustande gekommen.[1][2]
Da der anapside Schädel ein ursprüngliches Merkmal ist, in beiden Hauptlinien der Reptilien auftritt und bei den Schildkröten offenbar sogar sekundär neu erworben wurde, besitzt die Abwesenheit von Schläfenfenstern bei Amnioten heute keine systematische Relevanz mehr. Der Begriff „Anapsida“ verschwindet daher zunehmend aus der zoologischen und paläontologischen Literatur.
Synapsider Schädel


Der synapside Schädel besitzt ein einzelnes Schädelfenster im Schläfenbereich (Temporal- oder Schläfenfenster) auf jeder Schädelseite. Es wird gerahmt von den Schädelknochen Jugale, Postorbitale und Squamosum sowie in einigen Fällen auch vom Quadratojugale. Die von Jugale und Quadratojugale oder Squamosum in Form eines meist relativ schmalen Knochenstegs gebildete untere Umgrenzung des Schläfenfensters wird auch unterer Temporal- oder Schläfenbogen genannt.
Diese Konfiguration findet sich im Grundbauplan des Schädels der Synapsida, jener Gruppe, die auch die Säugetiere beinhaltet. Bei den Säugern ist das Schädelfenster jedoch sekundär geschlossen und seine vormalige Präsenz nur anhand des nach wie vor vorhandenen unteren Schläfenbogens, nunmehr als Jochbogen (Arcus zygomaticus) bezeichnet, erkennbar.
Auch bei einigen Vertretern der Parareptilia (Bolosaurier, Acleistorhinus und eventuell Mesosaurus[3]) treten solche Schläfenfenster auf. Daher sind diese allein kein verlässliches Merkmal für die Zuordnung eines Amnioten zu den Synapsiden.
Diapsider Schädel
„Grundform“
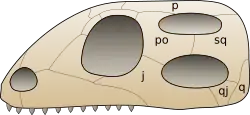
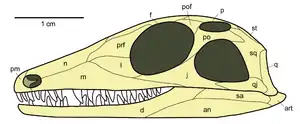
Der diapside Schädel ist durch jeweils zwei Schädelfenster in der Schläfenregion auf jeder Schädelseite gekennzeichnet: Ein oberes Schläfenfenster (Fenestra supratemporalis, Supratemporalfenster), das meist oben auf dem Schädeldach sitzt und von den Schädelknochen Parietale, Postorbitale und Squamosum sowie ursprünglich auch Postfrontale gerahmt wird, und ein unteres Schläfenfenster (Fenestra infratemporalis, Infratemporalfenster) in der Schädelseitenwand, eingefasst vom Jugale, Postorbitale, Squamosum und Quadratojugale. Der untere Schläfenbogen wird wie bei den Synapsiden vom Jugale und Quadratojugale gebildet. Der oft ebenfalls relativ schmal ausgebildete Knochensteg, der unteres und oberes Schläfenfenster voneinander trennt und vom Postorbitale und Squamosum gebildet wird, heißt oberer Schläfenbogen.
Diese Konfiguration repräsentiert den Schädelbau des gemeinsamen Vorfahren wahrscheinlich aller heutigen Reptilien, einschließlich der Vögel (Diapsiden-Kronengruppe).
Bei den Brückenechsen ist das Quadratojugale zugunsten einer Beteiligung des Squamosums von der Bildung des unteren Schläfenbogens ausgeschlossen. Daher wird angenommen, dass Brückenechsen von Vorfahren mit reduziertem unterem Temporalbogen abstammen und der untere Schläfenbogen der Brückenechsen neu entstanden ist. Auch der untere Temporalbogen der Archosaurier (Vögel, Krokodile und deren fossile Verwandte) ist wahrscheinlich eine Neubildung[4] (siehe katapsider Schädel).
„Triapsider“ Schädel

.jpg.webp)
Der „triapside“ Schädel ist eine Unterform des diapsiden. Er ist durch ein weiteres Schädelfenster vor der Augenhöhle gekennzeichnet, das sogenannte Prä- oder Antorbitalfenster. Dieses wird in der Regel von Lacrimale und Maxillare, teilweise auch von der vorderen Partie des Jugale gerahmt. Zusätzlich weist dieser Schädeltyp oft auch eine vergrößerte Nasenöffnung auf. Dieser Typus ist der ursprüngliche Schädeltyp der Archosauria, deren rezente Großgruppen die Krokodile und Vögel sind. Sowohl bei Krokodilen als auch bei Vögeln ist der triapside Schädel sekundär abgewandelt: Krokodile besitzen kein Präorbitalfenster mehr und der Vogelschädel ist so stark umgebildet, dass keines der Schädelfenster noch klar erkennbar ist. Deutlich als solche erkennbare triapside Schädel finden sich daher nur bei fossilen Archosauriern, so z. B. bei frühen engeren Verwandten der Krokodile (u. a. den Rauisuchiden) und bei vielen Dinosauriern.
Euryapsider Schädel


Der euryapside Schädel ist ein diapsider Schädel, bei dem das untere Schläfenfenster sekundär verschlossen ist. Die „Euryapsiden“, deren Name sich von diesem Fensterungstyp ableitet, sind eine Gruppe großer, ausgestorbener Meeresreptilien aus dem Mesozoikum, welche die Ichthyosaurier und die Sauropterygier (Nothosaurier, Plesiosaurier und Placodontier) beinhaltet. Einige jüngere Untersuchungen der Schädelanatomie haben jedoch erbracht, dass Ichthyosaurier und Sauropterygier möglicherweise nicht auf einen gemeinsamen Vorfahren zurückgehen, der euryapside Schädel also in beiden Gruppen unabhängig voneinander entstanden sein könnte.[5] Die Verwandtschaft der „Euryapsida“ scheint sogar so weitläufig zu sein, dass die Gruppe als polyphyletisch betrachtet werden müsste. Aber auch das landlebende Reptil Araeoscelis aus dem Perm von Texas, das weder mit Ichthyosauriern noch mit Sauropterygiern näher verwandt ist, hat einen euryapsiden Schädel.[5]
Parapsider oder metapsider Schädel


Beim parapsiden oder metapsiden Schädel handelt es sich ebenfalls um einen modifizierten Diapsidenschädel mit ausschließlich oberem Schläfenfenster. Die untere oder vielmehr äußere (laterale) Umgrenzung des oberen Schläfenfensters wird dabei nicht von Postorbitale und Squamosum, sondern von Postfrontale und Supratemporale gebildet, weshalb angenommen wird, dass der „echte“ euryapside und der parapside Schädel unabhängig voneinander entstanden sind. Die parapside Konfiguration ist typisch für Ichthyosaurier.[6]
In der Literatur des frühen 20. Jahrhunderts wird der Begriff „parapsid“ gleichbedeutend mit „euryapsid“ und „katapsid“ (siehe unten) verwendet und bezeichnet alle Reptilienschädel ohne unteren Temporalbogen, unabhängig davon, ob dieser Zustand das Ergebnis einer Reduktion des Temporalbogens oder eines kompletten Verschlusses des unteren Schläfenfensters ist. Der Taxonname „Parapsida“ leitet sich von dieser Bedeutung des Begriffes ab und bezeichnet eine Gruppe, die sowohl Ichthyosaurier als auch Araeoscelis und die Schuppenkriechtiere (Squamata) umfasst.[7] Eine solche Gruppierung gilt aber bereits seit langem als nicht natürlich und die Bezeichnung „Parapsida“ ist entsprechend veraltet.
„Katapsider“ Schädel


Auch der katapside Schädel ist ein abgeleiteter Diapsidenschädel. Bei ihm ist der untere Schläfenbogen reduziert. Er kommt rezent ausschließlich bei Schuppenkriechtieren (Squamata) vor, ist aber vermutlich relativ früh in der Evolution der Diapsiden entstanden und dann in verschiedenen Entwicklungslinien (Archosauria, Brückenechsen) konvergent wieder zu einem „echten“ diapsiden Schädel umgebildet worden.[4] Die Reduktion des unteren Schläfenbogens ging mit einer Größenreduktion des Quadratojugale einher. Eine Besonderheit des katapsiden Schädels der Squamaten ist die gelenkige Verbindung zwischen Quadratum und Squamosum sowie eine vollständige Reduktion des Quadratojugale.
Siehe auch
Literatur
- Michael J. Benton: Paläontologie der Wirbeltiere. Übersetzung der 3. englischsprachigen Auflage (Übersetzer: Hans-Ulrich Pfretzschner). Verlag Dr. Friedrich Pfeil, München 2007, ISBN 978-3-89937-072-0
Einzelnachweise
- Rafael Zardoya, Axel Meyer: Complete mitochondrial genome suggests diapsid affinity of turtles. Proceedings of the National Academy of Sciences of the United States of America. Bd. 25, Nr. 19, 1998, S. 14226–14231, doi:10.1073/pnas.95.24.14226
- Ylenia Chiari, Vincent Cahais, Nicolas Galtier, Frédéric Delsuc: Phylogenomic analyses support the position of turtles as the sister group of birds and crocodiles (Archosauria). BMC Biology 2012, 10:65, doi:10.1186/1741-7007-10-65 (Open Access)
- Graciela Piñeiro, Jorge Ferigolo, Alejandro Ramos, Michel Laurin: Cranial morphology of the Early Permian mesosaurid Mesosaurus tenuidens and the evolution of the lower temporal fenestration reassessed. Comptes Rendus Palevol. Bd. 11, Nr. 5, 2012, S. 379–391, doi:10.1016/j.crpv.2012.02.001
- Johannes Müller: Early loss and multiple return of the lower temporal arcade in diapsid reptiles. Naturwissenschaften. Bd. 90, Nr. 10, 2003, S. 473–476, doi:10.1007/s00114-003-0461-0
- Michel Laurin, Jacques A. Gauthier: Diapsid Phylogeny. Tree Of Life web project, 2000
- Michael W. Maisch: Phylogeny, systematics, and origin of the Ichthyosauria – the state of the art. Paleodiversity. Bd. 3, 2010, S. 151–214, online (PDF; 1,3 MB)
- Samuel W. Williston: The Phylogeny and Classification of Reptiles. The Journal of Geology. Bd. 25, Nr. 5, 1917, S. 411–421